

Abstract
Molecular automata are mixtures of molecules that undergo precisely defined structural changes in response to sequential interactions with inputs1–4. Previously studied nucleic acid-based-automata include game-playing molecular devices (MAYA automata3,5) and finite-state automata for analysis of nucleic acids6 with the latter inspiring circuits for the analysis of RNA species inside cells7,8. Here, we describe automata based on strand-displacement9,10 cascades directed by antibodies that can analyze cells by using their surface markers as inputs. The final output of a molecular automaton that successfully completes its analysis is the presence of a unique molecular tag on the cell surface of a specific subpopulation of lymphocytes within human blood cells.
The problem of labeling a narrow subpopulation within a much larger population of related cells occurs often because of the need to specifically tag a particular cell type for the purpose of elimination11, analysis and isolation12, or imaging13. The problem could be readily addressed in a direct manner if targeted subpopulations could have some unique cell-surface marker13 against which antibodies can be raised. However, as best illustrated through an example of a cancer therapy utilizing antibody-drug conjugates (ADCs), markers are most often shared by non-targeted cells, leading, in this case, to off-target toxicities13. In order to uniquely target cells that do not have any distinctive marker on their surfaces, we need to use a set of multiple markers for each subpopulation in a Boolean manner. Molecular automata with structural changes (“state transitions”) coupled to the sequential recognition of a selected set of cell surface markers might be able to contract the set into a single tag and thus provide a unique handle for the targeted cells. Or, in the language of molecular computing14,15, these molecular devices would autonomously, i.e., without any human participation, evaluate Boolean functions on cell surfaces with surface markers as inputs and a tag as an output.
We chose to utilize blood cells as targets for molecular automata, because these are the most exhaustively studied examples of cells16 with lineages and stages of differentiation defined by the presence or absence of multiple cell-surface markers. They are commonly characterized by flow cytometry via different levels of expression of multiple cell surface markers14 known as Clusters of Differentiation or CDs, with CD45, CD20, CD3, and CD8 used as examples in this work. We show in Fig. 1 the basic design principles for automata that will tag lymphocytes with targeted CD markers characteristic for B-cells, i.e., CD45+CD20+ cells, in the presence of non-targeted CD45+CD20− cells (e.g., CD45+CD3+, T-cells).
Figure 1. Design considerations for automata operating on cell surfaces.

a, Scheme of automata operating on a B-cell with C45+CD20+ phenotype (target), and on an example of a non-targeted cell with CD45+CD20− phenotype, e.g., T-cell. Oligonucleotide components (coloured horizontal lines) attached to antibodies (Y-shaped structures) are brought together on some cells and not others (for example, αCD45-1●2 and αCD20-3●4 are together only on B cells), leading to a cascade of oligonucleotide transfers driven by an increase in complementarity. The transfers result in a unique single stranded oligonucleotide 4 being displayed only on targeted cells; b, Scheme of a typical strand displacement reaction used in the automata: 0 + 1●2 + 3●4 → 0●1 + 2●3 + 4, controlled via a sequential exposure of toeholds (T1 then T3): single-stranded oligonucleotide 0 displaces oligonucleotide 2 from its complex with 1 via toehold interactions, that is, stronger complementarity and kinetic enablement due to the additional complementarity with exposed T1. This generates an oligonucleotide stretch in strand 2 complementary to a toehold T3 in strand 3 that can extend the reaction cascade by displacing oligonucleotide 4 from 3●4; this in turn generates the next oligonucleotide stretch complementary to toehold T5, that can be used to extend the cascade to 5●6 (not shown) and so on (as indicated by double dotted arrows) or label the cell with 5 carrying fluorescein. Without T3, the cascade stops. c, An example of oligonucleotide sequences used in the automata. Complete oligonucleotide sequences are available in Supplementary Materials.
