

Abstract
Parasitic nematodes are potent modulators of immune reactivity in mice and men. Intestinal nematodes live in close contact with commensal gut bacteria, provoke biased Th2 immune responses upon infection, and subsequently lead to changes in gut physiology. We hypothesized that murine nematode infection is associated with distinct changes of the intestinal bacterial microbiota composition. We here studied intestinal inflammatory and immune responses in mice following infection with the hookworm Heligmosomoidespolygyrus bakeri and applied cultural and molecular techniques to quantitatively assess intestinal microbiota changes in the ileum, cecum and colon. At day 14 post nematode infection, mice harbored significantly higher numbers of γ-Proteobacteria/Enterobacteriaceae and members of the Bacteroides /Prevotella group in their cecum as compared to uninfected controls. Abundance of Gram-positive species such as Lactobacilli, Clostridia as well as the total bacterial load was not affected by worm infection. The altered microbiota composition was independent of the IL-4/-13 – STAT6 signaling axis, as infected IL-4Rα-/- mice showed a similar increase in enterobacterial loads. In conclusion, infection with an enteric nematode is accompanied by distinct intestinal microbiota changes towards higher abundance of gram-negative commensal species at the small intestinal site of infection (and inflammation), but also in the parasite-free large intestinal tract. Further studies should unravel the impact of nematode-induced microbiota changes in inflammatory bowel disease to allow for a better understanding of how theses parasites interfere with intestinal inflammation and bacterial communities in men.
Introduction
Nematodes are the most prevalent worms worldwide dwelling the intestine of humans. Infections with Ascaris lumbricoides , Trichuris trichura , Ancylostoma duodenale and Necator americanus generally lead to persistent intestinal colonization [1]. Our study aimed to analyse the influence of infections with the murine hookworm model Heligmosomoidespolygyrus bakeri ( H . p . bakeri ) [2] on the composition of the bacterial microbiota in distinct compartments of the murine intestinal tract. H . p . bakeri inhabits an environment colonized by a large number of commensal and symbiotic microorganisms. The commensal host microbiota is essential for physiological processes, such as nutrient acquisition, maintenance of epithelial barrier function, and mucosal immune homeostasis [3,4]. Information regarding potential quantitative and qualitative microbiota changes induced by intestinal worm infections are scarce, but of major interest, given that distinct commensal species have been shown to be involved in development and perpetuation of inflammatory bowel disease (IBD) in humans [5,6]. In addition, parasitic nematodes are considered as a therapeutic option for the treatment of IBD [7] given that trials applying helminth eggs to patients with Crohn’s disease and ulcerative colitis resulted in disease remission [8]. Suppression of intestinal inflammation and induction of regulatory immune responses by parasitic nematode infections have been demonstrated in various IBD models (reviewed in 9). Furthermore, enteropathogenic infections and murine experimental colitis lead to dramatic changes in the intestinal ecosystem with alterations in the microbiota composition [10]. Although these models reflect a multifactorial etiology of intestinal inflammation, the resulting changes in the composition of the intestinal microbial community are consistently seen as a preferential overgrowth of the Gram-negative Enterobacteriaceae. Hence, it is under debate whether the phenomenon of “dysbiosis” in animal models and human IBD is primarily resulting from intestinal inflammation or initiating and perpetuating the inflammatory condition per se (“hen and egg”) [11–13].
In the presented study, we investigated the alterations of the intestinal bacterial microbiota composition following a gastrointestinal hookworm infection applying cultural and PCR-based techniques. We here show that i) H . p . bakeri infection was accompanied by a shift towards increased abundance of Gram-negative commensal species. ii) Remarkably, the most distinct changes of the microflora were determined distal from the site of acute nematode infection, i.e. in the cecum and colon. iii) IL-4Rα-/- mice showed similar changes in the microbiota composition after nematode infection, revealing that changes induced by the IL-4/-13 – STAT-6 signalling axis in the intestines are not involved in permitting the outgrowth of Gram-negative bacteria.
Materials and Methods
Ethics Statement
All animal experiments were performed in strict accordance with the national animal protection guidelines and approved by the appropriate ethics committee (Landesamt für Gesundheit und Soziales, Berlin, Germany; Permit Number G0363/10).
Mice, parasites and infections
Female C57BL/6 mice (purchased from Charles River) and IL-4Rα-/- mice (C57BL/6 background, a kind gift from M. Löhning, DRFZ, Berlin) were kept under specific pathogen-free conditions (light/dark cycles of 12h) in ventilated cabinets in cages equipped with standard bedding, filter tops, environmental enrichment and fed standard diet and autoclaved water ad libitum. 200 H . p . bakeri infective larvae (L3) were applied in 100µl water orally to 6-8 week old mice. Control mice received water in which the L3 had been stored at 4°C. Mice were sacrificed by cervical dislocation after inhalational (isoflurane) anesthesia on day 6, 14 or 28 after H . p . bakeri infection.
Cell culture, gut homogenates and cytokine detection
Mesenteric lymph node cells (mLNC) were isolated aseptically and single cell suspensions were plated in RPMI 1640 containing 5% fetal calf serum (both from BioChrom, Germany), 20 mM L-glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin (PAA, Germany) as triplicates on 96-well plates. Cultures were kept at 37°C and 5% CO2 with 3.5×105 cells in a total volume of 200 µl. Stimulation with concanavalin A (ConA, 2 µg/ml) was performed for 48 h, cultures with adult worm antigen extracts (40 µg/ml) were kept for 72 h. Supernatants were harvested and stored at -80°C. Culture supernatants were assayed by ELISA (kits from BD Biosciences and R&D Systems) for IL-4, IL-13, IL-10, IL-12p70, IL-17A, IFN-γ, and TGF-β1 according to the manufacturer’s instructions.
Histopathology
Intestinal tissue samples were fixed in 3.7% phosphate-buffered formalin, embedded in paraffin and used for cross sections. The histopathological score in response to infections was determined using hematoxylin/eosin stained cross sections. The grade of inflammation of small intestine, cecum and colon was evaluated according to the following parameters: (1) epithelial damage, (2) edema development, (3) inflammatory cell infiltration, (4) crypt depth/villus length. Goblet cells were quantified in periodic acid and Schiff’s reagent stained sections (10 high power fields/section).
Cultural analysis of the intestinal microflora
Quantitative cultural analyses were performed as described earlier [14]. In brief, ileal, cecal and colonic luminal contents were immediately resuspended in sterile PBS. Serial dilutions were plated onto respective solid media, bacteria were grown at 37 °C for two days under aerobic and obligate anaerobic conditions and bacterial loads expressed as colony forming units (CFU) per gram luminal content (wet weight).
Molecular analysis of the intestinal microbiota
DNA extractions from intestinal luminal contents were prepared as described previously [14]. In brief, DNA extracts and plasmids were quantified by using Quant-iT PicoGreen reagent (Invitrogen, UK) and all adjusted to 1 ng DNA/µl.
The abundance of specific intestinal bacterial groups was measured by qPCR with group-specific 16S rRNA gene primers (Tib MolBiol, Germany) as decribed previously [15,16]. As reference for quantification standard curves with tenfold serial dilutions of plasmids (ranging from 2×108 to 2×102 copies) were generated for each run. The real-time PCR primers were first used to amplify cloned 16S rDNA of reference strains (see Table 1). The number of 16S rRNA gene copies / ng DNA of each sample was determined. Frequencies of the given bacterial groups were calculated proportionally to the eubacterial (V3) amplicon.
