

ABSTRACT
One of the greatest threats to the conservation of wild cat populations may be dogs or, at least, one of their viruses. Canine distemper virus (CDV), a single-stranded RNA virus in the Paramyxoviridae family and genus Morbillivirus, infects and causes disease in a variety of species, not just canids. An outbreak of CDV in wild lions in the Serengeti, Tanzania, in 1994 was a wake-up call for conservationists, as it demonstrated that an infectious disease could swiftly impact a previously healthy felid population. To understand how this virus causes disease in noncanid hosts, researchers have focused on specific mutations in the binding site of the CDV hemagglutinin gene. Now, Seimon et al. provide information on CDV in its latest feline victim, the endangered wild Amur tiger (Panthera tigris altaica) [T. A. Seimon et al., mBio 4(4):e00410-13, 2013, doi:10.1128/mBio.00410-13]. Their findings of CDV strains infecting tigers, in combination with recent information from other felids, paints a different picture, one in which CDV strains from a variety of geographic lineages and with a variety of amino acid residues in the hemagglutinin gene binding site can infect cats and cause disease. Although CDV has been known as a multihost disease since its discovery in domestic dogs in 1905, perhaps it is time to reconsider whether these noncanid species are not just incidental or “spillover” hosts but, rather, a normal part of the complex ecology of this infectious disease.
Commentary
Despite its name, canine distemper virus (CDV) is known to infect a variety of species in the order Carnivora (1). More recently, even nonhuman primates have been fatally infected with CDV, raising concern over potential human infection (2). Felids were thought to be mostly resistant to CDV until a series of fatal infections occurred in captive tigers, lions, leopards, and jaguars living in zoological parks in the United States (3). Then, in 1994, approximately one-third of the lion population in the Serengeti died or disappeared. Their deaths, as well as deaths in a variety of carnivores within the Serengeti ecosystem, were attributed to CDV (4). Since then, there have been cases or outbreaks in other populations of free-ranging felids, such as bobcat, Canada lynx, Eurasian lynx, the critically endangered Iberian lynx, and now Amur tigers (5–8). What causes this “spillover” into novel hosts has been an area of increasing research interest.
The signaling lymphocyte activation molecule (SLAM) is the common receptor used by morbilliviruses to gain entry into immune cells. Amino acid variations in both SLAM and the CDV hemagglutinin (HA) protein that binds SLAM are thought to be important in species specificity. Mutations, particularly at amino acid residues 530 and 549 of the HA protein, may account for variations in the infectivity and pathogenicity of different CDV strains in novel hosts (9). In vitro experiments using Vero cells that expressed lion SLAM had increased syncytia when exposed to CDV HA proteins with histidine at residue 549 (549H) compared to the results for exposure to CDV HA proteins with 549Y (10), suggesting that CDV strains that express 549H are more “generalist” pathogens. However, analysis of the CDV HA gene in CDV isolated from a wide variety of canids and noncanids indicated that strains isolated from the noncanids tested (including animals from the families Felidae, Mustelidae, Ailuridae, Ursidae, and Procyonidae within the order Carnivora) showed no particular bias to 549Y or 549H (11). A closer look at known pathogenic CDV strains from captive and wild felids, including those just described from the Amur tiger, demonstrate that both Y and H amino acids occur at this residue (Fig. 1). Similarly, there is no apparent association between the amino acid residue at another putative critical site, 530G/E, in strains affecting felids (Fig. 1). Despite variations from domestic dog SLAM at 5 of the 21 amino acid residues involved in viral binding, Vero cells expressing domestic cat SLAM demonstrated cytopathic effects and syncytia similar to those in cells expressing domestic dog SLAM when they were exposed to canid and mustelid Asia-1 strains and the Onderstepoort strain of CDV (12). Therefore, additional factors may be more important than SLAM or the H protein in determining the infectivity and pathogenicity of CDV strains in various hosts.
FIG 1 .