The exact “program” (i.e., conditional sequential transitions) that the automata will execute on the surfaces of lymphocytes, will be defined by sets of antibodies against CD markers Mi which direct the cascade (see Fig. 1 with CD45 and CD20 as Mi’s). We started with the well-established off-the-shelf antibodies targeting CD markers: αCD45; αCD45RA; αCD20 (Rituximab); αCD3; and αCD8. All of these antigens are present at 80,000–200,000 copies per cell surface on targeted subpopulations of lymphocytes, ensuring a strong signal when measured by flow cytometry. These antibodies were conjugated17 with a set of partially complementary oligonucleotides (1●2, 3●4, and 5●6) optimized to execute modified strand-displacement cascades9 when triggered with oligonucleotide 0 (Fig. 1b,c, Supplementary Fig. S1–S6). Once turned on, the automata based on these antibody conjugates would perform ‘if yesMi then proceed’ or‘if notMi then proceed’ assessments of Mi markers on the surface of individual cells via oligonucleotide transfers enabled by sequential exposure of new toeholds (cf. Figure 1b) and driven by the formation of more strongly complementary oligonucleotides (0●1, 2●3, and 4●5).
The first step in demonstrating automata is to test their ability to evaluate two surface markers (see Fig. 2a for yesCD45yesCD20 experiment, functionally equivalent to Boolean CD45andCD20) and to selectively label one targeted subpopulation within a population of peripheral blood mononuclear cells (PBMCs). We constructed all possible automata that could assess combinations of two out of three markers, CD45 (a marker of nucleated hematopoietic cells), CD20 (a B-cell marker), and CD3 (a pan-T-cell marker). Two of these automata are capable of successful completion of their program: yesCD45yesCD20 would operate (label) only on B-cells (Fig. 2a) and yesCD45yesCD3 would operate only on T-cells (Supplementary Fig. S7). The third possible two-step automaton, yesCD3yesCD20 is a negative control, because no subpopulation displays these two markers at the same time (Supplementary Fig. S7c). The operation of these automata is equivalent to asking: “Is this cell a nucleated hematopoietic cell?” (yesCD45) followed by, in the case of the first automaton, “Is this a nucleated hematopoietic cell from a B-cell lineage?” (yesCD20) and, in the case of the second automaton, “Is this nucleated hematopoietic cell from the T-cell lineage?” (yesCD3). In all these automata, if both questions are answered positively in a row, the reaction performed, on an example of B-cells, will be: 0 + 1●2αCD45 + 3●4αCD20 → 0●1 + αCD452●3 + αCD204, with targeted subpopulations displaying a newly uncovered single-stranded oligonucleotide, 4. This one marker then contains the same information as traditional multicolor labeling with the same antibodies that we used in construction of automata and that we would otherwise use to characterize the immunological phenotype of these cells (e.g., as CD45+CD20+). Additionally, we set up a system so that the output oligonucleotide (4) would interact with a solution-phase label as in: αCD204 + 5●6 → αCD204●5 + 6 (where 5 is labeled with fluorescein), thus allowing direct analysis by flow cytometry of the response of targeted cells within a heterogeneous population of cells to the cascade. In order to assess the full operation of automata, we labeled 1 with Cy5, so both its removal and subsequent acquisition of fluorescein by αCD204, on the cell surface, could be monitored simultaneously in real time.
Figure 2. Demonstration of automata assessing the presence of two cell surface markers.

a, Scheme of yesCD45yesCD20 automata implementing reaction cascade 0 + 1●2αCD45 + 3●4αCD20 + 5●6 → 0●1 + αCD452●3 + αCD204●5 + 6 on the cell surface; 1 is labeled with Cy5 and 0 with a quencher (QCy5 = Iowa Black RQ) for Cy5; 5 is labeled with fluorescein (F), and 6 labeled with a quencher (QF = Iowa Black FQ) for fluorescein. b, Flow cytometry monitoring of the yesCD45yesCD20 cascade (each dot represents the fluorescence signal level from a single cell at the time of measurement, with dot density representing the number of cells, shown as increasing from blue-through-red). Time course of the cascade reaction on CD20+ B-cells; Left panel: monitoring the removal of Cy5-1 occurring on CD45+ cells after the triggering reaction with 0 ; Right panel: Fluorescein-labeled 5 is taken up from solution by CD20+ B-cells – this is used for monitoring the acquisition of 5 by 4 enabled by prior removal of 3 from 4. The addition of 5●6 (indicated by first red arrow) produces an immediate fluorescence increase on all cells due to incomplete quenching of fluorescein; the addition of 0 (indicated by the second red arrow) triggers the cascade and separation of the subpopulations of cells. c, Monitoring of a cascade on individual subpopulations within PBMCs using fluorescently labeled monoclonal antibodies targeting the same CDs but different non-overlapping epitopes for identification of cell subpopulations (PerCP-CD45 antibody, clone 2D1 and Pacific Blue-CD20 antibody, clone 2H7). These results confirm that all CD45+CD20+ cells (right gate, i.e., right box on bottom left panel) are labeled by automata (i.e., there is an increase in fluorescein uptake from solution, cf. bottom middle and right panels) and that cells that are CD45+CD20− (left gate, i.e., left box on bottom left panel) are not (upper left and central panels). It is seen that ~0.5% of the cells that are gated (box at the central up panel) as CD45+CD20− may react with a delay (upper right panel). Red arrows have the same meaning as in b.