Table 1. 16S rRNA gene group-specific primer for quantitative Real Time-PCR a .
| Target | Reference Strain | Primer sequence (5´ to 3´) and Orientationb | Reference |
|---|---|---|---|
| Domain Bacteria (targets 16S V3 region) | Escherichia coli ATCC 25922 | F: CGGYCCAGACTCCTACGGG, R:TTACCGCGGCTGCTGGCAC | [50] |
| Clostridium leptum subgroupc | Clostridium leptum DSMZ 753 | F: TTACTGGGTGTAAAGGG, R: TAGAGTGCTCTTGCGTA | [51] |
| Clostridium coccoides - Eubacterium rectale subgroupd | Clostridium coccoides DSMZ 935 | F: AAATGACGGTACCTGACTAA, R: CTTTGAGTTTCATTCTTGCGAA | [52] |
| Bacteroides groupe | Bacteroides ovatus DSMZ 1896 | F: GAAGGTCCCCCACATTG, R: CAATCGGAGTTCTTCGTG | [53] |
| γ-Proteobacteria/ Enterobacteriaceae | Escherichia coli ATCC 25922 | F: AAACTCAAATGAATTGACGG, R: CTTTTGCAACCCACTCC | [54] |
| Lactobacillus groupf | Lactobacillus acidophilus DSM 20079 | F: CACCGCTACACATGGAG, R: AGCAGTAGGGAATCTTCCA | [55,56] |
| Bifidobacterium genus | Bifidobacterium sp. (murine origin) | F: CTCCTGGAAACGGGTGG, R: GGTGTTCTTCCCGATATCTACA | [52,55,56] |
| Enterococcus genus | Enterococcus faecalis DSM 20478 | F: CCTTATTGTTAGTTGCCATCATT, R: ACTCGTTGTACTTCCCATTGT | [57] |
| Mouse Intestinal Bacteroides | MIB plasmid 16-1 | F: CCAGCAGCCGCGGTAATA, R: CGCATTCCGCATACTTCTC | [58] |
Modified from: Ref. [59]
F, Forward R, Reverse
including Faecalibacterium (Fusobacterium) prausnutzii Clostridium 16S rRNA cluster IV
Clostridium 16S rRNA cluster XIVa/b
including Prevotella and Porphyromonas
including Leuconostoc , Pediococcus , Aerococcus and Weissella but not Enterococcus or Streptococcus
Genetic fingerprints were generated by PCR-based denaturing gradient gel electrophoresis (PCR-DGGE) as described previously [14].
Statistical analysis
All experiments were performed with 4 to 6 mice per group and representative data are shown for at least two independent experiments as mean + SEM unless stated otherwise. Statistical analysis was performed using GraphPad Prism software using the two-tailed Mann–Whitney U-test. Values of p<0.05 were considered to be statistical significant.
Results
Histopathological mucosal changes following H . p . bakeri infection
Infections with H . p . bakeri are generally well tolerated by mice and do not lead to signs of morbidity such as weight loss [17]. As the intestinal flora is perceptive to inflammatory changes of the gut [10,12,14], we surveyed the proximal small intestine as the site of nematode infection and, additionally, the worm-free large intestine histopathologically for signs of inflammation. Inflammatory scores based on morphological changes and the size of inflammatory cell infiltrates were compiled with cross sections from the duodenum, ileum, cecum and colon on day 6 (larval development), 14 (acute adult stage) and 28 post infection (chronic adult stage). When compared to naive controls, mice infected with H . p . bakeri for 6 and 14 days displayed a rather mild, but significantly elevated inflammatory score of the duodenum, but not ileum, cecum and colon (Figure 1 A, C, D). The locally restricted enteritis of the small intestine peaked during the larval and acute adult stage infection and declined thereafter during chronicity. In addition, development of enteritis was accompanied by local duodenal goblet cell hyperplasia while goblet cells counts remained unaltered in distal parts of the intestines (Figure 1 B, C, D).
Figure 1. H . p . bakeri -induced intestinal immunopathology.

(A) Enteritis development was evaluated at day 6, 14 and 28 post infection and naïve controls according to a scoring system based on epithelial damage, edema development, villus length, crypt depth and the size of inflammatory cell infiltrates. (B) Goblet cells counts as detected in intestinal cross sections by periodic acid/Schiff stain. Mean + SEM is shown for 5 mice per group; * p < 0.05. Representative cross sections from duodenum (C) and colon (D) stained with H&E (upper row) and periodic acid/Schiff (lower row, goblet cells marked by intense purple stain) for histopathological scoring and goblet cell quantification.
Thus, infection with H . p . bakeri led to the development of a mild enteritis and increase of mucus producing goblet cells in the proximal small intestine, whereas distal parts of the intestinal tract appeared morphologically unaltered.
Cytokine response on the peak of pathology
In order to quantify the local immune response to H . p . bakeri we analyzed cells from the gut-draining mesenteric lymph nodes (mLN) and intestinal tissue of infected mice. On day 14 days post infection the key Th2 cytokines IL-4 and IL-13 as well as anti-inflammatory IL-10 were produced in significantly higher amounts by mLN cells from infected mice in response to mitogen and parasite antigen (Figure 2A, 2B, 2D). The levels of TGF-β did not change in acutely infected mice (Figure 2E). IFN-γ was significantly increased in mitogen-treated cultures from H . p . bakeri infected mice, but as expected, only negligible amounts (< 50 pg/ml) were secreted in response to parasite extracts (Figure 2C). IL-17A levels were generally very low (Figure 2 F).
Figure 2. Cytokineproduction by mLN cells during acute H . p . bakeri infection.

IL-4 (A), IL-13 (B), IFN-γ (C), IL-10 (D), TGF-β (E) and IL-17A (F) were measured in mLN culture supernatants with (ConA, H.p.b-Ag) or without (-) stimulation. Cells were derived from naïve controls and mice at day 14 post infection. Mean + SEM is shown for 5 mice per group. H.p.b. Ag: H. p. bakeri antigen; Con A: concanavalin A; * p < 0.05; ** p < 0.005.
Intestinal cytokine responses were analyzed in whole tissue from duodenum and cecum. Trends for increased IL-4/-13 and IL-10 production (n.s.) were detected in ileum as well as cecum tissue whereas IL-12, IFN-γ and TGF-β expression were detected at similar levels, irrespective of the infection status (data not shown).
Cultural analysis of the intestinal microflora after nematode infection
Given that the most pronounced small intestinal immunopathology correlated with increased levels of Th2 cytokines and IL-10 produced in mLN, we were further interested whether the acute phase of infection was accompanied by distinct shifts within the composition of the intestinal microbiota. To address this, we next quantitatively assessed the cultivable microbiota in ileal, cecal and colonic luminal contents taken on day 14 following H . p . bakeri infection. Infected mice displayed a trend towards higher total loads of cultivable bacteria (n.s.) as compared to uninfected controls, irrespective of the intestinal compartment analyzed (Figure 3). Strikingly, the enterobacterial loads increased significantly by 2-3 orders of magnitude in ileum, cecum as well as colon upon acute infection (Figure 3 A–C).
Figure 3. Quantification of aerobic intestinal bacterial groups.
