

Abstract
The development of reading skills in typical students is commonly described as a rapid growth across early grades of active reading education, with a slowing down of growth as active instruction tapers. This study examined the extent to which genetics and environments influence these growth rates. Participants were 371 twin pairs, aged approximately 6 through 12, from the Western Reserve Reading Project. Development of word-level reading, reading comprehension, and rapid naming was examined using genetically sensitive latent quadratic growth curve modeling. Results confirmed the developmental trajectory described in the phenotypic literature. Furthermore, the same shared environmental influences were related to early reading skills and subsequent growth, but genetic influences on these factors were unique.
Reading ability is an essential foundation of all other academic skills (National Reading Panel, 2000). Consequently, understanding the developmental course and successful acquisition and application of reading skills has important academic and clinical implications. If beginning readers do not develop into mature, fluent, and comprehending readers, there are serious lifelong implications, including a greater likelihood of high school dropout, increased substance abuse, and greater incidence of behavior problems (Lyon, 2002; Morgan, Farkas, Tuffs, & Sperling, 2008; Snow, Burns, & Griffin, 1998).
Reading skills in kindergarten and first grade are often significant predictors of later reading skills, and are also associated with the rate of change in reading skill (Butler, Marsh, Sheppard, & Sheppard, 1985; Cunningham & Stanovich, 1997; Schatschneider, Fletcher, Francis, Carlson, & Foorman, 2004; Wagner & Torgesen, 1987). Although average patterns of development are observable, research has also demonstrated that there is considerable variability in the initial reading level when children begin formalized instruction, and also in the rate of subsequent growth (Parrila, Aunola, Leskinen, Nurmi, & Kirby, 2005). Research has identified several constructs that influence the variability in children’s reading skills, including environmental influences such as socioeconomic status (Reardon & Robinson, 2007), the type and quality of teacher instruction (Connor et al., 2009), and parental beliefs and behaviors (Phillips & Lonigan, 2005). Cognitive measures such as phonological awareness (Wagner & Torgesen, 1987) and letter knowledge (Scanlon & Vellutino, 1996) have also shown significant relations with reading ability, for both concurrent and predictive measurement (Schatschneider et al., 2004).
In general, growth in reading skills from early reading to mature, fluent reading has been found to be fastest during the early grades, and relatively slower after third grade (Foorman, Francis, Fletcher, Schatschneider, & Mehta, 1998). More specifically, a study by Hill, Bloom, Black, and Lipsey (2007) obtained the effect size of annual expected gain at each of 13 years of school by averaging across seven nationally normed reading tests. The average expected gain was very large in kindergarten (d = 1.52) but shrank to a moderate effect size by early elementary school (d = 0.60 in Grade 2) and to an even smaller effect size by middle school (d = 0.23 in Grade 6). By Grade 11, almost no yearly gains were expected on standardized reading tests (d = 0.06; Hill et al., 2007). Although this study only provides cross-sectional evidence, it implies that there are changes in reading (and growth in reading) such that the growth rate of reading slows down as children age.
Other studies have approached the question of reading development using growth models. These studies have demonstrated individual differences in the growth rates of children’s reading (and related component) skills (Foorman et al., 1998; McCoach, O’Connell, Reis, & Levitt, 2006; Skibbe et al., 2008). Research also suggests that individual differences in student reading growth are related to both cognitive and environmental influences (Foorman et al., 1998; McCoach et al., 2006; Nye, Konstantopoulos, & Hedges, 2004; Torgesen, Wagner, Rashotte, Burgress, & Hecht, 1997; Torgesen et al., 1999).
Reading ability and development have also been studied in the quantitative genetics literature. Quantitative genetics studies have established that variance in reading ability, reading disability, and their underlying skills (such as vocabulary, phonological awareness, decoding, and fluency) are significantly influenced by both genetics and the environment (Byrne et al., 2005; Petrill, Deater-Deckard, Thompson, DeThorne, & Schatschneider, 2006b; Stevenson, Graham, Fredman, & McLoughlin, 1987). Recent quantitative genetics studies have also begun to examine how genetic and environmental attributions to variance in reading ability (and its underlying skills) vary as children get older and become more fluent readers (Betjemann et al., 2008; Byrne et al., 2007; Harlaar, Dale, & Plomin, 2007; Petrill et al., 2007; Wadsworth, DeFries, Olson, & Willcutt, 2007). For example, reading fluency has been shown to demonstrate a consistently significant influence of genetics across developmental ages, while phonological awareness has shown an increasing proportion of variance attributable to genetics as children get older (Byrne et al., 2005; Petrill et al., 2007). Other studies have directly assessed the stability of reading over time, and these studies suggest the variance in reading skill stability is primarily attributable to genetic effects, with some shared environmental influences, particularly in early reading (Betjemann et al., 2008; Byrne et al., 2007; Harlaar et al., 2007; Petrill et al., 2007; Wadsworth et al., 2007).
To review, the phenotypic reading development literature suggests that substantial individual differences in growth exist, and these growth rates are predicted from, and influenced by, indices of the learning environment. At the same time, behavioral genetic studies suggest that the majority of the variance in reading outcomes is attributable to significant genetic effects, and these effects are stable over time. These two results appear to be at odds with each other. To elucidate this question, it is necessary to combine the two approaches: to examine the behavior genetic influences in the development of reading skills over time.
To date, only one study has examined the genetic and environmental influences on children’s reading growth. Petrill et al. (2010) examined the growth in several early reading skills at ages 6, 7, and 8 using latent growth curve models. As expected from the larger quantitative genetic literature, the authors found significant genetic and environmental influences on reading skills of children at age 6. This study also demonstrated that the slope (or rate of change) was influenced by both genetic and environmental factors for phonological awareness and rapid naming, but only by the shared environment for letter identification, word identification, and phonemic decoding (Petrill et al., 2010). However, there were several shortcomings. First, Petrill et al. included three measurement occasions, and thus could not estimate the curvilinear growth trajectory of reading, as suggested by the phenotypic literature. More important, Petrill et al. focused on early reading skills, and thus did not examine a broader span in which reading skills are acquired and consolidated. Finally, because of the focus on early reading, Petrill et al. did not examine growth in reading comprehension, which is the ultimate goal of reading acquisition.
For this reason, this study expands upon the work of Petrill et al. (2010) to conduct the first quantitative genetic analysis of the growth of reading skill from early elementary school into early adolescence. This is important as this developmental period covers a span of schooling from early intensive reading instruction to instruction not focused on reading but instead on using reading to learn other subjects (Chall, 1983). Using a twin sample, which allows for the general description of the genetic and environmental influences during this key developmental period, we will be able to understand the etiology of the trajectories of developing readers from preschool through fifth grade. Furthermore, the current study is the first to assess the genetic and environmental influences of growth in reading comprehension, which is the ultimate goal of reading development. In addition, using six annual assessment points, we will also be the first to examine the genetic and environmental influences on a growth model of academic skill development, which allows us to model the commonly described phenotypic pattern of a slowing down of reading development. Finally, we have the opportunity to examine the extent to which genetic and environmental influences on the intercept are unique from or overlap with influences on subsequent growth, which will provide insight into the potential sources of variance that are influencing each of these factors.