In our experiments, the first two automata successfully labeled only surfaces of either B-(CD45+CD20+) or T-(CD45+CD3+) cells (Fig. 2a and b and Supplementary Fig. S7, S8), with each outcome confirmed three or more times on individual human blood samples and monitored by multicolor flow cytometry. From these same components we also made an automaton that could label the surfaces of both B- and T-cells by using 3●4αCD20 and 3●4αCD3 in the same solution (cf., Supplementary Fig. S7e); a possible presentation of this automaton is the demonstration of an or function, as in yesCD45(yesCD20oryesCD3). In control experiments, we also confirmed that the automata worked on enriched cell subpopulations with correct marker combinations (B- or T-cells).
We then studied in more detail negative controls, that is, antibody directed cascades that could occur only between markers on separate cells (between two subpopulations). Using the third possible two-step automaton introduced above, yesCD3yesCD20, we observed no labeling within the time-frame of our experiment, indicating that the T-cells did not visibly exchange elements with B-cells either through diffusion or through direct physical contact of cells (Supplementary Fig. S9). We also separated T- and B-cells, labeling the former with 1●2−αCD3, the latter with 3●4−αCD20. Upon remixing the cells, we observed no crosstalk between different lineages, within the detection limits of the flow cytometer (Supplementary Fig. S9). Finally, we demonstrated that automata yesCD20yesCD45, with the inverted order of assessing the cell, worked without labeling any CD45+CD20− cells (Supplementary Fig. S10). All of these experiments demonstrate low noise in the automata in the absence of an excess of elements in the solution-phase (i.e., they demonstrate minimal tagging of cells via diffusion or by direct contact between cells). In order to estimate the effects of washing away excess of antibody conjugates, we studied automata yesCD3yesCD20 and yesCD3(yesCD20ORyesCD8) without prior removal of the excess components from the solution. In both cases we observed changes in the fluorescence of non-target cells, albeit several-fold weaker than in the case of targeted cells (Supplementary Fig. S9), indicating that proximity-based interactions on a single cell were dominant.
The structures comprising these two-step automata were adjusted to enable labeling cells with fluorescent oligonucleotides only in the absence of a CD marker, that is, an ‘if notMi then proceed’ function (notMi, Fig. 3, Supplementary Fig. S11). During the differentiation of T-cells, from naïve to memory, there is a transition in expression of two different isoforms of CD45, CD45RA and CD45RO, and we created an automaton assessing the presence of isoforms of CD45 on CD8+ T-Cells, with the CD45RA isoform inhibiting the cascade. The automaton yesCD8notCD45RA consisted of 3●4αCD8 and 5*●6*αCD45RA triggered by 2 in the presence of solution-phase 5●6 (where 5 is labeled with fluorescein). All cells that responded to the automaton and acquired 5 from solution-phase strongly expressed CD45RO, that is, they were CD45RA− cells (Fig. 3b, Supplementary Fig. S11b). This was in contrast with CD8+CD45RA+ T-cells, namely CD45RO− or CD45ROdim, which were hindered in acquiring 5 due to competition with 5* from CD45RA in proximity to CD8-displaying 4, instead forming 5*●4αCD8 (Fig. 3, Supplementary Fig. S11). It should be noted that the ‘if notMi then proceed’ function is limited, until a threshold9 function is introduced, by the ratio of levels of expression of individual markers on the cell surface.
