

Abstract
To help understand the elusive mechanisms of zebrafish sex determination, we studied the genetic machinery regulating production and breakdown of retinoic acid (RA) during the onset of meiosis in gonadogenesis. Results uncovered unexpected mechanistic differences between zebrafish and mammals. Conserved synteny and expression analyses revealed that cyp26a1 in zebrafish and its paralog Cyp26b1 in tetrapods independently became the primary genes encoding enzymes available for gonadal RA-degradation, showing lineage-specific subfunctionalization of vertebrate genome duplication (VGD) paralogs. Experiments showed that zebrafish express aldh1a2, which encodes an RA-synthesizing enzyme, in the gonad rather than in the mesonephros as in mouse. Germ cells in bipotential gonads of all zebrafish analyzed were labeled by the early meiotic marker sycp3, suggesting that in zebrafish, the onset of meiosis is not sexually dimorphic as it is in mouse and is independent of Stra8, which is required in mouse but was lost in teleosts. Analysis of dead-end knockdown zebrafish depleted of germ cells revealed the germ cell-independent onset and maintenance of gonadal aldh1a2 and cyp26a1 expression. After meiosis initiated, somatic cell expression of cyp26a1 became sexually dimorphic: up-regulated in testes but not ovaries. Meiotic germ cells expressing the synaptonemal complex gene sycp3 occupied islands of somatic cells that lacked cyp26a1 expression, as predicted by the hypothesis that Cyp26a1 acts as a meiosis-inhibiting factor. Consistent with this hypothesis, females up-regulated cyp26a1 in oocytes that entered prophase-I meiotic arrest, and down-regulated cyp26a1 in oocytes resuming meiosis. Co-expression of cyp26a1 and the pluripotent germ cell stem cell marker pou5f1(oct4) in meiotically arrested oocytes was consistent with roles in mouse to promote germ cell survival and to prevent apoptosis, mechanisms that are central for tipping the sexual fate of gonads towards the female pathway in zebrafish.
Introduction
A critical stage in vertebrate sex determination is entry of the bipotential gonadal primordium into a developmental pathway leading to ovary or testis (reviewed in 1). Sex-specific programs depend upon cell signaling between developing germ cells and somatic cells. Discovery of Sry (sex-determining region Y), the major sex-determining gene in mammals [2,3], stimulated the search for genetic mechanisms that control the sexual fate of somatic and germ cells during vertebrate gonadogenesis. In some vertebrates, however, such as zebrafish, developmental genetic mechanisms that control sexual fate remain unknown [4–9], and the identification of sex-biasing genes has been elusive [10,11]. For a zebrafish stock recently derived from India, a major sex determinant was identified on a chromosome that has several features of a sex chromosome [12], but genetic analyses of several laboratory strains identified different polygenic factors in different strains [12–15].
In mammals, male and female germ cells enter meiosis at different developmental stages (reviewed in 16–19). In female embryonic mouse gonads, germ cells enter into meiosis at 13.5 days post-coitum (dpc) and concomitantly, the somatic gonadal primordium initiates ovarian differentiation by developing into granulosa and theca cells. In contrast, in male embryonic mouse gonads, SRY-expressing pre-Sertoli cells initiate testicular differentiation and germ cells arrest in the G0/G1 phase of the mitotic cell cycle, postponing meiosis until after birth [20,21]. Furthermore, meiotic XX germ cells antagonize testicular development in XY gonads in tissue co-culture experiments, leading to the hypothesis that germ cells committed to meiosis reinforce ovarian fate by antagonizing the testis pathway [22].
Recent studies showed that in mouse, retinoic acid (RA) in females or its absence in males -- regulated by the RA-destroying Cyp26 Cytochrome P450 enzyme family -- reinforces germ cell fate by controlling the sex-specific initiation of meiosis ( [23–27] reviewed in 18,28; although see 29 for an alternative hypothesis). Retinoic acid is an active derivative of vitamin A that diffuses through tissues and binds to heterodimers of the nuclear receptors RAR-RXR, which recognize RA-response elements (RAREs) in DNA to control the expression of RA-target genes [30–33]. Because Rar and Rxr are widely expressed in embryos, the regulation of RA action often occurs at the metabolic level, in which the balance of RA-synthesizing enzymes (i.e. the Aldh1a retinaldehyde dehydrogenase family) and RA-degrading enzymes (i.e. members of the Cyp26 P450-cytochrome family) determines the precise spatial-temporal distribution of RA [34]. This is the case in mouse gonadogenesis, in which male-specific up-regulation of Cyp26b1 expression leads to degradation of RA and protects germ cells from entering into meiosis in developing testes, while female-specific down-regulation of Cyp26b1 expression allows RA to induce germ cells to enter into meiosis in embryonic ovaries [23,24,35]. The central role of Cyp26b1 in preventing entry into meiosis was further supported by evidence showing that disruption of Cyp26b1 expression in embryonic mouse testes, or addition of CYP inhibitors to wild-type embryonic testes, induced germ cells to express the pre-meiotic marker Stra8 (stimulated by retinoic acid-8), which is required for the initiation of meiosis in mammals, followed by the expression of the early meiotic marker Sycp3 (synaptonemal complex protein 3), and the down-regulation of the pluripotent stem cell marker Pou5f1 (POU class 5 homeobox 1, or Oct4) [23,24,35–45].
In mice, expression of the RA-synthesizing enzyme Aldh1a2 in the embryonic mesonephros (but not in the gonads) as females initiate meiosis led to the suggestion of a source-sink regulatory system. According to this model, RA synthesized in the mesonephros enters the neighboring gonad and causes germ cells to enter meiosis in embryonic ovaries, which lack the RA-degrading enzyme Cyp26 ( [23,24,42]; reviewed in 18). Recently, male-specific expression of the Aldh1a2 paralog Aldh1a1 has been reported in somatic cells of embryonic mouse testes [46]. The expression of Aldh1a1, however, has been suggested to act as a buffer to maintain low levels of RA that might be needed for general testis morphogenesis rather than the high levels of RA needed for germ cells to enter meiosis [46].
RA plays a role in the onset of meiosis not only in mammals, but also in other tetrapods, including birds and amphibians [47–49]. Whether the role of RA during gonad development is a tetrapod innovation, however, or whether it is shared with other non-tetrapod vertebrates, including teleost fishes, remains unknown [50]. Similarities and differences in the mechanisms of gonadogenesis in teleosts and tetrapods suggest the question: Is RA action important for meiosis and gonadogenesis in zebrafish? Several considerations motivate this problem. First, in contrast to mouse, the gonads of a zebrafish do not lie adjacent to the mesonephros during the critical period for gonadal sex determination; consequently the source-sink regulatory system from the mesonephros to the gonad postulated in mouse is unlikely to apply to zebrafish. Second, in contrast to mouse, all zebrafish juveniles, regardless of their definitive sex, initially develop an ovary-like gonad with immature oocytes; in females, these oocytes continue to develop and reinforce the differentiation of mature ovaries, but in males, oocytes die by apoptosis and the gonads become testes [4,5,7,8,51–53]. We do not yet know, however, whether differences in the timing of the onset of meiosis exist between zebrafish males and females. Third, genomic surveys of the RA-metabolic genetic machinery have shown that some aldh1a family genes (i.e. aldh1a1) have been lost in zebrafish and other teleosts, and that this gene loss has altered the functional evolution of the surviving aldh1a paralogs [54–57]. Whether these gene losses had functional consequences in gonad development is not yet known.
To address these questions and to test the hypothesis that meiotic control mechanisms in mouse apply to zebrafish, we performed a comprehensive genomic and expression analysis of the genetic machinery that regulates the synthesis and degradation of RA during gonadogenesis, and studied genetic markers for meiosis and somatic cells of the gonad to investigate the role of RA in the onset of meiosis and sexual fate determination in zebrafish. Results revealed shared underlying regulatory themes between zebrafish and mammals but important genomic and developmental differences in the mechanisms of RA-regulated gonadogenesis and sex determination.
Results
1. In zebrafish, cyp26a1, rather than Cyp26b1 as in mammals, is the main RA-degradation gene expressed during gonad development
Tetrapods and zebrafish have three Cyp26 paralogs: Cyp26a1, Cyp26b1, and Cyp26c1. Recent studies in mouse [23,24], chicken [47,49] and urodele amphibians [48] implicated the RA-degrading enzyme Cyp26b1 as the pivotal sex-specific factor regulating the differential timing of meiotic onset in females and males. Our first question was whether male-specific up-regulation of cyp26b1 occurs during gonad development in zebrafish as it does in tetrapods.
To address this question, we investigated expression patterns of cyp26b1, as well as the other two gene family members cyp26a1 and cyp26c1, in adjacent histological sections of developing zebrafish gonads by in situ hybridization (ISH) (Figure 1A–I). Zebrafish cyp26a1 expression was detected in several cells in the gonadal primordium at 15 dpf before the critical period for sex determination (Figure 1A), became more abundant and mostly restricted to the dorsal surface of the gonad close to the body cavity by 19 dpf during early sex differentiation (Figure 1D), and was highly up-regulated and broadly expanded by 31 dpf during the immature testis stage (Figure 1G). This result revealed that cyp26a1 was expressed in zebrafish gonads early and continuously through the critical time window for gonadal sex determination (19 dpf - 31dpf) [4,5,8,58–60]. Unexpectedly, expression of zebrafish cyp26b1 was detected only in a few scattered cells in undifferentiated gonads at 15 dpf (Figure 1B), and in contrast to its ortholog in tetrapods, cyp26b1 was not detected in gonads of animals at the stages in which sex determination occurs (19 dpf -31 dpf, Figure 1E,H). No cyp26c1 expression was detected in developing gonads at any stage analyzed (Figure 1C,F,I), although positive controls did show cyp26c1 expression in the hindbrain of zebrafish embryos (data not shown) and in the retina of 15 dpf juveniles. These results revealed that, in contrast to tetrapods, in which cyp26b1 is the major regulator of RA availability during early gonad development, cyp26a1 is the only paralog up-regulated in developing zebrafish gonads during the time of sexual differentiation of the gonads.
Figure 1. In zebrafish, cyp26a1 is the main Cyp26 paralog expressed in gonads during the critical period for sex-determination.

Undifferentiated gonads at 15 days post-fertilization (dpf) expressed cyp26a1 (A) and cyp26b1 (B), but did not express cyp26c1 at detectable levels (C) (A-C: n=4). In bipotential gonads at 19 dpf, cyp26a1 expression became restricted mainly to the dorsal margin of the gonad (D) but expression of cyp26b1 and cyp26c1 was not detected (E, F) (D–F: n=9). In differentiating testes at 31 dpf, cyp26a1 expression up-regulated (G) and in contrast to mouse testes, which up-regulate cyp26b1, neither cyp26b1 nor cyp26c1 expression was detected in maturing zebrafish gonads (H, I) (G-I: n=10). Differentiating testes were assigned by morphological features and assessed by the expression of the male specific Amh marker (see Figure 4). We conclude that in zebrafish, Cyp26a1 is expressed at the time and place necessary to provide an RA-degrading function equivalent to Cyp26b1 in tetrapods. These results suggest independent subfunction partitioning of ancestral cyp26 regulatory elements in lineages leading to zebrafish and mouse. Arrows point to examples of expressing cells. Dashed lines outline gonads. Scale bar: 0.05mm.
