

Abstract
Cocaine generates drug-seeking behavior by creating long-lasting changes in the reward pathway. The role of the growth factor, brain-derived neurotrophic factor (BDNF) in facilitating these changes was investigated in the present report with a genetic rat model. Using conditioned place preference, the current study investigated the hypothesis that a partial knockout of the BDNF gene in rats (BDNF+/−) would attenuate the rewarding effects of cocaine. Wildtype rats exposed to cocaine exhibited normal cocaine-seeking responses one day after conditioning and cocaine-seeking behavior was reinstated with drug priming following drug abstinence. In contrast, BDNF+/− rats did not show cocaine-seeking behavior one day after conditioning, nor did they respond to drug priming. A median split of rats based on BDNF levels in sera collected prior to behavioral procedures revealed that wildtype rats with high BDNF levels showed stronger conditioned place preference and reinstatement to cocaine. Together, the results support the hypothesis that a partial knockout of the BDNF gene attenuates the rewarding properties of cocaine. Additionally, individual differences in BDNF levels may predict future cocaine-seeking behavior. An underlying mechanism of these effects may be a reduction of the amount of synaptic changes made in the reward pathway.
Keywords: brain-derived neurotrophic factor, knockout rats, conditioned place preference, addiction, reinstatement, extinction
1. Introduction
Cocaine dependent individuals persistently perform cocaine-seeking behavior despite negative consequences [21] and relapse is a tremendous challenge in treating addiction [6]. Abstinent human addicts presented with stimuli previously paired with drug use report increased drug craving [10]. Rats, like humans, exhibit addictive phenotypes, performing cocaine-seeking behaviors in the presence of aversive conditioned stimuli [21] and vulnerability to relapse after extinction of cocaine-seeking behavior [7]. For example, a priming injection of cocaine reinstates cocaine-seeking behavior in rats that have undergone extinction [7,15]. The occurrence of relapse, even after long periods of abstinence, is made possible by long-lasting changes in the reward pathway and the persistence of learned associations [11]. Drug-induced neuroplasticity in areas of the reward pathway, such as the ventral tegmental area (VTA) and nucleus accumbens (NA), as well as in the prefrontal cortex (PFC), amygdala, and hippocampus, generate drug-seeking behavior [16] and are mediated by growth factors [16,19].
Accumulating evidence suggests an integral role for the growth factor, brain-derived neurotrophic factor (BDNF) in cocaine addiction [18]. For example, a single infusion of BDNF into the VTA increased cocaine-seeking [13] and intra-PFC BDNF infusions regulate glutamate activity in NA after cocaine self-administration leading to decreased drug-induced reinstatement [3]. BDNF also improved dopamine neurotransmission in the VTA-NA pathway in the development of locomotor sensitization to cocaine [1] and BDNF injections slowed the extinction of drug-seeking behavior during drug abstinence and resulted in stronger tendencies to relapse [18]. Genetically modified mice heterozygous for the BDNF gene (BDNF+/−) had less cocaine-induced locomotion and reduced conditioned place preference (CPP) to cocaine, suggesting a reduction in sensitivity to the rewarding effects of cocaine [9].
In the present study, we evaluated the effects of a mono-allelic deletion of the BDNF gene on the rewarding effects of cocaine in a rat CPP model. We expected BDNF+/− rats to exhibit less cocaine-seeking behavior than wildtype rats and to exhibit a reduction in relapse susceptibility after an 8-day period of abstinence. Additionally, blood samples taken before behavioral procedures began were used to examine the relationship between serum BDNF levels and cocaine-seeking behavior.
2. Materials and Methods
2.1 Animals and colony conditions
The subjects were adult female Sprague-Dawley rats (N=40), bred on-site at Colby College by mating heterozygous male studs with wildtype females obtained from Sigma Advanced Genetic Engineering Labs (Sage®; Sigma-Aldrich Co., St. Louis, MO). Only 1–2 females from a litter were used. Rats were weaned into pairs on postnatal day 24 and housed in individually-ventilated clear polycarbonate cages (30.8 × 30.8 × 18.7 cm; Thoren Caging Systems, Hazleton, PA) with ad libitum access to food and water. The colony was maintained on a 12 h light/dark cycle with lights on at 7:00; the colony temperature was 22±2 °C with 40–60% humidity. All procedures were carried out during the rats’ light phase and were reviewed and approved by the Institutional Animal Care and Use Committee of Colby College.