The goals of this study are to (a) examine phenotypic growth models of reading development, including reading comprehension, across 6 years of development; (b) to identify the extent to which genetic and environmental influences contributed to variance in the initial level of reading, and the linear and quadratic rates of growth; and (c) identify the extent to which the genetic and environmental influences are shared between (or unique to) initial status with linear growth, initial status with quadratic growth, and linear growth with quadratic growth.
Method
Participants
Participants of this study are enrolled in an ongoing longitudinal twin study in Ohio, the Western Reserve Reading Project (WRRP). In the WRRP, pairs of twins are assessed in their home on reading and mathematics. Six annual visits focused on reading, with an additional supplemental visit focusing on mathematics. This mathematics visit occurred at the 6-month point between the third and fourth reading visits. Recruiting was conducted through Ohio state birth records, school nomination, and media announcements. At the time of each home visit, parents are informed of the study’s main goals and asked to consent to their children’s participation. Children are also asked to assent to their own participation.
The data used in this study were collected during the six waves of annual home visit testing that focused on reading skills (referred to throughout as Assessments 1–6). At the first assessment point, twin pairs were in preschool (about to enter kindergarten, n = 82), kindergarten (n = 155), or first grade (n = 78), and were on average approximately 6 years old (Mage = 6.1 years, SD = 0.68, range = 5.2–7.9 years). At the sixth assessment period, children were approximately 12 years old (Mage = 12.17, SD = 1.02, range = 10.0–14.6 years). The final number of same-sex twin pairs included 371 monozygotic (MZ; n = 158) and 213 dizygotic (DZ) twin pairs. Although slightly negatively skewed (skew = −0.07), parent education levels varied widely and were similar for fathers and mothers: 10% had a high school education or less, 16% attended some college, 32% had a bachelor’s degree, 20% had some postgraduate training, and 5% did not specify. Most families were two-parent households (92%) and nearly all were European American (92% of mothers, 94% of fathers).
Procedure and Measures
In this study, we examined growth of reading and related skills, including rapid naming, word identification, and nonword decoding, and higher level reading comprehension skill. All test sessions were conducted in the twins’ home in separate rooms by separate testers, and the total time to complete all testing was approximately 90 min per child.
Rapid naming was assessed using the Rapid Letter and Number Naming subtests from the Comprehensive Test of Phonological Processing (Wagner, Torgesen, & Rashotte, 1999). The rapid letter and digit naming assess the length of time required for participants to name presented sets of letters and digits, and have high published reliability (α = .83 and α = .73, respectively). As the Letter and Number Naming subtests were highly correlated (r = .73), they were combined into a composite score. This was then reverse scored so that a higher score reflected a better score. Word identification and nonword decoding were assessed with two subtests of the Woodcock Reading Mastery Test (WRMT; Woodcock, 1987). The Word Identification subtest requires students to recognize and read real words aloud. Published split half reliability for this subtest is .95 (Woodcock, 1987). The Nonword Reading sub-test of the WRMT requires participants to decode (pronounce) a list of nonwords (e.g., baf). The published reliability for the test is .94 (Woodcock, 1987). Reading comprehension was also assessed with the WRMT. The Reading Comprehension subtest of the WRMT requires children to read a short passage and identify a missing key word to complete a sentence at the end of the passage. The published median reliability for the test is .83.
Results
The primary goal of this study was to examine the genetic and environmental influences on growth in reading skills across several years of development. First, phenotypic models of growth were estimated. Second, latent quadratic growth models were fit in a genetically sensitive design to determine what proportion of the variance in the initial status and rate of change in each reading outcome was attributable to genetic and environmental components. Finally, the model was examined to determine whether genetic and environmental influences were shared between intercept, linear slope, and quadratic slope, or whether they were unique.
Prior to modeling, the subtests from the WRMT were all converted to W scores using the WRMT scoring protocol. W scores are based on item response theory, and as a result, all scores have equal measurement intervals (i.e., it takes the same amount of ability to move between a score of 110 and a score of 111 as it does to move between a score of 210 and 211). W scores were not available for the rapid naming test, so raw composite scores were used. The mean W scores and raw scores for the sample used in this study are reported in Table 1. In addition, additive genetic estimates of heritability (h2) and shared environment (c2) are reported for the raw scores of each measure at each time point (the method for deriving these estimates is described in detail in subsequent sections). For example, 30% of the variance in word attack at Assessment 1 was attributable to heritability and 58% of the variance was attributable to shared environmental effects (Table 1).
Table 1.
Means and Standard Deviations of W-Scores and Raw Scores, Estimates of Heritability and Shared Environment, and Results of Significance Tests for Each Measure at Each Assessment Point
|
W score
|
Raw
|
h2 | c2 | |||
|---|---|---|---|---|---|---|
| M | SD | M | SD | |||
| WRMT: Word Identification | ||||||
| Assessment 1 | 386.56 | 40.1 | 15.3 | 17 | 0.2* | 0.74* |
| Assessment 2 | 438.31 | 37.27 | 39.6 | 19 | 0.33* | 0.6* |
| Assessment 3 | 470.73 | 29.11 | 56.7 | 15 | 0.4* | 0.5* |
| Assessment 4 | 495.81 | 20.88 | 70.2 | 11 | 0.59* | 0.25* |
| Assessment 5 | 506.31 | 17.32 | 76.1 | 9 | 0.66* | 0.15 ns |
| Assessment 6 | 514.09 | 15.1 | 80.6 | 8 | 0.82* | 0 ns |
| WRMT: Word Attack | ||||||
| Assessment 1 | 457.84 | 19.66 | 5.3 | 7 | 0.3* | 0.58* |
| Assessment 2 | 481.15 | 18.77 | 15.3 | 10 | 0.27* | 0.52* |
| Assessment 3 | 495.19 | 16.18 | 23.4 | 10 | 0.58* | 0.23 ns |
| Assessment 4 | 504.41 | 14.29 | 29.2 | 8 | 0.75* | 0 ns |
| Assessment 5 | 509.21 | 13.51 | 31.8 | 7 | 0.47* | 0.24 ns |
| Assessment 6 | 510.88 | 11.94 | 32.9 | 6 | 0.71* | 0.01 ns |
| CTOPP: Rapid Naming | ||||||
| Assessment 1 | — | — | 382.02 | 31 | 0.51* | 0.25 ns |
| Assessment 2 | — | — | 355.39 | 21 | 0.61* | 0.22 ns |
| Assessment 3 | — | — | 343.75 | 13 | 0.44* | 0.3* |
| Assessment 4 | — | — | 335.59 | 9 | 0.31* | 0.33* |
| Assessment 5 | — | — | 331.41 | 7 | 0.69* | 0.02 ns |
| Assessment 6 | — | — | 328.71 | 6 | 0.45* | 0.26 ns |
| WRMT: Reading Comprehension | ||||||
| Assessment 1 | 448.37 | 20.18 | 11.01 | 9 | 0.39* | 0.39 ns |
| Assessment 2 | 469.41 | 20.61 | 21.47 | 10 | 0.44* | 0.47* |
| Assessment 3 | 486.97 | 17.2 | 31.01 | 9 | 0.32* | 0.5* |
| Assessment 4 | 500.6 | 13.99 | 38.44 | 7 | 0.72* | 0.05 ns |
| Assessment 5 | 508.24 | 11.83 | 42.68 | 6 | 0.55* | 0.23 ns |
| Assessment 6 | 513.92 | 12.47 | 45.97 | 7 | 0.82* | 0 ns |
Note. WRMT = Woodcock Reading Mastery Test; CTOPP = Comprehensive Test of Phonological Processing; h2 = proportion of raw-score variance attributable to heritability; c2 = proportion of raw-score variance attributable to shared environment. Mean ages at Assessments 1–6 were 6.1, 7.2, 8.2, 9.8, 10.9, and 12.1.
p < .05.
