

Abstract
Progranulin (PGRN) is a soluble factor that regulates cell proliferation, motility and inflammation. A role for PGRN in the progression of ovarian and breast cancers is well established. However, the expression and subsequent consequences of PGRN on the progression of gastrointestinal tumors is not well recognized. This review briefly summarizes our current knowledge of the mechanisms of action of PGRN and highlights the role of this signaling molecule in various gastrointestinal cancers.
Keywords: Cholangiocarcinoma, hepatocellular carcinoma, colorectal cancer, esophageal cancer, growth factor
Background
Progranulin (PGRN), also known as granulin-epithelin precursor, proepithelin, acrogranin and GP88/PC-cell derived growth factor, is a secreted glycoprotein conserved in most eukaryotes (1). It has been shown to mediate cell cycle progression, cell motility (2) and inflammatory processes (3). Structurally, it belongs to none of the well-established growth factor families, and inhibition of the known growth factor receptors such as IGF and EGF receptors fails to prevent the actions of PGRN (2).
PGRN has been implicated in a number of disease states. For example, a loss- of-function mutation in PGRN has been associated with the onset of frontotemporal lobar degeneration (4). Conversely, PGRN over-activity occurs in many types of cancer (2,5–8) and is thought to confer a more aggressive phenotype to the tumor cells (6). However, how the dysregulation of PGRN function leads to these pathogenic states is largely unknown.
Processing of PGRN to granulins
The human PGRN gene contains 12 protein-coding exons that result in 3 isoforms (9,10) and contains a signal sequence and a number of granulin-like domains. These domains are composed of highly conserved tandem repeats of a unique 12-cysteine sequence (11). The full-length PGRN protein is approximately 68.5 kDa, though it is heavily glycosylated and will therefore migrate as though it is much heavier (1). After secretion, the full-length protein can be proteolytically cleaved between the granulin-like domains by metalloproteinases such as matrix metalloproteinase-9 (MMP-9) (12), MMP-14 (13) and ADAMTS-7 (14). Complete cleavage of full length PGRN results in granulin peptides (GRN A–G and paragranulin) that are approximately 6 kDa in size, though intermediary cleavage products have also been identified (1). The full-length PGRN protein, the intermediaries and the GRN peptides all have biological activity, although it remains to be clarified if these are distinct or overlapping. Regardless, the proteolytic processing of full-length PGRN can be inhibited when secretory leukocyte protease inhibitor (SLPI) or high-density lipoprotein/apolipoprotein A-1 (HDL/ApoA-1) binds to full-length PGRN, preventing its cleavage (15). A schematic representation of the proteolytic cleavage of PGRN is described in Figure 1.
Figure 1.
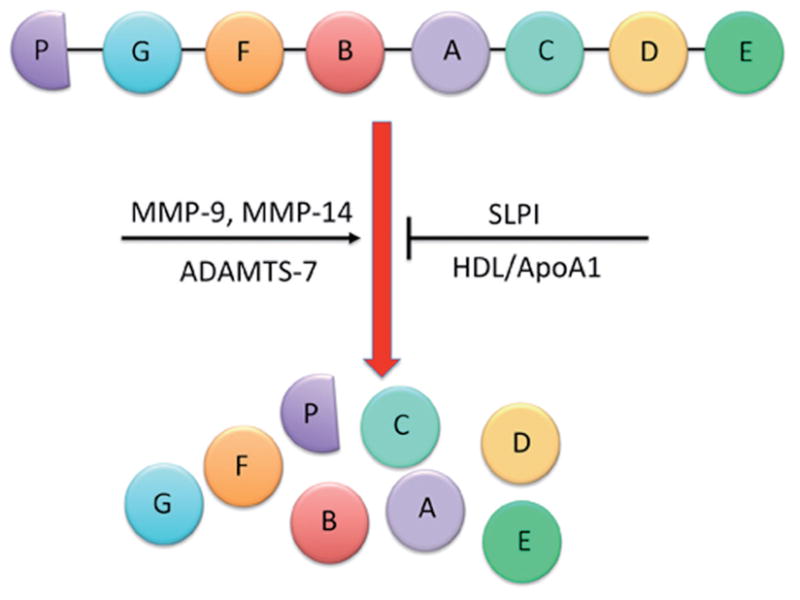
Proteolytic cleavage of PGRN. The human PGRN gene contains a number of granulin-like domains. After secretion, the full-length protein can be proteolytically cleaved between the granulin-like domains by metalloproteinases such as matrix metalloproteinase-9 (MMP-9), MMP-14, and ADAMTS-7. Complete cleavage of full-length PGRN results in granulin peptides (GRN A–G and paragranulin) that are approximately 6 kDa in size, though intermediary cleavage products have also been identified. The proteolytic processing of full-length PGRN can be inhibited when secretory leukocyte protease inhibitor (SLPI) or high-density lipoprotein/apolipoprotein A-1 (HDL/ApoA-1) binds to full-length PGRN to prevent its cleavage