Figure 3. Demonstration of automata assessing the absence of a cell surface marker.
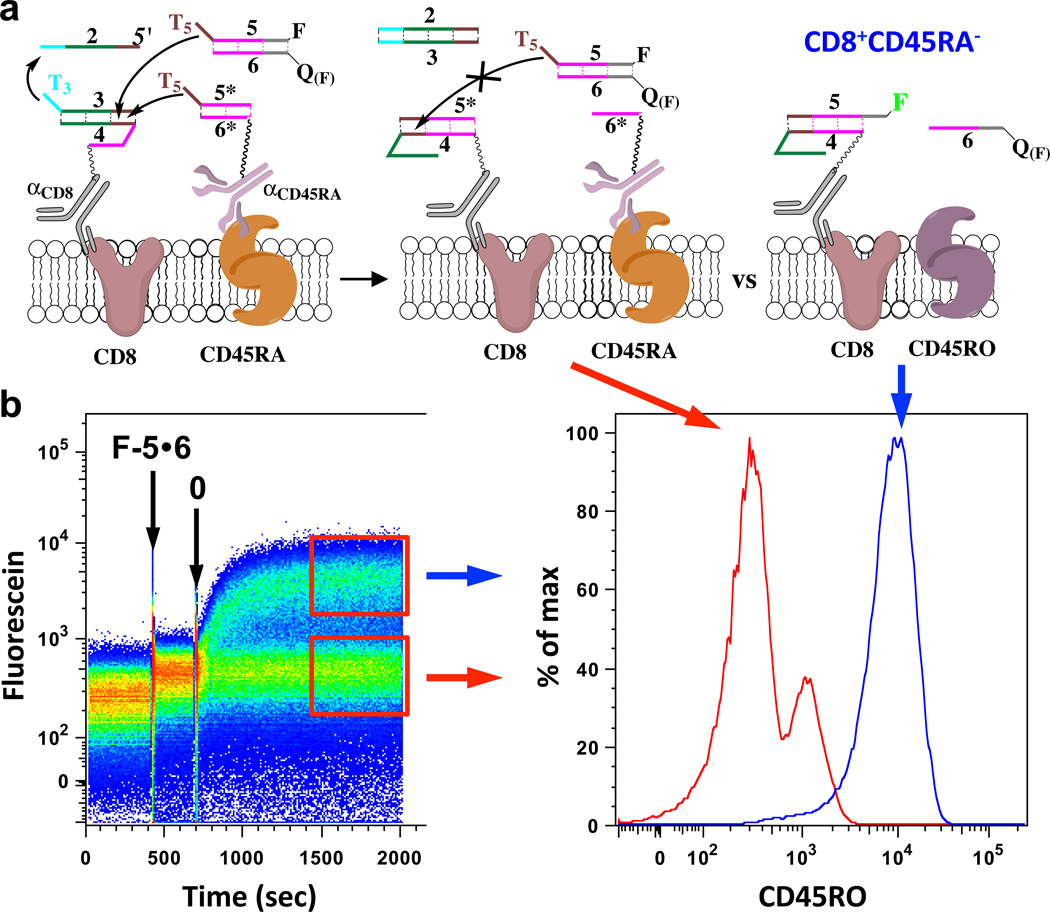
a, Scheme of a yesCD8notCD45RA cascade protecting naïve CD8+CD45RA+ T-cells; in these cells the CD45RA isoform prevents the targeting of CD8. This automaton works by: if cell is CD8 positive and if cell is CD45RA positive (or CD45ROneg) then reaction is 2 + 3●4αCD8 + 5*●6*αCD45RA + 5●6 → 2●3 + 5*●4αCD8 + 6*αCD45RA + 5●6, resulting in no labeling (red trace on right panel of b), else, when cell is CD45RA− (CD45RO isoform), the reaction is: 2 + 3●4αCD8 + 5●6 → 2●3 + 5●4αCD8 + 6; as a result, fluorescein-labeled 5 is removed from quencher (QF = Iowa Black FQ) strand 6 and is taken up from the solution in a simple yesCD8 response (blue trace on right panel of b). b, Monitoring of the yesCD8notCD45RA cascade: Left panel shows time-course of cascade reaction on the surface of CD8+ T-cells from peripheral blood: Right panel, histograms (or frequency distributions) of memory CD8+ T-cells responding to automata (upper gate/box on left panel,; blue trace on right panel, CD8+CD45RO+ or CD45RA−) while naïve CD8+ T-cells are being protected from the automaton (lower gate/box on left panel; red trace, CD8+CD45RO−/CD45RA+). For gating strategy, please see Supplementary Fig. S11.