2. Genomic synteny conservation and independent subfunction evolution of the Cyp26 family in teleosts and tetrapods
The apparent convergence of the roles of Cyp26a1 in zebrafish and Cyp26b1 in mouse could have either of two explanations: first, it might represent a case of independent subfunctionalization of gene roles possessed by the ancestral Cyp26 gene before the two rounds of vertebrate genome duplications (alias VGD1 and VGD2); alternatively, it might be that zebrafish gene orthologies had been incorrectly assigned. To rule out the possibility of error in orthology assignments, we examined gene orthologies by first investigating the phylogenetic relationships between zebrafish Cyp26a1 [61], Cyp26b1 [62] and Cyp26c1 [63] (originally called Cyp26d1 [64]) and their respective mouse paralogs [65–67]. Phylogenetic analysis by Maximum-likelihood showed that the three Cyp26 paralogs of various teleosts (zebrafish, medaka, stickleback and fugu) grouped within the clades of their correspondingly named tetrapod Cyp26 paralogs with high bootstrap support (Figure 2A), corroborating the recently proposed Cyp26 nomenclature [63].
Figure 2. Evolutionary relationships of CYP26 family members in zebrafish and mouse.

(A) Phylogenetic analyses inferred by maximum likelihood (ML) indicate that Cyp26 paralogs of teleosts (i.e. zebrafish (Dre), medaka (Ola), stickleback (Gac) and fugu (Tru)) grouped with their correspondingly named Cyp26 paralogs of tetrapods (i.e. mouse (Mmu), human (Hsa) and chicken (Gga)). Numbers indicate bootstrap values supporting each node (n=100), and no significant differences were found between ML and NJ analyses. (B) A circleplot shows graphically orthologous relationships of cyp26 genomic neighborhoods shared between zebrafish and mouse. Grey circumferential arcs represent chromosomes, with Cyp26b1 in green on Mmu6, Cyp26a1 in red on Mmu19, and Cyp26c1 in blue on Mmu19. Colored lines link orthologous regions in zebrafish chromosomes Dre7, Dre12 and Dre17, the sites of cyp26b1, cyp26a1 and cyp26c1 genes, respectively. (C–E) Clusters of synteny conservation reveal the presence of many gene neighbor orthologs shared between each Cyp26 genomic neighborhood in mouse and zebrafish cyp26b1 (C: cluster ID#265419 according to the Synteny Database), cyp26a1 (D: cluster ID#258723) and cyp26c1 (E: cluster ID#265367). These results rule out the possibility of reciprocal gene losses in zebrafish and mouse that could mask actual orthologous relationships in artifactual phylogenetic trees, and provide strong support for the conclusion that the zebrafish/mouse gene pairs called cyp26a1/Cyp26a1 and cyp26b1/Cyp26b1 are in fact orthologs. F: Gene clusters of synteny conservation (cluster ID#191383) between Dre12 and Dre17 reveal that the genomic neighborhoods of cyp26a1 and cyp26c1 are paralogous due to the teleost genome duplication (TGD, R3) that preceded the divergence of the teleost lineage after it split from the tetrapod branch. Duplicates of cyp26a1 and cyp26c1 in Dre17 and Dre12 were lost reciprocally after the teleost genome duplication (labeled with crosses) in contrast to, for instance, reep3 paralogs (in pink) that were maintained adjacent to cyp26 paralogs.
Phylogenetic analyses can sometimes give erroneous orthology assignments in cases of reciprocal gene loss after ancient gene duplications [68–70]; for example, four Cyp26 genes would have been produced in VGD1 and VGD2 [71], but zebrafish and tetrapods both have three rather than four Cyp26 genes, raising the possibility that different VGD paralogs were lost in the teleost and tetrapod lineages.
To critically test the hypothesis that reciprocal gene loss occurred in the Cyp26 family in the zebrafish and mouse lineages leading to erroneous orthology assignments, we examined a data set independent of Cyp26 amino acid sequence by examining syntenic conservation within the genomic neighborhoods (GN) surrounding Cyp26 genes in zebrafish and mouse (Figure 2B–F) and medaka (Figure S1, Figure S2). In mouse, the three Cyp26 paralogs reside on two chromosomes: Cyp26b1 is on mouse chromosome 6 (Mmu6), and Cyp26a1 and Cyp26c1 are adjacent and transcribed in the same orientation on Mmu19, consistent with the origin of Cyp26a1 and Cyp26c1 by tandem duplication rather than by genome duplication. In teleosts, however, the three Cyp26 paralogs are on three different chromosomes (i.e. cyp26a1 on zebrafish (Danio rerio) linkage group 12 (Dre12) and on medaka (Oryzias latipes) chromosome 19 (Ola19); cyp26b1 on Dre7 and Ola18; and cyp26c1 on Dre17 and Ola15). Genomic analysis at the chromosomal level using circleplots [69] revealed orthology relationships between each Cyp26 chromosomal neighborhood in mouse with its corresponding orthologous chromosomal region in zebrafish (Figure 2B) and medaka (Figure S2A): The mouse Cyp26b1 genomic neighborhood connected to teleost cyp26b1 neighborhoods; and the mouse Cyp26a1/Cyp26c1 neighborhood connected to teleost neighborhoods that harbor cyp26a or cyp26c1, respectively (Figure 2B and Figure S2A). Local analysis of each Cyp26 genomic neighborhood using the Synteny Database (version Ens56 [69]) identified conserved gene neighbors near each Cyp26 ortholog: Cyp26b1 with Dysf (Figure 2C and Figure S2B), cyp26a1 with Pde6c (Figure 2D and Figure S2C), and cyp26c1 with Tbc1d12 (Figure 2E and Figure S2D). These results provide robust evidence that rules out the hypothesis that reciprocal Cyp26 paralog loss occurred between zebrafish and mouse.
Comparative analysis between zebrafish genomic neighborhoods surrounding cyp26a1 in Dre12 and cyp26c1 in Dre17 revealed that these two regions are indeed paralogons (Figure 2F). This finding suggests the hypothesis that prior to the tetrapod-teleost divergence, cyp26a1 and cyp26c1 were already adjacent in an ancestral chromosome due to a tandem duplication, and that after the teleost genome duplication (TGD) event (reviewed in 72), cyp26a1 and cyp26c1 duplicates survived reciprocally in each paralogon, leading to the present situation in which cyp26a1 and cyp26c1 are on different teleost chromosomes (Figure 2F and Figure S2E).
Overall, our phylogenetic and comparative genomic analyses provided robust evidence that rules out the hypothesis of reciprocal Cyp26 gene losses in the zebrafish and mouse lineages and supports the recently postulated Cyp26 ontogeny [63]. We conclude, therefore, that the most parsimonious explanation is that the Cyp26 pro-ortholog predating the expansion of the gene family was already involved in the regulation of RA levels during gonad development, and that both Cyp26a1 and Cyp26b1 maintained this function in the last common ancestor of zebrafish and mouse. Subsequently, independent subfunction partitioning [72,73] likely led to the reciprocal retention of the gonad function of Cyp26a1 in the lineage leading to zebrafish and Cyp26b1 in the tetrapod lineage.
We wondered whether independent partitioning of subfunctions of the ancestral Cyp26 gene that were unrelated to the gonad occurred in the same manner as subfunction partitioning in the gonad. Comparative analysis of the three cyp26 paralogs during late eye development in zebrafish (Figure 3A) and mouse [74] revealed that the zebrafish ortholog of each of these three genes has the same expression pattern in the retina as its mouse ortholog. This result suggests that at least some ancestral subfunctions (i.e. in the retina) partitioned the same way in the three Cyp26 genes, perhaps before the divergence of zebrafish and mouse lineages, but the gonadal subfunction partitioned reciprocally in the two lineages after they diverged (Figure 3B).
Figure 3. Expression patterns of cyp26 gene family members in late developing retina are conserved between zebrafish and mouse.
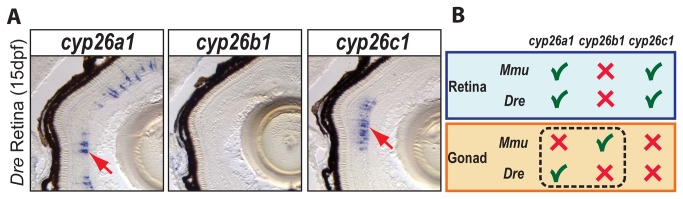
To learn if the divergent subfunction partitioning between zebrafish cyp26a1 and mouse cyp26b1 in gonad development applies to other organs, we studied the expression of cyp26 paralogs during retina development in 15-dpf zebrafish. Results revealed expression of cyp26a1 and cyp26c1 in different layers of the retina, but no detectable expression of cyp26b1 (A: n=1). This result agrees with the Cyp26 expression profile described in mouse retina (B), in which Cyp26a1 and Cyp26c1 genes were expressed in the inner nuclear layer during post-natal eye development while Cyp26b1 was not expressed in the eye [74]. We conclude that independent subfunction partitioning related to gonad development occurred after the teleost-tetrapod lineage divergence, while subfunctions related to at least one other organ, the retina, are still conserved between Cyp26 orthologs in different vertebrate lineages (B).
3. Comparative expression analysis of genes encoding RA-metabolic machinery and somatic and germ cell markers during zebrafish gonadogenesis
To learn whether RA-producing and RA-degrading enzymes are sexually dimorphic in developing gonads, we compared male and female expression patterns of cyp26 genes, which encode RA degrading enzymes, and the two zebrafish aldh1a paralogs (aldh1a2, and aldh1a3), which encode RA synthesizing enzymes. Because no robust genetic marker for sex is yet available for zebrafish [12,13], to assess sexual fate of zebrafish gonads, we examined expression of the early male-specific somatic marker amh (anti-Mullerian hormone) [59] and the germ cell-specific marker vasa [75]. We performed in situ hybridization analyses on adjacent gonadal sections from animals representing three key stages of gonad development: i) bipotential gonads (20 days post-fertilization (dpf)) when gonads are sexually undifferentiated and can become either ovary or testis (Figure 4 A–E); ii) transitional gonads (26 dpf), when gonads are in the process of transitioning from the juvenile ovary-like stage to enter the female or male pathway and differentiate into ovaries or testes (Figure 4F–O); and iii) differentiated but immature gonads (33 dpf and 41 dpf), after sexual fate selection but before gametes mature (Figure 4P–Y and Z–I’ respectively).
Figure 4. Expression of genes encoding enzymes for the synthesis and degradation of RA during zebrafish gonad development.

In situ hybridization on adjacent sections of animals representing the three key stages of gonad development: (A–E: n=7) bipotential, sexually undifferentiated gonads with an ovary-like morphology at 20 days post-fertilization (dpf); (F–O) gonads transitioning to testes or ovaries (26 dpf) (F-J: n=4; K–O: n=3); (P–Y and Z–I’) gonads sexually differentiated but still immature (33 dpf and 41 dpf) (P–T: n=3; U–Y: n=4; Z-D’: n=2; E’–I’: n=2). Images show expression patterns of the gene encoding the RA-synthesizing enzyme Aldh1a2 (A, F, K, P, U, Z, E’), the gene encoding the RA-degrading enzyme Cyp26a1 (C, H, M, R, W, B’,G’) and combined probes for cyp26b1 and cyp26c1 (E, J, O, T, Y, D’, I’) together with the early male gonadal marker amh (anti-Müllerian hormone: B, G, L, Q, V & A’, F’) and the germ-line specific marker vasa (D, I, N, S, X, C’, H’) at four different stages. Expression of aldh1a2 was detected in somatic cells in both male and female gonads throughout development (A, F, K, P, U, Z, E’). Expression of aldh1a3 was not detected at all in gonads but was detected in retina cells (data not shown) and the ortholog of Aldh1a1 was lost in the teleost lineage [54–56]. The expression pattern of cyp26a1, however, showed a distinct sexual dimorphism, as gonadal somatic cells from males (H, R and B’), but not from females (M, W, G’), up-regulated its expression during gonad development. Interestingly, in females, oocytes that had transitioned from early stage IB (yellow arrowhead in M,W,G’) to late stage IB (green arrowheads in M,W,G’) up-regulated the expression of cyp26a1 in the ooplasm, which was maintained at later stages (e.g. red arrowhead stage II in G’). The observed expression pattern of cyp26a1 in oocytes is compatible with a function in inhibiting meiotic progression and facilitating the meiotic arrest at diplotene stage. Expression of cyp26b1 and cyp26c1 in gonads was not detected at any of the stages analyzed in either sex (E, J, O, T, Y, D’, I’) Arrows point to examples of expressing cells. Scale bar shown per each row: 0.1mm.