Modes of action of progranulin
To date, no unique receptor for PGRN has been identified. However, PGRN has been shown to bind to the membrane proteins sortilin and tumor necrosis factor receptors (TNFR) 1 and 2 (1). Sortilin was originally identified as a regulator of lysosomal enzyme trafficking (16), but has also been shown to bind and regulate neurotrophic factors such as neurotensin (17) and pro-nerve growth factor (18). It is thought that sortilin mediates the uptake of extracellular PGRN and regulates its internalization within the cell (19,20). The binding of PGRN to the TNFRs, on the other hand, is thought to antagonize TNF-α binding, thereby exerting anti-inflammatory effects (21).
Some of the downstream signaling pathways activated by PGRN include the extracellular signal-regulated kinase (ERK) signaling cascade via the activation of Src homologous and collagen (Shc) protein and p44/42 MAPK, as well as phosphatidylinositol-3 kinase (PI3K) via the activation of Akt (6–8). Activation of these two pathways results in increased expression of Cyclin D1 and Cyclin B and an enhanced proliferation rate (7,22). In addition, PGRN has been shown to activate focal adhesion kinase (FAK) (7,23) and promote the formation of the paxillin/FAK/ERK complex, which promotes cell migration and invasion (23).
Progranulin in cancer
Elevated PGRN expression has been demonstrated in a number of tumor types including ovarian, breast, prostate, renal, liver and esophageal cancers (2,5–8,23–27). In experimental systems, PGRN confers an aggressive phenotype on poorly tumorigenic epithelial cancer cells (2). The malignancy of highly tumorigenic PGRN-expressing cell lines depends on the expression level, since attenuating PGRN mRNA levels greatly inhibits tumor progression (2). Furthermore, PGRN expression has recently been identified as a potential prognostic biomarker for predicting the progression-free survival and overall survival of patients with epithelial ovarian cancer, as patients having significantly higher PGRN levels display poorer prognosis (5,28).
Hepatocellular carcinoma background
Hepatocellular carcinoma (HCC) is the most common type of liver cancer, arising from the malignant transformation of hepatocytes, occurring in most cases as a secondary consequence to hepatitis infections or liver cirrhosis. The incidence of HCC is highest in Asia and Africa where there is high prevalence of Hepatitis B and C, although prevalence in western countries is on the rise (29).
The median survival rate of HCC is approximately 6 months. The prognosis is poor due, in part, to the late presentation of symptoms, large tumors and lack of effective treatment options (29). However, the outcome of patients with HCC has been improved with the introduction of Sorafenib as a test chemotherapeutic agent. Sorafenib is a small inhibitor molecule that targets a number of receptor tyrosine kinases (e.g., VEGF and PCDGF) as well as the Raf kinases (30). In clinical trials, Sorafenib has improved median survival by approximately 3 months (31), although in other trials it proved ineffective as an adjunct therapy (32).