At this point, we had established three types of transitions that could be used to build larger automata, yesMi, notMi, and or (the last function consisting of adding to the cells two antibodies conjugated to identical oligonucleotide components). As an example of the feasibility of building more complex automata from these simple transitions, we proceeded to build an automaton with a three-step cascade, evaluating the presence of up to three markers, and executing on the cell surface yesCD45yesCD3yesCD8 (the third question: “Is this nucleated hematopoietic cell of T-Cell lineage a CD8 positive cell?”). In this automaton, the surface of CD8+ cells enabled the following reaction: 0 + 1●2αCD45 + 3●4αCD3 + 5●6αCD8 + 7●8 → 0●1 + αCD452●3 + αCD34●5 + αCD86●7 + 8. The labeling scheme allowed us to monitor each step in this cascade via flow cytometry in real time (Fig. 4,b, Supplementary Fig. S12). This automaton was successfully demonstrated on targeted cells, with changes in fluorescence on cells being consistent with changes in distances between various components upon each step in the cascade (the first step is monitored by the removal of Pacific Blue, second by the drop in Cy5 due to quenching, and third by the acquisition of fluorescein from solution).
Figure 4. Demonstration of automata assessing the presence of three markers (CD45, CD3 and CD8) on the surface of the cell.

a, Scheme of a yesCD45yesCD3yesCD8 cascade (individual antibody conjugates: αCD45 with 1●2 complex, αCD3 with 3●4, and αCD8 with 5●6). Oligonucleotides are labeled with fluorescent dyes and quenchers to facilitate monitoring of multiple events in parallel (1 with Pacific Blue or PB, 3 with Cy5, 7 with fluorescein or F; 0 with a quencher (Q(PB) = Iowa Black FQ) for Pacific Blue, 2 with a quencher (Q(Cy5) = Iowa Black RQ) for Cy5, and 8 with a quencher (Q(F) = Iowa Black FQ) for fluorescein;). b, Flow-cytometry monitoring in three colors of the state transition (y-axis: fluorescence intensity, x-axis: time, with arrows showing events, i.e., the additions of oligonucleotides 7●8 and 0. The first step is observed in the left panel, with pacific blue removed from the cell surface by 0; in the middle panel, we observe the second step, i.e., transfer of 3 to 2, by quenching of Cy5; the third step is, transfer of 5 to 4, is not observed directly, but by an uptake of 7 from a solution by 6 (right panel). In the final step (right panel - fluorescein), the separation of CD45+CD3+CD8+ from all other lymphocytes is clearly shown.
Finally, we decided to test our automata under conditions that could lead to applications. We were able to demonstrate: (1) that we could isolate with a purity equivalent to a standard isolation protocol, fluorescein-labeled cells after a yesCD45yesCD3 automaton; for this we used a standard method for the isolation of cells (Fig. 5a, using anti-fluorescein antibody conjugated to magnetic beads); and (2) that an automaton (we used yesCD3yesCD8) can function in whole blood such that we could simply add automata components to the mixture all together prior to triggering the reaction (Fig. 5b). The former demonstration was important, because it showed that there is no detectable decrease in purity of isolated cells between a single step automaton-based procedure (in situ cascade) and the standard separation protocol based on individual separation steps for each CD marker. The latter experiment also established that blood components did not interfere with the cascades. Together with demonstrations that interactions via solution-phase information transfer do not represent major pathways in labeling cells (vide supra and Supplementary Fig. S9) this experiment opens up the possibility of using automata for labeling and eventually eliminating cells in vivo, depending on the pharmacokinetic properties of our conjugates.
Figure 5. Demonstrations of a potential for practical applications.