In undifferentiated bipotential gonads at 20 dpf, aldh1a2 was expressed throughout most cells of the ovary-like gonad of juveniles, but was not expressed in the largest germ cells, which were probably developing oocytes judging by their morphology (Figure 4A). No gonadal expression was observed for zebrafish aldh1a3 (data not shown), which is the only other remaining aldh1a paralog in zebrafish due to the loss of aldh1a1 early in the evolution of the teleost lineage [56]. In bipotential gonads, amh was already expressed, but in only a few scattered somatic cells (Figure 4B) [7,8,59]. At 20 dpf, cyp26a1 maintained its expression at the surface of the gonad near the body cavity in all individuals (Figure 4C) as observed at 19 dpf (Figure 1D). Comparison of this thin superficial domain of cyp26a1 expression to the broadly distributed domain of vasa expression in germ cells (Figure 4D) suggested that cyp26a1 was expressed either in specific somatic cells or in a sub-set of germ cells (i.e. germ cell stem cells or gonia). Analysis by double fluorescent in situ hybridization in sections of bipotential gonads resolved this question, showing that cyp26a1 was expressed in somatic cells but not in germ cells labeled by vasa (Figure 5A,B). No expression of cyp26b1 or cyp26c1 was detected in the gonad at this undifferentiated stage (Figure 4E).
Figure 5. Three color fluorescent detection of cyp26a1 and vasa expression during zebrafish gonad development.

Fluorescence detection of cyp26a1 (green) expression and vasa (red) expression by double in situ hybridization on gonad sections at the bipotential gonad stage at 15 dpf (A: n=1) and 20 dpf (B: n=2), and on immature gonads developing into testes (C: n=1) or ovaries (D: n=1) at 33 dpf. Cyp26a1 expression occurs in somatic cells and does not co-localize with the germ cell marker vasa in bipotential gonads (A,B). In immature testes, cyp26a1 expression was also expressed in somatic cells (arrow) and not in germ cells (C). In immature ovaries, however, cyp26a1 was expressed in large oocytes that had reached diplotene (arrowhead in D).
Gonads transitioning from the juvenile ovary-like stage into definitive testis or ovary pathways (26dpf) showed broad expression of aldh1a2 in gonads of both sexes (Figure 4F,K). Sex assignment was based on gonad morphology and the presence or absence of expression of the early male-specific somatic marker amh (Figure 4G,L). Comparison of the expression pattern of aldh1a2 (Figure 4F,K) with the distribution of vasa-expressing germ cells (Figure 4I,N) suggested that the expression of aldh1a2 was mostly restricted to somatic cells.
In transitional gonads, cyp26a1 expression became strikingly sexually dimorphic. In males, expression of cyp26a1 had extended from its prior thin superficial expression domain to include cells broadly distributed throughout the gonad (Figure 4H), likely somatic cells judged by the position of vasa-positive germ cells (Figure 4I). In contrast to males, females did not show detectable cyp26a1 expression in somatic cells (Figure 4M). Instead, cyp26a1 signal in females appeared restricted to the ooplasm of few large oocytes that had progressed to the diplotene stage of meiosis (i.e. late stage IB oocytes (lIB); green arrowhead in Figure 4M), but not in small, early stage oocytes (i.e. early stage IB oocytes (eIB); yellow arrowhead in Figure 4M). Because meiosis arrests at diplotene in late stage IB zebrafish oocytes – as occurs in human diplotene oocytes for as much as 40 or 50 years -- the up-regulation of cyp26a1 in late stage IB is compatible with a potential role of Cyp26a1 in arresting oocytes during meiosis. Expression of cyp26b1 and cyp26c1 was not detected at this crucial stage for gonad fate decision in ovaries or testis (Figure 4J,O).
Gonads that had differentiated as immature ovaries or immature testes (33dpf and 41dpf) continued to express aldh1a2 in somatic cells of gonads of both sexes (Figure 4P,U,Z,E’). In males, aldh1a2 expression domain appeared to be restricted to a set of somatic cells surrounding testis cysts (Figure 4P,Z), judging from the broader expression of amh in developing Sertoli cells (Figure 4Q, A’). In females, aldh1a2 expression continued to be restricted to somatic cells surrounding oocytes (Figure 4U,E’). These results suggest the continued potential for gonadal RA production in both sexes.
Differentiated but immature gonads continued to show a clear sexually dimorphic expression pattern for cyp26a1 (Figure 4R,W,B’, G’), suggesting likely sex-specific patterns of RA degradation. In males, the broad cyp26a1 expression domain observed at 33dpf (Figure 4R) became localized to a sub-set of somatic cells surrounding cysts of germ cells by 41dpf (Figure 4B’), judging by its more restricted distribution than that of amh (Figure 4A’) and its different distribution compared to aldh1a2-expressing cells (Figure 4Z). Expression of cyp26a1 and vasa did not co-localize by double fluorescent in situ hybridization, corroborating the conclusion that cyp26a1 was expressed in somatic but not germ cells in developing male gonads (Figure 5C). In 33dpf to 41dpf females, expression of cyp26a1 continued to be restricted to the ooplasm of large oocytes that had reached diplotene (lIB green arrowhead in Figure 4W, G’ and white arrowhead in Figure 5D), while no cyp26a1 expression was detected in the ooplasm of the smallest oocytes (eIB, yellow arrowhead in Figure 4W,G’ and Figure 5D). At 41-dpf, some oocytes had reached stage II (enlarged cells due to the formation of cortical alveoli) and consequently the cyp26a1 expression signal became diluted (red arrowhead in Figure 4G’). No expression of cyp26b1 and cyp26c1 was detected in immature testes or ovaries at 31dpf and 41dpf (Figure 4T,Y,D’, I’).
4. Somatic expression of aldh1a2 and cyp26a1 is independent of germ cell signaling
Germ cell signaling is essential for sex determination in zebrafish [6–8]. To test the hypothesis that germ cell signaling controls the somatic expression of aldh1a2 and cyp26a1, we investigated the expression of both genes in knockdown zebrafish that had been depleted of germ cells by injection of antisense-morpholino (MO) against dead end (dnd), a gene essential for germ cell survival [76,77]. In situ hybridization experiments were performed on adjacent sections of gonads representing three different stages of gonad development: i) bipotential gonads at 19 dpf (Figure 6A–C); ii) transitioning gonads at 25dpf (Figure 6D–F); and iii) differentiated immature gonads at 37dpf (Figure 6G–I). The absence of expression of the germ cell specific marker vasa verified total depletion of germ cells in dnd-MO-injected animals (Figure 6C, F, I). Results revealed that aldh1a2 (Figure 6A,D,G) and cyp26a1 (Figure 6B,E,H) were both expressed in somatic cells despite the lack of germ cells at all stages examined. The aldh1a2 expression domain spanned most of the gonad, except for a few islands that did not show signal, suggesting again the existence of a small subset of somatic cells that did not express aldh1a2 as observed in male gonads (Figure 4F,P and Z). Expression of cyp26a1 also occurred in the absence of germ cells, but in a smaller subset of somatic cells compared to the broader somatic expression domain of aldh1a2.
Figure 6. Expression of aldh1a2 and cyp26a1 is independent of germ cell signaling.

Animals depleted of germ cells that develop into sterile males were generated by dead end (dnd) morpholino knockdown to study the expression of aldh1a2 and cyp26a1 in gonads at (A-C: n=1) bipotential stage (19dpf), (D–F: n=1) transitioning stage (25dpf), and (G-I: n=1) differentiated testes (37dpf). Results showed that aldh1a2 was widely expressed in somatic cells of the gonads at the three different stages analyzed (A,D,G) while cyp26a1 expression was detected in a subset of somatic cells also in the three stages analyzed (arrows in B, E, H). These results demonstrate that the onset as well as the maintenance of aldh1a2 and cyp26a1 expression, at least until 37dpf, is independent of germ cell signaling. Expression of the germ cell specific marker vasa was not detected at any stage (C,F,I), confirming the total depletion of germ cells by dnd morpholino injection in all animals. Gonads are outlined by a dashed line (B,C, E, F,H,I). Scale bar: 0.05 mm (A).
These experiments ruled out the hypothesis that the initiation or maintenance of aldh1a2 and cyp26a1 expression by somatic cells of the gonads depends on germ cell signaling in zebrafish at least until 37dpf.
5. Expression of RA-metabolic genes and somatic and germ-line markers in adult gonads
To learn if RA plays a role in gonads beyond the time of sexual fate determination and maturation, we analyzed the expression patterns of genes encoding RA-metabolic machinery in mature gonads of 6-month old adult male and female zebrafish (Figure 7A–L).
Figure 7. Expression patterns of aldh1a2, cyp26a1, cyp26b1 and cyp26c1 in adult gonads.

In adult testes (n=3), aldh1a2 expression was detected in somatic cells surrounding cysts in a localization expected for Sertoli cells (A) and cyp26a1 expression was detected in a subset of cells in a localization expected for Leydig cells (B). The presence of germ cells expressing cyp26a1 could not be discarded. Expression of cyp26b1 (C) and cyp26c1 (D) was not detected in adult testes. In adult ovaries (n=2), aldh1a2 was detected in somatic cells but not in the oocytes (E,I). Expression of cyp26a1 was restricted to the ooplasm of oocytes and was not detected in somatic cells (F,J). In oocytes, cyp26a1 expression varied according to the stage of meiosis: it was barely detectable in early stage IB oocytes (eIB, yellow arrowhead in F, prior to the diplotene stage of meiosis), was up-regulated in late stage IB oocytes that entered meiotic arrest at diplotene stage (lIB, green arrowhead in F), was maintained in stage II (red arrowhead in F) and stage III oocytes (green arrowhead in J) and was not detected in stage IV oocytes (J) coinciding with the resumption of meiosis I. Expression of cyp26b1 was detected solely in a subset of cells in the somatic tissue surrounding the oocytes (arrows in G, K) and no expression of cyp26c1 was detected in the ovary (H,L). Scale bar: 0.1mm.
In mature testes, aldh1a2 expression localized to cells surrounding cysts in a position characteristic of Sertoli cells (Figure 7A). Expression of cyp26a1 in the testis was restricted to a subset of cells, probably Leydig cells according to their localization, although expression in a small subset of germ cells (i.e. germ line stem cells) cannot be discarded (Figure 7B). Expression of cyp26b1 (Figure 7C) and cyp26c1 (Figure 7D) was not detected in mature zebrafish testes.
In mature ovaries, aldh1a2 expression was restricted to somatic cells surrounding oocytes, and no expression was detected in oocytes themselves (Figure 7E,I). Reciprocally, cyp26a1 expression was not detected in somatic cells, but appeared to be restricted to the ooplasm of oocytes (Figure 7F,J). Likewise in 41-dpf adults (Figure 4G’), ovaries expressed cyp26a1 at a barely detectable level in early stage IB oocytes (eIB, yellow arrowhead Figure 7F), but then strongly up-regulated expression in late stage IB oocytes (lIB, green arrowhead Figure 7F), which enter meiotic arrest in diplotene; oocytes then maintained this high level until stage III (green arrowhead in Figure 7J). Interestingly, expression of cyp26a1 down-regulated in oocytes at stage IV (Figure 7J), coincident with the resumption of meiosis. Note that, although the expression of cyp26b1 had not been observed in developing ovaries (Figure 4), in mature adult ovaries, a subset of somatic cells did express cyp26b1 in a punctuate pattern (Figure 7G,K), which suggests the possibility that cyp26b1 might be involved in adult ovary homeostasis. Expression of cyp26c1, however, was not detected at any time in ovaries (Figure 7H,L), suggesting that while this zebrafish paralog may play a role in other organs –for example, in the retina (Figure 3A)-, it does not act in gonads.