Phenotypic Latent Growth Modeling
Typical applications of latent growth modeling occur in two phases: an unconditional phase, wherein the goal is to identify the underlying shape of growth, and a conditional phase, wherein the variability around the mean intercept, mean linear growth, and mean quadratic growth is explained through the use of covariates. The unconditional phase is important because it is well documented in the developmental literature that if the shape of growth is not correctly defined in the unconditional phase, the results of the conditional phase are untrustworthy and inaccurate (Raudenbush & Bryk, 2002). One hallmark of the latent growth curve is the ability to fit individual growth curves for each participant. In other words, a latent quadratic growth curve model can estimate an intercept, a slope, and a quadratic slope for each participant in the study. The intercept represents each person’s estimated score at whatever point the linear and quadratic terms are set to zero, the linear slope represents each person’s estimated rate of change at the centering point, and the quadratic slope parameter allowed each person’s growth trajectory to be curvilinear: to accelerate or decelerate.
In the unconditional phase of the current study, latent growth curve modeling was used to determine the shape of phenotypic growth. For these models, both twins were included and potential shared family variance was ignored because such nonindependence only impacts the standard errors of the estimates, and the main concern in these analyses identifying the shape of growth (not identifying significant predictors of model parameters). Phenotypic growth models were estimated separately for each reading outcome using Mplus software (Muthén & Muthén, 1998–2004), and data from all participants were included in each model. The shape of growth was determined through model fit comparisons. Results of the chi-square model comparisons tests for the unconditional phase of each outcome indicated that a model that allows for randomly varying intercepts, slopes, and quadratic terms yielded the best fit to the data. The depiction of the structural equation model for the final phenotypic growth model is presented in the bottom half of Figure 1. Specifically, the model contains a latent intercept (I), linear slope (S), and quadratic slope (Q), each loading on to the six assessment points. Because the intercept factor is similar to the intercept parameter in a typical regression equation, each of the unstandardized loadings to this factor was fixed at 1.0. This is in contrast to the linear slope, where the loadings increase incrementally by one at each time point, which results in the slope parameter representing the amount of change in reading score in 1 year. The quadratic function (allowing the slope to accelerate or decelerate) is modeled by weighting the quadratic pathways to each assessment point as squares of the weights given to the linear pathways.
Figure 1.

Genetically sensitive latent quadratic growth model. I = intercept; S = linear slope; Q = quadratic slope; ASMT1 = Assessment 1.
The parameter estimates for the final phenotypic growth models are presented in Table 2. Each of the four examined reading outcomes suggested a positive linear growth term but a significantly negative quadratic term. This indicates that while the general trend of growth was positive across the six time points, there was also a significant deceleration effect as children get older (Table 2). This basic shape was the same across all four examined outcomes. For example, the fitted estimates for growth in Word Identification have been plotted in the Figure 2. Note that the trajectory of growth is steep in the first three assessment points, but begins to decelerate at the older ages. Note also that the fitted estimates presented in Figure 2 map on almost perfectly to the observed means at each assessment point (presented in Table 1).
Table 2.
Phenotypic Quadratic Growth Model Estimates for the Four Examined Reading Outcomes
| Intercept | Linear slope | Quadratic slope | |
|---|---|---|---|
| Word Identification | |||
| M | 384 | 53 | −5.6 |
| SE | 1.65 | 0.83 | 0.12 |
| Word Attack | |||
| M | 457 | 23 | −2.6 |
| SE | 0.79 | 0.46 | 0.07 |
| Rapid Naming | |||
| M | −76 | 19 | −2.05 |
| SE | 1.22 | 0.61 | 0.08 |
| Reading Comprehension | |||
| M | 439 | 28 | −2.8 |
| SE | 11.34 | 0.73 | 0.11 |
Figure 2.

Phenotypic fitted growth curve of Word Identification across six waves of data collection.
It is important to highlight that the reported coefficients in these models are estimated mean intercept, linear slope, and quadratic slope values. Because an individual growth curve is fitted for each person in the latent growth modeling process, considerable variability can exist around each of these estimates (denoted, for one, by the standard errors around each estimate in Table 2). The intercept and growth rates (linear and quadratic) can be correlated with one another, but are not necessarily so. This means that the rank order of individuals on the intercept (where they start) and the rank order for the speed at which they grow (linear slope) and the rank order of the speed with which their individual growth rate accelerates or decelerates (quadratic slope) are not perfectly related. For example, the negative correlation between the intercept and slope indicates that students who began the study high on decoding ability tended to have a slower growth rate than those who started the study with low decoding skills. While these correlations are interesting, it is only the variance in intercepts and slopes that is predicted from covariates during the conditional phase of model building.
In this study, we took an alternative approach to the use of covariates to explain the variability in intercepts and slopes. In the conditional phase of growth modeling, we used cotwin relatedness as a predictor of this variability, which allowed us to partition the variability in intercepts and slopes into proportions due to genetics, shared environment, and nonshared environment.
Genetically Sensitive Latent Growth Modeling
Quantitative genetic models estimate the proportion of variance due to genetic effects by examining how much more similar MZ (identical) twins are than DZ (fraternal) twins. Because MZ twins share 100% of their segregating genes, the extent to which the MZ twins are more similar than the DZ twins is an indication of the importance of genetic influences. Because MZ and DZ twins share 100% of their shared environments (live in the same homes, share the same parents, etc.), the extent to which MZ and DZ twins are equally as similar to one another is an indication of shared environmental influences.
Analyses were conducted with individual-level data using Mx (Neale, Boker, Xie, & Maes, 2006). Four different models were fitted, one for each reading outcome. Although model comparisons in the phenotypic stage indicated that a quadratic model was the best fit to the data, that assumption was also tested in the genetically sensitive model. Results aligned and indicated that a quadratic model, which required 10 additional paths to be estimated over the linear model, fit the data significantly better than a linear growth model, Word Identification, χ2(10) = 504, p < .01; Word Attack, χ2(10) = 224, p < .01; Rapid Naming, χ2(10) = 928, p < .01; and Reading Comprehension; χ2(10) = 376, p < .01.