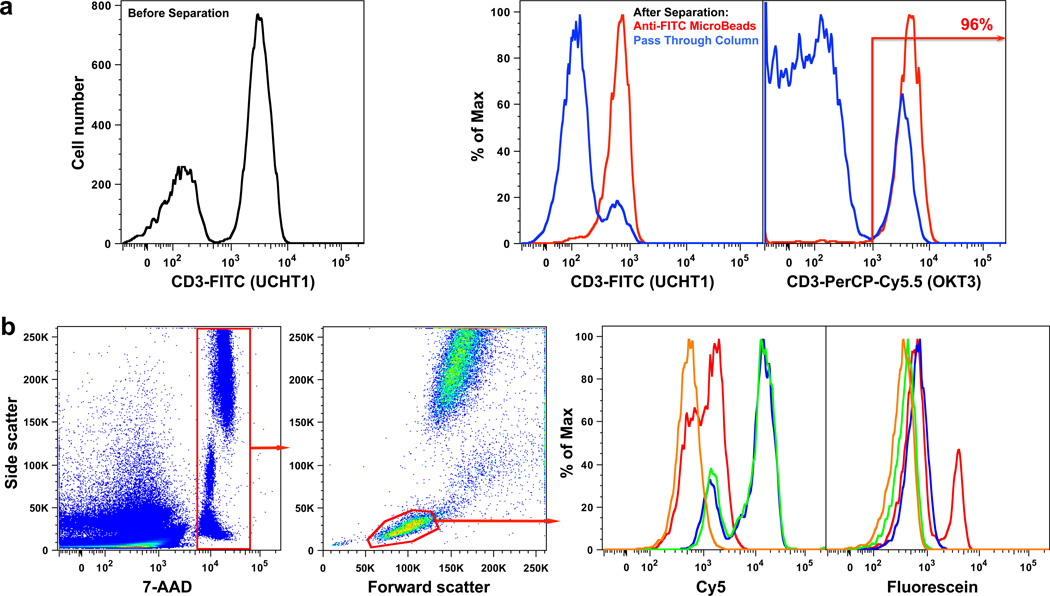
a, Magnetic separation of CD3+ cells from PBMCs based on results of a yesCD45yesCD3 automaton. Before the cascade (left panel), we see the mixture of cells with different CD3+ status. After the cascade, cells were incubated with magnetic microbeads conjugated with anti-FITC antibodies (Miltenyi Biotec) and applied on a MACS Column (Miltenyi Biotec; purity of the post-isolation preparation was >95%). The “pass-through” fraction (blue lines in middle panel) and the magnetically labeled cells (red lines in middle panel) were re-analyzed with different clones of αCD3antibodies (i.e., different epitopes of the same CD) to confirm purity (right panel). b, yesCD3yesCD8 automaton was examined in whole blood: Flow cytometry analysis with gating strategy shown (left and middle left panels); nucleated cells were gated by staining their DNA with 7-AAD, with lymphocytes selected based on forward and side scatter; The histograms show end points of two steps of the cascade as performed in blood, Cy5 fluorescence is used to show that the first step was accomplished, while fluorescein is used to demonstrate that the second step was accomplished, as in Figure 2). Lines on histograms: yellow – unlabeled blood sample; green – blood sample incubated for 15 min with αCD3 conjugated with duplex 1●2 and αCD8 conjugated with duplex 3●4; blue – same, but with 5●6 added and also incubated for 15 min; red – subsequent addition of 0 after incubation for 15 min.
In conclusion, we have established that a combination of antibodies and oligonucleotide-based reaction cascades can operate as molecular automata to assess the presence or absence of cell surface markers on living human cells. From the perspective of molecular automata, these results extend their use beyond the analysis by transfecting oligonucleotides into cells7,8, thereby permitting new operations on the surfaces of native cells. Unlike previous utilizations of proximity principles such as bispecific antibodies18 or proximity ligation reactions19 our approach can be readily expanded to more complex logic operations and to the protection of cells through a not transition. The demonstrated systems contribute as well to the emerging field of molecular robotics20–22. One approach to cell analysis with molecular robots is to increase the complexity of individual nanoparticles using self-assembly of DNA nanoobjects displaying multiple aptameric locks22. We offer here an alternative and potentially simpler method: we employ a number of elementary components that are brought together by the cell surface to execute more complex programmable (automata) functions, an approach that is conceptually similar to that of distributed robotics paradigms23.