6. Expression analysis of genes encoding the RA-metabolic machinery during the onset of meiosis
To learn whether the sexually dimorphic expression pattern of zebrafish cyp26a1 correlates with the onset of meiosis, we compared the expression pattern of cyp26a1 to that of zebrafish orthologs of tetrapod meiosis markers (reviewed in 18). In mouse, the RA-producing duct and tubules of the mesonephros are initially connected to the anterior of the bipotential gonad, and as ovaries down-regulate Cyp26b1, the RA target gene Stra8 (Stimulated by Retinoic Acid gene 8) [78–80], which is required for the initiation of meiosis and thereby serves as a pre-meiotic marker [24,39,44,45,81], experiences an anterior–posterior wave of up-regulation accompanied by the down-regulation of the pre-meiotic pluripotent cell marker Pou5f1(Oct4) in germ cells entering meiosis and the up-regulation of the early meiotic marker Sycp3 ( Synaptonemalcomplex protein 3) [82], which is required to assemble the synaptonemal complex during meiotic prophase [83].
To our surprise, in silico screening of the zv9 version of the zebrafish genome database [15] by BLAST analysis using either mouse or human STRA8 protein sequence did not return any sequences with significant similarity (e-value<1). Likewise, a survey of four other teleost reference genomes (stickleback, Tetraodon, fugu, medaka) as well as the basally diverging spotted gar genome did not identify Stra8. We did, however, find an ortholog for Stra8 (fgenesh2_pg.scaffold_222000016, reciprocal best blast hit e-value of 8e-09 vs. human) in the genome of the basally diverging chordate amphioxus Branchiostoma floridae , and in the elephant shark [84], supporting the hypothesis that stra8 was present in stem chordates and stem gnathostomes, but was lost secondarily in the ray fin fish lineage [84] at least prior to the divergence of spotted gar before the TGD [85]. Future analysis of Stra8 in cartilaginous fish is needed to test if its ancestral function was related to the initiation of meiosis, or alternatively, if Stra8 function in meiosis was a tetrapod innovation. In any case, the absence of Stra8 in teleosts suggests that if RA plays a role in the regulation of the initiation of meiosis in teleosts, it does not act through Stra8 as it does in mammals [81].
Analysis of the early meiotic marker Sycp3 by in situ hybridization in adjacent sections showed that all zebrafish analyzed at the bipotential stage (20 dpf, n=8) showed germ cells expressing sycp3 throughout the gonad, while cells expressing cyp26a1 were mostly restricted to the dorsal surface (Figure 8A,B and Figure 4C). Transitioning and immature gonads (26 dpf and 29 dpf) expressed sycp3 and cyp26a1 in a complementary, non-overlapping fashion. In males, germ cells expressing sycp3 were located in specific regions (black arrowhead in Figure 8C, G) in which no somatic cyp26a1 expression was observed (red arrowhead in Figure 8D, H) as predicted by the hypothesis that Cyp26a1 inhibits meiosis. In females, sycp3 expression was observed only in oocytes (black arrowhead in Figure 8E, I) that had not yet reached late stage IB, and no somatic cyp261a1 expression was observed. Interestingly, females showed cyp26a1 expression in the ooplasm of late stage IB oocytes (Figure 8F,J), but not in early oocytes that expressed sycp3 (Figure 8E,I), revealing a complementary non-overlapping expression pattern of sycp3 and cyp26a1 during oocyte maturation, as would be expected if Cyp26a1 inhibited meiosis.
Figure 8. Complementary expression of the meiotic recombination marker sycp3 and cyp26a1 in developing gonads.

In bipotential gonads at 20 dpf (A,B: n=8), germ cells expressed the meiotic recombination marker sycp3 (black arrowhead in A) in a non-overlapping pattern with cyp26a1 expression, which was mostly restricted to the dorsal part of the gonad (revealing that sycp3-expressing cells did not express cyp26a1 (red arrowhead in B). Expression of the meiotic marker sycp3 was detected in bipotential gonads of all animals analyzed (A, n=8) suggesting that some germ cells entered meiosis in all juveniles regardless of their definitive sex. In differentiating testes at 26 dpf (C,D: n=2) and 29 dpf (G, H: n=2), islands of germ cells that expressed sycp3 (black arrowheads in C, G) were found in an area in which RA was likely not degraded due to lack of cyp26a1 expression (red arrowheads in D,H). In contrast, in differentiating ovaries at 26 dpf (E, F: n=2) and 29 dpf (I, J: n=2), sycp3 was expressed in small germ cells (black arrowheads in E, I) that did not express cyp26a1 (red arrowheads in F, J). The expression of cyp26a1 was restricted to the ooplasm of oocytes that reached diplotene stage and entered in meiotic arrest (F,J). Scale bar: 0.1mm.
Finally, the expression of the pluripotent marker pou5f1 and the early-meiotic marker sycp3 in bipotential gonads at 20 dpf showed non-overlapping expression domains in groups of germ cells, as shown by vasa expression (Figure 9A–C). This complementary pattern is expected if pre-meiotic germ cells down-regulate pou5f1 as they enter meiosis and begin to express sycp3 [18]. Immature testes at 24 dpf showed groups of germ cells expressing sycp3, but no cells expressing pou5f1 (Figure 9 E, F), which suggests the presence of abundant cysts of germ cells undergoing meiosis, but not obvious pre-meiotic spermatogenic cells at this stage. Immature ovaries at 24 dpf contained oocytes at early stage IB progressing through meiosis and expressing sycp3 but not pou5f1 (Figure 9G–I red arrowheads). Interestingly, oocytes at late stage IB resumed pou5f1 expression (Figure 9G–I black arrow), which is compatible with the proposed role of Pou5f1 in promoting primordial germ cell survival by preventing apoptosis [86], a mechanism that has been shown to be central in late stage IB oocytes for tipping the sexual fate of the gonad towards the female pathway in zebrafish [5,8,87].
Figure 9. Complementary expression of the meiotic recombination marker sycp3 and the pluripotent marker pou5f1(oct4) in developing gonads.

In bipotential gonads at 20 dpf, comparison of the expression of the germ cell marker vasa (A), the synaptonemal complex marker sycp3 (arrow in B) and the pluripotent gene pou5f1 (arrow in C) revealed that sycp3 and pou5f1 were both expressed in germ cells but in a complementary non-overlapping fashion (red arrowheads and dashed lines in B and C). In differentiating testes at 24 dpf, expression of vasa was detected in germ cells (D) revealing that only some of the germ cells expressed sycp3 (arrows in E) but none of them expressed pou5f1 (F). In differentiating ovaries at 24 dpf, vasa labeled germ cells (G), and sycp3 only labeled those germ cells that were small (arrows in H), which interestingly did not express pou5f1 (red arrowheads and dashed lines in I) and had not reached the late stage IB. Complementarily, the larger oocytes that had reached diplotene stage expressed pou5f1 (arrow in I). Scale bar indicated per each raw: 0.1mm.
Discussion
This work provides, to our knowledge, the first comprehensive genomic and molecular analysis of the genetic machinery that regulates the synthesis and degradation of RA at the time that zebrafish gonads tip their sexual fate towards the male or female pathway. Our findings reveal several significant differences between RA-regulated gonadogenesis in zebrafish and tetrapods, including which cells express RA-synthesizing enzymes, which paralog encodes gonadal RA-degrading enzymes, whether RA-degrading enzymes are expressed in a dimorphic fashion, whether Stra8 regulates entry into meiosis, and whether the onset of meiosis is sexually dimorphic.
1. During the critical time window for gonadal sex determination, aldh1a2, which encodes an RA-synthesizing enzyme, is expressed in the gonad in zebrafish rather than in the mesonephros as in mouse
In mouse, Aldh1a2 is not expressed in the gonad as it is in zebrafish, but is strongly expressed in the adjacent mesonephros at the bipotential stage, leading to the hypothesis that mesonephros-derived RA provides a regulatory system that drives an anterior–posterior wave of meiotic progression in the fetal ovary [23] (Figure 10A). In contrast to mouse, zebrafish gonadal somatic cells express aldh1a2 throughout gonadogenesis, suggesting that the gonad can provide a continuing source of RA that controls the development of the germline (Figure 7B). This model is consistent with the fact that in zebrafish, the mesonephros lies distant from the gonad and does not contact the gonad during the critical time window of sex determination as it does in mouse. In chicken, as in zebrafish, Aldh1a2 is expressed in the embryonic gonad [47,49], which suggests by parsimony that the expression of Aldh1a2 in the gonads is likely the ancestral condition in vertebrates and that the lack of aldh1a2 expression in the mouse gonad is probably an evolutionary innovation in the mammalian lineage (Figure 10A). Somatic cells in mouse testis begin to express Aldh1a1 shortly after the onset of Sry expression, which implies a local source of RA in the testis in addition to the Aldh1a2-derived RA from the mesonephros [46]. Male-specific Aldh1a1 expression in gonads may provide low levels of RA that might facilitate early events in testes development or may prefigure later postnatal spermatogenesis, rather than functioning in the dimorphic induction of the onset of meiosis [46]. In zebrafish, it is likely that the secondary loss of the aldh1a1 gene, a paralog of aldh1a2, which occurred during the evolution of the teleost lineage [56], was not lethal or deleterious because its function in the gonad was redundantly covered by aldh1a2. The loss of aldh1a1 in teleosts, however, might have decreased the evolvability of aldh1a2, which became the only source of RA during gonad development, given our evidence that aldh1a3 is not expressed in the zebrafish gonad.
Figure 10. Model for the role of retinoic acid and meiotic progression during gonadogenesis in mouse and zebrafish.

In mouse (A), Aldh1a2 (yellow) in the mesonephros provides the RA-source that regulates gonad development (reviewed in [18]), while in zebrafish (B), aldh1a2 is expressed by somatic cells within the gonad (yellow), and thereby provides an internal RA-source. In mouse, non-meiotic germ cells (black circles) in bipotential gonads are protected from RA by high expression of Cyp26b1 (blue), while in zebrafish, Cyp26a1 expression (blue) is restricted to cells at the dorsal surface near the body cavity, and thereby germ cells elsewhere in the gonad are not protected from RA (red) and they are able to enter into meiosis (green circles). In mouse, sexually dimorphic expression of Cyp26b1 causes low levels of RA (blue) in testes (males, top in A) and high levels of RA (red) that diffuses from the mesonephros to the ovaries in an anterior to posterior wave (females, bottom in A), which results in a sexually dimorphic onset of meiosis. The onset of meiosis in mouse follows an anterior–posterior wave, accompanied by up-regulation of Stra8, and Sycp3 and down-regulation of Pou5f1(Oct4) [18]. In zebrafish, the sexually dimorphic expression of cyp26a1 differs in time and location from those of Cyp26b1 in mouse. In zebrafish males (top in B), somatic cells up-regulate cyp26a1, and meiotic cells expressing sycp3 localize to somatic islands (red) that lack cyp26a1 expression. This model is consistent with a role for Cyp26a1 in degrading RA and thereby protecting nearby germ cells from progressing through meiosis. Consistent with this model, in females (bottom in B), cyp26a1 is not expressed in somatic cells, but it is up-regulated in the ooplasm (blue) of oocytes at late stage IB that have reached diplotene stage and have entered in meiotic arrest, and also express pou5f1(oct4). The co-expression of cyp26a1 and pou5f1(oct4) is compatible with the proposed roles of these genes in mouse on promoting germ cell survival and preventing apoptosis [41,86], mechanisms that have been shown to be central for tipping the sexual fate of the gonad toward the female pathway in zebrafish [5,8,87].