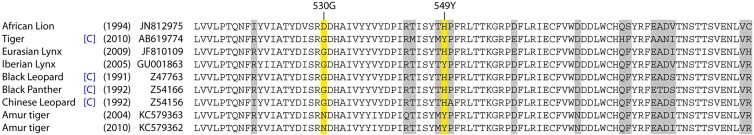
Comparison of amino acid sequences from positions 510 to 597 of the hemagglutinin protein of canine distemper virus isolates from felids (accession numbers JN812975, AB619774, JF810109, GU001863, Z47763, Z54166, Z54156, KC579363, and KC579362). Isolates from captive animals are denoted by [C], and the date in parentheses is the date of sampling. Variable residues are highlighted (gray), and sites 530G and 549Y, previously thought to be important for spillover into noncanid hosts, are highlighted yellow.
CDV infection is not universally fatal in felids. There is serologic evidence of CDV infection in wild felids throughout their natural habitats worldwide, including in South America, where fatal cases have not (yet) been reported (13–17). Retrospective serosurveys of the Serengeti lion population identified multiple “silent” CDV epidemics that were not associated with mortalities, suggesting that CDV infection was not always fatal and was present in the ecosystem long before 1994 (18). After a second high-mortality outbreak occurred in the adjacent Ngorongoro Crater lion population in 2001, comparison of the 1994 and 2001 outbreaks led to the finding that hemoparasitism was a contributing mortality factor. The levels of the tick-borne hemoparasite Babesia correlated with mortality rates such that lion prides with the highest levels of Babesia suffered the highest mortalities (>67%) (18). However, CDV infection has caused death in felids in the absence of coinfections, primarily due to interstitial pneumonia and encephalitis (6, 8, 19). The reasons for this apparent variation in pathogenicity are not known.
The question of how Amur tigers are exposed to CDV remains to be fully addressed. Because CDV spreads through aerosol droplets and contact with infected body fluids, predation is a possible means of infection for large felids. Tigers could be infected via exposure to unvaccinated domestic dogs, which are commonly implicated in wildlife CDV outbreaks. Habitat loss and poaching could lead to an increase in the number of human and dog incursions into tiger territories, but sick and old tigers, as well as individual tigers seeking new territories, are also known to trespass into human-dominated landscapes. It is possible that an increase in contact rates between wild and domestic carnivores could account for the emergence of CDV in tigers. However, CDV exposure has been noted in some felid populations that lack direct interaction with domestic dogs (20); hence, other mesocarnivores or peridomestic wild carnivores must also be considered a possible source of infection for apex predators.
Whether the virus can spread from tiger to tiger (e.g., point exposure and spread in the population) or if infection is more likely to occur from multiple spillover events in a spatial wave of multihost infection represents another unknown. The second hypothesis seems plausible due to the low number of endangered tigers, low contact rates between these territorial, solitary individuals, and the wide geographic and temporal distribution of cases (8). To disentangle these two hypotheses, one could use mathematical modeling approaches in combination with empirical data on the estimated densities, contact rates, CDV prevalence, and home ranges for tigers and conspecific carnivores in order to investigate challenging issues of CDV persistence and dynamics, as has been done in other ecosystems (21, 22). In regard to management strategies, if it turns out that a large multihost wild-carnivore community in the Russian Far East can maintain CDV, simply vaccinating domestic dogs will not protect endangered tigers from the threat of infection. Other options might include direct vaccination of tigers, but vaccinating secretive rare felids is no simple task either.
As researchers continue to search for molecular signatures and ecological factors that characterize fatal CDV infections in wild felids, all of the above begs the question: Has CDV “emerged” as a new pathogen for felids or have felids always been susceptible to CDV? Are the increased numbers of cases due to better detection methods or more-efficient monitoring of felids of conservation concern? For instance, a retrospective survey of captive-held large felids identified cases back to 1972, suggesting that CDV infection in cats was older and more widespread than previously thought (23). In contrast, serologic data from the Russian Far East suggest that Amur tigers lacked exposure to CDV prior to 2001, although the sample numbers were relatively small (14). Population monitoring of wild animals is challenging in general and even more challenging for the detection of illness or death due to disease. Hundreds of lions disappeared in the Serengeti ecosystem in the 1994 outbreak, but less than 1% of the population was either seen with signs or retrieved as carcasses. In the 2001 Ngorongoro Crater outbreak, 40% of the lion population disappeared, but scientists retrieved only 2 carcasses despite a dedicated lion research project and high tourist activity. The Crater is a much smaller and more open area than the Russian Far East, so on this score, the researchers studying the Amur tiger should be commended for their diligence in monitoring the population and extracting valuable information from the 5 carcasses presented in this study. Most felids are not as intensively monitored (nor as physically large) as the Amur tiger or Serengeti lion. It is easy to see how a solitary, elusive, and smaller wild felid could be infected from an unknown source, develop CDV, and die without detection. Irrespective of whether CDV in felids is a new phenomenon due to viral changes and changing ecological dynamics or whether it has been occurring all along, Seimon et al. (8) have shown that, once again, canine distemper virus is an important threat to the conservation of wild felids.