A visual representation of the decomposed quadratic growth model that was fitted to each outcome is presented in Figure 1. The bottom portion of Figure 1 is the phenotypic growth model described previously. The top portion of Figure 1 represents the genetic decomposition of the latent intercept, latent linear slope, and latent quadratic slope factors. Note that each of these latent variables is decomposed into genetic (A1, A2, and A3; represented by the letter A for “additive genetics”), shared environment (C1, C2, and C3; represented by the letter C for “common environment”), and nonshared environment (E1, E2, and E3) effects. In addition, path estimates were included to estimate whether the genetic, shared environmental, and nonshared environmental influences on each latent variable (I, S, and Q) were shared or unique. To account for age, age at Assessment 1 (centered at age 6) was modeled as a definition variable. This allowed the model to account for age differences within assessment points while also accounting for the covariance among the latent intercept, latent linear slope, and latent quadratic slope (Neale, Boker, Xie, & Maes, 2006). Because intercept and growth were parameterized as latent constructs, the random measurement error was minimized; thus, the influences in growth attributable to the specific environment (E1, E2, and E3) was expected to be minimal. Note that the data were also modeled with correlated cotwin errors, as is done in Reynolds et al. (2005). No differences in the parameter estimates or significance patterns of genetic decomposition of the growth models were observed between the model with correlated errors and the presented model for any examined outcome.
Due to the complexity of the model, Figure 1 represents only one twin. When the model is estimated, this model is fit simultaneously to each of two twins in a pair, and both twins are represented with this same model. The covariance between twins is modeled through model constraints. Specifically, paths between latent variables representing heritability are constrained to be equal for MZ twins, but 50% for DZ twins. By contrast, paths between the latent variables representing shared environmental influences (represented by the letter C) are constrained to be equal within each twin pair. Petrill et al. (2010) presents a graphical depiction of a more basic growth model, which includes a detailed view of the cotwin constraints.
The estimated proportions of variance in the intercept, linear slope, and quadratic slope are presented with confidence intervals in Table 3. The results of the quadratic growth models suggested significant genetic, shared environmental, and nonshared environmental influences on the intercepts of each measured reading outcome (Table 3, columns 1, 2, and 3). There were also significant genetic influences present for the linear slope of Word Identification, Word Attack, and Rapid Naming. Shared environmental influences on the linear slope were significant for all four examined outcomes. In addition, both Reading Comprehension and Rapid Naming showed significant nonshared environmental influences on the linear slope. Finally, the pattern of significance for the decomposition of the quadratic slope was the same as that of the linear slope (Table 3, columns 7, 8, and 9).
Table 3.
Estimates and Confidence Intervals (in Parentheses) for Genetic and Environmental Components of Variance in Latent Quadratic Growth Model Parameters
| Intercept
|
Linear slope
|
Quadratic slope
|
|||||||
|---|---|---|---|---|---|---|---|---|---|
| Genetic | Shared env | Nonshared | Genetic | Shared env | Nonshared | Genetic | Shared env | Nonshared | |
| WID | 0.35 (0.22, 0.51)* | 0.62 (0.46, 0.74)* | 0.03 (0.01, 0.06)* | 0.18 (0.06, 0.37)* | 0.82 (0.63, 0.94)* | 0.0005 (0.00, 0.02) | 0.18 (0.04, 0.42)* | 0.82 (0.57, 0.96)* | 0.00 (0.00, 0.02) |
| WATT | 0.37 (0.16, 0.61)* | 0.57 (0.34, 0.76)* | 0.05 (0.01, 0.11)* | 0.51 (0.17, 0.93)* | 0.48 (0.07, 0.81)* | 0.02 (0.00, 0.08) | 0.45 (0.08, 0.97)* | 0.52 (0.01, 0.89)* | 0.03 (0.00, 0.18) |
| RN | 0.55 (0.17, 0.79)* | 0.2 (0.001, 0.43)* | 0.24 (0.17, 0.34)* | 0.41 (0.08, 0.73)* | 0.33 (0.05, 0.59)* | 0.25 (0.15, 0.38)* | 0.38 (0.02, 0.71)* | 0.36 (0.07, 0.63)* | 0.25 (0.14, 0.40)* |
| Comp | 0.38 (0.18, 0.61)* | 0.55 (0.32, 0.74)* | 0.06 (0.02, 0.11)* | 0.24 (0.00, 0.58) | 0.68 (0.36, 0.91)* | 0.07 (0.01, 0.18)* | 0.31 (0.00, 0.73) | 0.58 (0.22, 0.94)* | 0.06 (0.001, 0.23)* |
Note. WID = Woodcock Reading Mastery Test (WRMT) Word Identification; WATT = WRMT Word Attack; Comp = WRMT Reading Comprehension; RN = Comprehensive Test of Phonological Processing, Rapid Naming.
p < .05.
Prior to examining the overlap in genetic and environmental influences on the intercept and slopes, we first acknowledge that a larger proportion of estimated variance of the intercept (centered at Assessment 1) is attributable to shared environment than would be expected from previous studies. To investigate, we fit five additional growth models for Word Identification recentering the intercept to each assessment time point. The resulting proportions of variance on the intercept from the reanalysis are presented in Figure 3, with each bar representing the results of a different analysis. In Figure 3, the total height of the bars represents the total variance in the intercept at each assessment, which substantially decreases over time. Turning to the proportions of variance in Figure 3, the amount of estimated variance due to genetic factors (labeled A) does not substantively change at each assessment point. In contrast, the amount of variance due to shared environment (labeled C) is smaller at each assessment point. As a result, the proportion of total variance attributable to genetics (the heritability) increases at later assessments (from about 35% at the first assessment to about 95% by the sixth assessment). Therefore, the observed increase in the proportion of variance attributable to heritability at later waves is not due to the emergence of additional genetic effects, but due primarily to a decrease in total variance, driven by a decrease in the amount of variance accounted for by the shared environment.
Figure 3.
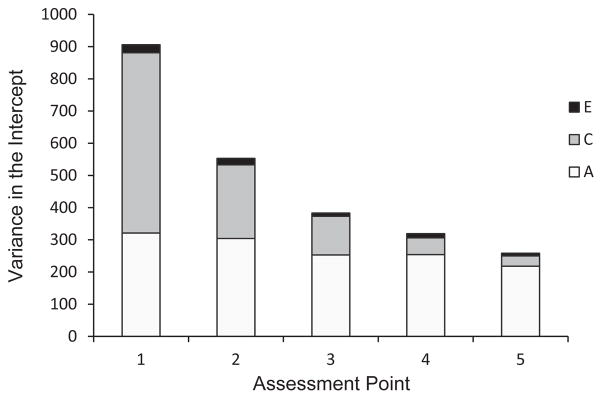
Results from recentering procedure. Bars represent genetic (A), shared environmental (C), and nonshared environmental (E) variance in the intercept when the model is centered at each of five assessment points.
Genetic and Environmental Contributions to the Overlap Between Intercept and Slope
It was also of interest to determine whether the genetic and environmental influences were unique to, or shared between, the intercept, linear slope, and quadratic parameters. Beginning with the genetic portions of the variance, the only significant overlap observed was between the intercepts and slopes of rapid naming (Table 4), suggesting that the genetic influences on children’s word-level reading, decoding, and reading comprehension at the first assessment point were unique from those related to how quickly the students’ grew in these skills, but that those influences did significantly overlap for rapid naming. There was some significant genetic overlap observed between the linear and quadratic slopes for Word Identification (−.17), Word Attack (−.47), and Rapid Naming (−.40). The direction of the covariance was negative, suggesting that the genetic influences related to faster growth were also associated with more deceleration of growth.