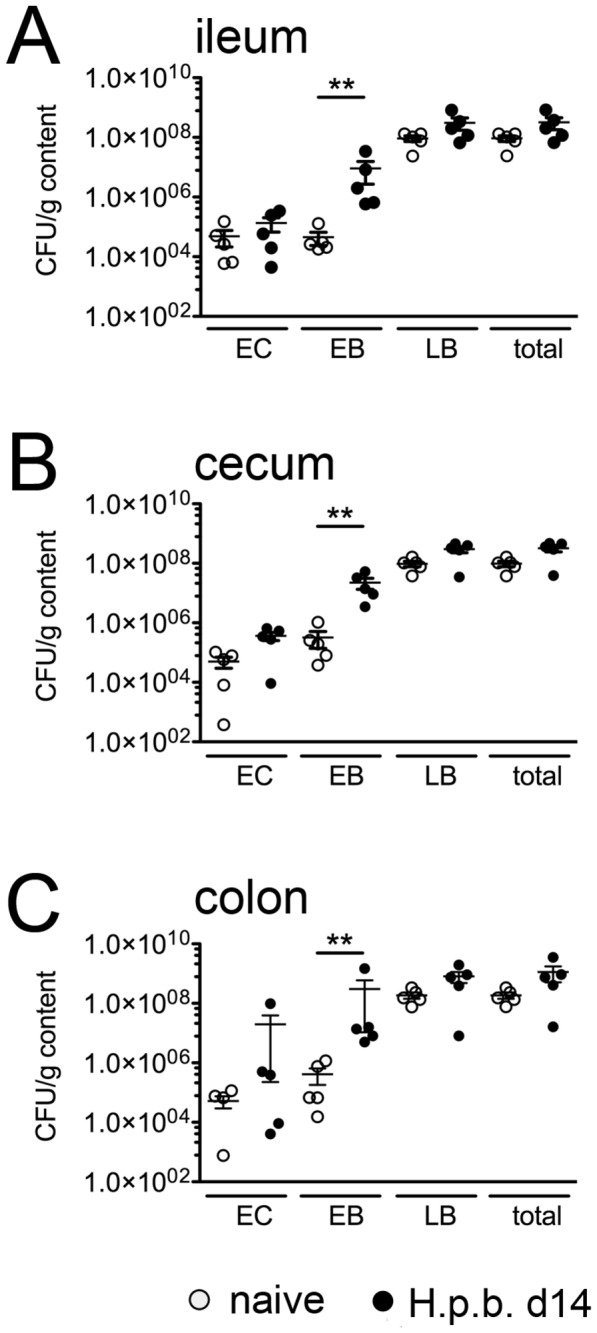
Counts of colony forming units (CFU) derived from luminal content of (A) ileum, (B) cecum and (C) colon. Open circles: naïve controls; filled circles: day 14 post H . p . bakeri infection. Mean ± SEM of 5 mice per group is shown. EC: Enterococci, EB: Enterobacteriaceae, LB: Lactobacilli; ** p < 0.005.
Molecular analysis of the microbiota composition during H . p . bakeri infection
We next performed a comprehensive molecular survey of the microbiota composition of ileum, cecum and colon upon H . p . bakeri infection applying quantitative 16S rRNA technique. Irrespective of the intestinal compartment, total bacterial abundance did not change in infected mice (Figure 4). Whereas abundances of Gram-positive bacterial groups such as Lactobacilli and Clostridium clusters were similar in cecum and colon of naïve and H . p . bakeri infected mice (< 2 fold increase, Figure 4B, 4C), a trend towards higher loads was detected in the small intestine of infected mice (3.7-7.0 fold increase, n.s.; Figure 4A). Significantly increased loads of gram-negative γ-Proteobacteria/Enterobacteriaceae were detected in cecal contents of H . p . bakeri colonized animals as compared to naive mice (32.7 fold increase, Figure 4B), whereas only a trend towards higher abundance of Gram-negative enterobacterial groups upon infection could be detected in the colon (Figure 4C). In addition, the cecum of infected mice harboured significantly higher loads of the Gram-negative obligate anaerobic Bacteroides group (93.6 fold increase, Figure 4B) whereas members of the operational taxonomic unit mouse intestinal Bacteroides (MIB), had increased in the ceca and colons following infection (Figure 4B and 4C). Irrespective of the infection, Enterococci were abundant at comparable low levels in ceca and colons (Figure 4B and 4C), whereas Bifidobacteria could not be detected at all.
Figure 4. Analysis of 16S rRNA gene-based real-time PCR amplification of intestinal bacterial groups.
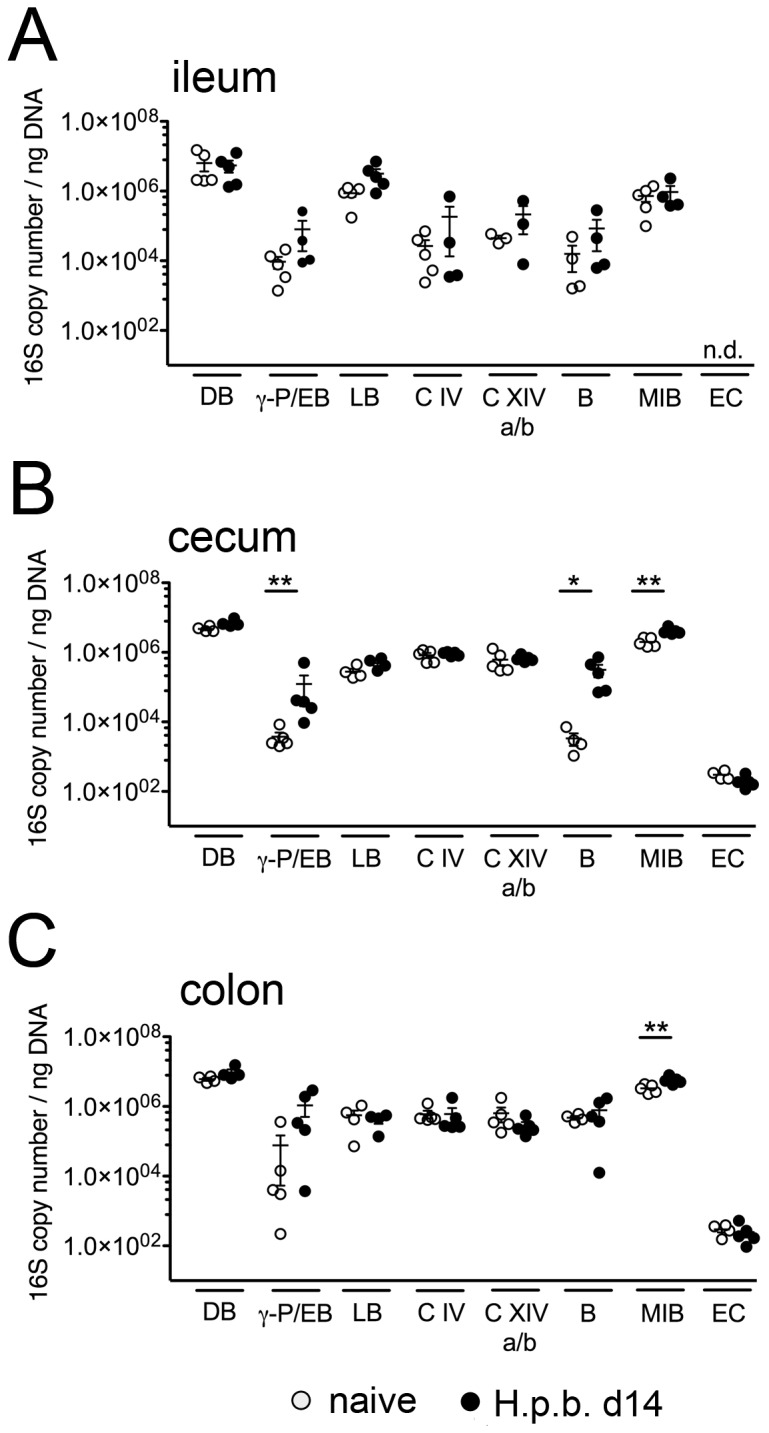
Detected DNA levels in luminal content from (A) ileum, (B) cecum and (C) colon of naive (open circles) and acutely infected (day 14 post infection, black circles) mice were calculated as 16S rRNA copy numbers per ng DNA extract. Mean ± SEM is shown for 4-5 mice per group. DB: domain bacteria, γ-p/EB: γ-Proteobacteria/Enterobacteriaceae, LB: Lactobacilli, C IV: Clostridium cluster IV, C XIV a/b: Clostridium cluster XIV a/b, B: Bacteroides group, MIB: mouse intestinal Bacteroides , EC: Enterococci, n.d.: not detectable. * p < 0.05; ** p < 0.005 compared to naïve.