Progranulin in HCC
Using a cDNA microarray approach, PGRN was identified as being upregulated in HCC samples compared to adjacent non-tumor tissue (33,34). Strong PGRN expression was associated with large tumor size, venous infiltration and early intrahepatic recurrence (34). In vitro, the reduction of PGRN expression resulted in decreased cell proliferation, invasion and migration (34). Furthermore, blocking PGRN function in an HCC cell line, using a PGRN-specific neutralizing antibody, inhibited proliferation in vitro and tumor growth in vivo via a mechanism involving the inhibition of the p44/42 MAPK and Akt pathways (26). In addition, treatment with the PGRN-specific neutralizing antibody reduced tumor angiogenesis and vascular endothelial growth factor (VEGF) levels (25).
Cholangiocarcinoma background
Cholangiocarcinoma is a tumor that arises from the malignant transformation of the epithelial cells of the intrahepatic or extrahepatic bile ducts (35). This type of liver cancer has very poor prognosis and is extremely aggressive, with symptoms unobservable until there is bile duct blockage by the tumor (36). Treatment of cholangiocarcinoma by chemotherapy and radiation therapy is not very effective; surgical resection of the tumor is the only treatment option (36). Further study of the factors that lead to tumor initiation, promotion, and progression is necessary for designing alternative treatments for this devastating illness.
The incidence of intrahepatic and extrahepatic cholangiocarcinoma varies by geographic region, with the highest being in Asian countries. Intrahepatic cholangiocarcinoma mortality rates have continuously increased since 1970, conversely, deaths due to extrahepatic cholangiocarcinoma have been decreasing in most countries. Men are slightly more likely to develop cholangiocarcinoma, while incidence increases with age in both sexes (37). The geographic variation of cholangiocarcinoma incidence is partly due to the distribution of risk factors by region and ethnic groups (38). Regional risk factors share the involvement of chronic inflammation and biliary irritation (39). In Asian countries, prevalence of this disease is associated with infection by pathogens including liver flukes, Hepatitis B and Hepatitis C. Meanwhile, in Western countries, 90% of patients diagnosed with cholangiocarcinoma lack any of the reported risk factors (38). However, certain factors are associated with the remaining 10% of cases, including chronic inflammation, primary sclerosing cholangitis, obesity, hepatolithiasis, bacterial infection and/or bile stasis-related chronic cholangitis (40–42).
Progranulin in cholangiocarcinoma
The expression and secretion of PGRN is increased in vitro in cholangiocarcinoma cell lines compared to their non-malignant counterparts (Figure 2) (25). Immunohistochemical analysis of human liver biopsy samples indicated that there is also increased PGRN immunoreactivity in cholangiocarcinoma samples compared to non-malignant controls (Figure 2) (24). In parallel, increased PGRN levels could be detected in the serum (but not bile) from patients with cholangiocarcinoma compared to non-malignant controls (Figure 2) (24).
Figure 2.

