

Abstract
To investigate whether palmitic acid can be responsible for the induction of inflammatory processes, HaCaT keratinocytes were treated with palmitic acid at pathophysiologically relevant concentrations. Secretion levels of interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), NF-κB nuclear translocation, NF-κB activation, Stat3 phosphorylation, and peroxisome proliferator-activated receptor alpha (PPARα) mRNA and protein levels, as well as the cell proliferation ability were measured at the end of the treatment and after 24 hours of recovery. Pyrrolidine dithiocarbamate (PDTC, a selective chemical inhibitor of NF-κB) and goat anti-human IL-6 polyclonal neutralizing antibody were used to inhibit NF-κB activation and IL-6 production, respectively. Our results showed that palmitic acid induced an upregulation of IL-6, TNF-α, IL-1β secretions, accompanied by NF-κB nuclear translocation and activation. Moreover, the effect of palmitic acid was accompanied by PPARα activation and Stat3 phosphorylation. Palmitic acid-induced IL-6, TNF-α, IL-1β productions were attenuated by NF-κB inhibitor PDTC. Palmitic acid was administered in amounts able to elicit significant hyperproliferation and can be attenuated by IL-6 blockage. These data demonstrate for the first time that palmitic acid can stimulate IL-6, TNF-α, IL-1β productions in HaCaT keratinocytes and cell proliferation, thereby potentially contributing to acne inflammation and pilosebaceous duct hyperkeratinization.
1. Introduction
Acne is a chronic inflammation of the pilosebaceous units in certain area, including face and trunk, that mainly occurs in adolescence. Its pathogenesis is complex and is dependent on the interplay of multiple factors such as genetic predisposition, excess of sebum production, abnormal follicular proliferation, and development of inflammation [1]. Inflammation is indicated as a key component of the pathogenesis of acne [2]. An immunological reaction to the gram-positive microbe Propionibacterium acnes may play a major role in the initiation of the inflammatory reaction [3]. However, some published studies also indicate that in addition to Propionibacterium acnes, some components such as free fatty acid (FFA), arachidonic acid, linoleic acid, and some proinflammatory cytokines are associated with acne inflammation, and inflammation-inducing effects may not depend on the presence of Propionibacterium acnes. In addition, peroxisome proliferator-activated receptor (PPAR)α and neural factors are also related to acne inflammation [4–7].
Human sebaceous glands secrete a lipid mixture containing squalene and wax esters, as well as cholesterolesters, triglycerides, and possibly some free cholesterol [8–10]. Sebaceous lipids are responsible for the three-dimensional organization of skin surface lipids and the integrity of the skin barrier [11]. Besides, sebaceous lipids and its products were detected to express proinflammatory and anti-inflammatory properties [6, 12]. The early study found that FFA induces skin inflammation and stimulates sebaceous duct hyperkeratosis in animal models. Zouboulis evaluated the experimental results of patients with inflammatory lesions successfully treated with a new anti-inflammatory agent that specifically blocks the formation of leukotriene (LT) B4, demonstrating a significant reduction of FFA in sebum [12]. The decrease in FFA directly correlated with the improvement of inflammatory lesions. It is known that bacterial hydrolases convert some of the triglycerides to FFA on the skin surface [13]; however, there is also evidence indicating that sebaceous glands can also synthesize considerable amounts of FFA [12]. The main ingredients of FFA secreted by sebaceous glands are linoleic acid (LA), palmitic acid (PA), and oleic acid (OA). The composition of comedonal free fatty acids has been studied, demonstrating that the proportion of LA is markedly decreased in acne comedones, while PA is significantly increased [14, 15]. Akamatsu et al. have found that the decreased levels of LA in acne comedones contribute, in part, to the worsening of acne inflammation by the failure of low level of LA to inhibit neutrophil ROS generation and phagocytosis [16]. Further studies have indicated that PA can reduce the neutrophils to produce hydrogen peroxide; by their role in oxidative stress and damage to the epidermal barrier function, the proinflammatory mediators thereby more easily pass through the hair follicles into the dermis and aggravate acne inflammation [17]. However, the mechanisms of FFA in inducing acne inflammation have not been thoroughly studied.
A number of proinflammatory cytokines, including interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and interleukin-1β (IL-1β), have been implicated in the inflammatory process of acne [18, 19]. IL-6 has been shown to be a key player in acute and chronic inflammation [20]. Serum IL-6 levels were significantly higher in acne patients than that in normal population, suggesting a role for IL-6 in the pathogenesis of acne [21]. TNF-α and IL-1β can be induced by NF-κB activation [22]. These two cytokines (TNF-α and IL-1β) propagate the acne inflammatory response by acting on endothelial cells to elaborate adhesion molecules to facilitate recruitment of inflammatory cells into the skin [23, 24].
The aim of this study was to investigate the possible role of PA in the initiation and development of inflammatory events using human HaCaT keratinocytes as model. We evaluated the effect of PA on IL-6, TNF-α, and IL-1β secretion in HaCaT cells. We also focused on the activation of NF-κB, which coordinates the expression of different proinflammatory genes, secretion of the cytokine IL-6, TNF-α, and IL-1β, and induction of PPARα. The latter inhibits the synthesis of proinflammatory molecules via a decreased activity of the NF-κB signaling pathway [25].
2. Materials and Methods
2.1. Materials
Palmitic acid (PA) powder was bought from Sigma Co. Ltd. PA powder was added to a 10% solution of fatty acid free BSA and dissolved by shaking gently overnight at 37°C to yield an 200 mmol/L solution of PA complexed to BSA. Antibodies for PPAR-α, p-Stat3, and total-Stat3 were bought from Cell Signaling Technology, California, USA; antibody for LaminB was bought from Santa Cruz Biotechnology, CA, USA; antibodies for NF-κB p65, IKKα, IκBα, β-Actin, Cy3-conjugated mouse anti-rabbit immunoglobulins, pyrrolidine dithiocarbamate (PDTC, a selective chemical inhibitor of NF-κB), cell culture supplies, CCK-8 Kit, and BCA Protein Assay Kit were all bought from Beyotime Institute of Biotechnology; Trizol was bought from Invitrogen, Carlsbad, CA; real-time PCR Assay Kits were bought from Nanjing KGI Bioteknologi Development Co, Ltd. IL-6 Sandwich ELISA Kit was bought from JingMei Bioengineer Company, Shenzhen. Helenalin was bought from Merlin Standard Chemicals, Singapore. Goat anti-human IL-6 polyclonal neutralizing antibody was obtained from R&D Systems, Minneapolis, MN, USA.
2.2. Method
2.2.1. Cell Culture
Keratinocyte line HaCaT cells were cultured in a cell incubator at 37°C, 5% CO2, in DMEM medium containing 10% fetal bovine serum and 1% penicillin and streptomycin. After cells became polygon arranging as a single layer, they were vaccinated at the density of 1 × 109/L with 0.25% trypsin solution. The cultured cells were used for experiment when they adhered to the culture plate and the confluence reached 70%~80%.
2.2.2. Experiment Grouping and Treatment of Cells
HaCaT keratinocytes were either without or with pretreatment for 1 hour with 10 μM PDTC and then treated in serum-free conditions for 24 hours with PA at concentrations of 75, 100, 125, and 150 μmol/L. For certain experiments, goat anti-human IL-6 polyclonal neutralizing antibody was added into cell culture system at a concentration of 10 μg/mL. At the end of the treatment, fresh medium was added. Cells were collected at the end of the treatment or after 24 hours from the addition of fresh medium. As for IL-6, IL-1β, and TNF-α release detection, at the end of the treatment, supernatants were collected, fresh medium was added, and free cell supernatants were collected after 24 hours.