In the following we calculated the relative abundance of the respective bacterial groups (Figure 5). The amplification with primers specific for the γ-Proteobacteria/Enterobacteriaceae, Lactobacillus group, Clostridium cluster IX, Clostridium cluster XIVa/b, MIB and Bacteroides group covered 81-96% of the total intestinal bacterial microbiota except for the ileal content of naïve mice, where only about 44% of the bacterial load was covered (Figure 5A). The higher coverage for ileal samples from nematode infected mice was mainly due to an increase of the Lactobacillus group from 26% ± 12 in naïve to 62% ± 16 in H . p . bakeri -infected mice (Figure 5A). Furthermore, frequencies of the cecal Bacteroides group were barely detectable in naïve controls but increased to about 4% in acutely infected mice (Figure 5B). Remarkably, the most distinct proportional increase for γ-Proteobacteria/Enterobacteriaceae was detected in the colon (from 1% ± 1 in naive to 7% ± 5 in acutely infected mice (Figure 5C)). Taken together, the data show that the applied PCR technique covered the majority of bacterial strains present in the murine intestinal tract, except for the small intestinal microbiota in naïve mice. Furthermore, the most prominent findings were increased abundances and proportions of Gram-negative γ-Proteobacteria/Enterobacteriaceae, Bacteroides and MIB in intestinal parts distal of the site of worm infection.
Figure 5. Proportions of intestinal bacterial group based on 16S rRNA gene real-time PCR.

Pie charts represent the mean frequencies for the main bacterial phyla in (A) ileum, (B) cecum and (C) colon as detected in 4-5 mice per group. Abundance was calculated using the phylum-specific 16S gene copy numbers per ng DNA in relation to the amounts of the DNA copy numbers for the domain Bacteria set to 100%. p.i.: post H . p . bakeri infection.
Given that intestinal inflammation is associated with decreased bacterial diversity [14,18,19] we finally applied denaturing gradient gel electrophoresis (DGGE), another PCR-based technique to generate genetic fingerprints of the intestinal microbiota according to the intestinal bacterial molecular band profiles before and after worm infection (Figure 1). Interestingly, six additional bands could be detected in the ileal samples derived from mice 14 day pi, which were absent in naïve controls and during the very early and chronic phase of infection (day 6 and 28 p.i., respectively) (Figure S1A). Similarly, the most distinct / overt differences were detected in cecum and colon samples at day 14 p.i. (Figure S1B, S1C). Remarkably, changes in bacterial diversity patterns of the cecal microbiota were not exclusively found during acute, but also in the early and chronic phase of infection.
DGGE bands only present in the DNA extracted from ileal contents of naïve mice could be identified as Paenibacillus spp. (Figure S2A), whereas additional bands present in cecal content of acutely infected mice were identified as Gram-negative Prevotella spp. and Porphyromonadaceae belonging to the Bacteroides group (Figure S2B). Members of Lachnospiraceae were detected in the cecum of all acutely infected mice, while only 2 out of 5 naïve mice showed the respective bands (Figure S2B).
Increase in Enterobacteria after H . p . bakeri infection is independent of IL-4Rα-/- signalling
As physiological gut functions such as mucosal permeability, chloride secretion, smooth muscle contractility and glucose absorption of the small and large intestine are altered in nematode infected mice and this is mainly mediated by IL-4/-13 [20–22], we asked whether the microbiota changes detected in the acute phase of infection with H . p . bakeri were depending on the IL-4/-13 – STAT6 signalling axis. Hence we analyzed the intestinal pathology and the cultivable flora of naïve and acutely infected IL-4Rα-/- mice. Expectedly, IL-4Rα-/- mice showed a substantially decreased Th2 cytokine response as indicaticated by significantly downreguated IL-4/-13 production of mLN cells and a diminished IL-10 response to parasite-specific restimulation as compared to wild type controls, whereas the IFN-γ levels did not differ (Figure 6 A–D). Worm burdens were comparable in wild type and IL-4Rα-/- mice (Figure 6I). Interestingly, IL-4Rα-/- mice displayed virtually no sign of duodenal goblet cell hyperplasia in response to H . p . bakeri infection (Figure 6E, 6G, 6H), but developed a locally restricted duodenal enteritis indistinguishable from the inflammatory response in infected wild type mice (Figure 6F, 6G, 6H).
Figure 6. Immune response, histopathology and parasite burden of infected wild type versus IL-4Rα-/- mice.

Levels of IL-4 (A), IL-13 (B), IL-10 (C) and IFN-γ (D) produced by mLN cells from naïve controls and mice at day 14 post infection in response to adult worm antigen. (E) Goblet cell counts in duodenal cross sections. (F) Enteritis scores for duodenal tissue. Representative duodenum cross sections from naïve (G) and acutely infected (H) wild type and IL-4Rα-/- mice stained with H&E (upper row) and periodic acid/Schiff (lower row) for histopathological scoring and goblet cell quantification. (I) Adult worm counts of WT and IL-4Rα-/- mice at day 14 post infection. Mean + SEM of 5-6 mice per group is shown. * p < 0.05; ** p < 0.005.
Remarkably, the main shifts in intestinal microbiota composition observed in infected IL-4Rα-/- mice were also due to increased abundances of Enterobacteriaceae (Figure 7) as indicated by higher enterobacterial loads in both ileum and colon, but not in the cecum (Figure 7) as compared to uninfected controls. Lactobacilli numbers also increased significantly in the ileal lumen of IL-4Rα-/-, but not wild type mice (Figure 7 A and data not shown). Taken together, the increase of intestinal Enterobacteriaceae after H . p . bakeri infection occurs independent of IL-4/-13-induced changes in the gut.
Figure 7. Quantification of aerobically cultivable bacteria in wild type and IL-4Rα-/- mice.