2.2 Genotyping and Sera BDNF determination
One week before the onset of behavioral procedures, 2–3 mm samples of tissue from the tip of the tail were collected, stored in 70% alcohol at room temperature, and shipped overnight to Sage Laboratories for BDNF genotyping. Samples were analyzed by standard PCR, treated with Exo-SAP, and sent directly to sequencing. Based on the genotyping, 18 rats were heterozygous for the BDNF gene (BDNF+/−) and 22 were wildtype (WT). At the time of tissue collection, approximately 200 μl of tail blood was collected from each rat and extracted sera was stored at −80 °C until assayed for BDNF levels in duplicate under manufacturer’s instructions using a BDNF Sandwich ELISA kit (ChemiKine, cat. no. CYT306). It should also be noted that WT rats weighed significantly less than BDNF+/− rats at the study’s start (WT: M=239.5 g, SD=35.0, BDNF+/−: 277.3 g, SD=35.4; t[38]=−3.376, p=0.002) and end (WT: 255.4 g, SD=34.35, BDNF+/−: 300.0 g, SD=24.5; t[38]=−4.619, p=0.001).
2.3 Conditioned place preference apparatus
There were 4 identical CPP apparatuses constructed of painted wood (58.5 × 28.5 × 26 cm) with a clear Plexiglas lid that allowed for video recording and software tracking (ANY-maze, Stoelting Co., Wood Dale, IL) via a HD digital video recorder (JVC Everio, Model No. GZ-MG620) mounted directly overhead. Each apparatus was separated into two equal chambers with a removable divider. The two chambers were visually and tactically distinct.
2.4 Experimental procedures
The CPP procedures are shown in Figure 1 and described below. There were four stages occurring over a 14-day period: habituation; conditioning; extinction; and priming. Rats were handled regularly by the experimenters during the weeks before CPP and weighed daily during testing to ensure accurate dosing of drugs on conditioning days and maintain consistent experiences during each stage of CPP. For all stages, rats spent 15 minutes (or 900 seconds) in the apparatus.
Figure 1.
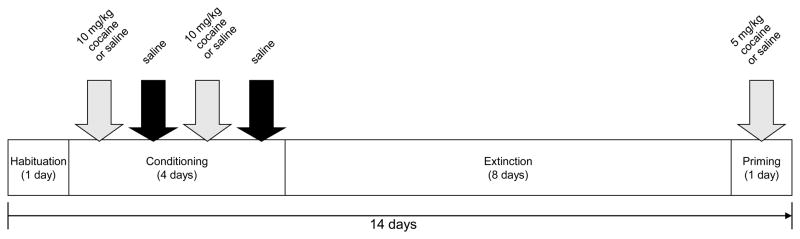
Experimental timeline of CPP procedures.
2.4.1 Habituation
All rats received one day of habituation to the apparatus. Each rat was placed into the apparatus with the divider removed and allowed to freely explore both chambers. Two rats showed a strong preference for one chamber over the other during habituation (>700 of the 900 seconds spent in it) and were later conditioned in their non-preferred chamber (one WT-cocaine; one BDNF+/− saline). Otherwise, rats were randomly assigned to a conditioning chamber (one of the two sides of the apparatus) as well as drug condition (saline or cocaine), and the combination of chamber-drug pairings were counterbalanced within and between groups.
2.4.2 Conditioning trials
On the day following habituation, conditioning began and continued for 4 consecutive days. On all conditioning trials, rats in the saline groups were injected with 0.9% saline (i.p.) and placed immediately into the apparatus. On days 1 and 3, they were confined to one chamber of the apparatus with the divider wall present; on days 2 and 4 they were confined to the opposite chamber. On days 1 and 3, rats in the cocaine groups were injected with cocaine hydrochloride (10 mg/kg, i.p.; from Sigma-Aldrich Co.) and placed in their assigned chamber. On days 2 and 4, these rats received an i.p. injection of saline and were placed in the opposite chamber. These procedures are well-established in the field [7] and ensure equal exposure to both chambers of the apparatus while, for cocaine-treated rats, only one chamber is paired with drug (Paired) and the other side with saline (Unpaired). Note that these terms were also used for saline-treated rats though it was arbitrary since they had the same experience in both chambers. The study design resulted in 4 experimental conditions: WT/saline (n=10); BDNF+/−/saline (n=8); WT/cocaine (n=12); and BDNF+/−/cocaine (n=10).