Table 4.
Estimates and Related Confidence Intervals (in Parentheses) of Genetic and Environmental Components of Covariance in the Latent Quadratic Growth Model Parameters
| Genetic
|
Shared env
|
Nonshared env
|
||||
|---|---|---|---|---|---|---|
| Intercept | Linear slope | Intercept | Linear slope | Intercept | Linear slope | |
| Word Identification | ||||||
| Slope | −0.01 (−0.14, 0.1) | — | −0.60 (−0.73, −0.46)* | — | −0.004 (−0.03, 0.01) | — |
| Quadratic | −0.08 (−0.22, 0.06) | −0.17 (−0.36, −0.02)* | 0.50 (0.32, 0.65)* | −0.81 (−0.93, −0.59)* | −0.003 (−0.02, 0.02) | 0.0004 (−0.02, 0.002) |
| Word Attack | ||||||
| Slope | −0.06 (−0.33, 0.16) | — | −0.43 (−0.65, −0.16)* | — | 0.02 (−0.03, 0.05) | — |
| Quadratic | 0.02 (−0.26, 0.35) | −0.47 (−0.94, −0.11)* | 0.36 (0.03, 0.63)* | −0.48 (−0.84, −0.02)* | −0.04 (−0.08, 0.02) | −0.01 (−0.12, 0.003) |
| Reading Comprehension | ||||||
| Slope | −0.2 (0.46, 0.01) | — | −0.61 (−0.80, −0.36)* | — | −0.06 (−0.03, −0.01)* | — |
| Quadratic | 0.21 (−0.05, 0.50) | −0.29 (−0.66, 0.07) | 0.56 (0.28, 0.80)* | −0.64 (−0.92, −0.29)* | 0.05 (0.01, 0.14) | −0.07 (−0.20, −0.01)* |
| Rapid Naming | ||||||
| Slope | −0.44 (−0.71, −0.14)* | — | −0.24 (−0.48, −0.01)* | — | −0.24 (−0.36, −0.16)* | — |
| Quadratic | 0.40 (0.10, 0.67)* | −0.40 (−0.72, −0.06)* | 0.25 (0.01, 0.48)* | −0.34 (−0.61, −0.06)* | 0.24 (0.16, 0.36)* | −0.25 (−0.39, −0.15)* |
Note. Quadratic = Quadriatic Slope; Word Identification = Woodcock Reading Mastery Test (WRMT) Word Identification; Word Attack = WRMT Word Attack; Reading Comprehension = WRMT Reading Comprehension; Rapid Naming = Comprehensive Test of Phonological Processing, Rapid Naming.
p < .05.
For the shared environment, there was significant shared variance observed between all three parameter estimates for each reading outcome. Just as with the phenotypic results the directions of these associations were negative between the intercept and linear slope, negative between the linear slope and quadratic slope, but positive between the intercept and quadratic slope. The significant overlap suggests that the shared environmental influences on children’s scores at the first assessment point were also important for how quickly they grew, as well as how much they decelerated. Finally, the significant nonshared environmental influences for Reading Comprehension and Rapid Naming suggested that the non-shared environmental influences on students’ skill in reading comprehension and rapid naming were also significantly related to how quickly the students’ grew in each of these areas.
Discussion
The first goal of this study was to identify the shape of growth in reading skills over 6 years of development, from the initiation of formalized instruction into early adolescence. As expected from previous work (Hill et al., 2007), the results of these models supported a positive general trend of growth across the six assessment periods, but the significant negative quadratic effect suggests that the growth rate slows as children get older. It is unknown if this changing growth pattern reflects our literacy education pattern itself, in that intensive literacy instruction drives the rapid growth, while the deceleration is temporally linked with when explicit reading instruction is no longer provided. Alternatively, the rapid growth followed by the deceleration could reflect a true maturation period in the development of reading, and that our education system is best served to intervene with younger readers to maximize the potential of more rapid growth. These possibilities can be informed somewhat through the decomposition of the individual differences variance into genetic and environmental components.
Turning to the relative importance of genetics on early reading skills and growth, the results demonstrated that significant proportions of variance in estimated reading scores at age 6 were attributable to genetic influences for all four examined outcomes (Word Identification, Word Attack, Rapid Naming, and Reading Comprehension). For the slopes, significant proportions of variance were attributable to genetic influences for most of the examined outcomes. The significant genetic influences on the slope of growth suggest that the growth rate identified in the phenotypic growth model and confirmed by the genetically sensitive growth model reflects, at least in part, a natural maturation process. We also found that for Word Identification, Word Attack, and Reading Comprehension, the genetic influences on the slope were unique from those on the intercept. This suggests that the genetic influences related to how quickly or slowly a student grows in their reading skill are not the same as the genetic influences on their skill in at the first assessment. In other words, some new genetic component related to growth is coming online after that first assessment wave, and it is influencing development. In addition, the finding that genetic influences on growth are unique from those on the intercept may indicate that different genetic factors may be important for students who demonstrate early risk for word-level reading disability than those students who are classified as having a reading disability because they do not make gains as expected in response to instruction (i.e., the RTI model; Al Otaiba & Fuchs, 2006; Torgesen, 2000).
This is in contrast to the rapid naming results, where some significant genetic overlap was observed between the intercept and slopes. This indicates that the same genetic factors were related to how quickly children performed the task at the first assessment point as were related to the rate of growth and deceleration across time. Recent work suggests that fluency may be genetically distinct from the domain being measured (Petrill et al., 2012). Thus, it is possible that the significant overlap between the intercept and slopes for rapid naming represents the common element of fluency (speed) of performance.
Similar to the overall genetic results, the shared environmental proportions of variance on the intercepts (when centered at Assessment 1) and slopes were also significant for all four examined outcomes. However, in contrast to the genetic results, the shared environmental overlap between the intercept and linear slope was significantly negative, and that between the intercept and quadratic slope was significantly positive. This suggests that the shared environmental influences on the initial assessment point are also important for how students grow in their reading skills throughout school (both growth rate and deceleration rate). Furthermore, the directions of these relations suggest that the environmental influences on high scores at initial assessment point were also significantly related to slower growth in reading throughout school, and faster deceleration of growth throughout school. This is not surprising given that students with higher reading scores are often found to grow more slowly in those skills than students with lower initial scores; students with lower initial scores have more room to grow (Skibbe et al., 2008). These findings highlight the importance of early environmental influences on shaping children’s reading performance in later years. Several phenotypic studies suggest that early environmental influences can be causally related to increased reading performance at later grades (e.g., shared book reading; Piasta, Justice, McGinty, & Kaderavek, 2012). These findings suggest that though concurrently measured shared environmental factors can be important, early environmental factors, such as the curriculum used in the kindergarten classroom, or home factors such as the presence of books in the home or parental literacy practices could be particularly salient for future reading performance.