PGRN expression and secretion is increased in cholangiocarcinoma. PGRN levels were assessed in four cholangiocarcinoma cell lines as well as non-malignant cholangiocyte cell lines H69 and HIBEC by real time PCR and immunoblotting (A). For real time PCR, data are expressed as average ± SEM (n=4). (*P<0.05 compared to PGRN in H69 cells). Representative PGRN immunoblots are shown (lower panel); β-Actin is shown as a loading control. PGRN levels were also assessed in biopsy samples from 48 cholangiocarcinoma patients and non-malignant controls by immunohistochemistry. Representative photomicrographs of PGRN immunoreactivity are shown (B; magnification ×40). Staining intensity was assessed as described in the methods and expressed as an average ± SEM of all cholangiocarcinoma patients compared to control samples (B; *P<0.05 compared with PGRN immunoreactivity in control biopsy samples). PGRN levels in the supernatant of cell suspensions of cholangiocarcinoma cell lines and the non-malignant cholangiocyte cell lines H69 and HIBEC were determined by EIA after 6 hr (C). Data are expressed as average PGRN concentration (ng/mL) ± SEM (n=3; *P<0.05 compared with PGRN levels secreted from H69 cells). PGRN levels in bile samples from cholangiocarcinoma and intrahepatic cholelithiasis patients were assayed by EIA (D). Data are expressed as average PGRN concentration (ng/mL) ± SEM. (reprinted with permission from Frampton, et al. Gut, 2012;61:268–77) (43)
Interleukin-6 (IL-6) is overexpressed in cholangiocarcinoma and shares a long-standing association with the neoplastic transformation of cholangiocytes to cholangiocarcinoma cells (44,45). Therefore, we hypothesized that IL-6 signaling might be driving the increase in PGRN expression seen in cholangiocarcinoma. Indeed, we demonstrated that increased PGRN expression in cholangiocarcinoma cells is driven by the IL-6-mediated activation of the ERK1/2/RSK1/C/EBP pathway (24).
Increased PGRN secretion was demonstrated to exert subsequent growth-promoting effects on cholangiocarcinoma cells via the activation of Akt and subsequent phosphorylation and nuclear extrusion of Forkhead box protein O1 (FOXO1) (24). A summary of our findings can be found in Figure 3 (24). These data suggest that the upregulation of PGRN may be a key feature associated with the progression of cholangiocarcinoma; inhibiting PGRN expression or function may be a viable target for the development of an effective adjunct therapy to treat this deadly disease.
Figure 3.

Schematic diagram summarizing the major actions of PGRN in CCA. PGRN expression is upregulated in cholangiocarcinoma via IL-6-driven activation of ERK1/2 and RSK1. These signal transduction events in turn activate C/EBP, which is a predominant transcription factor known to regulate PGRN expression. PGRN is then secreted, where it may act on the surrounding cells (autocrine or paracrine) via an unknown receptor, leading to increased Akt activity and subsequent increased phospho-FOXO1. Phosphorylation of FOXO1 causes nuclear extrusion into the cytoplasm, where it is known to bind the chaperone protein 14-3-3. Cytoplasmic location of FOXO1 inhibits its activity as a transcription activator, thereby changing the expression of certain target genes, which ultimately has growth-promoting properties on the cholangiocarcinoma cells (reprinted with permission from Frampton, et al. Gut, 2012; 61:268–77) (43)
Interestingly, we have also demonstrated that PGRN expression and secretion is increased during hyperplastic cholangiocyte proliferation in a rodent model of extrahepatic biliary obstruction (43). Furthermore, PGRN exerted growth-promoting effects on cholangiocytes via the nuclear extrusion and inhibition of FOXO1, although the mechanism by which this occurred is different (43). Specifically, FOXO1 was extruded after phosphorylation via a mechanism involving the Akt signaling pathway in cholangiocarcinoma, whereas in hyperplastic cholangiocyte proliferation PGRN inhibited the expression of the deacetylase Sirt1 and subsequently increased acetylation of FOXO1, which also results in nuclear extrusion (43). One explanation for the difference in the mechanism by which FOXO1 is inhibited may be due to the increased reliance on Akt signaling by cholangiocarcinoma compared to their non-malignant counterparts (46–48).
Progranulin in other GI cancers
Esophageal squamous cell carcinoma
There are two types of esophageal cancer. Adenocarcinoma arises from glandular cells present at the junction of the esophagus and stomach, and squamous cell carcinoma arises from the cells that line the upper part of the esophagus. Research into the role of PGRN in esophageal cancer is sparse. However, Chen et al. (24) demonstrated that PGRN is upregulated in esophageal squamous cell carcinoma compared to normal mucosa, similar to other tumors. PGRN expression correlated with the depth of tumor invasion, lymph node metastasis and TNM classification, as well as microvessel density and VEGF expression (24). The precise mechanism by which PGRN expression is upregulated, and the downstream signaling pathways activated by PGRN in esophageal squamous cell carcinoma are unknown. No information exists concerning the role of PGRN in esophageal adenocarcinoma.