2.4.3 Extinction trials
On the day after conditioning day 4, extinction trials began and lasted for 8 consecutive days. On each extinction trial, rats were placed in the center of the apparatus with the center divider removed. The time rats spent in each chamber was measured. The chamber times on extinction day 1 and 8 were of particular interest: chamber times on extinction day 1 were used to assess the extent to which cocaine-treated rats preferred the chamber paired with cocaine over the chamber not paired with cocaine, whereas chamber times on extinction day 8 were used to assess the extent to which any conditioned place preferences in cocaine-treated rats had dissipated as a result of the 7 previous extinction trials. It was expected that, overall, rats given saline on all conditioning trials would not show any chamber preferences on either day.
2.4.4 Priming
On the day after extinction day 8, relapse susceptibility was assessed by giving a priming dose of cocaine (5 mg/kg; i.p.) to those rats previously conditioned with it; rats in the saline conditions received a saline injection. As on extinction days, the center divider was removed and rats explored the apparatus freely. The times in each chamber were again recorded and expected to reveal the extent to which the priming dose of cocaine led to a reinstatement of the previously extinguished CPP.
3. Results
3.1 Extinction
For extinction day 1, A 2×2×2 mixed factorial ANOVA conducted on chamber times with the factors Genotype, Drug, and Chamber revealed a significant main effect of Chamber (F[1,35]=4.749, p=0.035 and a significant interaction of Chamber and Drug (F[1,35]=8.663, p=0.006), but no other significant effects. Planned, paired t-tests, 2-tailed, were used to analyze chamber preferences on extinction day 1, and revealed that only WT cocaine-treated rats spent significantly more time in the paired chamber than the unpaired chamber (t[10]=3.623, p=0.005; see Fig. 2A). BDNF+/− cocaine-treated rats also spent more time in the paired than unpaired chamber but this difference was not statistically significant (t[8]=1.952, p=0.087). As expected, the difference in time spent in each chamber of the apparatus was not statistically significant for WT or BDNF+/− rats treated only with saline. There were no significant differences in overall distance traveled in the apparatus during this session (WT-Saline M=15.74, SE=2.37; BDNF+/− Saline M=14.99, SE=2.09; WT-Cocaine M=15.97; SE=2.99; BDNF+/− Cocaine M=14.81, SE=1.99). For extinction day 8, the ANOVA did not detect any significant effects and planned comparisons confirmed that the WT-cocaine-treated rats no longer showed a significant preference for the cocaine-paired chamber (t[10]=1.216, p=0.252; see Fig. 2B); the other 3 groups continued to not show a significant preference for the paired chamber.
Figure 2.

WT, but not BDNF+/−, rats given cocaine during conditioning spent significantly more time in the chamber aired with the drug than in the unpaired chamber on extinction day 1 (A) and priming day (C). On extinction day 8, no groups favored the paired chamber (B). Error bars are ± SEM. * p < 0.05
3.2 Priming
A 2×2×2 ANOVA conducted on chamber times after the priming injection of cocaine showed a significant interaction between Chamber and Drug (F[1,35]=6.767, p=0.014) but no other significant effects. Planned comparisons showed that cocaine priming in WT and BDNF+/− rats led to reinstatement of CPP in WT (t[10]=4.187, p=0.002), but not BDNF+/− rats (t[8]=1.212, p=0.260; see Figure 2C). As expected, giving saline to WT and BDNF+/− rats previously given only saline during conditioning did not produce a place preference on the priming day.
