

The increasing worldwide prevalence of obesity and its commonly associated disease type 2 diabetes has a dramatic influence on morbidity and mortality in the human population [1]. Among currently available obesity treatments, bariatric surgery achieves the best substantial and permanent weight loss [1]. Interestingly, surgeries inducing a rapid reduction in appetite and a normalization of glycemic index (e.g. Roux-en-Y gastric bypass) are notably associated with an increase in the anorexigenic gut peptides PYY and GLP-1 [1]. These two hormones act as peripheral sensory inputs integrated by the central nervous system (CNS) to modulate appetite and energy expenditure. The pharmacological mimicry of the hormonal milieu after bariatric surgery has thus been proposed as a potential strategy for the treatment of obesity [1]. However, such therapies will require a complete understanding of the mechanisms by which gut hormonal signals interact and integrate with other peripheral and central informations to modulate appetite.
PYY is an amidated peptide of 36-amino acids composed of tyrosine residues (Y) at each end. PYY is highly homologous to another gut hormone (pancreatic polypeptide) and to the central neurotransmitter NPY. Whereas PYY1-36 is an agonist of three mammalian NPY receptors (Y1, 2 and 5), PYY3-36, resulting from the digestion of PYY1-36 by the enzyme dipeptidyl peptidase 4 (DPP-4) is a selective Y2 receptor (Y2R) agonist. PYY is synthesized after the meal by the L endocrine cells of the small intestine in proportion to energy intake. The anorectic effect of PYY is specifically mediated by Y2R via interactions with both the vagus nerve and the arcuate nucleus of the hypothalamus [2]. In animals and humans, intravenous infusion of PYY3-36 reduces food intake in both lean and obese individuals [3]. Whereas the role of PYY3-36 in appetite control has been well characterized, its role in glucose homeostasis remains controversial.
In this issue, Chandarana and colleagues address the role of peripheral PYY on glucose homeostasis, including the context of bariatric surgery in mice [4]. The administration of PYY3-36 during an intraperitoneal glucose tolerance test (IPGTT) had a lowering effect on glycemia, which depended on Y2R. On the contrary, PYY1-36 administration had no effect. Moreover, the lowering effect of peripheral PYY3-36 on glycemia was not inhibited by the central administration of an Y2R antagonist, but only by the peripheral (IP) injection of such an antagonist. The lowering effect of PYY3-36 on glycemia occurred in parallel to an increase in blood insulin levels. However, whilst PYY1-36 had an inhibitory effect on insulin secretion as already described, PYY3-36 had no direct effect on insulin secretion. Interestingly, exogenous PYY3-36 decreased blood glucose levels only in a postprandial context. This is in line with the fact that endogenous PYY is secreted in response to lipids or glucose by intestinal L-cells [1]. The authors thus hypothesized that the postprandial increase in PYY levels measured in the portal vein after a meal could have a role in regulating postprandial insulin secretion via an incretin effect. Enterogastric anastomosis (EGA) performed in obese mice is an operation that mimics RYGBP surgery in humans [5]. The authors used this model to decipher the effect of PYY3-36 on glucose homeostasis. In wild-type EGA mice, the increase in PYY3-36 paralleled an increase in GLP-1. This was abrogated in PYY null mice. This suggested that PYY3-36 might regulate GLP-1 secretion. Accordingly, the authors showed that PYY3-36 enhanced glucose-stimulated and meal-stimulated GLP-1 secretion in the portal vein of control mice. Moreover, this effect was dependent on peripheral Y2R receptors. The use of exendin 9-39, an antagonist of GLP-1 receptor, confirmed that the positive effect of PYY3-36 on glucose homeostasis was, at least in part, due to GLP-1 action.