2. Independent partitioning of gonadal subfunctions between Cyp26a1 and Cyp26b1 paralogs in teleosts and tetrapods
Our study revealed that zebrafish and mouse express different Cyp26 paralogs during early gonad development; mainly Cyp26a1 in zebrafish and mainly Cyp26b1 in mouse. Our phylogenetic analysis and study of conserved syntenies between zebrafish, medaka, and mouse support the proposed gene nomenclature [63] and show that orthologies are correctly assigned.
We propose that the ancestral vertebrate Cyp26 pro-ortholog was already responsible for the regulation of RA action during gonadogenesis, and that after expansion of the Cyp26 family in stem vertebrates [54,88], independent subfunction partitioning events [72,73] led to different paralogs — cyp26a1 and Cyp26b1- functioning as the main players of RA degradation in the gonads of zebrafish and tetrapods, respectively. Comparative analysis of the three cyp26 paralogs during late eye development in zebrafish (Figure 3) and mouse [74] suggests that the independent subfunction partitioning we found in gonads does not necessarily extend to other organs. The modular fashion in which gonadal and retinal subfunctions partitioned in teleosts and tetrapods predicts that independent transcriptional regulatory elements control each subfunction.
Interestingly, a recent work detected cyp26b1 transcript by RT-PCR in 100-day post hatching gonads of Japanese flounder males induced by high temperature [89]. No data, however, is available concerning the cellular distribution of cyp26b1 expression in flounder, nor whether cyp26a1 and cyp26c1 are expressed in flounder gonads. Future analysis of all three cyp26 paralogs in the Japanese flounder and other teleosts, together with a comparative analysis of the regulatory regions of the Cyp26 paralogs of teleosts and tetrapods will clarify the evolution of cyp26 subfunctions during the evolution of teleosts and tetrapods.
3. Expression of cyp26a1 in zebrafish does not suggest a role in the dimorphic onset of meiosis as in tetrapods, but correlates with a role in the differential progression of meiotic oocytes in both males and females
In mammals, birds, and amphibians, the sexually dimorphic expression of Cyp26b1 appears to regulate the timing of meiotic onset, which differs in oocytes and spermatocytes [18,23,24,47,48]. In early mouse gonads (11.5 dpc), males and females both express Cyp26b1 throughout the gonad, but after the expression of Sry and gonads begin sexual differentiation (12.5 dpc), Cyp26b1 expression up-regulates in males but down-regulates in females [23,24] (Figure 9A). In male mice, up-regulation of Cyp26b1 in gonadal somatic cells is at a time and place appropriate to protect germ cells from RA, and thereby to prevent oogenesis in males by retarding Stra8 expression and postponing the onset of meiosis until postnatal stages [23,24] (Figure 9A). In female mice, the absence of Cyp26b1 leads to the activation of Stra8 in germ cells, and promotes the onset of meiosis and oocyte development, thereby reinforcing the female pathway (Figure 9A).
In mouse females, it has been hypothesized that the presence of germ cells committed to meiosis reinforces ovarian fate by antagonizing the testis pathway [22]. We wondered if this hypothesis also applied to zebrafish, and so tested whether some of zebrafish juveniles (presumably females) have an early onset of meiosis that could reinforce the ovarian fate of the gonad, while others (presumably males) have a delayed onset of meiosis and develop testes. Our results reveal that the meiotic marker sycp3 is expressed in bipotential gonads of all juvenile zebrafish analyzed, suggesting that, in contrast to mammals, the onset of meiosis in zebrafish gonads is not sexually dimorphic and therefore the onset of meiosis does not appear to be a cue that biases gonads towards the female or male pathway (Figure 8). Although Sycp3 is an essential component of the synaptonemal complex in meiotic cells [40] and is generally used as a meiotic marker [23,90], we cannot verify that all sycp3 expressing cells in zebrafish are progressing through meiosis [91].
Our expression analyses comparing genes that encode enzymes that regulate RA synthesis and degradation markers for somatic and germ cells and markers for meiotic and pluripotent cells are compatible with a model in which RA — or, less likely [28] a non-RA Cyp26-digested molecule [29] — is involved in the progression of meiosis in zebrafish (Figure 10). First, all juveniles with bipotential gonads containing sycp3-expressing meiotic germ cells express aldh1a2 broadly throughout the gonad but express cyp26a1 in a few peripheral cells, suggesting that most germ cells, including those randomly scattered pre-meiotic germline stem cells labeled by nanos2 [92], are developing in a cellular environment that according to gene expression patterns should contain RA (Figure 9B). Second, the result that germ cells expressing the meiotic cell marker sycp3 appear only at a substantial distance from cells expressing cyp26a1 in the same gonad suggests that the cyp26a1-expressing cells inhibit meiosis. Third, the finding that the onset and maintenance of aldh1a2 and cyp26a1 expression occurs in gonads containing no germ cells suggests that in bipotential zebrafish gonads, the gonadal soma rather than the germ line regulates RA and the onset of meiosis.
These considerations lead us to suggest the hypothesis that the continuous availability of RA to germ cells in bipotential gonads prohibits the sexually dimorphic onset of meiosis in zebrafish and is consistent with the initial development of meiotic oocytes in all juvenile zebrafish (Figure 10B). This model contrasts with mouse, in which the female-specific down-regulation of Cyp26b1 triggers the onset of meiosis, while the male-specific up-regulation of Cyp26b1 delays the onset of meiosis until after birth (Figure 10A). Our model for zebrafish is compatible with the teleost-specific loss of stra8, the “meiotic gatekeeper” [39] gene that in tetrapods mediates sexually dimorphic onset of meiosis (Figure 10B).
Interestingly, we observed that zebrafish express cyp26a1 dimorphically in gonads committed to the male or female pathway, consistent with a role of RA regulating meiotic progression during spermatogenesis and oogenesis (Figure 9B). In zebrafish males, the localized presence of sycp3-positive germ cells entering meiosis only where associated with somatic cells free of cyp26a1 expression suggests the hypothesis that cells expressing cyp26a1 degrade RA and thus prevent nearby germ cells from entering RA-induced meiosis. This hypothesis is consistent with a model in which RA availability is important for the progression of meiosis, and Cyp26a1 acts as a meiosis-inhibiting factor [23]. In females, cyp26a1 expression up-regulates in oocytes that have completed recombination (late stage IB) and are entering meiotic arrest in diplotene. This observation, together with the fact that cyp26a1 expression disappears late in oocyte development at stage IV when meiosis resumes, is compatible with the Cyp26a1-mediated degradation of RA in oocytes that prevents the progression of meiosis and maintains meiotic arrest. This finding is consistent with results in mouse, in which RA prevents meiotic arrest in testes of 13.5 dpc embryos [93]. Interestingly, in zebrafish developing gonads, oocytes entering meiotic arrest at diplotene also express both the meiosis-inhibiting gene cyp26a1 and the pluripotent marker pou5f1 (oct4). The convergence of cyp26a1 and pou5f1 expression in zebrafish oocytes at meiotic arrest is compatible with the proposed roles of Pou5f1 and Cyp26b1 in mouse germ cells in promoting survival and preventing apoptosis [41,86], mechanisms that tip the sexual fate of the gonad toward the female pathway in zebrafish [5,8,87].
Overall, this work provides the knowledge base on RA metabolic gene expression necessary for the design of experiments that alter RA signaling in zebrafish developing gonads to test the model stemming from our results for the role of RA signaling in the progress of meiosis. In addition, further work is required to understand the relationship of RA signaling to the Fgf9/Wnt4 seesaw that helps regulate gonad fate in mouse [90,94–98]. Especially intriguing with respect to the evolution of sex determination is the lack of an fgf9 gene in zebrafish and other teleosts [99,100] and the presence of two wnt4 genes [101], genomic variations with interesting consequences for zebrafish sex determination.
Materials and Methods
1. Animals and Ethics Statement
AB strain zebrafish were used in all experiments. Animals were reared and collected under standard conditions and were handled in accordance with good animal practice. The University of Oregon Institutional Animal Care and Use Committee approved all animal work (Animal Welfare Assurance Number A-3009-01, IACUC protocol 08-13).
2. dead end morpholino injections
To obtain animals lacking germ cells, wild-type zebrafish embryos were injected at the 1-2 cell stage with antisense morpholino oligonucleotide (Gene Tools, Oregon) directed against dead end as described [77]. Sibling non-injected embryos and a fraction of dnd MO-injected embryos were fixed at 24 hours post-fertilization to confirm the presence or absence of germ cells by whole-mount in situ hybridization using vasa probe as described [75].
3. In situ hybridization
In situ hybridization experiments on zebrafish cryosections were performed as described [59]. Adjacent sections of gonads were obtained by placing five consecutive 16-μm sections of the gonad on five different slides. Generally, between two to nine animals at each stage were analyzed per each probe, as indicated in each figure legend. Probes for aldh1a2 and aldh1a3 were made as described [56]; amh probe was made as described and used as a somatic cell marker to distinguish differentiating testes and ovaries when morphological features were not clear [59]; and probe for vasa was made from its 3’ end as described [75]. Fragments from cDNA of cyp26a1 (nucleotides 507-1848 of NM_131146), cyp26b1 (nucleotides 518-1548 of NM_212666), cyp26c1 (nucleotides 198-1533 of NM_001029951), pou5f1 (oct4) (nucleotides 705-1482 of NM_131112), and sycp3 (nucleotides 265-884 of NM_001040350) were cloned in TOPO vector (Invitrogen) and used to synthesize DIG-labeled riboprobes (Boehringer Mannheim). The protocol for the three-color fluorescent in situ hybridization is described in wiki.zfin.org/display/prot/3+color+Fluorescent+in+situ+on+sections.
4. Phylogenetic Tree and Analysis of synteny conservation
An alignment of Cyp26 proteins from vertebrates and cephalochordates was generated with clustalX [102]. This alignment was used to generate phylogenetic trees inferred by Neighbor-Joining using the MEGA2 package [103] and Maximum likelihood (ML) using PhyML [104] following a LG+I+G model, the alpha parameter of the gamma distribution and a proportion of invariable sites were estimated from the sample, and four categories of substitution rates were taken into account. The topology, branch lengths, and rate parameters of the tree were optimized. Tree topologies obtained by ML and NJ were identical. Confidence of tree topologies (NJ and ML) was inferred by 100 replications to calculate bootstrap values supporting each node of the tree, and differences between the two methods were minor. Cyp26 proteins from cephalochordates [54], which diverged at the base of chordate phylogeny, were used as the outgroup to root the tree. Genomic database accession numbers from NCBI (www.ncbi.nlm.nih.gov), Ensembl (www.ensembl.org) and JGI (www.jgi.doe.gov): Tetrapods: Homo sapiens (Hsa CYP26A1, NP_000774.2; CYP26B1, NP_063938.1; CYP26C1, NP_899230.2); Mus musculus (Mmu: cyp26a1, NP_031837.1; cyp26b1, NP_780684.1; cyp26c1, NP_001098671.1); Gallus gallus (Gga: Cyp26a1, NP_001001129.1; Cyp26b1, XP_426366.2; Cyp26c1, XP_421678.2); Teleosts: Danio rerio (Dre: cyp26a1, NP_571221.2; cyp26b1, NP_997831.1; cyp26c1, NP_001025122.2); Takifugu rubripes (Tru: Cyp26a1, ENSTRUG00000002005; Cyp26b1, ENSTRUG00000012613; Cyp26c1, ENSTRUG00000012794); Gasterosteus aculeatus (Gac: Cyp26a1, ENSGACG00000015370; Cyp26b1, ENSGACG00000018809; Cyp26c1, ENSGACG00000011080); Oryzias latipes (Ola: Cyp26a1, ENSORLG00000014516; Cyp26b1, ENSORLG00000003465; Cyp26c1, ENSORLG00000002036); Cephalochordates: Branchiostoma floridae (Cyp26a, Bfl Brafl1:87630; Cyp26b, Brafl1|124944 as in JGI inferred by [54]). Circleplots graphically represent user-selected chromosomes as arcs along the circumference of a circle. The origins of lines connecting positions along the arcs represent pairs of orthologous genes between two different species. Relationships of orthology and plots were generated by the Synteny Database (version Ens56; http://syntenydb.uoregon.edu/synteny_db/ [69]). Each genomic neighborhood consists of a 5Mb window centered on each Cyp26 gene: CYP26B1 in Mmu6 (82,022-87,022 kb); CYP26A1 and CYP26C1 in Mmu19 (35,260-40,272 Mb); cyp26a1 in Ola19 (19,025-24,025 Kb); cyp26b1 in Ola18 (2,121-7,121 Kb); and cyp26c1 in Ola15 (6,956-11,956 Kb). Gene loci that are close to each other may appear to overlap as a single connecting line in circle-plots due to the selected graph resolution, and lines based on best reciprocal blast hits that were not significantly different were not considered. Clusters of conserved synteny were created by coupling results from the reciprocal best hit BLAST pipeline with the use of a 100-gene sliding-window analysis that links chromosome segments with conserved synteny (for details see 69). Clusters that link chromosomal segments between different species represent orthologous syntenic conservation (e.g. Figure 2C–E), and clusters that link chromosomal segments within the same species represent paralogous syntenic conservation (e.g. Figure 2F).