3.3 CPP as a function of pre-conditioning BDNF levels
A 2×2 ANOVA conducted on pre-conditioning BDNF levels in sera revealed a significant main effect of Genotype (F[1,35]=32.62, p<0.001). As expected, BDNF+/− rats had significantly lower BDNF levels than WT rats (see Fig. 3A). Neither the main effect of Drug nor the interaction between Genotype and Drug were statistically significant indicating that there were no pre-existing differences amongst the groups that went into the cocaine and saline conditions. To investigate if initial BDNF levels predicted CPP outcomes, we performed a median split in the WT and BDNF+/− cocaine-treated groups (see Fig. 3B) and reassessed preferences for the cocaine-paired chamber on extinction day 1 and priming day. Rats below the median in each group were classified as “low BDNF” (WT range: 88.4–131.3 pg/mL, BDNF+/− range: 64.1–80.3 pg/mL) and rats above the median were classified as “high BDNF” (WT range: 132.1–264.4 pg/mL, BDNF+/− range: 91.2–102.2 pg/mL). A 2×2 ANOVA conducted with the factors of Genotype and BDNF category (“high BDNF” or “low BDNF”) on BDNF levels revealed significant main effects of Genotype (F[1,16]=22.559, p<0.001) and Category (F[1,16]=11.672, p=0.004) but a non-significant interaction. As shown in Fig. 3B, there were significantly lower levels of BDNF in rats categorized as low BDNF for both WT and BDNF+/− rats (t[9]=−2.910, p=0.017 and t[7]=−5.521, p=0.001, respectively, Bonferroni corrected).
Figure 3.

Serum BDNF levels were significantly lower in BDNF+/− rats compared to WT and there were no pre-existing differences among the drug conditions (A). A median split into low and high BDNF groups for cocaine-treated WT and BDNF+/− groups revealed significant differences between those categorized into “low” and “high” BDNF levels (B). Only high BDNF WT rats displayed a significant preference for the cocaine-paired chamber on extinction day 1 (C) and showed a significantly larger increase in time spent in the paired chamber from extinction day 8 to priming (D). Error bars are ± SEM. * p < 0.05
Using these BDNF categories the results from extinction day 1 and priming day were reanalyzed for those rats given cocaine conditioning trials. A 2×2×2 ANOVA with factors of Genotype, BDNF Category, and Chamber conducted on chamber times revealed significant main effects of Chamber on both days (F[1,16]=13.026, p=0.002 and F[1,16]=9.702, p=0.007, respectively) but no other significant effects or interactions. Planned comparisons showed that both high and low BDNF WT rats spent more time in the cocaine-paired chamber than in the unpaired chamber, but the difference was only statistically significant for the WT rats with high BDNF levels (t[5]=2.981, p=0.031; see Fig. 3C) and not for the low BDNF WT rats (t[4]=2.215, p=0.091). Among the BDNF+/− rats, neither low (t[4]=1.338, p=0.252) nor high (t[3]=1.264, p=0.295) BDNF rats spent significantly more time in the cocaine-paired chamber on extinction day 1 (see Fig. 3C). On priming day, both low (t[4]=2.613, p=0.059) and high (t[5]=2.317, p=0.068) BDNF WT rats spent more time in the cocaine-paired chamber than the saline-paired chamber, but neither was statistically significant (data not shown). Similarly, on priming day, neither high nor low BDNF BDNF+/− rats given cocaine spent more time in the cocaine-paired chamber. Interestingly, a regression analysis did not compelling reveal that BDNF levels, overall, could account for CPP levels on extinction day 1 (R2[20]=0.013, p=0.314). Two strong coefficients of determination emerged when each group was considered individually: in the low BDNF WT rats, BDNF levels significantly accounted for CPP levels on extinction day 1 (R2[5]=0.650; p=0.050); in the high BDNF BDNF+/− rats the coefficient was also high, though not statistically significant (R2[4]=0.724; p=0.075). Analyses of all others were not statistically significant.
To examine the relation of BDNF levels to the effects of drug-induced reinstatement we calculated difference scores of cocaine-paired chamber times: time spent in conditioned chamber on extinction day 8 minus time spent there on priming day. Values increasing over zero would reflect greater reinstatement of the preference for the cocaine-paired chamber. A 2×2 ANOVA with factors Genotype and BDNF grouping did not reveal any significant main effects or interactions but planned comparisons revealed a significantly larger, positive difference score for WT rats categorized as high BDNF when compared to low BDNF WT rats (t[9]=−2.382, p=0.041; see Fig. 3D). There were no significant differences between the low and high BDNF rats in the BDNF+/− group.