The views expressed in this Commentary do not necessarily reflect the views of the journal or of ASM.
Footnotes
Citation Terio KA, Craft ME. 2013. Canine distemper virus (CDV) in another big cat: should CDV be renamed carnivore distemper virus? mBio 4(5):e00702-13. doi:10.1128/mBio.00702-13.
REFERENCES
- 1. Williams ES. 2001. Canine distemper, p 50–59 In Williams ES, Barker IK. (ed), Infectious diseases of wild mammals, 3rd ed. Iowa State Press, Ames, IA [Google Scholar]
- 2. Sakai K, Yoshikawa T, Seki F, Fukushi S, Tahara M, Nagata N, Ami Y, Mizutani T, Kurane I, Yamaguchi R, Hasegawa H, Saijo M, Komase K, Morikawa S, Takeda M. 2013. Canine distemper virus associated with a lethal outbreak in monkeys can readily adapt to use human receptors. J. Virol. 87:7170–7175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Appel MJ, Yates RA, Foley GL, Bernstein JJ, Santinelli S, Spielman LH, Miller LD, Arp LH, Anderson M, Barr M, Pearce-Kelling S, Summers BA. 1994. Canine distemper virus epizootic in lions, tigers, and leopards in North America. J. Vet. Diagn. Invest. 6:277–288 [DOI] [PubMed] [Google Scholar]
- 4. Roelke-Parker ME, Munson L, Packer C, Kock R, Cleaveland S, Carpenter M, O’Brien SJ, Pospischil A, Hofmann-Lehmann R, Lutz H, Mwamengele GL, Mgasa MN, Machange GA, Summers BA, Appel MJ. 1996. A canine distemper virus epidemic in Serengeti lions (Panthera leo). Nature 379:441–445 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Daoust PY, McBurney SR, Godson DL, van de Bildt MW, Osterhaus AD. 2009. Canine-distemper virus-associated encephalitis in free-living lynx (Lynx canadensis) and bobcats (Lynx rufus) of eastern Canada. J. Wildl. Dis. 45:611–624 [DOI] [PubMed] [Google Scholar]
- 6. Origgi FC, Plattet P, Sattler U, Robert N, Casaubon J, Mavrot F, Pewsner M, Wu N, Giovannini S, Oevermann A, Stoffel MH, Gaschen V, Segner H, Ryser-Degiorgis MP. 2012. Emergence of canine distemper virus strains with modified molecular signature and enhanced neuronal tropism leading to high mortality in wild carnivores. Vet. Pathol. 49:913–929 [DOI] [PubMed] [Google Scholar]
- 7. Meli ML, Simmler P, Cattori V, Martínez F, Vargas A, Palomares F, López-Bao JV, Simón MA, López G, León-Vizcaino L, Hofmann-Lehmann R, Lutz H. 2010. Importance of canine distemper virus (CDV) infection in free-ranging Iberian lynxes (Lynx pardinus). Vet. Microbiol. 146:132–137 [DOI] [PubMed] [Google Scholar]
- 8. Seimon TA, Miquelle DG, Chang TY, Newton AL, Korotkova I, Ivanchuk G, Lyubchenko E, Tupikov A, Slabe E, McAloose D. 2013. Canine distemper virus: an emerging disease in wild endangered Amur tigers (Panthera tigris altaica). mBio 4(4):e00410-13. 10.1128/mBio.00410-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. McCarthy AJ, Shaw MA, Goodman SJ. 2007. Pathogen evolution and disease emergence in carnivores. Proc. Biol. Sci. 274:3165–3174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Nikolin VM, Osterrieder K, von Messling V, Hofer H, Anderson D, Dubovi E, Brunner E, East ML. 2012. Antagonistic pleiotropy and fitness trade-offs reveal specialist and generalist traits in strains of canine distemper virus. PLoS One 7:e50955. 10.1371/journal.pone.0050955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Nikolin VM, Wibbelt G, Michler FU, Wolf P, East ML. 2012. Susceptibility of carnivore hosts to strains of canine distemper virus from distinct lineages. Vet. Microbiol. 156:45–53 [DOI] [PubMed] [Google Scholar]