Supporting Information
Cyp26 orthologies between zebrafish and medaka were supported by clusters of conserved synteny, which extends conclusions from zebrafish to other teleost models. A: cyp26a1 in Dre12 and Ola19; B: cyp26b1 in Dre7 and Ola18; and cyp26c1 in Dre17 and Ola15. Cyp26 orthologs have been labeled with larger fonts, and names of gene neighbors can be surfed in the high-resolution pdf electronic files.
(PDF)
Comparative genomic analysis of synteny conservation between mouse and medaka supports conclusions from comparison of zebrafish and mouse (Figure 2), ruling out the possibility of reciprocal gene loss in different lineages, and supports the notion that teleost cyp26a1 and tetrapod cyp26b1 are not orthologs. B: cyp26b1 in Mmu6 and Ola18; B: Cyp26a1 in Mmu19 and Ola19; and Cyp26c1 in Mmu19 and Ola15. Large fonts label cyp26 orthologs and names of gene neighbors are legible in the high-resolution pdf electronic files.
(PDF)
Acknowledgments
We are grateful to A. Rapp, M. McFadden, R. Montgomery, T. Mason and the University of Oregon Zebrafish Facility for providing animals and excellent fish care. We thank C.M. Rivera and M. Fajer for helping with sample collection. We thank P.K. Loi, N. Banning and B. Wiskow from the University of Oregon Histology Facility for sample sectioning.
Funding Statement
Funding support to the University of Oregon Zebrafish Facility came from grant 5P01HD022486, to CC came from grant 2009SGR336 from Generalitat de Catalunya and grant BFU2010-14875 from the Ministerio de Economía y Competitividad (Spain), and to JHP came from grants 1R01GM085318 and 2R01RR020833 (alias R01OD011116) from the National Institutes of Health. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Brennan J, Capel B (2004) One tissue, two fates: molecular genetic events that underlie testis versus ovary development. Nat Rev Genet 5: 509-521. doi:10.1038/nrm1449. PubMed: 15211353. [DOI] [PubMed] [Google Scholar]
- 2. Koopman P (2001) Sry, Sox9 and mammalian sex determination. EXS: 25-56. PubMed: 11301599. [DOI] [PubMed] [Google Scholar]
- 3. Lovell-Badge R, Robertson E (1990) XY female mice resulting from a heritable mutation in the primary testis-determining gene, Tdy. Development 109: 635-646. PubMed: 2401216. [DOI] [PubMed] [Google Scholar]
- 4. Takahashi H (1977) Juvenile hermaphroditism in the zebrafish, Brachydanio rerio. Bull Fac Fish Hokkaido Univ 28: 57–65. [Google Scholar]
- 5. Uchida D, Yamashita M, Kitano T, Iguchi T (2002) Oocyte apoptosis during the transition from ovary-like tissue to testes during sex differentiation of juvenile zebrafish. J Exp Biol 205: 711-718. PubMed: 11914381. [DOI] [PubMed] [Google Scholar]
- 6. Slanchev K, Stebler J, de la Cueva-Méndez G, Raz E (2005) Development without germ cells: the role of the germ line in zebrafish sex differentiation. Proc Natl Acad Sci U S A 102: 4074-4079. doi:10.1073/pnas.0407475102. PubMed: 15728735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Siegfried KR, Nüsslein-Volhard C (2008) Germ line control of female sex determination in zebrafish. Dev Biol 324: 277-287. doi:10.1016/j.ydbio.2008.09.025. PubMed: 18930041. [DOI] [PubMed] [Google Scholar]
- 8. Rodríguez-Marí A, Cañestro C, Bremiller RA, Nguyen-Johnson A, Asakawa K et al. (2010) Sex reversal in zebrafish fancl mutants is caused by Tp53-mediated germ cell apoptosis. PLOS Genet 6: e1001034 PubMed: 20661450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Rodríguez-Marí A, Postlethwait JH (2011) The role of Fanconi anemia/BRCA genes in zebrafish sex determination. Methods Cell Biol 105: 461-490. doi:10.1016/B978-0-12-381320-6.00020-5. PubMed: 21951543. [DOI] [PubMed] [Google Scholar]
- 10. Herpin A, Schartl M (2011) Sex determination: switch and suppress. Curr Biol 21: R656-R659. doi:10.1016/j.cub.2011.07.026. PubMed: 21920296. [DOI] [PubMed] [Google Scholar]
- 11. Graves JA (2013) How to evolve new vertebrate sex determining genes. Dev Dyn 242: 354-359. doi:10.1002/dvdy.23887. PubMed: 23074164. [DOI] [PubMed] [Google Scholar]
- 12. Anderson JL, Rodríguez-Marí A, Braasch I, Amores A, Hohenlohe P et al. (2012) Multiple sex-associated regions and a putative sex chromosome in zebrafish revealed by RAD mapping and population genomics. PLOS ONE 7: e40701. doi:10.1371/journal.pone.0040701. PubMed: 22792396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Bradley KM, Breyer JP, Melville DB, Broman KW, Knapik EW et al. (2011) An SNP-Based Linkage Map for Zebrafish Reveals Sex Determination Loci Bethesda)1. p. G3 pp. 3-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Liew WC, Bartfai R, Lim Z, Sreenivasan R, Siegfried KR et al. (2012) Polygenic sex determination system in zebrafish. PLOS ONE 7: e34397. doi:10.1371/journal.pone.0034397. PubMed: 22506019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Howe K, Clark MD, Torroja CF, Torrance J, Berthelot C et al. (2013) The zebrafish reference genome sequence and its relationship to the human genome. Nature, 496: 498–503. PubMed: 23594743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hunt PA, Hassold TJ (2002) Sex matters in meiosis. Science 296: 2181-2183. doi:10.1126/science.1071907. PubMed: 12077403. [DOI] [PubMed] [Google Scholar]
- 17. Morelli MA, Cohen PE (2005) Not all germ cells are created equal: aspects of sexual dimorphism in mammalian meiosis. Reproduction 130: 761-781. doi:10.1530/rep.1.00865. PubMed: 16322537. [DOI] [PubMed] [Google Scholar]
- 18. Bowles J, Koopman P (2007) Retinoic acid, meiosis and germ cell fate in mammals. Development 134: 3401-3411. doi:10.1242/dev.001107. PubMed: 17715177. [DOI] [PubMed] [Google Scholar]
- 19. Lesch BJ, Page DC (2012) Genetics of germ cell development. Nat Rev Genet 13: 781-794. doi:10.1038/nrg3294. PubMed: 23044825. [DOI] [PubMed] [Google Scholar]
- 20. Hilscher B, Hilscher W, Bülthoff-Ohnolz B, Krämer U, Birke A et al. (1974) Kinetics of gametogenesis. I. Comparative histological and autoradiographic studies of oocytes and transitional prospermatogonia during oogenesis and prespermatogenesis. Cell Tissue Res 154: 443-470. PubMed: 4442109. [DOI] [PubMed] [Google Scholar]
- 21. McLaren A (1984) Meiosis and differentiation of mouse germ cells. Symp Soc Exp Biol 38: 7-23. PubMed: 6400220. [PubMed] [Google Scholar]
- 22. Yao HH, DiNapoli L, Capel B (2003) Meiotic germ cells antagonize mesonephric cell migration and testis cord formation in mouse gonads. Development 130: 5895-5902. doi:10.1242/dev.00836. PubMed: 14561636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Bowles J, Knight D, Smith C, Wilhelm D, Richman J et al. (2006) Retinoid signaling determines germ cell fate in mice. Science 312: 596-600. doi:10.1126/science.1125691. PubMed: 16574820. [DOI] [PubMed] [Google Scholar]
- 24. Koubova J, Menke DB, Zhou Q, Capel B, Griswold MD et al. (2006) Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc Natl Acad Sci U S A 103: 2474-2479. doi:10.1073/pnas.0510813103. PubMed: 16461896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Lin Y, Gill ME, Koubova J, Page DC (2008) Germ cell-intrinsic and -extrinsic factors govern meiotic initiation in mouse embryos. Science 322: 1685-1687. doi:10.1126/science.1166340. PubMed: 19074348. [DOI] [PubMed] [Google Scholar]
- 26. Le Bouffant R, Guerquin MJ, Duquenne C, Frydman N, Coffigny H et al. (2010) Meiosis initiation in the human ovary requires intrinsic retinoic acid synthesis. Hum Reprod 25: 2579-2590. doi:10.1093/humrep/deq195. PubMed: 20670969. [DOI] [PubMed] [Google Scholar]
- 27. Li H, Clagett-Dame M (2009) Vitamin A deficiency blocks the initiation of meiosis of germ cells in the developing rat ovary in vivo. Biol Reprod 81: 996-1001. doi:10.1095/biolreprod.109.078808. PubMed: 19587328. [DOI] [PubMed] [Google Scholar]
- 28. Griswold MD, Hogarth CA, Bowles J, Koopman P (2012) Initiating meiosis: the case for retinoic acid. Biol Reprod 86: 35. doi:10.1095/biolreprod.111.096610. PubMed: 22075477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Kumar S, Chatzi C, Brade T, Cunningham TJ, Zhao X et al. (2011) Sex-specific timing of meiotic initiation is regulated by Cyp26b1 independent of retinoic acid signalling. Nat Commun 2: 151. doi:10.1038/ncomms1136. PubMed: 21224842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Chambon P (1996) A decade of molecular biology of retinoic acid receptors. FASEB J 10: 940-954. PubMed: 8801176. [PubMed] [Google Scholar]
- 31. Mark M, Ghyselinck NB, Chambon P (2006) Function of retinoid nuclear receptors: lessons from genetic and pharmacological dissections of the retinoic acid signaling pathway during mouse embryogenesis. Annu Rev Pharmacol Toxicol 46: 451-480. doi:10.1146/annurev.pharmtox.46.120604.141156. PubMed: 16402912. [DOI] [PubMed] [Google Scholar]
- 32. Duester G (2008) Retinoic acid synthesis and signaling during early organogenesis. Cell 134: 921-931. doi:10.1016/j.cell.2008.09.002. PubMed: 18805086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Niederreither K, Dollé P (2008) Retinoic acid in development: towards an integrated view. Nat Rev Genet 9: 541-553. doi:10.1038/nrg2340. PubMed: 18542081. [DOI] [PubMed] [Google Scholar]
- 34. Reijntjes S, Blentic A, Gale E, Maden M (2005) The control of morphogen signalling: regulation of the synthesis and catabolism of retinoic acid in the developing embryo. Dev Biol 285: 224-237. doi:10.1016/j.ydbio.2005.06.019. PubMed: 16054125. [DOI] [PubMed] [Google Scholar]
- 35. Snyder EM, Small C, Griswold MD (2010) Retinoic acid availability drives the asynchronous initiation of spermatogonial differentiation in the mouse. Biol Reprod 83: 783-790. doi:10.1095/biolreprod.110.085811. PubMed: 20650878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Bouillet P, Oulad-Abdelghani M, Vicaire S, Garnier JM, Schuhbaur B et al. (1995) Efficient cloning of cDNAs of retinoic acid-responsive genes in P19 embryonal carcinoma cells and characterization of a novel mouse gene, Stra1 (mouse LERK-2/Eplg2). Dev Biol 170: 420-433. doi:10.1006/dbio.1995.1226. PubMed: 7649373. [DOI] [PubMed] [Google Scholar]