2.2.3. Immunofluorescence and Confocal Microscopy Detection of NF-κB p65 in HaCaT Cells
The cells were washed with 0.01 M phosphate-buffered saline (PBS) and fixed in 4% formaldehyde for 30 min at room temperature. After being permeabilized with 1% Triton X-100 for 10 min, the cells were blocked with PBS containing 5% bovine serum albumin for 30 min at room temperature and immunofluorescent staining was performed using a specific mouse polyclonal antibody against NF-κB p65 (dilution, 1 : 500) followed by Cy3-conjugated mouse anti-rabbit immunoglobulins (red). The slides were counterstained with Hoechst 33258 (blue). Finally, the cover slips were mounted on the slides and fluorescence was visualized using a confocal laser fluorescence microscope (Carl Zesis Zen 2008, Carl Zeiss Inc., Germany). Photographic images were taken from five random fields. Substitution of the primary antibody with a normal mouse IgG was used as control.
2.2.4. Preparation of Cytoplasmic and Nuclear Extracts
Briefly, cells were scraped from dishes in PBS, pelleted, washed in hypotonic buffer (10 mM HEPES buffer, pH 7.9, 1.5 mM MgCl2, 5 mM KCl, 1 mM PMSF, 1 mM dithiothreitol, 1 mM Na3VO4, 1 mM NaF), and lysed by resuspension in the same buffer with 0.1% Nonidet P-40. Cytoplasmic extracts were isolated by centrifugation at 10,000 rpm for 10 min. Nuclear pellets were washed in hypotonic buffer and resuspended in cold extraction buffer (20 mM HEPES, 25% glycerol, 450 mM KCl, 1 mM EDTA, 1 mM PMSF, 1 mM Na3VO4, 1 mM NaF), gently agitated at 4°C for 45 min, and spun at 13,000 rpm for 30 min at 4°C. Supernatants were collected, and protein concentrations were determined with BCA Protein Assay Kit.
2.2.5. Western Blotting Tests for the Protein Expression of NF-κB p65, IKKα, IκBα, p-Stat3, and PPAR-α in HaCaT Cells
An aliquot of protein extracted from cytoplasmic or nuclear extracts was subjected to 10% SDS-PAGE by electrophoresis under reducing conditions and transferred to PVDF membrane. The blotted membrane was then blocked with 5% nonfat dry milk in 1 × TBS (0.1% Tween 20) for 1 h at room temperature and incubated overnight at 4°C with primary antibodies to NF-κB p65 (dilution, 1 : 500), to IKKα (dilution, 1 : 250), to IκBα (dilution, 1 : 250), to PPAR-α (dilution, 1 : 500), to p-Stat3 (dilution, 1 : 500), to total-Stat3 (dilution, 1 : 500), to β-actin (dilution, 1 : 2,000), and to LaminB (dilution, 1 : 1,000). Following the incubation with horseradish peroxidase-conjugated sheep anti-mouse secondary antibodies (dilution, 1 : 2,000) for 1 h at room temperature, the blotted membrane was detected by Pierce ECL reagents (Thermo Fisher Scientific) and captured on X-ray film. Quantification of protein bands was established by Band-Scan software (PROZYME, San Leandro, California, USA) and expressed as a value relative to the density of the internal control (β-actin or LaminB).
2.2.6. Real-Time PCR Detection of the Expression of PPAR-α mRNA in HaCaT Cells
Trizol was added to break down the cells, followed by extraction of total RNA, measurement of concentration, and then measurement of purity. After ensuring that the quality met the requirements of the experiment, cDNA was obtained by reverse transcription. It was diluted 10 times and amplified according to a 20 μL reaction system. Primers were synthesized by Nanjing Kaiji Bio-tech Co., Ltd. (Table 1). Amplification conditions are as follows: pre-degeneration at 95°C for 5 min, entering reaction circles, degeneration at 95°C for 15 min, annealing for 30 s at 60°C, extending for 30 s at 72°C, keeping at 72°C for 10 min after 40 cycles.
Table 1.
Primers used in the real-time RT-PCR amplification of the human PPAR-α gene and GAPDH mRNAs.
| Gene name | primer sequences | |
|---|---|---|
| PPARα | Forward primer | 5′-TTCGCAATCCATCGGCGAG-3′ |
| Reverse primer | 5′-CCACAGGATAAGTCACCGAGG-3′ | |
|
| ||
| GAPDH | Forward primer | 5′-TGTTGCCATCAATGACCCCTT-3′ |
| Reverse primer | 5′-CTCCACGACGTACTCAGCG-3′ | |
2.2.7. ELISA Analysis of the Expression of IL-6, IL-1β, and TNF-α in Cell Supernatants
Measurement of IL-6, IL-1β, and TNF-α were performed using commerical ELISA kits. This assay uses the quantitative sandwich immunoassay technique. The standard curve demonstrated a direct relationship between OD and secreted cytokine levels.
2.2.8. Cell Proliferation Assay
Cell proliferation was assayed using a CCK-8 Kit. In brief, 100 μL of cells (2 × 103 cells/well) was transferred into 96-well plates after digestion with trypsin, and five parallel wells were used for each treatment. After attachment, the cells were subjected to the different treatments and then cultured for 24 h in a 5% CO2 incubator at 37°C. Subsequently, 10 μL of CCK-8 was added to each well, and the cells were cultured for another 3 h. Cell density was determined by measuring the absorbance at 450 nm using a Varioskan Flash (Thermo Scientific, USA).
2.3. Statistical Analysis
SPSS13.0 software was used for data analysis, and the form of average ± standard deviation was used to indicate measurement data. ANOVA was used for intergroup comparison, P < 0.05 was considered statistically significant.
3. Results
3.1. Induction of IL-6, IL-1β, and TNF-α Secretion in HaCaT Keratinocytes by PA
We observed, by means of ELISA analysis, an increase in a dose-dependent manner in the release of IL-6, IL-1β, and TNF-α in HaCaT keratinocyte supernatant treated with PA at a concentration of 75, 100, 125, and 150 μmol/L 24 hours after PA removal (Figures 1(a), 1(b) and 1(c)). The obtained results demonstrate that PA is able to induce an inflammatory stimulus in HaCaT keratinocytes by increasing IL-6, IL-1β, and TNF-α secretion.
Figure 1.