However, these results must be balanced against another finding of the current study (and the larger behavioral genetic literature) that the influence of shared environment decreases over time. This finding may map on to the changing role of home and school environments during the examined developmental period (Figure 3). Specifically, at school entry, much of the variance in reading scores represents what has been occurring in the home (e.g., amount of shared book reading, visits to the library). This is likely more variable that what occurs in schools that often use a standardized curriculum to teach reading and reading related skills. As students enter school, they then receive instruction in word reading, decoding, letter recognition, and even reading comprehension. These environmental factors increase mean reading performance across all children. As such, entry into school represents a decrease in the variability in the shared environment, and therefore less total attribution of shared environment to the total variance in reading (as shown in Figure 2).
Limitations
There are several limitations to consider when interpreting these findings. First, although the confidence intervals of the parameter estimates indicate significance from zero, most are large and make statistical comparisons between estimates difficult. Relatedly, the large confidence intervals observed may indicate that some nonsignificant estimates (e.g., the genetic contribution to growth in Reading Comprehension) could be found to be significant given a larger sample size. Second, three of the assessments, Word Attack, Word Identification, and Reading Comprehension, demonstrated floor effects. In other words, some students scored very poorly on these measures at the first assessment point. Because floor effects make mono- and dizygotic twins more similar to one another, they can result in a trait appearing to have an overly large contribution to the shared environment. These effects are unfortunately inherent in the assessment of a skill that is just beginning to come online and would be difficult to tease apart in future studies.
Although the model indicated that there were significant environmental influences on children’s reading status and growth, the model used did not attempt to identify them. Future work should seek to identify these environmental influences, and how they interact with genetic factors. For example, future studies could directly measure the amount of shared book reading that occurs in children’s homes. It is likely that those students with more shared book reading would have stronger intercepts (read better) and/or slopes (grow faster) than those students with less book reading. Direct measurement of the environment would enhance our understanding of which environmental attributions influence growth in these skills over time.
It should be noted that the significant shared environmental variance attributable to word identification and word attack at the intercept was larger in magnitude than other findings reported in the extant literature. For example, Harlaar, Spinath, Dale, and Plomin (2005) reported a shared environmental estimate of about 20% for their sample of 7-year-old students, and a similar proportion of variance was reported by Taylor and Schatschneider (2010) in a large sample of kindergarten and first-grade students in Florida. The International Longitudinal Survey of Twins (Samuelsson et al., 2008) found even smaller shared environmental estimates, most indicating less than 10% of the variance in reading skill was due to the shared environment. The larger attribution to the shared environment in the estimates observed in this study could be due to the relatively wide educational range of participants (the first assessment point contains students in entering kindergarten to first grade). This could also be related to the comparatively strong environmental attribution found on the slopes found in Petrill et al. (2010). The results of this study demonstrated that the quadratic model was a more accurate representation of the true form of growth. Such an improvement will result in parameter estimates that are more accurate. Thus, the significant genetic and shared environmental influences observed for the growth parameters in this study are likely a more accurate representation of these influences than those reported in Petrill et al.
This is further evidenced by an examination of the univariate estimates at each individual time point. At each wave of data collection, the attribution to the shared environment declines, while the proportion due to heritability increases (Table 1). It should be noted, however, that we included age as a definition variable in the current study, which is highly correlated (r = .88) with months of schooling at Assessment 1 (Petrill, Deater-Deckard, Thompson, DeThorne, & Schatschneider, 2006a).
Finally, it is very important to highlight that although our results indicate that 35% of the variability in word identification is attributable to genetics, that does not mean that any one person has an initial reading score that is 35% predetermined by genetics. Rather, it indicates that 35% of the variability in word identification scores in the present sample could be accounted for by the extent to which MZ twins were more similar than DZ twins. These statistics are reliant on the sample from which they were drawn, and particularly on the environmental variability to which the sample has been exposed. Replications of the current study are needed with large sample sizes, perhaps those selected for particular reading levels, and wide ranges of environmental variance to best determine the proportions of variance attributable to genes and environments in the population.
Conclusions and Future Directions
Research in the area of reading development consistently finds that reading skills show rapid change during the first few years of development, but then that change slows considerably (Hill et al., 2007). Variability in reading and growth in reading have been shown to be significantly related to environmental factors (Torgesen et al., 1999). At the same time, results from the extant behavior genetics literature have suggested that the majority of the variance in reading is influenced by stable genetic effects. One possible reason for this seeming disconnect in the literature is that the behavior genetics literature on reading development has been based primarily on static time points or other basic methods of examining longitudinal change over time. For example, behavioral genetics studies have compared genetic and environmental contributions at multiple static time points, used autoregressive twin models (Byrne et al., 2007; Samuelsson et al., 2008), or cross-lagged twin models (Ebejer et al., 2010; Harlaar et al., 2007) to examine development of reading and its component skills. In other words, the conclusion that reading development is primarily influenced by genetics has been based on studies that do not directly model individual student growth.
In this study, we combined these two methodological approaches. In so doing, we found evidence consistent with both the phenotypic and quantitative genetic bodies of literature. Our results suggested that, consistent with the behavior genetics literature, the primary influence on the static assessment points (intercepts) was found to be genetic, particularly at later ages. At the same time, growth in reading skills was found to be primarily driven by the shared environment, as would be expected based on the extant phenotypic literature on reading development. Through demonstrating this, we hope to enhance the general knowledge and understanding of individual differences in reading growth, and to further elucidate how phenotypic and behavioral genetic literatures complement each other.
Finally, the results of the current study have important implications for the timing of how we may intervene in reading education. We found that the shared environmental contributions on the slope were not unique to the slope, but overlapped significantly with the variance at school entry. This confirms the findings in the phenotypic reading literature suggesting that indices of the home environment are salient in early as opposed to later reading. In addition, although significant genetic variance was found in the slopes, it was unique from that at the static time points. Thus, children come to school with different experiences, but also with varying genetic risk and protective factors for reading.
Untangling these contributions will involve greater integration of quantitative genetic, molecular genetics (Hannula-Jouppi et al., 2005; Taipale et al., 2003), and educational neuroscience. In a recent review of the literature, Poelmans, Buitelaar, Pauls, and Franke (2011), suggested that 10 of the 14 most commonly cited dyslexia candidate genes influence two developmental processes: neuronal migration (where neurons move) and neurite outgrowth (how the dendrites and axons of neurons grow from the cell body). Because molecular genetic studies typically use static (sometimes singular) assessments of reading, the developmental timing of these candidate genes is poorly understood. Given the results of our study, differences in the developmental course of reading may extend beyond differences in instruction, or differences in preexisting genetic differences that are laid down prior to school, but may also be due to differences in genetic contributions to rate of reading growth in school. If this is the case, then a subset of the candidate genes related to neuronal migration and neurite outgrowth (or other processes) may affect differences in rates of growth (the slope) whereas other candidate genes may be related to the intercept.