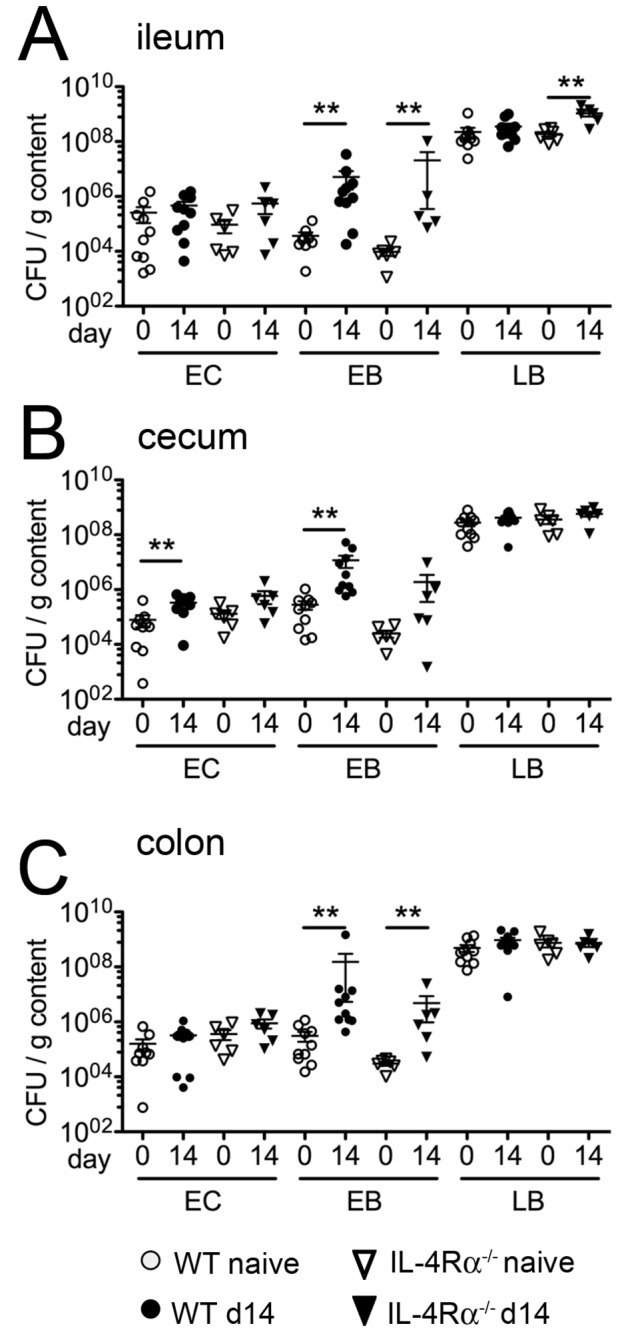
Counts of colony forming units (CFU) derived from luminal content of (A) ileum, (B) cecum and (C) colon of WT (naïve: open circles; d14 p.i. black circles) and IL-4Rα-/- mice (naïve: open triangles; d14 p.i. black triangles). Mean ± SEM of 5-6 mice per group is shown. CFU: colony forming units, EC: enterococci, EB: Enterobacteriaceae, LB: Lactobacilli; ** p < 0.005.
Discussion
In the present study we analyzed whether infection with the nematode H . p . bakeri dwelling the proximal small intestine and the subsequently induced immune responses are associated with distinct changes of the intestinal bacterial microbiota composition. A plethora of studies in mice and men revealed that intestinal inflammation is associated with alterations in the composition of the gut microbiota towards a lower diversity and overgrowth with mainly Gram-negative species such as enterobacterial and Bacteroides spp. by the cost of decreasing Lactobacilli [10,14,18,19,23]. We analyzed the bacterial microbiota composition during the acute phase of the nematode infection marked by the development of a locally restricted small intestinal enteritis and the highest expression of Th2 related cytokines [17,24]. The strong Th2 response leads to changes of the small and large intestinal physiology, including increased mucosal permeability, chloride secretion and smooth muscle contractility as well as decreased glucose absorption [20–22]. We thus aimed to dissect the contribution of the highly biased Th2 immune response to alterations of the microbiota.
The identification of intestinal bacteria is impeded by the fact that most species are non-cultivable or restricted to specialized cultivation conditions. Hence, in order to analyze the complexity of microbial communities we combined classical cultivation techniques with culture-independent molecular approaches. Beside denaturing-gradient-gel-electrophoresis (DGGE) demonstrating the high diversity of the species present in the intestinal tract, a quantitative PCR technique based on the detection of 16S rRNA gene copy numbers was used for the analysis of dominant microbial groups. The latter technique quantifying γ-Proteobacteria/Enterobacteriaceae, the Lactobacillus group, Clostridium clusters IX and XIVa/b, the operational taxonomic unit MIB and the Bacteroides group permitted the detection of 81-96% of the total bacterial microbiota within the cecum and colon of naïve controls and acutely worm-infected mice. Of note, we restricted our analyses of infection-related changes of the microbiome to bacteria, thus possible alterations in archaeal and eukaryotic species remain to be investigated.
Intestinal inflammation may alter the total intestinal bacterial loads [12,18,19] and highly pathogenic intestinal inflammation models such as Toxoplasma gondii-driven ileitis and Citrobacter - or Salmonella -induced colitis are characterized by decreased loads of Gram-positive bacteria such as Lactobacilli and Clostridium groups [12,14,25]. We detected no significant effect of infections with H . p . bakeri on the total bacterial load in infected animals in our experiments.
In our study, the strongest alterations following acute H . p . bakeri infections were detected for Gram-negative bacteria. We found significant increases of cultivable Enterobacteria in all examined intestinal compartments, a finding confirmed by quantitative PCR for γ-Proteobacteria/Enterobacteriaceae. IBD-like inflammation models, regardless whether induced by infections, chemicals or genetic predisposition [10,14,19], as well as human IBD [26–28] are characterized by overgrowth of Gram-negative Enterobacteriaceae and such changes in the intestinal microbiota may be associated with the induction or perpetuation of the inflammatory response in IBD. It thus is interesting that a nematode infection leading to a mild and locally restricted inflammatory response also led to a significant increase in bacteria associated with inflammatory disorders like IBD. Of note, infections with H . p . bakeri have been shown to suppress inflammation in several models of colitis (reviewed in [9]) which are associated with increased intestinal loads of Gram-negative Enterobacteriaceae [10,29]. Future work will have to elucidate whether the nematode infection also affects the microbiota composition in models of colitis.
A study by Walk and colleagues previously described alterations in the gut microbiota after infection with H . p . bakeri [30]. In this study, a significantly increased abundance of members of the Lactobacillaceae family was detected with a 16s rRNA clone library generated from terminal ileum tissue samples from acutely worm infected mice after removal of the luminal content. Our data based on the analysis of luminal content also show an increase in the relative distribution of Lactobacillaceae members in the ileum. Sequencing of DGGE-derived DNA-bands detectable predominantly or exclusively in acutely nematode-infected mice revealed an increase in Lachnospiraceae and Porphyromonaceae members after infection, which confirms the data published by Walk et al. [30]. Differing compositions of the gut-wall-associated versus luminal flora and the relatively low abundance of Enterobacteriaceae in the gut of naïve and infected mice may explain why we detected changes concerning this bacterial group that have not been detected in the previously published study.
It is up to speculation whether inflammation-driven changes in the gut physiology after H . p . bakeri infection directly facilitate the outgrowth of Gram-negative bacteria or act indirectly by creating conditions less well tolerated by other groups of commensals. Enterobacteriaceae as facultative anaerobes use mixed-acid fermentation under anaerobic conditions to produce lactate, succinate, ethanol, acetate and carbon dioxide. These pathways require low levels of hydrogen. One factor facilitating the outgrowth of Enterobacteriaceae in the inflamed gut may be that the members of this group are relatively resistant to oxidative stress mediated by reactive oxygen metabolites, which are produced during inflammatory processes [31,32]. Another reason might be their high efficiency in metabolizing glucose, which is absorbed to a lesser extent during intestinal inflammation [33]. Studies in pigs infected with the nematode Trichuris suis demonstrated changes in both microbial composition and metabolic potential in the lumen of the colon [34,35]. Members of the Enterobacteriaceae, such as E. coli, might profit from an increased fluid influx and impeded glucose absorption induced by infection with H . p . bakeri [20]. These effects, together with an increased mucus production and gut peristaltic, largely depend on the key Th2 cytokines IL-4/-13 and STAT-6 signalling. However, we found no evidence for a role of IL-4/-13 in the detected microbiota changes, as IL-4Rα-/- mice showed similar alterations after infection with H . p . bakeri .