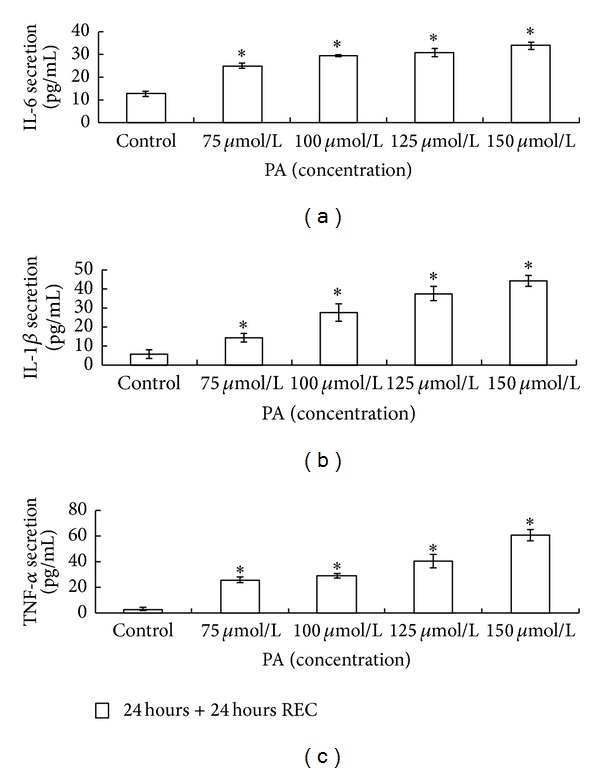
PA induces an upregulation of IL-6, IL-1β, and TNF-α secretion in HaCaT keratinocytes. HaCaT keratinocytes were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours under serum-free conditions, and collected 24 hours after PA depletion. IL-6 (a), IL-1β (b), and TNF-α (c) releases were determined by ELISA kits. Results are expressed as average mean of protein concentration (pg/mL) ± SD and represent the mean of three experiments in duplicate. Asterisks (*) indicate significant differences of P < 0.05, respectively, between the PA-treated groups and nontreated group.
3.2. Induction of NF-κB Nuclear Translocation in HaCaT Keratinocytes by PA
We used immunofluorescence staining to examine the localization of NF-κB p65 in HaCaT keratinocytes. NF-κB p65 was stained with Cy3-conjugated mouse anti-rabbit immunoglobulins (red) and nuclei were stained with Hoechst 33342 (blue). We found that NF-κB p65 positive staining was predominantly localized in control cytoplasm (Figure 2). Interestingly, NF-κB p65 staining significantly shifted to the nuclei with PA stimulation immediately after the treatment (Figure 2(a)) and 24 hours after the PA depletion (Figure 2(b)). The nuclear translocation of NF-κB p65 subunit was further confirmed by the results derived from the western blotting studies. Upregulation of nuclear NF-κB p65 protein levels in a dose-dependent manner was observed in cells treated with PA at 75, 100, 125, and 150 μmol/L compared to control, immediately after the treatment (Figure 3(a)). The level of nuclear NF-κB p65 protein expression in treated cells also increased dose dependently 24 hours after PA removal (Figure 3). The results obtained in these results demonstrate that PA is able to induce NF-κB nuclear translocation in HaCaT keratinocytes.
Figure 2.
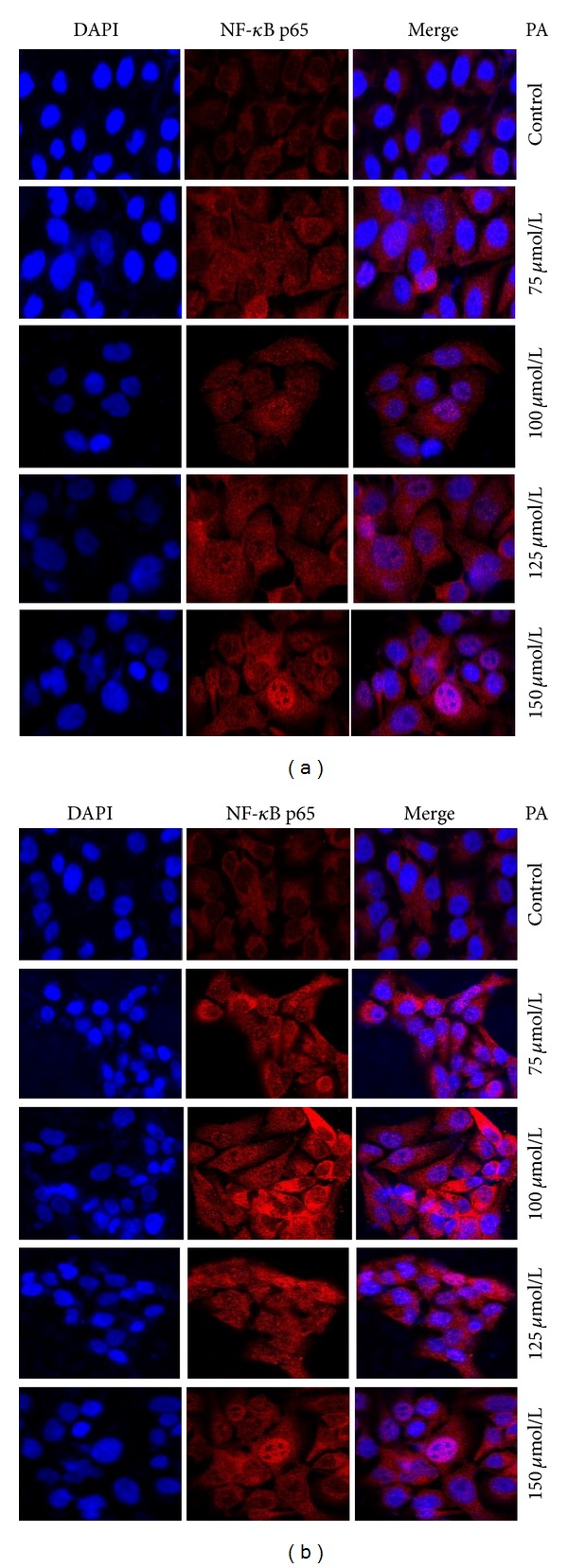
PA-induced nuclear translocation of NF-κB p65 subunit in HaCaT cells immediately after the treatment and 24 hours after the PA depletion. Keratinocytes were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours under serum-free conditions. Immunostaining was performed with specific mouse anti-p65 antibody followed by Cy3-conjugated mouse anti-rabbit immunoglobulins (red) immediately after the treatment (a) and 24 hours after the PA depletion (b). Images are representative of three independent experiments.
Figure 3.
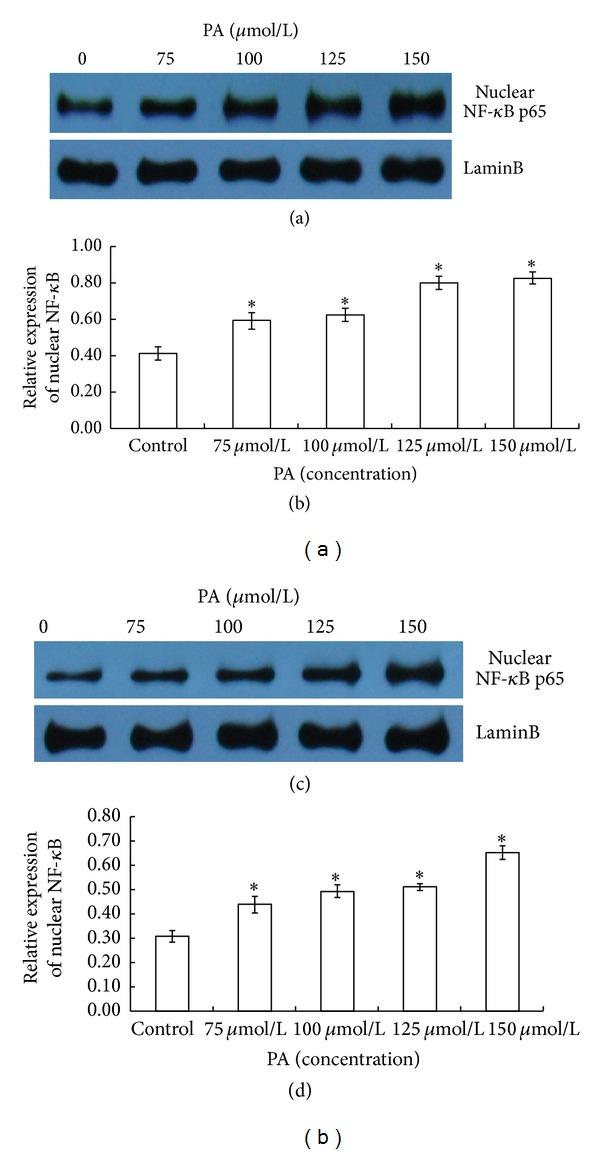
PA induces nuclear translocation of NF-κB p65 subunit in HaCaT cells measured by western blotting. Keratinocytes were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours under serum-free conditions. The cells were harvested, and nuclear extracts were analyzed with NF-κB p65 by western blot immediately after the treatment (a) and 24 hours after the PA depletion (c). The densitometry values are means ± SD of three independent experiments ((b) and (d)). Asterisks (*) indicate significant differences of P < 0.05, respectively, between the PA-treated groups and nontreated group.