In addition, neuroimaging research has found that neurobiological correlates of word recognition in reading disabled groups show perisylvian region dysfunction, including under activation of left temporoparietal and occipitotemporal regions and greater right temporoparietal and occipitotemporal activation as compared to typically developing readers (Pugh et al., 2000; Richlan, Kronbichler, & Wimmer, 2009; Rumsey, Nace, Donohue, & Wise, 1997). Importantly, these differences appear even before formalized reading instruction begins (Raschle, Chang, & Gabb, 2011). It is possible that individual differences in these brain regions may explain a portion of the heritability as well as the shared environmental variance of reading outcomes. Differences in these brain regions may also be partially explained by candidate genes related to reading outcomes. Examining the longitudinal interrelations among brain structure and function, molecular genetics, and quantitative genetics across the developmental course of reading growth would help address the larger questions of how and why reading disabilities develop, and how they can be prevented and treated in light of the substantial naturally occurring genetic and environmental variance that children bring with them to school and experience in school. Thus, the findings of this study suggest that there may be important, currently untapped, avenues for identifying risk, not only for poor initial reading, but also poor reading growth, beyond quality of the home environment and instruction.
Acknowledgments
This research was supported by the National Institute of Child Health and Human Development (NICHD) Grants HD38075 and HD68728 to The Ohio State University. The content of this publication does not necessarily reflect the views or policies of the NICHD, and mention of trade names, commercial products, or organizations does not imply endorsement by the United States government.
Contributor Information
Jessica A. R. Logan, The Ohio State University
Sara A. Hart, The Florida State University
Laurie Cutting, Vanderbilt University.
Kirby Deater-Deckard, Virginia Tech.
Chris Schatschneider, The Florida State University.
Stephen Petrill, The Ohio State University.
References
- Al Otaiba S, Fuchs D. Who are the young children for whom best practices in reading are ineffective? An experimental and longitudinal study. Journal of Learning Disabilities. 2006;39:414–431. doi: 10.1177/00222194060390050401. [DOI] [PubMed] [Google Scholar]
- Betjemann RS, Willcutt EG, Olson RK, Keenan JM, DeFries JC, Wadsworth SJ. Word reading and reading comprehension: Stability, overlap, and independence. Reading and Writing. 2008;21:539–558. [Google Scholar]
- Butler SR, Marsh HW, Sheppard MJ, Sheppard JL. Seven-year longitudinal study of the early prediction of reading achievement. Journal of Educational Psychology. 1985;77:349–361. [Google Scholar]
- Byrne B, Samuelsson S, Wadsworth S, Hulslander J, Corley R, DeFries JC, et al. Longitudinal twin study of early literacy development: Preschool through Grade 1. Reading and Writing. 2007;20:77–102. [Google Scholar]
- Byrne B, Wadsworth S, Corley R, Samuelsson S, Quain P, DeFries JC, et al. Longitudinal twin study of early literacy development: Preschool and kindergarten phases. Special Issue: Genes, Environment, and the Development of Reading Skills. 2005;9:219–235. [Google Scholar]
- Chall JS. Stages of reading development. New York: McGraw-Hill; 1983. [Google Scholar]
- Connor CMD, Piasta SB, Fishman B, Glasney S, Schatschneider C, Crowe E, et al. Individualizing student instruction precisely: Effects of child by instruction interactions on first graders’ literacy development. Child Development. 2009;80:77–100. doi: 10.1111/j.1467-8624.2008.01247.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham AE, Stanovich KE. Early reading acquisition and its relation to reading experience and ability 10 years later. Developmental Psychology. 1997;33:934–945. doi: 10.1037//0012-1649.33.6.934. [DOI] [PubMed] [Google Scholar]
- Ebejer JL, Coventry WL, Byrne B, Willcutt EG, Olson RK, Corley R, et al. Genetic and environmental influences on inattention, hyperactivity-impulsivity, and reading: Kindergarten to Grade 2. Scientific Studies of Reading. 2010;14:293–316. doi: 10.1080/10888430903150642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foorman BR, Francis DJ, Fletcher J, Schatschneider C, Mehta P. The role of instruction in learning to read: Preventing reading failure in at-risk children. Journal of Educational Psychology. 1998;90:37–55. [Google Scholar]
- Hannula-Jouppi K, Kaminen-Ahola N, Taipale N, Eklund R, Nopola-Hemmi J, Kaariainen H, et al. The axon guidance receptor gene ROBO1 is a candidate gene for developmental dyslexia. PLoS Genetics. 2005;1:467–474. doi: 10.1371/journal.pgen.0010050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harlaar N, Dale P, Plomin R. Reading exposure: A (largely) environmental risk factor with environmentally-mediated effects on reading performance in the primary school years. Journal of Child Psychology and Psychiatry. 2007;48:1192–1199. doi: 10.1111/j.1469-7610.2007.01798.x. [DOI] [PubMed] [Google Scholar]
- Harlaar N, Spinath FM, Dale PS, Plomin R. Genetic influences on early word recognition abilities and disabilities: A study of 7-year-old twins. Journal of Child Psychology and Psychiatry. 2005;46:373–384. doi: 10.1111/j.1469-7610.2004.00358.x. [DOI] [PubMed] [Google Scholar]
- Hill CJ, Bloom HS, Black AR, Lipsey MW. Empirical benchmarks for interpreting effect sizes in research. New York: MDRC; 2007. [Google Scholar]
- Lyon GR. Reading development, reading difficulties, and reading instruction educational and public health issues. Journal of School Psychology. 2002;40:3–6. [Google Scholar]
- McCoach DB, O’Connell AA, Reis SM, Levitt HA. Growing readers: A hierarchical linear model of children’s reading growth during the first 2 years of school. Journal of Educational Psychology. 2006;98:14–28. [Google Scholar]
- Morgan PL, Farkas G, Tuffs PA, Sperling RA. Are reading and behavior problems risk factors for each other? Journal of Learning Disabilities. 2008;41:417–436. doi: 10.1177/0022219408321123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muthén LK, Muthén B. Mplus user’s guide. 3. Los Angeles: Muthén & Muthén; 1998–2004. [Google Scholar]
- National Reading Panel. National Reading Panel report: Teaching children to read: An evidence-based assessment of the scientific research literature on reading and its implications for reading instruction (NIH Pub No 00-4769) Washington, DC: Author; 2000. [Google Scholar]
- Neale MC, Boker SM, Xie G, Maes HH. Mx: Statistical modeling. 7. Richmon, VA: Department of Psychiatry; 2006. [Google Scholar]
- Nye B, Konstantopoulos S, Hedges LV. How large are teacher effects? Educational Evaluation and Policy Analysis. 2004;26:237–257. [Google Scholar]
- Parrila R, Aunola K, Leskinen E, Nurmi JE, Kirby JR. Development of individual differences in reading: Results from longitudinal studies in English and Finnish. Journal of Educational Psychology. 2005;97:299–319. [Google Scholar]