4. Discussion
Until now, a CPP rat model had not been used to study the effects of a partial knockout of the BDNF gene on the rewarding effects of cocaine. The results of the present study can thus be used to further our understanding of the role of BDNF in cocaine addiction, particularly considering the extent of past research with rats in this domain. As expected, WT rats exposed to cocaine showed a place preference one day after conditioning and this preference was fully extinguished after 8 days of abstinence. A priming injection of cocaine reinstated cocaine-seeking behavior in WT rats. By contrast, BDNF+/− rats did not show a significant place preference following cocaine conditioning and consequently were also not responsive to the priming injection. Levels of BDNF are highly variable among individuals, thus we took serum BDNF samples, which are considered reflective of BDNF levels in the brain [17,12], before conditioning to examine their relation with rats’ behavior. Interestingly, high BDNF serum levels prior to conditioning predicted cocaine preference in WT rats on extinction day 1. Thus, rats with higher BDNF levels in the WT cocaine group were a substantial source of the significant CPP preference for extinction day 1. Additionally, WT rats with high BDNF levels had a significantly steeper increase in cocaine-seeking behavior from extinction day 8 to priming day compared to low BDNF WT rats. These patterns were not observed among the BDNF+/− rats. Overall, the results of this study demonstrated that rats bred to be heterozygous for the BDNF gene have diminished responses to the rewarding properties of cocaine and BDNF levels in WT rats may modulate cocaine CPP responses.
Conditioned place preference is generally used to understand the extent to which a substance is rewarding [14,20]. That BDNF+/− rats did not show a place preference for cocaine supports our hypothesis that reduced levels of BDNF reduce susceptibility to the acquisition of cocaine-seeking behaviors based on attenuation of the rewarding effect of cocaine. There is evidence that the role of BDNF in addiction and reward varies by brain region [18]. For example, injecting exogenous BDNF into the NA shell increases the amount of cocaine a rat will self-administer [8]. Conversely, a single BDNF infusion into the medial PFC attenuates cocaine-seeking [2]. Given that the effects of exogenous BDNF are complex and site-specific, future experiments could examine plasticity markers in areas thought to be affected by this genetic manipulation. One interpretation of our findings is that fewer neuroplastic changes to the reward pathway occurred as a result of reduced BDNF resulting in attenuation of the rewarding effects of cocaine. A possible alternative explanation is that the BDNF+/− cocaine rats did not exhibit a place preference because reduced BDNF levels caused general deficits in associative learning. However, a study using BDNF+/− mice suggests that a partial knockout of the BDNF gene does not abolish learning and memory: BDNF+/− mice lacked cocaine-seeking behavior in a CPP paradigm when administered 10 mg/kg cocaine, but showed a near normal pattern of behavior when administered a 20 mg/kg dose of cocaine [9]. This is evidence that general learning deficits are not responsible for reduced place preference in BDNF+/− mice, as they are capable of acquiring CPP; rather, this rightward shift in the dose-response curve implies a reduced sensitivity to the rewarding effects of cocaine. Additionally, the tendency for the BDNF+/− rats in the present study to show non-significant preferences for the cocaine-paired chamber suggests that a similar rightward shift in the dose-response curve likely would occur in BDNF+/− rats. Future experiments should be conducted to evaluate this hypothesis and elucidate the magnitude of the shift.
In the current study we measured relapse using priming injections of cocaine after the extinction of cocaine-seeking behavior. Relapse susceptibility is currently the best measure of cocaine addiction in humans [5]. A prospective study of human cocaine addicts seeking treatment revealed that elevated serum BDNF levels were associated with higher rates of relapse after 4 weeks of abstinence [5]. Therefore BDNF levels show promise as a predictor of cocaine relapse outcomes. One reason that we did not find significant differences based on BDNF serum levels on priming day could be that serum samples were collected prior to cocaine exposure and the critical measure may be the difference between baseline and post-conditioning levels. Interestingly, an increase in serum BDNF from baseline levels through a detoxification period correlated with increased craving to use cocaine in cocaine-dependent individuals [4]. Along this line BDNF levels could be collected at different stages of extinction and before a reinstatement paradigm. For this topic of research, BDNF knockout rats will be more useful than BDNF knockout mice because reinstating cocaine-seeking behavior via drug priming in mice is highly inconsistent [22].