3.3. Induction of IKKα Activation and IκBα Degradation in HaCaT Keratinocytes by PA
Upregulation of IKKα protein levels in a dose dependent manner were observed in cells treated with PA at 100, 125, and 150 μmol/L compared to control, immediately after the treatment and 24 hours after the PA depletion (Figures 4(a) and 4(b)). The level of IκBα protein expression in 75, 100, 125, and 150 μmol/L PA treated cells decreased dose dependently immediately after the treatment and 24 hours after the PA depletion (Figures 4(d) and 4(e)). The results obtained in these results demonstrate that PA is able to induce NF-κB activation in HaCaT keratinocytes.
Figure 4.
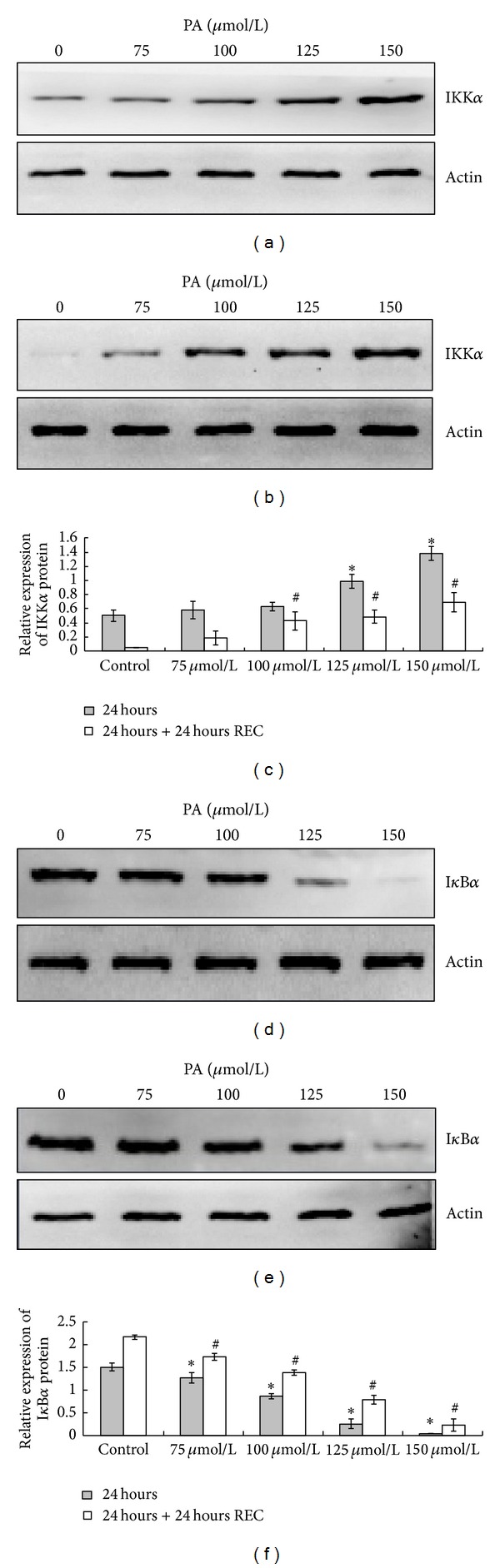
PA induces activation of IKKα and degradation of IκBα in HaCaT cells measured by western blotting. Keratinocytes were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours under serum-free conditions. The cells were harvested, and nuclear extracts were analyzed with IKKα by western blot immediately after the treatment (a) and 24 hours after the PA depletion (b). The densitometry values are means ± SD of three independent experiments (c). The protein expression of IκBα immediately after the treatment (d) and 24 hours after the PA depletion (e) were also determined by western blot. The densitometry values are means ± SD of three independent experiments (f). *indicate significant differences of P < 0.05, respectively, between the PA-treated groups and nontreated group immediately after the treatment groups (24 hours); #indicate significant differences of P < 0.05, respectively, between the PA-treated groups and nontreated group in 24 hours after the PA depletion groups (24 hours + 24 hours REC).
3.4. Induction of PPARα Expression and Phospho-Stat3 in HaCaT Keratinocytes by PA
Immediately after treatment, PA at 75, 100, 125, and 150 μmol/L caused an upregulation of PPARα mRNA levels compared to controls (Figure 5(a)). Afterwards, the level of PPARα gene expression in the treated cells decreased during the following 24 hours (Figure 5(a)). Significant upregulation of PPARα protein levels was observed by means of western blot analysis in cells treated with PA at 100, 125, and 150 μmol/L compared to controls, immediately and 24 hours after PA removal (Figures 5(b) and 5(c)). On the contrary, the treatment of HaCaT keratinocytes with PA at 75 μmol/L did not exert any effect on the expression pattern of PPARα, compared to untreated controls (Figures 5(b) and 5(c)). The PPARα upregulation supports the data mentioned above, which highlighted the possible induction of inflammatory response in HaCaT keratinocytes after the treatment with PA. Significant upregulation of levels of p-Stat3 was also observed by means of western blot analysis in cells treated with PA at 100, 125, and 150 μmol/L compared to total Stat3, immediately and 24 hours after PA removal (Figures 6(a) and 6(b)).
Figure 5.