The main interest of these findings is that they complement previous studies on the importance of intestinal signals in glucose homeostasis by providing a new link dealing with the regulation of insulin secretion by PYY. The authors first showed that PYY3-36 controlled glucose homeostasis though a regulation of insulin secretion and secondly that this regulation is independent of central Y2R. On the contrary, the anorectic action of peripheral administration of PYY3-36 has been shown to be dependent on brain Y2R [6]. The highly interesting point of this study concerns the link between the two gut hormones PYY3-36 and GLP-1. Data from Chandarana and colleagues suggest that PYY3-36 improves insulin secretion after a meal in a GLP-1 dependent manner (Figure 1). However, a perceived weakness is the lack of a comprehensive cellular mechanism linking the secretions of both hormones. Future studies will have to decipher this mechanism. Important questions remain unresolved. For example, does PYY3-36 act directly on Y2R present in intestinal L-cells to induce and/or improve GLP-1 secretion? Does PYY3-36 act on Y2R via the autonomous nervous system, which is very dense within the walls of the portal vein [7], to stimulate GLP-1 secretion? Is there a role in this chain of the central nervous system via a reflex arc? It is noteworthy that a neural mechanism has been suggested to account at least in part for the anorectic effect of PYY3-36 [2] or for the benefits of GLP-1 on glucose control [8,9]. Moreover, it is concerning that the interplay between PYY, GLP-1 and Y2R may be complicated by the possible involvement of DPP-4. Indeed, PYY3-36 is produced from the digestion of PYY1-36 by DPP-4. The data reported in this study suggested that PYY3-36 improves glucose homeostasis, whilst PYY1-36 does not. Thus, DPP-4 inhibitors, which are currently used as a treatment to prolong the GLP-1 incretin effect in type-2-diabetes patients, might oppose rather than contribute to the PYY3-36 positive effect. If the mechanism suggested here were of primary importance, it would be hardly reconcilable with the benefits of DDP-4 inhibitors on GLP-1 incretin effects in pathophysiological situations [10].
Figure 1.
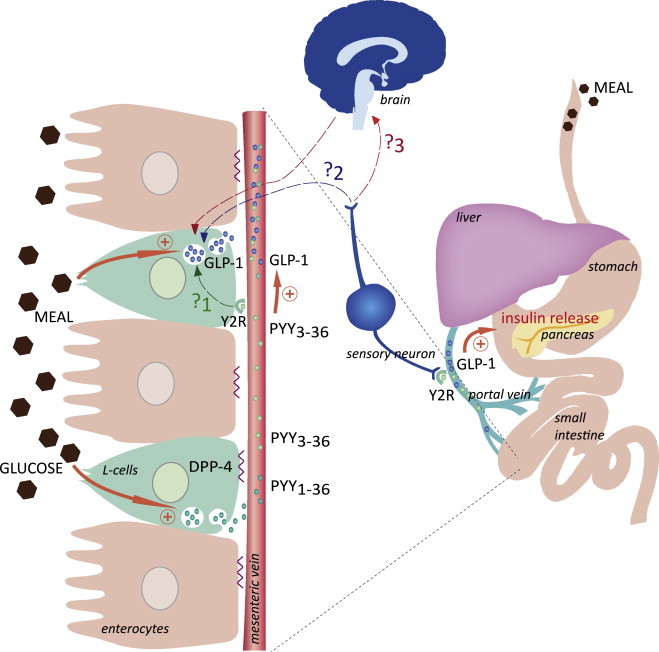
PYY3-36 improves insulin secretion after a meal in a GLP-1 dependent manner. The results obtained by Chandarana et al. are illustrated in orange (arrows): meal/glucose increase the levels of PYY3-36 in blood. PYY3-36, via the activation of peripheral Y2R, enhances GLP-1 secretion. As a result, GLP-1 levels increase in the portal vein leading to an increase in insulin secretion. Some questions that have to be solved to specify the cellular mechanisms linking the secretions of both hormones are illustrated in dotted lines.  : Does PYY3-36 act directly on Y2R expressed by intestinal L-cells to improve GLP-1 secretion?
: Does PYY3-36 act directly on Y2R expressed by intestinal L-cells to improve GLP-1 secretion?  : Does PYY3-36 act on Y2R expressed by neurons of the autonomic nervous system to improve GLP-1 secretion?
: Does PYY3-36 act on Y2R expressed by neurons of the autonomic nervous system to improve GLP-1 secretion?  : Is the brain involved in the enhanced GLP-1 secretion promoted by PYY3-36?
: Is the brain involved in the enhanced GLP-1 secretion promoted by PYY3-36?