- Petrill SA, Deater-Deckard K, Thompson LA, DeThorne LS, Schatschneider C. Genetic and environmental effects of serial naming and phonological awareness on early reading outcomes. Journal of Educational Psychology. 2006a;98:112–121. doi: 10.1037/0022-0663.98.1.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrill SA, Deater-Deckard K, Thompson LA, DeThorne LS, Schatschneider C. Reading skills in early readers: Genetic and shared environmental influences. Journal of Learning Disabilities. 2006b;39:48–55. doi: 10.1177/00222194060390010501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrill SA, Deater-Deckard K, Thompson LA, Schatschneider C, DeThorne LS, Vandenbergh DJ. Longitudinal genetic analysis of early reading: The Western Reserve Reading Project. Reading and Writing. 2007;20:127–146. doi: 10.1007/s11145-006-9021-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrill SA, Hart SA, Harlaar N, Logan J, Justice LM, Schatschneider C, et al. Genetic and environmental influences on the growth of early reading skills. Journal of Child Psychology and Psychiatry. 2010;51:660–667. doi: 10.1111/j.1469-7610.2009.02204.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrill SA, Logan JAR, Hart SA, Vincent P, Thompson L, Kovas Y, et al. Math fluency is etiologically distinct from untimed math performance, decoding fluency, and untimed reading performance: Evidence from a twin study. Journal of Learning Disabilities. 2012;45:371–381. doi: 10.1177/0022219411407926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phillips BM, Lonigan CJ. Social Correlates of Emergent Literacy. In: Snowling MJ, Hulme C, editors. The science of reading: A handbook. Oxford, England: Blackwell; 2005. pp. 173–187. [Google Scholar]
- Piasta SB, Justice LM, McGinty AS, Kaderavek JN. Increasing young children’s contact with print during shared reading: Longitudinal effects on literacy achievement. Child Development. 2012;83:810–820. doi: 10.1111/j.1467-8624.2012.01754.x. [DOI] [PubMed] [Google Scholar]
- Poelmans G, Buitelaar JK, Pauls DL, Franke B. A theoretical molecular network for dyslexia: Integrating available genetic findings. Molecular Psychiatry. 2011;16:365–382. doi: 10.1038/mp.2010.105. [DOI] [PubMed] [Google Scholar]
- Pugh KR, et al. Functional neuroimaging studies of reading and reading disability. Mental Retardation and Developmental Disabilities Research Reviews. 2000;6:207–213. doi: 10.1002/1098-2779(2000)6:3<207::AID-MRDD8>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- Raschle NM, Chang M, Gabb N. Structural brain alterations associated with dyslexia predate reading onset. Neuroimage. 2011;57:742–749. doi: 10.1016/j.neuroimage.2010.09.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raudenbush SW, Bryk AS. Hierarchical linear models: Applications and data analysis methods. 3. Thousand Oaks, CA: Sage; 2002. [Google Scholar]
- Reardon SF, Robinson J. Patterns and trends in racial/ethnic and socioeconomic academic achievement gaps. Upper Saddle River, NJ: Prentice Hall; 2007. [Google Scholar]
- Reynolds CA, Finkle D, McCardle JJ, Gatz M, Berg S, Pederson NL. Quantitative genetic analysis of latent growth curve models of cognitive abilities in adulthood. Developmental Psychology. 2005;41:3–16. doi: 10.1037/0012-1649.41.1.3. [DOI] [PubMed] [Google Scholar]
- Richlan F, Kronbichler M, Wimmer H. Functional abnormalities in the dyslexic brain: A quantitative meta-analysis of neuroimaging studies. Human Brain Mapping. 2009;30:3299–3308. doi: 10.1002/hbm.20752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rumsey JM, Nace K, Donohue B, Wise D. A positron emission tomographic study of impaired word recognition and phonological processing in dyslexic men. Archives of Neurology. 1997;54:562–574. doi: 10.1001/archneur.1997.00550170042013. [DOI] [PubMed] [Google Scholar]
- Samuelsson S, Byrne B, Olson RK, Hulslander J, Wadsworth S, Corley R, et al. Response to early literacy instruction in the United States, Australia, and Scandinavia: A behavior-genetic analysis. Learning and Individual Differences. 2008;18:289–295. doi: 10.1016/j.lindif.2008.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scanlon DM, Vellutino FR. Prerequisite skills, early instruction, and success in first-grade reading: Selected results from a longitudinal study. Mental Retardation and Developmental Disabilities Research Reviews. 1996;2:54–63. [Google Scholar]
- Schatschneider C, Fletcher JM, Francis DJ, Carlson CD, Foorman BR. Kindergarten prediction of reading skills: A longitudinal comparative analysis. Journal of Educational Psychology. 2004;96:265–282. [Google Scholar]
- Skibbe LE, Grimm KJ, Stanton-Chapman TL, Justice LM, Pence KL, Bowles RP. Reading trajectories of children with language difficulties from preschool through fifth grade. Language, Speech, and Hearing Services in Schools. 2008;39:475–486. doi: 10.1044/0161-1461(2008/07-0016). [DOI] [PubMed] [Google Scholar]
- Snow CE, Burns MS, Griffin P. Preventing reading difficulties in young children. In Reading research: Anthology: The why? of reading instruction. Novata, CA: Academic Therapy; 1999. pp. 148–155. [Google Scholar]
- Stevenson J, Graham P, Fredman G, McLoughlin V. A twin study of genetic influences on reading and spelling ability and disability. Journal of Child Psychology and Psychiatry. 1987;28:229–247. doi: 10.1111/j.1469-7610.1987.tb00207.x. [DOI] [PubMed] [Google Scholar]
- Taipale M, Kaminen N, Nopola-Hemmi J, Haltia T, Myllyluoma B, Lyytinen H, et al. A candidate gene for developmental dyslexia encodes a nuclear tetratricopeptide repeat domain protein dynamically regulated in brain. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:11553–11558. doi: 10.1073/pnas.1833911100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor J, Schatschneider C. Genetic influence on literacy constructs in kindergarten and first grade: Evidence from a diverse twin sample. Behavior Genetics. 2010;40:591–602. doi: 10.1007/s10519-010-9368-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torgesen JK. Individual differences in response to early interventions in reading: The lingering problem of treatment resisters. Learning Disabilities Research and Practice. 2000;15:55–64. [Google Scholar]
- Torgesen JK, Wagner RK, Rashotte CA, Burgress S, Hecht S. Contribution of phonological awareness and rapid automatic naming ability to the growth of word reading skills in second-to-fifth-grade children. Scientific Studies of Reading. 1997;1:161–185. [Google Scholar]
- Torgesen J, Wagner R, Rashotte C, Rose E, Lindamood P, Conway T, et al. Preventing reading failure in young children with phonological processing disabilities: Group and individual responses to instruction. Journal of Educational Psychology. 1999;91:579–593. [Google Scholar]
- Wadsworth SJ, DeFries JC, Olson RK, Willcutt EG. Colorado longitudinal twin study of reading disability. Annals of Dyslexia. 2007;57:139–160. doi: 10.1007/s11881-007-0009-7. [DOI] [PubMed] [Google Scholar]
- Wagner RK, Torgesen JK. The nature of phonological processing and its causal role in the acquisition of reading skills. Psychological Bulletin. 1987;101:192–212. [Google Scholar]
- Wagner RK, Torgesen JK, Rashotte CA. Comprehensive Test of Phonological Processing. Austin, TX: PRO-ED; 1999. [Google Scholar]
- Woodcock RW. Woodcock Reading Mastery Tests–Revised. Circle Pines, MN: American Guidance Service; 1987. [Google Scholar]