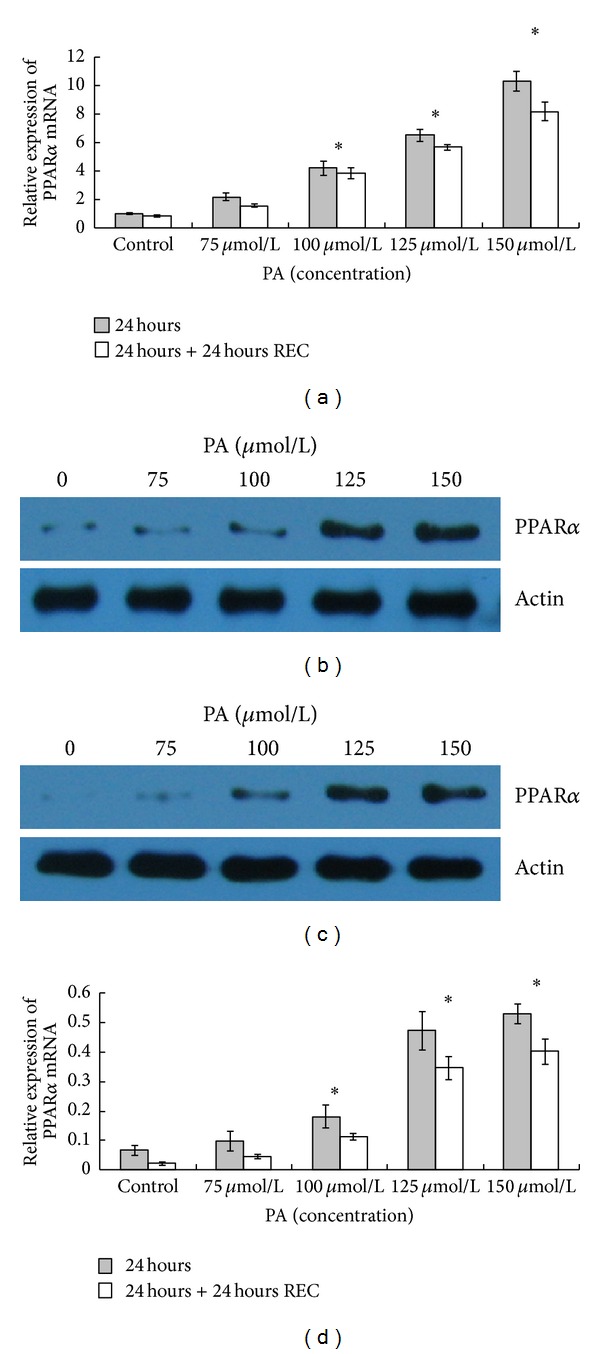
PA induces an upregulation of PPARα mRNA and protein levels in HaCaT keratinocytes. HaCaT keratinocytes were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours in serum-free conditions, and collected immediately and 24 hours after PA depletion. (a) PPAR-α mRNA levels were determined by real-time RT-PCR. The values shown represent mean ± SD of three experiments. Expression of PPARα after treated or untreated with PA immediately (b) and 24 hours (c) were detected by western blotting. The band intensities were evaluated by densitometric analysis (d). The values shown represent the mean ± SD of three experiments. Asterisks (*) indicate significant differences of P < 0.05, respectively, between the PA-treated groups and nontreated group.
Figure 6.
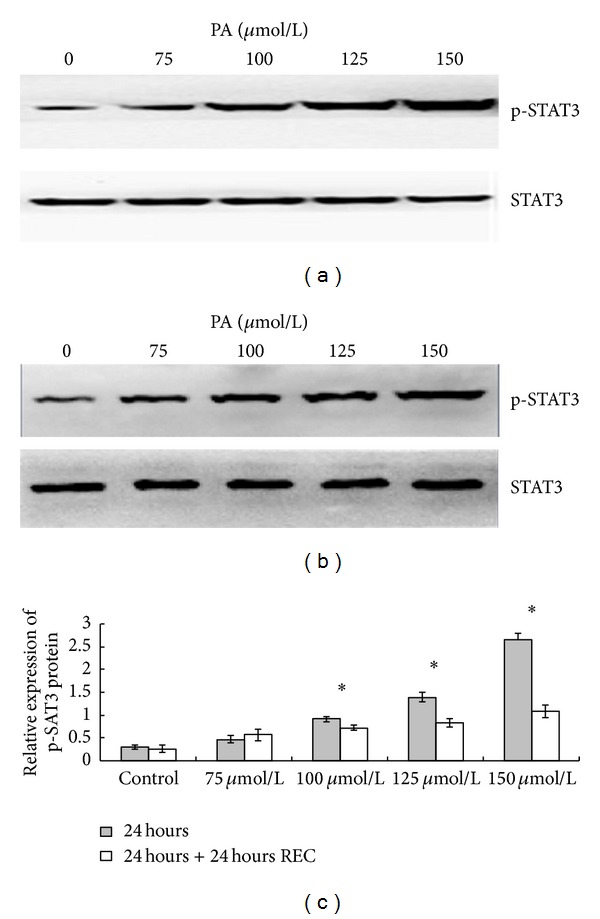
PA induces activation of p-Stat3 (Tyr705) in HaCaT cells measured by western blotting. Keratinocytes were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours under serum-free conditions. The cells were harvested, and nuclear extracts were analyzed with p-Stat3 and total Stat3 by western blot immediately after the treatment (a) and 24 hours after the PA depletion (b). The densitometry values are means ± SD of three independent experiments (c). Asterisks (*) indicate significant differences of P < 0.05, respectively, between the PA-treated groups and nontreated group.
3.5. PA-Induced IL-6, IL-1β, and TNF-α Production Is Attenuated by Inhibitor of NF-κB
We examined the effect of inhibition of NF-κB on IL-6, IL-1β, and TNF-α expression in response to PA. As shown by CCK-8 detection, pretreatment for 1 hour with 10 μM PDTC, a selective chemical inhibitor of NF-κB, had no obvious toxicity to HaCaT cell viability. Moreover, 10 μM PDTC was proved to inhibit PA-induced IKKα activation and IκBα degradation (see supplementary Figure S1 in Supplementary Material available online at http://dx.doi.org/10.1155/2013/530429). The solvent dimethylsulfoxide (0.1%) was used as a vehicle and control. PA-stimulated IL-6, IL-1β, and TNF-α productions were significantly attenuated by PDTC, showing that these PA-induced proinflammatory cytokine expressions involve activation of NF-κB activation (Figure 7). Dimethylsulfoxide had no effect on PA-induced IL-6, IL-1β, and TNF-α production.
Figure 7.
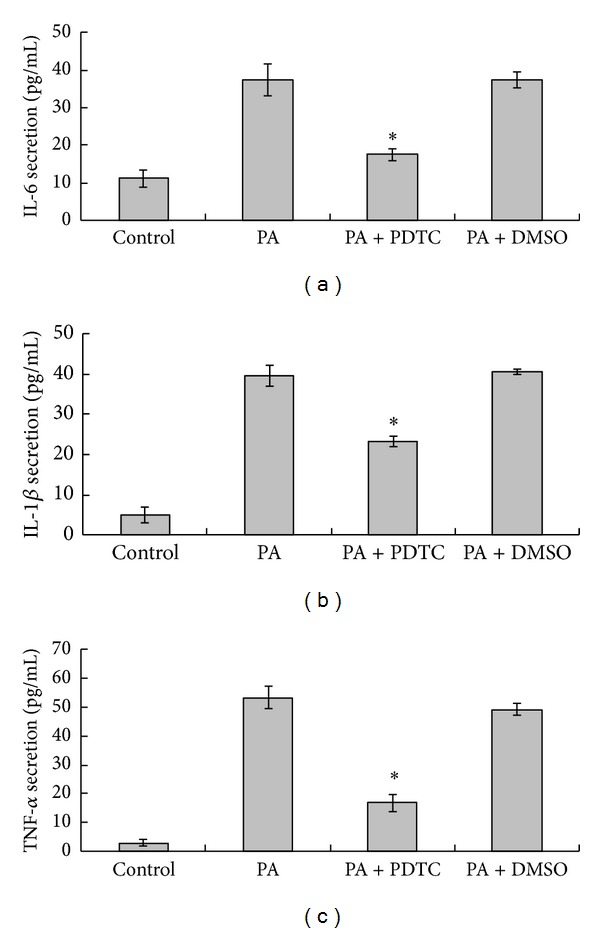
Effect of NF-κB inhibitor PDTC on PA-induced IL-6, IL-1β, and TNF-α production. HaCaT cells were untreated or exposed to 0.15 mM PA for 24 hours, either without or with pretreatment for 1 hour with 10 μmol/L PDTC or 0.1% dimethylsulfoxide (DMSO, as vehicle control). IL-6 (a), IL-1β (b), and TNF-α (c) productions were measured by ELISA. Each experiment was done in triplicate. Data are mean ± standard deviation. *P < 0.05 compared with PA.