BDNF is a protein that affects the brain in many ways, one of which is through long-term potentiation of dopaminergic synapses in the reward pathway [13]. As a result of having lower BDNF levels, BDNF+/− rats may have fewer neuroplastic changes in the reward pathway during cocaine exposure and thus did not develop a robust place preference for cocaine. That BDNF+/− rats did not relapse in a reinstatement paradigm strengthens this conclusion. Future study is warranted to investigate dose-dependent effects of cocaine in BDNF+/− rats, the specific effect of the BDNF partial knockout in the neural circuitry involved in reward, as well as the potential of BDNF levels to act as a predictor of relapse outcomes. The present findings also point to the utility of this genetic rat model in future studies of psychopathological processes related to dysfunction in the BDNF system, like addiction, but also including depression and schizophrenia.
Research Highlights.
BDNF+/− rats incompletely express brain derived neurotrophic factor
Wildtype rats show a cocaine preference in a CPP apparatus
BDNF+/− rats do not show a cocaine preference in a CPP apparatus
Reduced BDNF limits acquisition of cocaine seeking behaviors
Serum BDNF predicted cocaine-seeking behavior on extinction day 1
Acknowledgments
The authors would like to acknowledge SAGE® Labs (Sigma Advanced Genetic Engineering) in the use of the BDNF+/− rats (SD- Bdnf tm1sage, heterozygous; TGRS3850) and to thank Amanda Kimball for her editorial assistance on earlier drafts of this manuscript. This project was supported by grants from the National Center for Research Resources (5P20RR016463-12) and the National Institute of General Medical Sciences (8P20GM103423-12) from the National Institutes of Health to MJG.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Bahi A, Boyer F, Dreyer J. Role of accumbens BDNF and TrkB in cocaine-induced psychomotor sensitization, conditioned-place preference, and reinstatement in rats. Psychopharmacol. 2008;199:169–182. doi: 10.1007/s00213-008-1164-1. [DOI] [PubMed] [Google Scholar]
- 2.Berglind WJ, See RS, Fuchs RA, Ghee SM, Whitfield TM, Jr, Miller SW, McGinty JF. A BDNF infusion into the medial prefrontal cortex suppresses cocaine-seeking in rats. Euro J Neuro. 2007;26:757–766. doi: 10.1111/j.1460-9568.2007.05692.x. [DOI] [PubMed] [Google Scholar]
- 3.Berglind WJ, Whitfield TW, Jr, LaLumiere RT, Kalivas PW, McGinty JF. A single intra-PFC infusion of BDNF prevents cocaine-induced alterations in extracellular glutamate within the nucleus accumbens. J Neurosci. 2009;29:3715–3719. doi: 10.1523/JNEUROSCI.5457-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Corominas-Roso M, Roncero C, Eiroa-Orosa FJ, Gonzalvo B, Grau-Lopez L, Sanchez-Mora C, Ramos-Quiroga JA, Casas M. Brain-derived neurotrophic factor serum levels in cocaine-dependent patients during early abstinence. Eur Neuropsychopharmacol. 2012 doi: 10.1016/j.euroneuro.2012.08.016. http://dx.doi.org/10.1016/j.euroneuro.2012.08.016. [DOI] [PubMed]
- 5.D’Sa C, Fox HC, Hong AK, Dileone RJ, Sinha R. Increased serum brain-derived neurotrophic factor is predictive of cocaine relapse outcomes: a prospective study. Biol Psychiatry. 2011:706–711. doi: 10.1016/j.biopsych.2011.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.DeJong W. Relapse prevention: an emerging technology for promoting long-term drug abstinence. Int J Addict. 1994;29:681–705. doi: 10.3109/10826089409047904. [DOI] [PubMed] [Google Scholar]
- 7.Deroche-Gamonet V, Belin D, Piazza PV. Evidence for addiction-like behavior in the rat. Science. 2004;305:1014–1017. doi: 10.1126/science.1099020. [DOI] [PubMed] [Google Scholar]