Colorectal cancer
Chemoresistant colorectal cancer cells (CRCs), selected by chronic exposure to high levels of chemotherapeutic drugs, secrete soluble factors that can stimulate growth in the parental chemonaïve cells and make the chemonaïve cells less sensitive to chemotherapy treatment (49). To identify the soluble factors released from the chemoresistant CRCs, proteomic analysis of the conditioned media was performed, demonstrating that PGRN levels (among other factors) were significantly higher in the media from chemoresistant CRCs than the chemonaïve cells (49). Furthermore, treatment of the parental cells with recombinant PGRN conferred a chemoresistant phenotype, suggesting that chemoresistant tumor cells may promote resistance through the release of PGRN and other soluble factors that mediate survival in otherwise chemosensitive tumor cells (49).
Conclusions
Regardless of the tumor type, it is evident that PGRN is upregulated in cancer cells and confers growth-promoting and chemoresistant effects. A role for PGRN in migration, invasion and angiogenesis is also evident. Specific targeting of progranulin may represent an alternative target for the development of therapeutic strategies to combat these gastrointestinal cancers.
Acknowledgments
Portions of these studies were supported by the following grants awarded to Dr. Sharon DeMorrow: American Cancer Society Research Scholar Award (RSC118760), and NIH K01 Award (DK078532) and NIH R03 Award (DK088012). This material is the result of work supported with resources and the use of facilities at the Central Texas Veterans Health Care System in Temple, TX.
The authors acknowledges Mr. Gabriel Frampton for assistance in proof reading this review.
Footnotes
Disclosure: The author declares no conflict of interest.
References
- 1.De Muynck L, Van Damme P. Cellular effects of progranulin in health and disease. J Mol Neurosci. 2011;45:549–60. doi: 10.1007/s12031-011-9553-z. [DOI] [PubMed] [Google Scholar]
- 2.He Z, Bateman A. Progranulin (granulin-epithelin precursor, pc-cell-derived growth factor, acrogranin) mediates tissue repair and tumorigenesis. J Mol Med. 2003;81:600–12. doi: 10.1007/s00109-003-0474-3. [DOI] [PubMed] [Google Scholar]
- 3.Jian J, Konopka J, Liu C. Insights into the role of progranulin in immunity, infection, and inflammation. J Leukoc Biol. 2013;93:199–208. doi: 10.1189/jlb.0812429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cruts M, Van Broeckhoven C. Loss of progranulin function in frontotemporal lobar degeneration. Trends Genet. 2008;24:186–94. doi: 10.1016/j.tig.2008.01.004. [DOI] [PubMed] [Google Scholar]
- 5.Han JJ, Yu M, Houston N, et al. Progranulin is a potential prognostic biomarker in advanced epithelial ovarian cancers. Gynecol Oncol. 2011;120:5–10. doi: 10.1016/j.ygyno.2010.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.He Z, Ismail A, Kriazhev L, et al. Progranulin (pc-cell-derived growth factor/acrogranin) regulates invasion and cell survival. Cancer Res. 2002;62:5590–6. [PubMed] [Google Scholar]
- 7.Lu R, Serrero G. Mediation of estrogen mitogenic effect in human breast cancer mcf-7 cells by pc-cell-derived growth factor (pcdgf/granulin precursor) Proc Natl Acad Sci U S A. 2001;98:142–7. doi: 10.1073/pnas.011525198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zanocco-Marani T, Bateman A, Romano G, et al. Biological activities and signaling pathways of the granulin/epithelin precursor. Cancer Res. 1999;59:5331–40. [PubMed] [Google Scholar]