3.6. Induction of Cell Proliferation by PA Is Dependent of IL-6 Production
In order to evaluate the possible role of PA in the hyperkeratinization of the pilosebaceous duct, we focused on the proliferative response of HaCaT keratinocytes after treatment with PA in a concentration range of 75 to 150 μmol/L by means of CCK-8 test. We observed that PA concentrations between 75 and 150 μmol/L induced a significant proliferative stimulus (Figure 8(a)). These data suggest that PA may be involved in the induction of the hyperkeratosis of the pilosebaceous duct. Interestingly, PA-induced cell proliferative effect was significantly attenuated by the addition of 10 μg/mL goat anti-human IL-6 polyclonal neutralizing antibody in cell culture system (Figure 8(b)). These data demonstrate that autocrine IL-6 production is causally linked to cell proliferation in this in vitro model.
Figure 8.
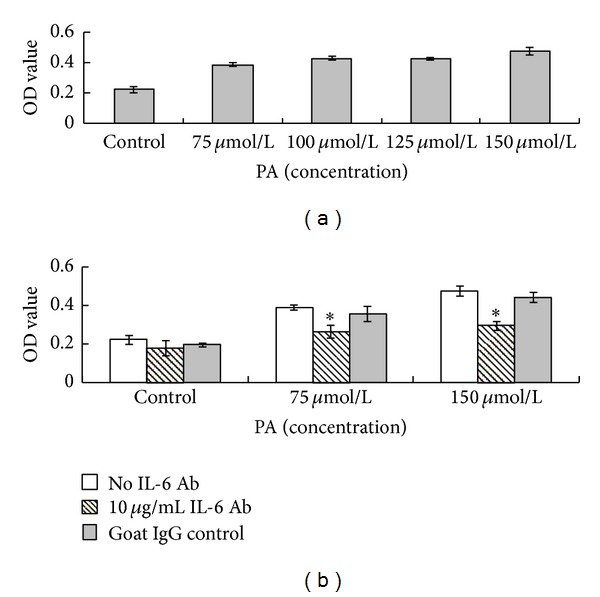
Induction of cell proliferation by palmitic acid is dependent of IL-6. (a) Cells were untreated or treated with PA (75, 100, 125, and 150 μmol/L) for 24 hours. Cell viability was measured by a CCK-8 assay kit. Data are expressed as mean ± standard deviation (n = 5). *P < 0.05 compared with control. (b) Cells were untreated or treated with PA (75 and 150 μmol/L) for 24 hours, with or without the presence of 10 μg/mL goat anti-human IL-6 polyclonal neutralizing antibody (IL-6 Ab) or control goat IgG in cell culture system. Cell viability was measured by a CCK-8 assay kit. Data are expressed as mean ± standard deviation (n = 5). *P < 0.05 compared with treatments without IL-6 Ab.
4. Discussion
In this study, we supplemented HaCaT cells with PA to mimic the influx of excess FFAs into keratinocytes. For experiments, we used the human keratinocyte cell line HaCaT, which retains biochemical and morphological properties characteristic of keratinocytes, and has proven useful for studying the possible roles of inflammatory mediators in acne vulgaris [25].
Our data demonstrate that exposure of keratinocytes (HaCaT cells) to pathophysiologically relevant concentrations of PA results in increased IL-6, IL-1β, and TNF-α secretion. This is in agreement with several studies that suggest a link between these three proinflammatory cytokines and PA-induced inflammation [26, 27]. For example, Staiger et al. reported palmitate-induced interleukin-6 expression in human coronary artery endothelial cells and suggested a potential contribution of palmitate to vascular inflammation [27]. Importantly, evidence from previous publications shows that IL-6 is elevated in patients with acne, suggesting that this inflammatory cytokine also may contribute to the development of acne [21]. It was reported that PA significantly decreased H2O2 generation both by neutrophils and in the xanthine-xanthine oxidase system, while neutrophil chemotaxis and phagocytosis as well as O2 − and OH∙ generation by both systems were not markedly affected in the presence of PA [17]. The authors suggested that PA may be involved in the pathogenesis of acne inflammation from a standpoint of oxidative tissue injury [17]. Besides, it is well documented that IL-6 phosphorylates transcription factor Stat3 at Tyr705 residue and has a role in inflammation [28–30]. Our present data showed that the p-Stat3 level was significantly induced following PA treatment. In this context, we suggested that PA may also contribute to acne inflammation via increasing IL-6 secretion.
Besides IL-6, we also found that TNF-α and IL-1β were increased following PA treatment, which are also critically important in acne inflammation. These two proinflammatory cytokines not only amplify the NF-κB signaling pathways that originally led to their production through cell surface receptor activation (an autocrine loop), but also will stimulate nearby cells in a paracrine manner. For example, TNF-α and IL-1β are known to upregulate adhesion molecules, such as ICAM-1 and VCAM-1 on endothelial cells [31, 32]. Thus, the observation that ICAM-1, E-selectin, and VCAM-1 expression levels on the luminal surface of endothelial cells are increased in inflammatory acne papules may be a consequence of TNF-α and IL-1β induction in the milieu [33]. The elaboration of adhesion molecules is necessary to slow the flow of circulating inflammatory cells for their eventual diapedesis into the inflamed tissue [31, 32].
NF-κB is known to be an important transcription factor for proinflammatory gene expression, and appears to regulate IL-6, IL-1β, and TNF-α secretions in a cell type-specific and stimulus-specific manner [34, 35]. Activation of NF-κB is mediated through the action of a family of serine/threonine kinases known as IκB kinase (IKK). The IKK (IKKα and/or IKKβ) phosphorylates IκB proteins and the members of the NF-κB family. In our current study, we observed that NF-κB p65 is activated in a dose-dependent manner in human HaCaT cells following PA exposure and subsequently translocated to the nucleus. PA exposure also resulted in an increased degradation of IκBα protein. This suggests that the activation of NF-κB p65 in HaCaT cells is mediated through the inhibition of IκBα protein proteolysis. It is well documented that IκBα is bound to NF-κB p65 through a protein-protein interaction and thus prevents migration of NF-κB p65 into the nucleus [36]. Additionally, the IKK complex is an important site for integrative signals that regulate the NF-κB pathway. We observed that PA exposure resulted in an increase in IKKα protein expression in HaCaT cells. These data suggest that PA-induced NF-κB activation and nuclear translocation of NF-κB p65 through greater activation of IKKα and degradation of IκBα proteins. Besides, PA-induced IL-6, IL-1β, and TNF-α expressions were attenuated by a NF-κB inhibitor, strongly implicating this transcription factor in the regulation of PA-induced IL-6, IL-1β, and TNF-α expressions. PPARα exerts a modulatory role in the control of the inflammatory response by antagonizing NF-κB signaling pathway. After the treatment with PA, HaCaT keratinocytes exhibited higher levels of PPARα transcripts probably as a feedback in anti-inflammatory response to the stimulus.