Selected Methods
Detailed protocols, all sequences and their optimization, and full characterization of all synthesized reagents are provided in the Supplementary Material. Briefly: Oligonucleotides were coupled to antibodies, unless stated otherwise, in a two-step procedure: (i) DTT was used under conditions that reduce interchain disulfide bonds; (ii) 5’-end maleimide-functionalized oligonucleotides were coupled to the reduced antibody sulfhydryls, and the products were purified using gel filtration. One biotinylated antibody was used in the notCD45RA cascade, with streptavidin used to cross-link it to biotinylated oligonucleotides; this procedure was performed directly on cells without purification of conjugates (in this case negative controls with no streptavidin and no antibody were successfully run as well). Reagents were added to cell suspensions, and in all experiments involving PBMC’s reagents were removed from solution by centrifugation. In whole blood experiments (Fig. 5), reagents were left in blood to mimic the conditions for potential in vivo applications.
Supplementary Material
Acknowledgements
The research presented in this paper, as well as past attempts that eventually led to the current design, were supported by: NIH (R21CA128452, RC2CA147925, R21EB014477, RGM104960) to SR and MNS, NSF (CCF-0218262, CCF-0621600, ECCS-1026591, CBET-1033288), NASA (NAS2-02039), and a fellowship by the Lymphoma and Leukemia Foundation (CPD Award) to MNS. We thank Drs. John Loeb, Eric Meffre, and Darko Stefanovic for their advice and comments on the manuscript.
Footnotes
Author’s contributions: MR was the principal experimenter on cells, while ST carried out conjugations and optimized cascades in solution phase; PP performed exploratory experiments, AD and SK model experiments on beads. SR is the responsible author in charge of flow cytometry experiments, ST and MNS of non-cell-based experiments. MNS and VB designed and put early proposals for implementation of molecular computing on cell surfaces and settled on lymphocytes as targets. MR, ST, SR, and MNS analyzed the data; SR and MNS were the primary designers of the experiments and are most responsible for the structure of the presentation in this paper, while MNS wrote the initial draft of this manuscript.
Supplementary Materials
Materials and Methods
Figs. S1 to S12
Supplementary References
References
- 1.Rothemund P. A DNA and restriction enzyme implementation of Turing Machines. DNA based computers. DIMACS Series in Discrete Mathematics and Theoretical Computer Science. 1996;(27):75–120. [Google Scholar]
- 2.Benenson Y, Paz-Elitzur T, Adar R, Keinan E, Livneh Z, Shapiro E. Programmable and autonomous computing machine made of biomolecules. Nature. 2001;414:430–434. doi: 10.1038/35106533. [DOI] [PubMed] [Google Scholar]
- 3.Stojanovic MN, Stefanovic D. A deoxyribozyme-based molecular automaton. Nature Biotechnol. 2003;21:1069–1074. doi: 10.1038/nbt862. [DOI] [PubMed] [Google Scholar]
- 4.Wang Z-G, Elbaz J, Remacle F, Levine RD, Willner I. All DNA finite-state automata with finite memory. Proc. Nat. Ac. Sci. (USA) 2010;107:21996–22001. doi: 10.1073/pnas.1015858107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pei R, Matamoros E, Liu M, Stefanovic D, Stojanovic MN. Training a molecular automaton to play a game. Nature Nanotech. 2010;5(11):773–777. doi: 10.1038/nnano.2010.194. [DOI] [PubMed] [Google Scholar]
- 6.Benenson Y, Gil B, Ben-Dor U, Adar R, Shapiro E. An Autonomous Molecular Computer for Logical Control of Gene Expression. Nature. 2004;429:423–429. doi: 10.1038/nature02551. [DOI] [PubMed] [Google Scholar]