Infections with another intestinal nematode, T . muris , alter the expression of anti-microbial products such as angiogenin 4 and cryptdins in the intestine [36,37]. We hence asked whether the changes in the microbiota composition after infection with H . p . bakeri correlated with altered expression of anti-microbial defence molecules and focussed on members of the cryptdins, angiogenins and cathelecidins with known activity against Gram-negative bacteria such as E. coli, Citrobacter rodentium and members of the Bacteroides family [38–40]. Analysing the cryptin DefB1, ang4 and CRAMP produced by Paneth cells and macrophages/neutrophils, respectively, we found an unaltered expression in worm-infected mice compared to naïve controls (data not shown). Thus it is unlikely that changes in the expression of innate defence molecules are crucially involved in permitting the detected changes in the composition of the intestinal bacterial community.
We and others reported on H . p . bakeri -driven modulations of the host immune response during concurrent intestinal infections [41–43]. On the one hand H . p . bakeri may impair host resistance to bacterial pathogens, such as Citrobacter rodentium and thereby enhance Citrobacter -induced colitis [42] with altered macrophages being incapable to kill internalized pathogenic bacteria. On the other hand, the nematode protects the host from Helicobacter pylori-driven inflammation [43]. Furthermore, H . p . bakeri effectively suppresses intestinal inflammation in murine models of human IBD [44–47]. The fact that H . p . bakeri infection leads to the atypical production of immunosuppressive TGF-β by intestinal CD4+ T cells in response to TLR-4 triggering by LPS [48] poses the question whether such mechanisms of immune modulation help to avoid undesired immune activation by microbiota changes resulting from the infection. It remains to be investigated whether nematodes also interfere with the composition of the intestinal microbiota in face of a dysregulated intestinal immune response. Evidence comes from a study showing that Trichuris trichiura infections in macaques suffering from chronic diarrhea alters the composition of the bacterial microbiota attached to the mucosa, which correlated with a clinical improvement of chronic diarrhea [49].
In conclusion, our study analyzing the ileal, cecal and colonic microbiota composition during an H . p . bakeri infection permits the following insights into the parasite-bacteria interplay: I) An intestinal nematode infection leads to moderate changes in the complexity of the intestinal microbiota and no significant alterations in the total number of intestinal bacteria. II) The peak of pathology induced by the intestinal nematode infection is accompanied by a significant increase in gram-negative bacteria including γ-Proteobacteria/Enterobacteriaceae. III) Changes in microbiota composition occur not only in the proximity of the nematode’s habitat, but also in distal regions of the gut. VI) The detected microbiota alterations in nematode infected mice are not a consequence of IL-4/-13-depending changes in the intestine. The impact of nematode-induced microbiota changes on IBD remains to be investigated, possibly adding to our understanding of how theses parasites interfere with intestinal inflammation.
Supporting Information
DGGE analysis of luminal contents of (A) ileum, (B) cecum and (C) colon. Band profiles shown derive from 3 naïve and infected mice at different time points after infection. Arrowheads mark additional bands during acute H . p . bakeri infection (14 d.p.i.). M: marker.
(TIF)
DGGE profiles and results of sequencing analysis of naive versus acutely infected mice. SYBR green stained gels of extracted DNA from (A) ileum and (B) cecum were used for DNA extraction and sequencing of the marked bands.
(TIF)
Acknowledgments
The authors thank B. Sonnenburg, S. Spiekermann and M. Müller for excellent technical assistance.
Funding Statement
This work was supported by grants of the collaborative research centers SFB 852 (to SH), SFB 650 (to SH), SFB 633 (to SB, AF and MMH) and the Heisenberg-Program (to SH) from the Deutsche Forschungsgemeinschaft and by the American Broad Foundation (to SH). The funders had no role in study design, datacollection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A et al. (2006) Soil-transmitted helminth infections: ascariasis, trichuriasis, and hookworm. Lancet 367: 1521-1532. doi:10.1016/S0140-6736(06)68653-4. PubMed: 16679166. [DOI] [PubMed] [Google Scholar]
- 2. Monroy FG, Enriquez FJ (1992) Heligmosomoides polygyrus: a model for chronic gastrointestinal helminthiasis. Parasitol Today 8: 49-54. doi:10.1016/0169-4758(92)90084-F. PubMed: 15463566. [DOI] [PubMed] [Google Scholar]
- 3. Kelly D, Conway S, Aminov R (2005) Commensal gut bacteria: mechanisms of immune modulation. Trends Immunol 26: 326-333. doi:10.1016/j.it.2005.04.008. PubMed: 15922949. [DOI] [PubMed] [Google Scholar]
- 4. Magalhaes JG, Tattoli I, Girardin SE (2007) The intestinal epithelial barrier: how to distinguish between the microbial flora and pathogens. Semin Immunol 19: 106-115. doi:10.1016/j.smim.2006.12.006. PubMed: 17324587. [DOI] [PubMed] [Google Scholar]
- 5. Sartor RB (2008) Microbial influences in inflammatory bowel diseases. Gastroenterology 134: 577-594. doi:10.1053/j.gastro.2007.11.059. PubMed: 18242222. [DOI] [PubMed] [Google Scholar]
- 6. Tlaskalová-Hogenová H, Stepánková R, Hudcovic T, Tucková L, Cukrowska B et al. (2004) Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunol Lett 93: 97-108. doi:10.1016/j.imlet.2004.02.005. PubMed: 15158604. [DOI] [PubMed] [Google Scholar]
- 7. Weinstock JV, Elliott DE (2009) Helminths and the IBD hygiene hypothesis. Inflamm Bowel Dis 15: 128-133. doi:10.1002/ibd.20633. PubMed: 18680198. [DOI] [PubMed] [Google Scholar]
- 8. Summers RW, Elliott DE, Urban JF Jr., Thompson RA, Weinstock JV (2005) Trichuris suis therapy for active ulcerative colitis: a randomized controlled trial. Gastroenterology 128: 825-832. doi:10.1053/j.gastro.2005.01.005. PubMed: 15825065. [DOI] [PubMed] [Google Scholar]
- 9. Whelan RA, Hartmann S, Rausch S (2012) Nematode modulation of inflammatory bowel disease. Protoplasma 249: 871-886. doi:10.1007/s00709-011-0342-x. PubMed: 22086188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Wohlgemuth S, Haller D, Blaut M, Loh G (2009) Reduced microbial diversity and high numbers of one single Escherichia coli strain in the intestine of colitic mice. Environ Microbiol 11: 1562-1571. doi:10.1111/j.1462-2920.2009.01883.x. PubMed: 19245530. [DOI] [PubMed] [Google Scholar]
- 11. Garrett WS, Gallini CA, Yatsunenko T, Michaud M, DuBois A et al. (2010) Enterobacteriaceae act in concert with the gut microbiota to induce spontaneous and maternally transmitted colitis. Cell Host Microbe 8: 292-300. doi:10.1016/j.chom.2010.08.004. PubMed: 20833380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Lupp C, Robertson ML, Wickham ME, Sekirov I, Champion OL et al. (2007) Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe 2: 119-129. doi:10.1016/j.chom.2007.06.010. PubMed: 18005726. [DOI] [PubMed] [Google Scholar]