Our study clearly demonstrates that exposure to excess PA induces HaCaT keratinocyte proliferation and IL-6 production in HaCaT keratinocytes. IL-6 is reported to stimulate keratinocyte proliferation and is therefore studied in diseases associated with epidermal hyperplasia and in wound healing [37–39]. Sebaceous duct keratinocytes from comedones exhibit a hyperproliferative response compared to normal keratinocytes [40, 41]. The hyperproliferative behavior of HaCaT keratinocytes induced by PA could support the idea that FFA may be involved in comedone formation and, in particular, that PA may be responsible for this event. However, whether IL-6 is responsible for the PA-induced HaCaT keratinocyte proliferation is not yet clear. Our data suggested that the PA-induced HaCaT cell proliferation was attenuated by adding IL-6 polyclonal neutralizing antibody into cell culture system, strongly indicating that autocrine of IL-6 is primarily responsible for regulation of PA-induced keratinocytes proliferation. Hence, the interrelation between PA-induced keratinocyte proliferation and IL-6 production may be an important factor in comedone formation.
In conclusion, we show for the first time that one major type of FFA, palmitic acid, stimulates human HaCaT keratinocytes to produce proinflammatory cytokines IL-6, IL-1β, and TNF-α in a dose-dependent manner, via activation of NF-κB. Besides, the elevated level of IL-6 may also contribute to PA-induced keratinocyte proliferation.
Supplementary Material
Supplementary Figure: Effect of NF-κB inhibitor PDTC on HaCaT cell viability and PA–induced NF-κB activation. (A) HaCaT cells were untreated or exposed to PDTC (10, 50, 100 μmol/L) for 1 hour. Cell viability was measured by a CCK-8 assay kit. Data are expressed as mean ± standard deviation (n = 5). ∗P < 0.05 compared with control. (B) HaCaT cells were untreated or exposed to 0.15 mM palmitic acid (PA) for 24 hours, either without or with pretreatment for 1 hour with 10 μmol/L PDTC or 0.1% dimethylsulfoxide (DMSO, as vehicle control). IKKα and IκBα expression levels were measured by western blotting. Each experiment was done in triplicate. Data are mean ± standard deviation. ∗P < 0.05 compared with PA.
Conflict of Interests
The authors declare that they have no conflict of interest.
Authors' Contribution
Bing-rong Zhou and Jia-an Zhang contributed equally to the paper.
Acknowledgments
This work was supported by Grant from the China National Natural Science Foundation (81000700 and 81171518), science project from traditional Chinese medicine Bureau of Jiangsu Province (LZ11084), and Jiangsu National Natural Science Foundation (BK2012877).
References
- 1.Zouboulis CC, Eady A, Philpott M, et al. What is the pathogenesis of acne? Experimental Dermatology. 2005;14(2):143–152. doi: 10.1111/j.0906-6705.2005.0285a.x. [DOI] [PubMed] [Google Scholar]
- 2.Fleischer AB., Jr. Inflammation in rosacea and acne: implications for patient care. Journal of Drugs in Dermatology. 2011;10(6):614–620. [PubMed] [Google Scholar]
- 3.Jappe U, Ingham E, Henwood J, Holland KT. Propionibacterium acnes and inflammation in acne; P. acnes has T-cell mitogenic activity. British Journal of Dermatology. 2002;146(2):202–209. doi: 10.1046/j.1365-2133.2002.04602.x. [DOI] [PubMed] [Google Scholar]
- 4.Lee WJ, Jung HD, Lee HJ, Kim BS, Lee S-J, Kim DW. Influence of substance-P on cultured sebocytes. Archives of Dermatological Research. 2008;300(6):311–316. doi: 10.1007/s00403-008-0854-1. [DOI] [PubMed] [Google Scholar]
- 5.Toyoda M, Nakamura M, Makino T, Kagoura M, Morohashi M. Sebaceous glands in acne patients express high levels of neutral endopeptidase. Experimental Dermatology. 2002;11(3):241–247. doi: 10.1034/j.1600-0625.2002.110307.x. [DOI] [PubMed] [Google Scholar]
- 6.Toyoda M, Morohashi M. New aspects in acne inflammation. Dermatology. 2003;206(1):17–23. doi: 10.1159/000067818. [DOI] [PubMed] [Google Scholar]
- 7.Sertznig P, Reichrath J. Peroxisome proliferator-activated receptors (PPARs) in dermatology: challenge and promise. Dermato-Endocrinology. 2011;3(3):130–135. doi: 10.4161/derm.3.3.15025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nikkari T. Comparative chemistry of sebum. Journal of Investigative Dermatology. 1974;62(3):257–267. doi: 10.1111/1523-1747.ep12676800. [DOI] [PubMed] [Google Scholar]
- 9.Ramasastry P, Downing DT, Pochi PE, Strauss JS. Chemical composition of human skin surface lipids from birth to puberty. Journal of Investigative Dermatology. 1970;54(2):139–144. doi: 10.1111/1523-1747.ep12257164. [DOI] [PubMed] [Google Scholar]
- 10.Thody AJ, Shuster S. Control and function of sebaceous glands. Physiological Reviews. 1989;69(2):383–416. doi: 10.1152/physrev.1989.69.2.383. [DOI] [PubMed] [Google Scholar]
- 11.Pilgram GSK, van der Meulen J, Gooris GS, Koerten HK, Bouwstra JA. The influence of two azones and sebaceous lipids on the lateral organization of lipids isolated from human stratum corneum. Biochimica et Biophysica Acta. 2001;1511(2):244–254. doi: 10.1016/s0005-2736(01)00271-1. [DOI] [PubMed] [Google Scholar]
- 12.Zouboulis CC. Is acne vulgaris a genuine inflammatory disease? Dermatology. 2001;203(4):277–279. doi: 10.1159/000051771. [DOI] [PubMed] [Google Scholar]
- 13.Shalita AR. Genesis of free fatty acids. Journal of Investigative Dermatology. 1974;62(3):332–335. doi: 10.1111/1523-1747.ep12724289. [DOI] [PubMed] [Google Scholar]
- 14.Perisho K, Wertz PW, Madison KC, Stewart ME, Downing DT. Fatty acids of acylceramides from comedones and from the skin surface of acne patients and control subjects. Journal of Investigative Dermatology. 1988;90(3):350–353. doi: 10.1111/1523-1747.ep12456327. [DOI] [PubMed] [Google Scholar]
- 15.Wertz PW, Miethke MC, Long SA, Strauss JS, Downing DT. The composition of the ceramides from human stratum corneum and from comedones. Journal of Investigative Dermatology. 1985;84(5):410–412. doi: 10.1111/1523-1747.ep12265510. [DOI] [PubMed] [Google Scholar]
- 16.Akamatsu H, Komura J, Miyachi Y, Asada Y, Niwa Y. Suppressive effects of linoleic acid on neutrophil oxygen metabolism and phagocytosis. Journal of Investigative Dermatology. 1990;95(3):271–274. doi: 10.1111/1523-1747.ep12484890. [DOI] [PubMed] [Google Scholar]
- 17.Akamatsu H, Niwa Y, Matsunaga K. Effect of palmitic acid on neutrophil functions in vitro. International Journal of Dermatology. 2001;40(10):640–643. doi: 10.1046/j.1365-4362.2001.01292.x. [DOI] [PubMed] [Google Scholar]
- 18.Graham GM, Farrar MD, Cruse-Sawyer JE, Holland KT, Ingham E. Proinflammatory cytokine production by human keratinocytes stimulated with Propionibacterium acnes and P. acnes GroEL. British Journal of Dermatology. 2004;150(3):421–428. doi: 10.1046/j.1365-2133.2004.05762.x. [DOI] [PubMed] [Google Scholar]