- 9.Bhandari V, Bateman A. Structure and chromosomal location of the human granulin gene. Biochem Biophys Res Commun. 1992;188:57–63. doi: 10.1016/0006-291x(92)92349-3. [DOI] [PubMed] [Google Scholar]
- 10.Bhandari V, Palfree RG, Bateman A. Isolation and sequence of the granulin precursor cdna from human bone marrow reveals tandem cysteine-rich granulin domains. Proc Natl Acad Sci U S A. 1992;89:1715–9. doi: 10.1073/pnas.89.5.1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Shoyab M, McDonald VL, Byles C, et al. Epithelins 1 and 2: Isolation and characterization of two cysteine-rich growth-modulating proteins. Proc Natl Acad Sci U S A. 1990;87:7912–6. doi: 10.1073/pnas.87.20.7912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Xu D, Suenaga N, Edelmann MJ, et al. Novel MMP-9 substrates in cancer cells revealed by a label-free quantitative proteomics approach. Mol Cell Proteomics. 2008;7:2215–28. doi: 10.1074/mcp.M800095-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Butler GS, Dean RA, Tam EM, et al. Pharmacoproteomics of a metalloproteinase hydroxamate inhibitor in breast cancer cells: Dynamics of membrane type 1 matrix metalloproteinase-mediated membrane protein shedding. Mol Cell Biol. 2008;28:4896–914. doi: 10.1128/MCB.01775-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bai XH, Wang DW, Kong L, et al. Adamts-7, a direct target of pthrp, adversely regulates endochondral bone growth by associating with and inactivating gep growth factor. Mol Cell Biol. 2009;29:4201–19. doi: 10.1128/MCB.00056-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhu J, Nathan C, Jin W, et al. Conversion of proepithelin to epithelins: Roles of slpi and elastase in host defense and wound repair. Cell. 2002;111:867–78. doi: 10.1016/s0092-8674(02)01141-8. [DOI] [PubMed] [Google Scholar]
- 16.Canuel M, Libin Y, Morales CR. The interactomics of sortilin: An ancient lysosomal receptor evolving new functions. Histol Histopathol. 2009;24:481–92. doi: 10.14670/HH-24.481. [DOI] [PubMed] [Google Scholar]
- 17.Mazella J. Sortilin/neurotensin receptor-3: a new tool to investigate neurotensin signaling and cellular trafficking? Cell Signal. 2001;13:1–6. doi: 10.1016/s0898-6568(00)00130-3. [DOI] [PubMed] [Google Scholar]
- 18.Nykjaer A, Lee R, Teng KK, et al. Sortilin is essential for prongf-induced neuronal cell death. Nature. 2004;427:843–8. doi: 10.1038/nature02319. [DOI] [PubMed] [Google Scholar]
- 19.Carrasquillo MM, Nicholson AM, Finch N, et al. Genome-wide screen identifies rs646776 near sortilin as a regulator of progranulin levels in human plasma. Am J Hum Genet. 2010;87:890–7. doi: 10.1016/j.ajhg.2010.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hu F, Padukkavidana T, Vaegter CB, et al. Sortilin-mediated endocytosis determines levels of the frontotemporal dementia protein, progranulin. Neuron. 2010;68:654–67. doi: 10.1016/j.neuron.2010.09.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tang W, Lu Y, Tian QY, et al. The growth factor progranulin binds to tnf receptors and is therapeutic against inflammatory arthritis in mice. Science. 