- 13. Sun L, Nava GM, Stappenbeck TS (2011) Host genetic susceptibility, dysbiosis, and viral triggers in inflammatory bowel disease. Curr Opin Gastroenterol 27: 321-327. doi:10.1097/MOG.0b013e32834661b4. PubMed: 21483258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Heimesaat MM, Bereswill S, Fischer A, Fuchs D, Struck D et al. (2006) Gram-negative bacteria aggravate murine small intestinal Th1-type immunopathology following oral infection with Toxoplasma gondii. J Immunol 177: 8785-8795. PubMed: 17142781. [DOI] [PubMed] [Google Scholar]
- 15. Bereswill S, Fischer A, Plickert R, Haag LM, Otto B et al. (2011) Novel murine infection models provide deep insights into the "menage a trois" of Campylobacter jejuni, microbiota and host innate immunity. PLOS ONE 6: e20953. doi:10.1371/journal.pone.0020953. PubMed: 21698299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Heimesaat MM, Nogai A, Bereswill S, Plickert R, Fischer A et al. (2010) MyD88/TLR9 mediated immunopathology and gut microbiota dynamics in a novel murine model of intestinal graft-versus-host disease. Gut 59: 1079-1087. doi:10.1136/gut.2009.197434. PubMed: 20639251. [DOI] [PubMed] [Google Scholar]
- 17. Rausch S, Huehn J, Loddenkemper C, Hepworth MR, Klotz C et al. (2009) Establishment of nematode infection despite increased Th2 responses and immunopathology after selective depletion of Foxp3+ cells. Eur J Immunol 39: 3066-3077. doi:10.1002/eji.200939644. PubMed: 19750483. [DOI] [PubMed] [Google Scholar]
- 18. Heimesaat MM, Fischer A, Jahn HK, Niebergall J, Freudenberg M et al. (2007) Exacerbation of murine ileitis by Toll-like receptor 4 mediated sensing of lipopolysaccharide from commensal Escherichia coli. Gut 56: 941-948. doi:10.1136/gut.2006.104497. PubMed: 17255219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Heimesaat MM, Fischer A, Siegmund B, Kupz A, Niebergall J et al. (2007) Shift towards pro-inflammatory intestinal bacteria aggravates acute murine colitis via Toll-like receptors 2 and 4. PLOS ONE 2: e662. doi:10.1371/journal.pone.0000662. PubMed: 17653282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Shea-Donohue T, Sullivan C, Finkelman FD, Madden KB, Morris SC et al. (2001) The role of IL-4 in Heligmosomoides polygyrus-induced alterations in murine intestinal epithelial cell function. J Immunol 167: 2234-2239. PubMed: 11490010. [DOI] [PubMed] [Google Scholar]
- 21. Su CW, Cao Y, Kaplan J, Zhang M, Li W et al. (2011) Duodenal helminth infection alters barrier function of the colonic epithelium via adaptive immune activation. Infect Immun 79: 2285-2294. doi:10.1128/IAI.01123-10. PubMed: 21444669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Zhao A, McDermott J, Urban JF Jr., Gause W, Madden KB et al. (2003) Dependence of IL-4, IL-13, and nematode-induced alterations in murine small intestinal smooth muscle contractility on Stat6 and enteric nerves. J Immunol 171: 948-954. PubMed: 12847266. [DOI] [PubMed] [Google Scholar]
- 23. Erridge C, Duncan SH, Bereswill S, Heimesaat MM (2010) The induction of colitis and ileitis in mice is associated with marked increases in intestinal concentrations of stimulants of TLRs. p. 2, PLOS ONE 5: [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Finney CA, Taylor MD, Wilson MS, Maizels RM (2007) Expansion and activation of CD4(+)CD25(+) regulatory T cells in Heligmosomoides polygyrus infection. Eur J Immunol 37: 1874-1886. doi:10.1002/eji.200636751. PubMed: 17563918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Stecher B, Robbiani R, Walker AW, Westendorf AM, Barthel M et al. (2007) Salmonella enterica serovar typhimurium exploits inflammation to compete with the intestinal microbiota. PLOS Biol 5: 2177-2189. PubMed: 17760501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Darfeuille-Michaud A, Boudeau J, Bulois P, Neut C, Glasser AL et al. (2004) High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 127: 412-421. doi:10.1053/j.gastro.2004.04.061. PubMed: 15300573. [DOI] [PubMed] [Google Scholar]
- 27. Swidsinski A, Ladhoff A, Pernthaler A, Swidsinski S, Loening-Baucke V et al. (2002) Mucosal flora in inflammatory bowel disease. Gastroenterology 122: 44-54. doi:10.1053/gast.2002.30294. PubMed: 11781279. [DOI] [PubMed] [Google Scholar]
- 28. Swidsinski A, Weber J, Loening-Baucke V, Hale LP, Lochs H (2005) Spatial organization and composition of the mucosal flora in patients with inflammatory bowel disease. J Clin Microbiol 43: 3380-3389. doi:10.1128/JCM.43.7.3380-3389.2005. PubMed: 16000463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Lee IA, Bae EA, Hyun YJ, Kim DH (2010) Dextran sulfate sodium and 2,4,6-trinitrobenzene sulfonic acid induce lipid peroxidation by the proliferation of intestinal gram-negative bacteria in mice. J Inflamm (Lond) 7: 7 PubMed: 20181058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Walk ST, Blum AM, Ewing SA, Weinstock JV, Young VB (2010) Alteration of the murine gut microbiota during infection with the parasitic helminth Heligmosomoides polygyrus. Inflamm Bowel Dis 16: 1841-1849. doi:10.1002/ibd.21299. PubMed: 20848461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Kruidenier L, Kuiper I, Lamers CB, Verspaget HW (2003) Intestinal oxidative damage in inflammatory bowel disease: semi-quantification, localization, and association with mucosal antioxidants. J Pathol 201: 28-36. doi:10.1002/path.1409. PubMed: 12950014. [DOI] [PubMed] [Google Scholar]
- 32. Sundaram U, Hassanain H, Suntres Z, Yu JG, Cooke HJ et al. (2003) Rabbit chronic ileitis leads to up-regulation of adenosine A1/A3 gene products, oxidative stress, and immune modulation. Biochem Pharmacol 65: 1529-1538. doi:10.1016/S0006-2952(03)00067-4. PubMed: 12732366. [DOI] [PubMed] [Google Scholar]
- 33. Sundaram U, Coon S, Wisel S, West AB (1999) Corticosteroids reverse the inhibition of Na-glucose cotransport in the chronically inflamed rabbit ileum. Am J Physiol 276: G211-G218. PubMed: 9886998. [DOI] [PubMed] [Google Scholar]
- 34. Li RW, Wu S, Li W, Navarro K, Couch RD et al. (2012) Alterations in the porcine colon microbiota induced by the gastrointestinal nematode Trichuris suis. Infect Immun 80: 2150-2157. doi:10.1128/IAI.00141-12. PubMed: 22493085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Wu S, Li RW, Li W, Beshah E, Dawson HD et al. (2012) Worm burden-dependent disruption of the porcine colon microbiota by Trichuris suis infection. PLOS ONE 7: e35470. doi:10.1371/journal.pone.0035470. PubMed: 22532855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. D’Elia R, DeSchoolmeester ML, Zeef LA, Wright SH, Pemberton AD et al. (2009) Expulsion of Trichuris muris is associated with increased expression of angiogenin 4 in the gut and increased acidity of mucins within the goblet cell. BMC Genomics 10: 492. doi:10.1186/1471-2164-10-492. PubMed: 19852835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Steenwinckel V, Louahed J, Lemaire MM, Sommereyns C, Warnier G et al. (2009) IL-9 promotes IL-13-dependent paneth cell hyperplasia and up-regulation of innate immunity mediators in intestinal mucosa. J Immunol 182: 4737-4743. doi:10.4049/jimmunol.0801941. PubMed: 19342650. [DOI] [PubMed] [Google Scholar]