In the obesity and diabetes field, it is obvious that implementing the knowledge on the peripheral effects of PYY3-36 could represent an important step in the understanding of the complex interplay between gut hormones, their maturation processes and their roles in regulating glucose and energy homeostasis. Further research in this field is warranted, since it may provide novel paradigms that could be useful in the elaboration of future approaches of prevention and/or treatment of metabolic diseases.
Conflict of interest
The authors wish to confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
Footnotes
This commentary refers to “Peripheral activation of the Y2-receptor promotes secretion of GLP-1 and improves glucose tolerance, by Keval Chandarana et al.” (10.1016/j.molmet.2013.03.001).
References
- 1.Field B.C., Chaudhri O.B., Bloom S.R. Bowels control brain: gut hormones and obesity. Nature Reviews Endocrinology. 2010;6:444–453. doi: 10.1038/nrendo.2010.93. [DOI] [PubMed] [Google Scholar]
- 2.Koda S., Date Y., Murakami N., Shimbara T., Hanada T., Toshinai K. The role of the vagal nerve in peripheral PYY3-36-induced feeding reduction in rats. Endocrinology. 2005;146:2369–2375. doi: 10.1210/en.2004-1266. [DOI] [PubMed] [Google Scholar]
- 3.Batterham R.L., Cohen M.A., Ellis S.M., Le Roux C.W., Withers D.J., Frost G.S. Inhibition of food intake in obese subjects by peptide YY3-36. New England Journal of Medicine. 2003;349:941–948. doi: 10.1056/NEJMoa030204. [DOI] [PubMed] [Google Scholar]
- 4.Chandarana K., Gelegen C., Irvine E.E, Choudhury A.I., Amouyal C., Andreelli F. Peripheral activation of the Y2-receptor promotes secretion of GLP-1 and improves glucose tolerance. Molecular Metabolism. 2013;2:142–152. doi: 10.1016/j.molmet.2013.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Troy S., Soty M., Ribeiro L., Laval L., Migrenne S., Fioramonti X. Intestinal gluconeogenesis is a key factor for early metabolic changes after gastric bypass but not after gastric lap-band in mice. Cell Metabolism. 2008;8:201–211. doi: 10.1016/j.cmet.2008.08.008. [DOI] [PubMed] [Google Scholar]
- 6.Abbott C.R., Small C.J., Kennedy A.R., Neary N.M., Sajedi A., Ghatei M.A. Blockade of the neuropeptide Y Y2 receptor with the specific antagonist BIIE0246 attenuates the effect of endogenous and exogenous peptide YY(3-36) on food intake. Brain Research. 2005;1043:139–144. doi: 10.1016/j.brainres.2005.02.065. [DOI] [PubMed] [Google Scholar]
- 7.Delaere F., Duchampt A., Mounien L., Seyer P., Duraffourd C., Zitoun C. The role of sodium-coupled glucose co-transporter 3 in the satiety effect of portal glucose sensing. Molecular Metabolism. 2013;2:47–53. doi: 10.1016/j.molmet.2012.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vahl T.P., Tauchi M., Durler T.S., Elfers E.E., Fernandes T.M., Bitner R.D. Glucagon-like peptide-1 (GLP-1) receptors expressed on nerve terminals in the portal vein mediate the effects of endogenous GLP-1 on glucose tolerance in rats. Endocrinology. 2007;148:4965–4973. doi: 10.1210/en.2006-0153. [DOI] [PubMed] [Google Scholar]
- 9.Mithieux G. Nutrient control of hunger by extrinsic gastrointestinal neurons. Trends in Endocrinology and Metabolism. 2013;24:378–384. doi: 10.1016/j.tem.2013.04.005. [DOI] [PubMed] [Google Scholar]
- 10.Crepaldi G., Carruba M., Comaschi M., Del Prato S., Frajese G., Paolisso G. Dipeptidyl peptidase 4 (DPP-4) inhibitors and their role in Type 2 diabetes management. Journal of Endocrinological Investigation. 2007;30:610–614. doi: 10.1007/BF03346357. [DOI] [PubMed] [Google Scholar]