- 7.Rinaudo K, Bleris L, Maddamsetti R, Subramanian S, Weiss R, Benenson Y. A universal RNAi based logic evaluator that operates in mammalian cells. Nature Biotechnology. 2007;25:795–801. doi: 10.1038/nbt1307. [DOI] [PubMed] [Google Scholar]
- 8.Xie Z, Wroblewska L, Prochazka L, Weiss R, Benenson Y. Multi-input RNAi-based logic circuit for identification of specific cancer cells. Science. 2011;333:1307–1311. doi: 10.1126/science.1205527. [DOI] [PubMed] [Google Scholar]
- 9.Seelig G, Soloveichik D, Zhang DY, Winfree E. Enzyme-Free Nucleic Acid Logic Circuits. Science. 2006;314:1585–1588. doi: 10.1126/science.1132493. [DOI] [PubMed] [Google Scholar]
- 10.Qian L, Winfree E, Bruck J. Neural network computation with DNA strand displacement cascades. Nature. 2011;475:368–372. doi: 10.1038/nature10262. [DOI] [PubMed] [Google Scholar]
- 11.Weiner LM, Surana R, Wang S. Monocolonal antibodies: versatile platforms for cancer immunotherapy. Nature Rev. Immunology. 2010;10:317–327. doi: 10.1038/nri2744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Welte Y, Adjaye J, Lehrach HR, Regenbrecht CR. A. Cancer Stem cells in solid tumors, elusive or illusive? Cell Communication and Signaling. 2010;8:6. doi: 10.1186/1478-811X-8-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ichise M, Harris PE. Imaging of β-cell mass and function. J. Nucl. Med. 2010;51:1001–1004. doi: 10.2967/jnumed.109.068999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.De Silva AP, Uchiayam S. Molecular Logic and Computing. Nature Nanotechnology. 2007;2:399–410. doi: 10.1038/nnano.2007.188. [DOI] [PubMed] [Google Scholar]
- 15.Katz E, Privman V. Enzyme-based logic systems for information processing. Chem. Soc. Rev. 2010;39:1835–1857. doi: 10.1039/b806038j. [DOI] [PubMed] [Google Scholar]
- 16.Bendall SC, Simonds EF, Qiu P, Amir el-AD, Krutzik PO, Finck R, Bruggner RV, Melamed R, Trejo A, Ornatsky OI, Balderas RS, Plevritis SK, Sachs K, Pe’er D, Tanner SD, Nolan GP. Single-cell mass cytometry of differential immune and drug responses across a human hematopoietic continuum. Science. 2011;332(6030):687–696. doi: 10.1126/science.1198704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu H, Chumsae C, Gaza-Bulseco G, Hurkmans K, Radziejewski CH. Ranking the susceptibility of disulfide Bonds in Human IgG1 antibodies by reduction, differential alkylation, and LC-MS analysis. Analytical Chemistry. 2010;82:5219–5226. doi: 10.1021/ac100575n. [DOI] [PubMed] [Google Scholar]
- 18.Holmes D. Buy buy bispecific antibodies. Nature Rev. Drug Discovery. 2011;10:798. doi: 10.1038/nrd3581. [DOI] [PubMed] [Google Scholar]
- 19.Söderberg O, Gullberg M, Jarvius M, Ridderstråle K, Leuchowius K-J, Jarvius J, Wester K, Hydbring P, et al. Direct observation of individual endogenous protein complexes in situ by proximity ligation. Nature Methods. 2006;3(12):995–1000. doi: 10.1038/nmeth947. [DOI] [PubMed] [Google Scholar]
- 20.Douglas SM, Bachelet I, Church GM. A logic-gated nanorobot for targeted transport of molecular payloads. Science. 2012;335:831–834. doi: 10.1126/science.1214081. [DOI] [PubMed] [Google Scholar]
- 21.Gu H, Chao J, Xiao S-J, Seeman NC. A proximity-based programmable DNA nanoscale assembly line. Nature. 2010;465:202–205. doi: 10.1038/nature09026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lund K, Manzo AJ, Dabby N, Michelotti N, Johnson-Buck A, Nangreave J, Taylor S, Pei R, Stojanovic MN, Walter NG, Winfree E, Yan H. Molecular robots guided by prescriptive landscapes. Nature. 2010;13:206–210. doi: 10.1038/nature09012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Distributed Autonomous Robotic Systems: The 10th International Symposium; 2012. (Springer Tracts in Advanced Robotics) [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.