- 12. Hara Y, Suzuki J, Noguchi K, Terada Y, Simoda J, Mizuno T, Maeda K. 26 May 2013, posting date Function of feline signalling lymphocyte activation molecule as a receptor of canine distemper virus. J. Vet. Med. Sci. [DOI] [PubMed] [Google Scholar]
- 13. Munson L, Terio KA, Ryser-Degiorgis MP, Lane EP, Courchamp F. 2010. Wild felid diseases: conservation implications and management strategies, p 237–259 In MacDonald DW, Loveridge AJ. (ed), Biology and conservation of wild felids. Oxford University Press, Oxford, United Kingdom [Google Scholar]
- 14. Goodrich JM, Quigley KS, Lewis JC, Astafiev AA, Slabi EV, Miquelle DG, Smirnov EN, Kerley LL, Armstrong DL, Quigley HB, Hornocker MG. 2012. Serosurvey of free-ranging Amur tigers in the Russian Far East. J. Wildl. Dis. 48:186–189 [DOI] [PubMed] [Google Scholar]
- 15. Furtado MM, de Ramos Filho JD, Scheffer KC, Coelho CJ, Cruz PS, Ikuta CY, de Almeida Jácoma AT, de Oliveira Porfírio GE, Silveira L, Sollmann R, Tôrres NM, Neto JS. 2013. Serosurvey for selected viral infections in free-ranging jaguars (Panthera onca) and domestic carnivores in Brazilian cerrado, pantanal and Amazon. J. Wildl. Dis. 49:510–521 [DOI] [PubMed] [Google Scholar]
- 16. Uhart MM, Rago MV, Marull CA, Ferreyra Hdel V, Pereira JA. 2012. Exposure to selected pathogens in Geoffroy’s cats and domestic carnivores from central Argentina. J. Wildl. Dis. 48:899–909 [DOI] [PubMed] [Google Scholar]
- 17. Nava AF, Cullen L, Jr, Sana DA, Nardi MS, Filho JD, Lima TF, Abreu KC, Ferreira F. 2008. First evidence of canine distemper in Brazilian free-ranging felids. Ecohealth 5:513–518 [DOI] [PubMed] [Google Scholar]
- 18. Munson L, Terio KA, Kock R, Mlengeya T, Roelke ME, Dubovi E, Summers B, Sinclair AR, Packer C. 2008. Climate extremes promote fatal co-infections during canine distemper epidemics in African lions. PLoS One 3:e2545. 10.1371/journal.pone.0002545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Nagao Y, Nishio Y, Shiomoda H, Tamaru S, Shimojima M, Goto M, Une Y, Sato A, Ikebe Y, Maeda K. 2012. An outbreak of canine distemper virus in tigers (Panthera tigris): possible transmission from wild animals to zoo animals. J. Vet. Med. Sci. 74:699–705 [DOI] [PubMed] [Google Scholar]
- 20. Alexander KA, McNutt JW, Briggs MB, Standers PE, Funston P, Hemson G, Keet D, van Vuuren M. 2010. Host pathogens and carnivore management in southern Africa. Comp. Immunol. Microbiol. Infect. Dis. 33:249–265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Almberg ES, Cross PC, Smith DW. 2010. Persistence of canine distemper virus in the greater Yellowstone ecosystem’s carnivore community. Ecol. Appl. 20:2058–2074 [DOI] [PubMed] [Google Scholar]
- 22. Craft ME, Volz E, Packer C, Meyers LA. 2009. Distinguishing epidemic waves from disease spillover in a wildlife population. Proc. Biol. Sci. 276:1777–1785 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Myers DL, Zurbriggen A, Lutz H, Pospischil A. 1997. Distemper: not a new disease in lions and tigers. Clin. Diagn. Lab. Immunol. 4:180–184 [DOI] [PMC free article] [PubMed] [Google Scholar]