- 8.Graham DL, Edwards S, Bachtell RK, DiLeone RJ, Rios M, Self DW. Dynamic BDNF activity in nucleus accumbens with cocaine use increases self-administration and relapse. Nat Neurosci. 2007;10:1029–1037. doi: 10.1038/nn1929. [DOI] [PubMed] [Google Scholar]
- 9.Hall FS, Drgonova J, Goeb M, Uhl GR. Reduced behavioral effects of cocaine in heterozygous brain-derived neurotrophic factor (BDNF) knockout mice. Neuropsychopharmacology. 2003;28:1485–1490. doi: 10.1038/sj.npp.1300192. [DOI] [PubMed] [Google Scholar]
- 10.Hyman SE, Malenka RC, Nestler EJ. Neural mechanisms of addiction: the role of reward-related learning and memory. Annu Rev Neurosci. 2006;29:565–598. doi: 10.1146/annurev.neuro.29.051605.113009. [DOI] [PubMed] [Google Scholar]
- 11.Kalivas PW, McFarland K. Brain circuitry and the reinstatement of cocaine-seeking behavior. Psychopharmacology (Berl) 2003;168:44–56. doi: 10.1007/s00213-003-1393-2. [DOI] [PubMed] [Google Scholar]
- 12.Klein AB, Williamson R, Santini MA, Clemmensen C, Ettrup A, Rios M, Knudsen GM, Aznar S. Blood BDNF concentrations reflect brain-tissue BDNF levels across species. Int J Neuropsychopharmacol. 2011;14:347–353. doi: 10.1017/S1461145710000738. [DOI] [PubMed] [Google Scholar]
- 13.Lu L, Dempsey J, Liu SY, Bossert JM, Shaham Y. A single infusion of brain-derived neurotrophic factor into the ventral tegmental area induces long-lasting potentiation of cocaine seeking after withdrawal. J Neurosci. 2004;24:1604–1611. doi: 10.1523/JNEUROSCI.5124-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lynch WJ, Nicholson KL, Dance ME, Morgan RW, Foley PL. Animal models of substance abuse and addiction: implications for science, animal welfare, and society. Comp Med. 2010;60:177–188. [PMC free article] [PubMed] [Google Scholar]
- 15.Mueller D, Stewart J. Cocaine-induced conditioned place preference: reinstatement by priming injections of cocaine after extinction. Behav Brain Res. 2000;115:39–47. doi: 10.1016/s0166-4328(00)00239-4. [DOI] [PubMed] [Google Scholar]
- 16.Nestler EJ. Historical review: Molecular and cellular mechanisms of opiate and cocaine addiction. Trends Pharmacol Sci. 2004;25:210–218. doi: 10.1016/j.tips.2004.02.005. [DOI] [PubMed] [Google Scholar]
- 17.Sartorius A, Hellweg R, Litzke J, Vogt M, Dormann C, Vollmayr B, Danker-Hopfe H, Gass P. Correlations and discrepancies between serum and brain tissue levels of neurotrophins after electroconvulsive treatment in rats. Pharmacopsychiatry. 2009;42:270–276. doi: 10.1055/s-0029-1224162. [DOI] [PubMed] [Google Scholar]
- 18.Schoenbaum G, Stalnaker TA, Shaham Y. A role for BDNF in cocaine reward and relapse. Nat Neurosci. 2007;10:935–936. doi: 10.1038/nn0807-935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Thomas MJ, Kalivas PW, Shaham Y. Neuroplasticity in the mesolimbic dopamine system and cocaine addiction. Br J Pharmacol. 2008;154:327–342. doi: 10.1038/bjp.2008.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Tzschentke TM. Measuring reward with the conditioned place preference paradigm: a comprehensive review of drug effects, recent progress and new issues. Prog Neurobiol. 1998;56:613–672. doi: 10.1016/s0301-0082(98)00060-4. [DOI] [PubMed] [Google Scholar]
- 21.Vanderschuren LJ, Everitt BJ. Drug seeking becomes compulsive after prolonged cocaine self-administration. Science. 2004;305:1017–1019. doi: 10.1126/science.1098975. [DOI] [PubMed] [Google Scholar]
- 22.Yan Y, Nabeshima T. Mouse model of relapse to the abuse of drugs: procedural considerations and characterizations. Behav Brain Res. 2009;196:1–10. doi: 10.1016/j.bbr.2008.08.017. [DOI] [PubMed] [Google Scholar]