- 38. Hooper LV, Stappenbeck TS, Hong CV, Gordon JI (2003) Angiogenins: a new class of microbicidal proteins involved in innate immunity. Nat Immunol 4: 269-273. doi:10.1038/ni888. PubMed: 12548285. [DOI] [PubMed] [Google Scholar]
- 39. Iimura M, Gallo RL, Hase K, Miyamoto Y, Eckmann L et al. (2005) Cathelicidin mediates innate intestinal defense against colonization with epithelial adherent bacterial pathogens. J Immunol 174: 4901-4907. PubMed: 15814717. [DOI] [PubMed] [Google Scholar]
- 40. Ouellette AJ (2011) Paneth cell alpha-defensins in enteric innate immunity. Cell Mol Life Sci 68: 2215-2229. doi:10.1007/s00018-011-0714-6. PubMed: 21560070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Rausch S, Held J, Stange J, Lendner M, Hepworth MR et al. (2010) A matter of timing: early, not chronic phase intestinal nematode infection restrains control of a concurrent enteric protozoan infection. Eur J Immunol 40: 2804-2815. doi:10.1002/eji.201040306. PubMed: 20809519. [DOI] [PubMed] [Google Scholar]
- 42. Chen CC, Louie S, McCormick B, Walker WA, Shi HN (2005) Concurrent infection with an intestinal helminth parasite impairs host resistance to enteric Citrobacter rodentium and enhances Citrobacter-induced colitis in mice. Infect Immun 73: 5468-5481. doi:10.1128/IAI.73.9.5468-5481.2005. PubMed: 16113263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Fox JG, Beck P, Dangler CA, Whary MT, Wang TC et al. (2000) Concurrent enteric helminth infection modulates inflammation and gastric immune responses and reduces Helicobacter-induced gastric atrophy. Nat Med 6: 536-542. doi:10.1038/75015. PubMed: 10802709. [DOI] [PubMed] [Google Scholar]
- 44. Elliott DE, Setiawan T, Metwali A, Blum A, Urban JF Jr. et al. (2004) Heligmosomoides polygyrus inhibits established colitis in IL-10-deficient mice. Eur J Immunol 34: 2690-2698. doi:10.1002/eji.200324833. PubMed: 15368285. [DOI] [PubMed] [Google Scholar]
- 45. Metwali A, Setiawan T, Blum AM, Urban J, Elliott DE et al. (2006) Induction of CD8+ regulatory T cells in the intestine by Heligmosomoides polygyrus infection. Am J Physiol Gastrointest Liver Physiol 291: G253-G259. doi:10.1152/ajpgi.00409.2005. PubMed: 16825660. [DOI] [PubMed] [Google Scholar]
- 46. Setiawan T, Metwali A, Blum AM, Ince MN, Urban JF Jr. et al. (2007) Heligmosomoides polygyrus promotes regulatory T-cell cytokine production in the murine normal distal intestine. Infect Immun 75: 4655-4663. doi:10.1128/IAI.00358-07. PubMed: 17606601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Sutton TL, Zhao A, Madden KB, Elfrey JE, Tuft BA et al. (2008) Anti-Inflammatory mechanisms of enteric Heligmosomoides polygyrus infection against trinitrobenzene sulfonic acid-induced colitis in a murine model. Infect Immun 76: 4772-4782. doi:10.1128/IAI.00744-07. PubMed: 18644879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Ince MN, Elliott DE, Setiawan T, Blum A, Metwali A et al. (2006) Heligmosomoides polygyrus induces TLR4 on murine mucosal T cells that produce TGFbeta after lipopolysaccharide stimulation. J Immunol 176: 726-729. PubMed: 16393954. [DOI] [PubMed] [Google Scholar]
- 49. Broadhurst MJ, Ardeshir A, Kanwar B, Mirpuri J, Gundra UM et al. (2012) Therapeutic helminth infection of macaques with idiopathic chronic diarrhea alters the inflammatory signature and mucosal microbiota of the colon. PLOS Pathog 8: e1003000 PubMed: 23166490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Lee DH, Zo YG, Kim SJ (1996) Nonradioactive method to study genetic profiles of natural bacterial communities by PCR-single-strand-conformation polymorphism. Appl Environ Microbiol 62: 3112-3120. PubMed: 8795197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Van Dyke MI, McCarthy AJ (2002) Molecular biological detection and characterization of Clostridium populations in municipal landfill sites. Appl Environ Microbiol 68: 2049-2053. doi:10.1128/AEM.68.4.2049-2053.2002. PubMed: 11916731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Matsuki T, Watanabe K, Fujimoto J, Miyamoto Y, Takada T et al. (2002) Development of 16S rRNA-gene-targeted group-specific primers for the detection and identification of predominant bacteria in human feces. Appl Environ Microbiol 68: 5445-5451. doi:10.1128/AEM.68.11.5445-5451.2002. PubMed: 12406736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Bartosch S, Fite A, Macfarlane GT, McMurdo ME (2004) Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl Environ Microbiol 70: 3575-3581. doi:10.1128/AEM.70.6.3575-3581.2004. PubMed: 15184159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Kühbacher T, Ott SJ, Helwig U, Mimura T, Rizzello F et al. (2006) Bacterial and fungal microbiota in relation to probiotic therapy (VSL#3) in pouchitis. Gut 55: 833-841. doi:10.1136/gut.2005.078303. PubMed: 16401690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Heilig HG, Zoetendal EG, Vaughan EE, Marteau P, Akkermans AD et al. (2002) Molecular diversity of Lactobacillus spp. and other lactic acid bacteria in the human intestine as determined by specific amplification of 16S ribosomal DNA. Appl Environ Microbiol 68: 114-123. doi:10.1128/AEM.68.1.114-123.2002. PubMed: 11772617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Walter J, Hertel C, Tannock GW, Lis CM, Munro K et al. (2001) Detection of Lactobacillus, Pediococcus, Leuconostoc, and Weissella species in human feces by using group-specific PCR primers and denaturing gradient gel electrophoresis. Appl Environ Microbiol 67: 2578-2585. doi:10.1128/AEM.67.6.2578-2585.2001. PubMed: 11375166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Rinttilä T, Kassinen A, Malinen E, Krogius L, Palva A (2004) Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J Appl Microbiol 97: 1166-1177. doi:10.1111/j.1365-2672.2004.02409.x. PubMed: 15546407. [DOI] [PubMed] [Google Scholar]
- 58. Barman M, Unold D, Shifley K, Amir E, Hung K et al. (2008) Enteric salmonellosis disrupts the microbial ecology of the murine gastrointestinal tract. Infect Immun 76: 907-915. doi:10.1128/IAI.01432-07. PubMed: 18160481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Wise MG, Siragusa GR (2007) Quantitative analysis of the intestinal bacterial community in one- to three-week-old commercially reared broiler chickens fed conventional or antibiotic-free vegetable-based diets. J Appl Microbiol 102: 1138-1149. PubMed: 17381758. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
DGGE analysis of luminal contents of (A) ileum, (B) cecum and (C) colon. Band profiles shown derive from 3 naïve and infected mice at different time points after infection. Arrowheads mark additional bands during acute H . p . bakeri infection (14 d.p.i.). M: marker.
(TIF)
DGGE profiles and results of sequencing analysis of naive versus acutely infected mice. SYBR green stained gels of extracted DNA from (A) ileum and (B) cecum were used for DNA extraction and sequencing of the marked bands.
(TIF)