- 19.Kim J. Review of the innate immune response in acne vulgaris: activation of toll-like receptor 2 in acne triggers inflammatory cytokine responses. Dermatology. 2005;211(3):193–198. doi: 10.1159/000087011. [DOI] [PubMed] [Google Scholar]
- 20.Gabay C. Interleukin-6 and chronic inflammation. Arthritis Research and Therapy. 2006;8(supplement 2):p. S3. doi: 10.1186/ar1917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Salamon M, Sysa-Jedrzejowska A, Lukamowicz J, Lukamowicz M, Swiatkowska E, Wozniacka A. Concentration of selected cytokines in serum of patients with acne rosacea. Przeglad Lekarski. 2008;65(9):371–374. [PubMed] [Google Scholar]
- 22.Kim BH, Roh E, Lee HY, et al. Benzoxathiole derivative blocks lipopolysaccharide-induced nuclear factor-κB activation and nuclear factor-κB-regulated gene transcription through inactivating inhibitory κB kinase β . Molecular Pharmacology. 2008;73(4):1309–1318. doi: 10.1124/mol.107.041251. [DOI] [PubMed] [Google Scholar]
- 23.Vowels BR, Yang S, Leyden JJ. Induction of proinflammatory cytokines by a soluble factor of Propionibacterium acnes: implications for chronic inflammatory acne. Infection and Immunity. 1995;63(8):3158–3165. doi: 10.1128/iai.63.8.3158-3165.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kang S, Cho S, Chung JH, Hammerberg C, Fisher GJ, Voorhees JJ. Inflammation and extracellular matrix degradation mediated by activated transcription factors nuclear factor-κB and activator protein-1 in inflammatory acne lesions in vivo. American Journal of Pathology. 2005;166(6):1691–1699. doi: 10.1016/s0002-9440(10)62479-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ottaviani M, Alestas T, Flori E, Mastrofrancesco A, Zouboulis CC, Picardo M. Peroxidated squalene induces the production of inflammatory mediators in HaCaT keratinocytes: a possible role in acne vulgaris. Journal of Investigative Dermatology. 2006;126(11):2430–2437. doi: 10.1038/sj.jid.5700434. [DOI] [PubMed] [Google Scholar]
- 26.Tian D, Qiu Y, Zhan Y, et al. Overexpression of steroidogenic acute regulatory protein in rat aortic endothelial cells attenuates palmitic acid-induced inflammation and reduction in nitric oxide bioavailability. Cardiovascular Diabetology. 2012;11(article 144) doi: 10.1186/1475-2840-11-144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Staiger H, Staiger K, Stefan N, et al. Palmitate-induced interleukin-6 expression in human coronary artery endothelial cells. Diabetes. 2004;53(12):3209–3216. doi: 10.2337/diabetes.53.12.3209. [DOI] [PubMed] [Google Scholar]
- 28.Stoian I, Manolescu B, Atanasiu V, Lupescu O, Buşu C. IL-6—STAT-3—hepcidin: linking inflammation to the iron metabolism. Romanian Journal of Internal Medicine. 2007;45(3):305–309. [PubMed] [Google Scholar]
- 29.Atreya R, Neurath MF. Signaling molecules: the pathogenic role of the IL-6/STAT-3 trans signaling pathway in intestinal inflammation and in colonic cancer. Current Drug Targets. 2008;9(5):369–374. doi: 10.2174/138945008784221116. [DOI] [PubMed] [Google Scholar]
- 30.Neurath MF, Finotto S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine and Growth Factor Reviews. 2011;22(2):83–89. doi: 10.1016/j.cytogfr.2011.02.003. [DOI] [PubMed] [Google Scholar]
- 31.Ledebur HC, Parks TP. Transcriptional regulation of the intercellular adhesion molecule-1 gene by inflammatory cytokines in human endothelial cells. Essential roles of a variant NF-κb site and p65 homodimers. Journal of Biological Chemistry. 1995;270(2):933–943. doi: 10.1074/jbc.270.2.933. [DOI] [PubMed] [Google Scholar]
- 32.Chen CC, Manning AM. Transcriptional regulation of endothelial cell adhesion molecules: a dominant role for NF-κB. Agents and Actions Supplements. 1995;47:135–141. doi: 10.1007/978-3-0348-7343-7_12. [DOI] [PubMed] [Google Scholar]
- 33.Jeremy AHT, Holland DB, Roberts SG, Thomson KF, Cunliffe WJ. Inflammatory events are involved in acne lesion initiation. Journal of Investigative Dermatology. 2003;121(1):20–27. doi: 10.1046/j.1523-1747.2003.12321.x. [DOI] [PubMed] [Google Scholar]
- 34.Ndisang JF. Role of heme oxygenase in inflammation, insulin-signalling, diabetes and obesity. Mediators of Inflammation. 2010;2010 doi: 10.1155/2010/359732.359732 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Olszowski T, Baranowska-Bosiacka I, Gutowska I, Chlubek D. Pro-inflammatory properties of cadmium. Acta Biochimica Polonica. 2012;59(4):475–482. [PubMed] [Google Scholar]
- 36.Baeuerle PA, Baltimore D. Nf-κB: ten years after. Cell. 1996;87(1):13–20. doi: 10.1016/s0092-8674(00)81318-5. [DOI] [PubMed] [Google Scholar]
- 37.Sato M, Sawamura D, Shinsuke I, Yaguchi T, Hanada K, Hashimoto I. In vivo introduction of the interleukin 6 gene into human keratinocytes: induction of epidermal proliferation by the fully spliced form of interleukin 6, but not by the alternatively spliced form. Archives of Dermatological Research. 1999;291(7-8):400–404. doi: 10.1007/s004030050429. [DOI] [PubMed] [Google Scholar]
- 38.Elder JT, Sartor CI, Boman DK, Benrazavi S, Fisher GJ, Pittelkow MR. Interleukin-6 in psoriasis: expression and mitogenicity studies. Archives of Dermatological Research. 1992;284(6):324–332. doi: 10.1007/BF00372034. [DOI] [PubMed] [Google Scholar]
- 39.Grossman RM, Krueger J, Yourish D, et al. Interleukin 6 is expressed in high levels of psoriatic skin and stimulates proliferation of cultured human keratinocytes. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(16):6367–6371. doi: 10.1073/pnas.86.16.6367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Knaggs HE, Holland DB, Morris C, Wood EJ, Cunliffe WJ. Quantification of cellular proliferation in acne using the monoclonal antibody Ki-67. Journal of Investigative Dermatology. 1994;102(1):89–92. doi: 10.1111/1523-1747.ep12371738. [DOI] [PubMed] [Google Scholar]
- 41.Hughes BR, Morris C, Cunliffe WJ, Leigh IM. Keratin expression in pilosebaceous epithelia in truncal skin of acne patients. British Journal of Dermatology. 1996;134(2):247–256. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure: Effect of NF-κB inhibitor PDTC on HaCaT cell viability and PA–induced NF-κB activation. (A) HaCaT cells were untreated or exposed to PDTC (10, 50, 100 μmol/L) for 1 hour. Cell viability was measured by a CCK-8 assay kit. Data are expressed as mean ± standard deviation (n = 5). ∗P < 0.05 compared with control. (B) HaCaT cells were untreated or exposed to 0.15 mM palmitic acid (PA) for 24 hours, either without or with pretreatment for 1 hour with 10 μmol/L PDTC or 0.1% dimethylsulfoxide (DMSO, as vehicle control). IKKα and IκBα expression levels were measured by western blotting. Each experiment was done in triplicate. Data are mean ± standard deviation. ∗P < 0.05 compared with PA.