- 37. Oulad-Abdelghani M, Bouillet P, Décimo D, Gansmuller A, Heyberger S et al. (1996) Characterization of a premeiotic germ cell-specific cytoplasmic protein encoded by Stra8, a novel retinoic acid-responsive gene. J Cell Biol 135: 469-477. doi:10.1083/jcb.135.2.469. PubMed: 8896602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Rosner MH, Vigano MA, Ozato K, Timmons PM, Poirier F et al. (1990) A POU-domain transcription factor in early stem cells and germ cells of the mammalian embryo. Nature 345: 686-692. doi:10.1038/345686a0. PubMed: 1972777. [DOI] [PubMed] [Google Scholar]
- 39. Baltus AE, Menke DB, Hu YC, Goodheart ML, Carpenter AE et al. (2006) In germ cells of mouse embryonic ovaries, the decision to enter meiosis precedes premeiotic DNA replication. Nat Genet 38: 1430-1434. doi:10.1038/ng1919. PubMed: 17115059. [DOI] [PubMed] [Google Scholar]
- 40. Di Carlo AD, Travia G, De Felici M (2000) The meiotic specific synaptonemal complex protein SCP3 is expressed by female and male primordial germ cells of the mouse embryo. Int J Dev Biol 44: 241-244. PubMed: 10794082. [PubMed] [Google Scholar]
- 41. MacLean G, Li H, Metzger D, Chambon P, Petkovich M (2007) Apoptotic extinction of germ cells in testes of Cyp26b1 knockout mice. Endocrinology 148: 4560-4567. doi:10.1210/en.2007-0492. PubMed: 17584971. [DOI] [PubMed] [Google Scholar]
- 42. Li H, MacLean G, Cameron D, Clagett-Dame M, Petkovich M (2009) Cyp26b1 expression in murine Sertoli cells is required to maintain male germ cells in an undifferentiated state during embryogenesis. PLOS ONE 4: e7501. doi:10.1371/journal.pone.0007501. PubMed: 19838304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Li H, Palczewski K, Baehr W, Clagett-Dame M (2011) Vitamin A deficiency results in meiotic failure and accumulation of undifferentiated spermatogonia in prepubertal mouse testis. Biol Reprod 84: 336-341. doi:10.1095/biolreprod.110.086157. PubMed: 20881313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Anderson EL, Baltus AE, Roepers-Gajadien HL, Hassold TJ, de Rooij DG et al. (2008) Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. Proc Natl Acad Sci U S A 105: 14976-14980. doi:10.1073/pnas.0807297105. PubMed: 18799751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Zhou Q, Li Y, Nie R, Friel P, Mitchell D et al. (2008) Expression of stimulated by retinoic acid gene 8 (Stra8) and maturation of murine gonocytes and spermatogonia induced by retinoic acid in vitro. Biol Reprod 78: 537-545. doi:10.1095/biolreprod.107.064337. PubMed: 18032419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Bowles J, Feng CW, Knight D, Smith CA, Roeszler KN et al. (2009) Male-specific expression of Aldh1a1 in mouse and chicken fetal testes: implications for retinoid balance in gonad development. Dev Dyn 238: 2073-2080. doi:10.1002/dvdy.22024. PubMed: 19618468. [DOI] [PubMed] [Google Scholar]
- 47. Smith CA, Roeszler KN, Bowles J, Koopman P, Sinclair AH (2008) Onset of meiosis in the chicken embryo; evidence of a role for retinoic acid. BMC Dev Biol 8: 85. doi:10.1186/1471-213X-8-85. PubMed: 18799012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wallacides A, Chesnel A, Chardard D, Flament S, Dumond H (2009) Evidence for a conserved role of retinoic acid in urodele amphibian meiosis onset. Dev Dyn 238: 1389-1398. doi:10.1002/dvdy.21948. PubMed: 19347951. [DOI] [PubMed] [Google Scholar]
- 49. Yu M, Yu P, Leghari IH, Ge C, Mi Y et al. (2013) RALDH2, the enzyme for retinoic acid synthesis, mediates meiosis initiation in germ cells of the female embryonic chickens. Amino Acids 44: 405-412. doi:10.1007/s00726-012-1343-6. PubMed: 22733143. [DOI] [PubMed] [Google Scholar]
- 50. Alsop D, Matsumoto J, Brown S, Van Der Kraak G (2008) Retinoid requirements in the reproduction of zebrafish. Gen Comp Endocrinol 156: 51-62. doi:10.1016/j.ygcen.2007.11.008. PubMed: 18158153. [DOI] [PubMed] [Google Scholar]
- 51. Uchida D, Yamashita M, Kitano T, Iguchi T (2004) An aromatase inhibitor or high water temperature induce oocyte apoptosis and depletion of P450 aromatase activity in the gonads of genetic female zebrafish during sex-reversal. Comp Biochem Physiol A Mol Integr Physiol 137: 11-20. doi:10.1016/S1095-6433(03)00178-8. PubMed: 14720586. [DOI] [PubMed] [Google Scholar]
- 52. Dranow DB, Tucker RP, Draper BW (2013) Germ cells are required to maintain a stable sexual phenotype in adult zebrafish. Dev Biol 376: 43-50. doi:10.1016/j.ydbio.2013.01.016. PubMed: 23348677. [DOI] [PubMed] [Google Scholar]
- 53. Rodríguez-Marí A, Wilson C, Titus TA, Cañestro C, BreMiller RA et al. (2011) Roles of brca2 (fancd1) in oocyte nuclear architecture, gametogenesis, gonad tumors, and genome stability in zebrafish. PLOS Genet 7: e1001357 PubMed: 21483806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Cañestro C, Postlethwait JH, Gonzàlez-Duarte R, Albalat R (2006) Is retinoic acid genetic machinery a chordate innovation? Evol Dev 8: 394-406. doi:10.1111/j.1525-142X.2006.00113.x. PubMed: 16925675. [DOI] [PubMed] [Google Scholar]
- 55. Pittlik S, Domingues S, Meyer A, Begemann G (2008) Expression of zebrafish aldh1a3 (raldh3) and absence of aldh1a1 in teleosts. Gene Expr Patterns 8: 141-147. doi:10.1016/j.gep.2007.11.003. PubMed: 18178530. [DOI] [PubMed] [Google Scholar]
- 56. Cañestro C, Catchen JM, Rodríguez-Marí A, Yokoi H, Postlethwait JH (2009) Consequences of lineage-specific gene loss on functional evolution of surviving paralogs: ALDH1A and retinoic acid signaling in vertebrate genomes. PLOS Genet 5: e1000496 PubMed: 19478994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Albalat R, Cañestro C (2009) Identification of Aldh1a, Cyp26 and RAR orthologs in protostomes pushes back the retinoic acid genetic machinery in evolutionary time to the bilaterian ancestor. Chem Biol Interact 178: 188-196. doi:10.1016/j.cbi.2008.09.017. PubMed: 18926806. [DOI] [PubMed] [Google Scholar]
- 58. Maack G, Segner H (2003) Morphological development of the gonads in zebrafish. J Fish Biol 62: 895-906. doi:10.1046/j.1095-8649.2003.00074.x. [Google Scholar]
- 59. Rodríguez-Marí A, Yan YL, Bremiller RA, Wilson C, Cañestro C et al. (2005) Characterization and expression pattern of zebrafish Anti-Mullerian hormone (Amh) relative to sox9a, sox9b, and cyp19a1a, during gonad development. Gene Expr Patterns 5: 655-667. doi:10.1016/j.modgep.2005.02.008. PubMed: 15939378. [DOI] [PubMed] [Google Scholar]
- 60. Wang XG, Bartfai R, Sleptsova-Freidrich I, Orban L (2007) The timing and extent of ‘juvenile ovary’ phase are highly variable during zebrafish testis differentiation. J Fish Biol 70: 33–44. doi:10.1111/j.1095-8649.2007.01363.x. [Google Scholar]
- 61. White JA, Guo YD, Baetz K, Beckett-Jones B, Bonasoro J et al. (1996) Identification of the retinoic acid-inducible all-trans-retinoic acid 4-hydroxylase. J Biol Chem 271: 29922-29927. doi:10.1074/jbc.271.47.29922. PubMed: 8939936. [DOI] [PubMed] [Google Scholar]
- 62. Zhao Q, Dobbs-McAuliffe B, Linney E (2005) Expression of cyp26b1 during zebrafish early development. Gene Expr Patterns 5: 363-369. doi:10.1016/j.modgep.2004.09.011. PubMed: 15661642. [DOI] [PubMed] [Google Scholar]
- 63. Hernandez RE, Putzke AP, Myers JP, Margaretha L, Moens CB (2007) Cyp26 enzymes generate the retinoic acid response pattern necessary for hindbrain development. Development 134: 177-187. doi:10.1242/dev.02706. PubMed: 17164423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Gu X, Xu F, Wang X, Gao X, Zhao Q (2005) Molecular cloning and expression of a novel CYP26 gene (cyp26d1) during zebrafish early development. Gene Expr Patterns 5: 733-739. doi:10.1016/j.modgep.2005.04.005. PubMed: 15979416. [DOI] [PubMed] [Google Scholar]
- 65. Ray WJ, Bain G, Yao M, Gottlieb DI (1997) CYP26, a novel mammalian cytochrome P450, is induced by retinoic acid and defines a newfamily. J Biol Chem 272: 18702-18708. doi:10.1074/jbc.272.30.18702. PubMed: 9228041. [DOI] [PubMed] [Google Scholar]
- 66. Abu-Abed SS, Beckett BR, Chiba H, Chithalen JV, Jones G et al. (1998) Mouse P450RAI (CYP26) expression and retinoic acid-inducible retinoic acid metabolism in F9 cells are regulated by retinoic acid receptor gamma and retinoid X receptor alpha. J Biol Chem 273: 2409-2415. doi:10.1074/jbc.273.4.2409. PubMed: 9442090. [DOI] [PubMed] [Google Scholar]
- 67. MacLean G, Abu-Abed S, Dollé P, Tahayato A, Chambon P et al. (2001) Cloning of a novel retinoic-acid metabolizing cytochrome P450, Cyp26B1, and comparative expression analysis with Cyp26A1 during early murine development. Mech Dev 107: 195-201. doi:10.1016/S0925-4773(01)00463-4. PubMed: 11520679. [DOI] [PubMed] [Google Scholar]
- 68. Postlethwait JH (2007) The zebrafish genome in context: ohnologs gone missing. J Exp Zool B Mol Dev Evol 308: 563-577. PubMed: 17068775. [DOI] [PubMed] [Google Scholar]
- 69. Catchen JM, Conery JS, Postlethwait JH (2009) Automated identification of conserved synteny after whole-genome duplication. Genome Res 19: 1497-1505. doi:10.1101/gr.090480.108. PubMed: 19465509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Bassham S, Cañestro C, Postlethwait JH (2008) Evolution of developmental roles of Pax2/5/8 paralogs after independent duplication in urochordate and vertebrate lineages. BMC Biol 6: 35. doi:10.1186/1741-7007-6-35. PubMed: 18721460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Dehal P, Boore JL (2005) Two rounds of whole genome duplication in the ancestral vertebrate. PLOS Biol 3: e314. doi:10.1371/journal.pbio.0030314. PubMed: 16128622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Postlethwait J, Amores A, Cresko W, Singer A, Yan YL (2004) Subfunction partitioning, the teleost radiation and the annotation of the human genome. Trends Genet 20: 481-490. doi:10.1016/j.tig.2004.08.001. PubMed: 15363902. [DOI] [PubMed] [Google Scholar]