2011;332:478–84. doi: 10.1126/science.1199214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lu R, Serrero G. Inhibition of pc cell-derived growth factor (pcdgf, epithelin/granulin precursor) expression by antisense pcdgf cdna transfection inhibits tumorigenicity of the human breast carcinoma cell line mda-mb-468. Proc Natl Acad Sci U S A. 2000;97:3993–8. doi: 10.1073/pnas.97.8.3993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Monami G, Gonzalez EM, Hellman M, et al. Proepithelin promotes migration and invasion of 5637 bladder cancer cells through the activation of erk1/2 and the formation of a paxillin/fak/erk complex. Cancer Res. 2006;66:7103–10. doi: 10.1158/0008-5472.CAN-06-0633. [DOI] [PubMed] [Google Scholar]
- 24.Chen XY, Li JS, Liang QP, et al. Expression of pc cell-derived growth factor and vascular endothelial growth factor in esophageal squamous cell carcinoma and their clinicopathologic significance. Chin Med J. 2008;121:881–6. [PubMed] [Google Scholar]
- 25.Frampton G, Invernizzi P, Bernuzzi F, et al. Interleukin-6-driven progranulin expression increases cholangiocarcinoma growth by an akt-dependent mechanism. Gut. 2012;61:268–77. doi: 10.1136/gutjnl-2011-300643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ho JC, Ip YC, Cheung ST, et al. Granulin-epithelin precursor as a therapeutic target for hepatocellular carcinoma. Hepatology. 2008;47:1524–32. doi: 10.1002/hep.22191. [DOI] [PubMed] [Google Scholar]
- 27.Pan CX, Kinch MS, Kiener PA, et al. PC cell-derived growth factor expression in prostatic intraepithelial neoplasia and prostatic adenocarcinoma. Clin Cancer Res. 2004;10:1333–7. doi: 10.1158/1078-0432.ccr-1123-03. [DOI] [PubMed] [Google Scholar]
- 28.Cuevas-Antonio R, Cancino C, Arechavaleta-Velasco F, et al. Expression of progranulin (acrogranin/pcdgf/granulin-epithelin precursor) in benign and malignant ovarian tumors and activation of mapk signaling in ovarian cancer cell line. Cancer Invest. 2010;28:452–8. doi: 10.3109/07357900903346455. [DOI] [PubMed] [Google Scholar]
- 29.Blechacz B, Mishra L. Hepatocellular carcinoma biology. Recent Results Cancer Res. 2013;190:1–20. doi: 10.1007/978-3-642-16037-0_1. [DOI] [PubMed] [Google Scholar]
- 30.Hotte SJ, Hirte HW. Bay 43-9006: Early clinical data in patients with advanced solid malignancies. Curr Pharm Des. 2002;8:2249–53. doi: 10.2174/1381612023393053. [DOI] [PubMed] [Google Scholar]
- 31.Bai W, Wang YJ, Zhao Y, et al. Sorafenib in combination with transarterial chemoembolization improves survival of unresectable hepatocellular carcinoma: A propensity-score matching study. J Dig Dis. 2013;14:181–90. doi: 10.1111/1751-2980.12038. [DOI] [PubMed] [Google Scholar]
- 32.Keating GM, Santoro A. Sorafenib: A review of its use in advanced hepatocellular carcinoma. Drugs. 2009;69:223–40. doi: 10.2165/00003495-200969020-00006. [DOI] [PubMed] [Google Scholar]
- 33.Chen X, Cheung ST, So S, et al. Gene expression patterns in human liver cancers. Mol Biol Cell. 2002;13:1929–39. doi: 10.1091/mbc.02-02-0023.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cheung ST, Wong SY, Leung KL, et al. Granulin-epithelin precursor overexpression promotes growth and invasion of hepatocellular carcinoma. Clin Cancer Res. 2004;10:7629–36. doi: 10.1158/1078-0432.CCR-04-0960. [DOI] [PubMed] [Google Scholar]
- 35.Alpini A, Prall RT, LaRusso NF. The pathobiology of biliary epithelia. In: Arias IM, Boyer JL, Chisari FV, et al., editors. The Liver; Biology & Pathobiology, 4E. Philadelphia, PA: Lippincott Williams & Wilkins; 2001. pp. 421–35. [Google Scholar]