- 73. Force A, Lynch M, Pickett FB, Amores A, Yan YL et al. (1999) Preservation of duplicate genes by complementary, degenerative mutations. Genetics 151: 1531-1545. PubMed: 10101175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Sakai Y, Luo T, McCaffery P, Hamada H, Dräger UC (2004) CYP26A1 and CYP26C1 cooperate in degrading retinoic acid within the equatorial retina during later eye development. Dev Biol 276: 143-157. doi:10.1016/j.ydbio.2004.08.032. PubMed: 15531370. [DOI] [PubMed] [Google Scholar]
- 75. Yoon C, Kawakami K, Hopkins N (1997) Zebrafish vasa homologue RNA is localized to the cleavage planes of 2- and 4-cell-stage embryos and is expressed in the primordial germ cells. Development 124: 3157-3165. PubMed: 9272956. [DOI] [PubMed] [Google Scholar]
- 76. Ciruna B, Weidinger G, Knaut H, Thisse B, Thisse C et al. (2002) Production of maternal-zygotic mutant zebrafish by germ-line replacement. Proc Natl Acad Sci U S A 99: 14919-14924. doi:10.1073/pnas.222459999. PubMed: 12397179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Weidinger G, Stebler J, Slanchev K, Dumstrei K, Wise C et al. (2003) dead end, a novel vertebrate germ plasm component, is required for zebrafish primordial germ cell migration and survival. Curr Biol 13: 1429-1434. doi:10.1016/S0960-9822(03)00537-2. PubMed: 12932328. [DOI] [PubMed] [Google Scholar]
- 78. Mu X, Wen J, Guo M, Wang J, Li G et al. (2013) Retinoic acid derived from the fetal ovary initiates meiosis in mouse germ cells. J Cell Physiol 228: 627-639. doi:10.1002/jcp.24172. PubMed: 22886539. [DOI] [PubMed] [Google Scholar]
- 79. Tedesco M, Desimio MG, Klinger FG, De Felici M, Farini D (2013) Minimal concentrations of retinoic Acid induce stimulation by retinoic Acid 8 and promote entry into meiosis in isolated pregonadal and gonadal mouse primordial germ cells. Biol Reprod 88: 145. doi:10.1095/biolreprod.112.106526. PubMed: 23636811. [DOI] [PubMed] [Google Scholar]
- 80. Menke DB, Koubova J, Page DC (2003) Sexual differentiation of germ cells in XX mouse gonads occurs in an anterior-to-posterior wave. Dev Biol 262: 303-312. doi:10.1016/S0012-1606(03)00391-9. PubMed: 14550793. [DOI] [PubMed] [Google Scholar]
- 81. Mark M, Jacobs H, Oulad-Abdelghani M, Dennefeld C, Féret B et al. (2008) STRA8-deficient spermatocytes initiate, but fail to complete, meiosis and undergo premature chromosome condensation. J Cell Sci 121: 3233-3242. doi:10.1242/jcs.035071. PubMed: 18799790. [DOI] [PubMed] [Google Scholar]
- 82. Bullejos M, Koopman P (2004) Germ cells enter meiosis in a rostro-caudal wave during development of the mouse ovary. Mol Reprod Dev 68: 422-428. doi:10.1002/mrd.20105. PubMed: 15236325. [DOI] [PubMed] [Google Scholar]
- 83. Yuan L, Liu JG, Zhao J, Brundell E, Daneholt B et al. (2000) The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol Cell 5: 73-83. doi:10.1016/S1097-2765(00)80404-9. PubMed: 10678170. [DOI] [PubMed] [Google Scholar]
- 84. Venkatesh B, Kirkness EF, Loh YH, Halpern AL, Lee AP et al. (2007) Survey sequencing and comparative analysis of the elephant shark (Callorhinchus milii) genome. PLOS Biol 5: e101. doi:10.1371/journal.pbio.0050101. PubMed: 17407382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Amores A, Catchen J, Ferrara A, Fontenot Q, Postlethwait JH (2011) Genome evolution and meiotic maps by massively parallel DNA sequencing: spotted gar, an outgroup for the teleost genome duplication. Genetics 188: 799-808. doi:10.1534/genetics.111.127324. PubMed: 21828280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Kehler J, Tolkunova E, Koschorz B, Pesce M, Gentile L et al. (2004) Oct4 is required for primordial germ cell survival. EMBO Rep 5: 1078-1083. doi:10.1038/sj.embor.7400279. PubMed: 15486564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Houwing S, Kamminga LM, Berezikov E, Cronembold D, Girard A et al. (2007) A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish. Cell 129: 69-82. doi:10.1016/j.cell.2007.03.026. PubMed: 17418787. [DOI] [PubMed] [Google Scholar]
- 88. Marlétaz F, Holland LZ, Laudet V, Schubert M (2006) Retinoic acid signaling and the evolution of chordates. Int J Biol Sci 2: 38-47. PubMed: 16733532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Yamaguchi T, Kitano T (2012) High temperature induces cyp26b1 mRNA expression and delays meiotic initiation of germ cells by increasing cortisol levels during gonadal sex differentiation in Japanese flounder. Biochem Biophys Res Commun 419: 287-292. doi:10.1016/j.bbrc.2012.02.012. PubMed: 22342721. [DOI] [PubMed] [Google Scholar]
- 90. Bowles J, Feng CW, Spiller C, Davidson TL, Jackson A et al. (2010) FGF9 suppresses meiosis and promotes male germ cell fate in mice. Dev Cell 19: 440-449. doi:10.1016/j.devcel.2010.08.010. PubMed: 20833365. [DOI] [PubMed] [Google Scholar]
- 91. Novak I, Lightfoot DA, Wang H, Eriksson A, Mahdy E et al. (2006) Mouse embryonic stem cells form follicle-like ovarian structures but do not progress through meiosis. Stem Cells 24: 1931-1936. doi:10.1634/stemcells.2005-0520. PubMed: 16644921. [DOI] [PubMed] [Google Scholar]
- 92. Beer RL, Draper BW (2013) nanos3 maintains germline stem cells and expression of the conserved germline stem cell gene nanos2 in the zebrafish ovary. Dev Biol 374: 308-318. doi:10.1016/j.ydbio.2012.12.003. PubMed: 23228893. [DOI] [PubMed] [Google Scholar]
- 93. Trautmann E, Guerquin MJ, Duquenne C, Lahaye JB, Habert R et al. (2008) Retinoic acid prevents germ cell mitotic arrest in mouse fetal testes. Cell Cycle 7: 656-664. doi:10.4161/cc.7.5.5482. PubMed: 18256537. [DOI] [PubMed] [Google Scholar]
- 94. Colvin JS, Green RP, Schmahl J, Capel B, Ornitz DM (2001) Male-to-female sex reversal in mice lacking fibroblast growth factor 9. Cell 104: 875-889. doi:10.1016/S0092-8674(01)00284-7. PubMed: 11290325. [DOI] [PubMed] [Google Scholar]
- 95. Naillat F, Prunskaite-Hyyryläinen R, Pietilä I, Sormunen R, Jokela T et al. (2010) Wnt4/5a signalling coordinates cell adhesion and entry into meiosis during presumptive ovarian follicle development. Hum Mol Genet 19: 1539-1550. doi:10.1093/hmg/ddq027. PubMed: 20106871. [DOI] [PubMed] [Google Scholar]
- 96. Chassot AA, Gregoire EP, Lavery R, Taketo MM, de Rooij DG et al. (2011) RSPO1/beta-catenin signaling pathway regulates oogonia differentiation and entry into meiosis in the mouse fetal ovary. PLOS ONE 6: e25641. doi:10.1371/journal.pone.0025641. PubMed: 21991325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Jameson SA, Lin YT, Capel B (2012) Testis development requires the repression of Wnt4 by Fgf signaling. Dev Biol 370: 24-32. doi:10.1016/j.ydbio.2012.06.009. PubMed: 22705479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Kim Y, Kobayashi A, Sekido R, DiNapoli L, Brennan J et al. (2006) Fgf9 and Wnt4 act as antagonistic signals to regulate mammalian sex determination. PLOS Biol 4: e187. doi:10.1371/journal.pbio.0040187. PubMed: 16700629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Itoh N (2007) The Fgf families in humans, mice, and zebrafish: their evolutional processes and roles in development, metabolism, and disease. Biol Pharm Bull 30: 1819-1825. doi:10.1248/bpb.30.1819. PubMed: 17917244. [DOI] [PubMed] [Google Scholar]
- 100. Sun YL, Zeng S, Ye K, Yang C, Li MH et al. (2012) Involvement of FGF9/16/20 subfamily in female germ cell development of the Nile tilapia, Oreochromis niloticus. Fish Physiol Biochem 38: 1427-1439. doi:10.1007/s10695-012-9630-4. PubMed: 22451340. [DOI] [PubMed] [Google Scholar]
- 101. Nicol B, Guerin A, Fostier A, Guiguen Y (2012) Ovary-predominant wnt4 expression during gonadal differentiation is not conserved in the rainbow trout (Oncorhynchus mykiss). Mol Reprod Dev 79: 51-63. doi:10.1002/mrd.21404. PubMed: 22125114. [DOI] [PubMed] [Google Scholar]
- 102. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25: 4876-4882. doi:10.1093/nar/25.24.4876. PubMed: 9396791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Kumar S, Tamura K, Jakobsen IB, Nei M (2001) MEGA2: molecular evolutionary genetics analysis software. Bioinformatics 17: 1244-1245. doi:10.1093/bioinformatics/17.12.1244. PubMed: 11751241. [DOI] [PubMed] [Google Scholar]
- 104. Guindon S, Lethiec F, Duroux P, Gascuel O (2005) PHYML Online--a web server for fast maximum likelihood-based phylogenetic inference. Nucleic Acids Res 33: W557-W559. doi:10.1093/nar/gki352. PubMed: 15980534. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Cyp26 orthologies between zebrafish and medaka were supported by clusters of conserved synteny, which extends conclusions from zebrafish to other teleost models. A: cyp26a1 in Dre12 and Ola19; B: cyp26b1 in Dre7 and Ola18; and cyp26c1 in Dre17 and Ola15. Cyp26 orthologs have been labeled with larger fonts, and names of gene neighbors can be surfed in the high-resolution pdf electronic files.
(PDF)
Comparative genomic analysis of synteny conservation between mouse and medaka supports conclusions from comparison of zebrafish and mouse (Figure 2), ruling out the possibility of reciprocal gene loss in different lineages, and supports the notion that teleost cyp26a1 and tetrapod cyp26b1 are not orthologs. B: cyp26b1 in Mmu6 and Ola18; B: Cyp26a1 in Mmu19 and Ola19; and Cyp26c1 in Mmu19 and Ola15. Large fonts label cyp26 orthologs and names of gene neighbors are legible in the high-resolution pdf electronic files.
(PDF)