- 36.Sirica AE. Cholangiocarcinoma: Molecular targeting strategies for chemoprevention and therapy. Hepatology. 2005;41:5–15. doi: 10.1002/hep.20537. [DOI] [PubMed] [Google Scholar]
- 37.Patel T. Worldwide trends in mortality from biliary tract malignancies. BMC Cancer. 2002;2:10. doi: 10.1186/1471-2407-2-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ben-Menachem T. Risk factors for cholangiocarcinoma. Eur J Gastroenterol Hepatol. 2007;19:615–7. doi: 10.1097/MEG.0b013e328224b935. [DOI] [PubMed] [Google Scholar]
- 39.Gores GJ. Cholangiocarcinoma: Current concepts and insights. Hepatology. 2003;37:961–9. doi: 10.1053/jhep.2003.50200. [DOI] [PubMed] [Google Scholar]
- 40.Catalano OA, Sahani DV, Forcione DG, et al. Biliary infections: Spectrum of imaging findings and management. Radiographics. 2009;29:2059–80. doi: 10.1148/rg.297095051. [DOI] [PubMed] [Google Scholar]
- 41.Chen MF. Peripheral cholangiocarcinoma (cholangiocellular carcinoma): Clinical features, diagnosis and treatment. J Gastroenterol Hepatol. 1999;14:1144–9. doi: 10.1046/j.1440-1746.1999.01983.x. [DOI] [PubMed] [Google Scholar]
- 42.de Groen PC, Gores GJ, LaRusso NF, et al. Biliary tract cancers. N Engl J Med. 1999;341:1368–78. doi: 10.1056/NEJM199910283411807. [DOI] [PubMed] [Google Scholar]
- 43.Frampton G, Ueno Y, Quinn M, et al. The novel growth factor, progranulin, stimulates mouse cholangiocyte proliferation via sirtuin-1-mediated inactivation of FOXO1. Am J Physiol Gastrointest Liver Physiol. 2012;303:G1202–11. doi: 10.1152/ajpgi.00104.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Park J, Tadlock L, Gores GJ, et al. Inhibition of interleukin 6-mediated mitogen-activated protein kinase activation attenuates growth of a cholangiocarcinoma cell line. Hepatology. 1999;30:1128–33. doi: 10.1002/hep.510300522. [DOI] [PubMed] [Google Scholar]
- 45.Yokomuro S, Tsuji H, Lunz JG, 3rd, et al. Growth control of human biliary epithelial cells by interleukin 6, hepatocyte growth factor, transforming growth factor beta1, and activin a: Comparison of a cholangiocarcinoma cell line with primary cultures of non-neoplastic biliary epithelial cells. Hepatology. 2000;32:26–35. doi: 10.1053/jhep.2000.8535. [DOI] [PubMed] [Google Scholar]
- 46.Kobayashi S, Werneburg NW, Bronk SF, et al. Interleukin-6 contributes to mcl-1 up-regulation and trail resistance via an akt-signaling pathway in cholangiocarcinoma cells. Gastroenterology. 2005;128:2054–65. doi: 10.1053/j.gastro.2005.03.010. [DOI] [PubMed] [Google Scholar]
- 47.Leelawat K, Leelawat S, Narong S, et al. Roles of the MEK1/2 and AKT pathways in CXCL12/CXCR4 induced cholangiocarcinoma cell invasion. World J Gastroenterol. 2007;13:1561–8. doi: 10.3748/wjg.v13.i10.1561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xu X, Kobayashi S, Qiao W, et al. Induction of intrahepatic cholangiocellular carcinoma by liver-specific disruption of Smad4 and Pten in mice. J Clin Invest. 2006;116:1843–52. doi: 10.1172/JCI27282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bose D, Zimmerman LJ, Pierobon M, et al. Chemoresistant colorectal cancer cells and cancer stem cells mediate growth and survival of bystander cells. Br J Cancer. 2011;105:1759–67. doi: 10.1038/bjc.2011.449. [DOI] [PMC free article] [PubMed] [Google Scholar]