

Abstract
Various co-signaling molecules on T cells can contribute to activation, inhibition, or exhaustion, depending on context. The SLAM family receptor CD244 (2B4/SLAMf4) has been shown to be capable of either inhibitory or enhancing effects upon engagement of its ligand CD48 (SLAMf2). We examined phenotypes of CD8 T cells from HIV+ and HIVneg human donors, specific for HIV and/or respiratory syncytialvirus. Cultured and ex vivo CD8 T cells expressed PD-1, CD244, and TIM-3. We found that ex vivo CD8 T cells downregulated CD244 in response to superantigen. Furthermore cognate peptide induced rapid downregulation of both CD244 and TIM-3, but not PD-1, on CD8 T cell clones. CD244 downmodulation required simultaneous signaling via both TCR and CD244 itself. Using a pH-sensitive fluorophore conjugated to avidin-antibody tetramers, we found that CD244 crosslinking in the presence of TCR signaling resulted in rapid transport of CD244 to an acidic intracellular compartment. Downregulation was not induced by PMA-ionomycin, or prevented by PI3K inhibition, implicating a TCR-proximal signaling mechanism. CD244 internalization occurred within hours of TCR stimulation, and required less peptide than was required to induce IFNγ production. The degree of CD244 internalization varied among cultured CD8 T cell lines of different specificities, and correlated with the enhancement of IFNγ production in response to CD48 blockade in HIV+, but not HIVneg subjects. Our results indicate that rapid CD244 internalization is induced by a two-signal mechanism and plays a role in modulation of antiviral CD8 T cell responses by CD48-CD244 signaling.
Introduction
In order to protect against infections and tumors while avoiding autoimmune and inflammatory pathology, T cells are regulated by complex networks of activating and inhibitory costimulatory/co-signaling molecules. The activity of each co-signaling receptor is regulated by factors such as post-translational modification, expression of secondary signaling molecules, concentration of cognate ligands, and co-expression of other co-signaling receptors. Chronic viral infection can lead to progressive exhaustion of T cells. Exhausted T cells, including those found in HIV+ subjects, are defective in many antiviral functions and express progressively higher levels of markers including PD-1, CD160, LAG-3, TIM-3, and CD244 (1). In some cases, blockade of one or more of these receptors can reverse the effects of exhaustion and restore immune function (2), thus proving that PD-1, for example, is not only a marker but also a mediator of T cell dysfunction in chronic infection.
CD244 (2B4/SLAMf4) presents an interesting case of “dual functions”, as blockade of CD244 signaling has been shown to enhance or inhibit T cell responses in different contexts. The SLAM (Surface-receptor-signaling Lymphocytic Activation Molecule)2 family of receptors (3, 4) consists of eight type I transmembrane proteins and one GPI-linked receptor (CD48/SLAMf2). With the exceptions of CD48 and CD244, which in humans are thought to be the unique cellular ligands for each other, SLAM family receptors bind in a homophilic manner, such that each receptor is the unique cellular ligand for itself. SLAM family receptors play many roles in regulating the differentiation and effector functions of hematopoetic cells (3). Blockade of CD244-CD48 interactions in vitro has been variously shown to enhance or inhibit (5) antiviral T cell responses in the PBMC of subjects with different viral infections.
All of the transmembrane SLAM family receptors for which intracellular signaling pathways have been identified can signal via the SLAM-associated protein (SAP) (3). SAP is encoded by SH2D1A, which is located on the X-chromosome, and various mutations of SH2D1A cause X-linked Lymphoproliferative disease (XLP) in male children, the hallmark of which is an inability to control EBV infection (6). The failure to control EBV infection is due to an inability of CTL to lyse EBV-infected B cells, which express high levels of ligands for SLAM family receptors expressed by CTL. Schlaphoff et al (7) reported that the degree of enhancement or inhibition of specific clonotypes of antiviral T cells by CD244-CD48 blockade correlated with the respective degree of SAP expressed by those clonotypes. This result implies that the effect of CD244 expression on T cell function is controlled both by the expression of CD48 and by the expression of an internal signaling molecule (SAP). CD244 can be recruited to the immune synapse in NK cells (8) and CD8 T cells (9). Recently Zhao et al (10) reported that SLAMf6 (also known as Ly109 in mice and NBTA in humans) can localize to the murine T cell synapse, and modulate T cell signaling by differential recruitment of Src homology region 2 domain-containing phosphatase-1 (SHP-1).
For the present study we investigated co-signaling receptor expression on CD8 T cells isolated from the blood of HIV+ or HIVneg subjects, and specific for a nonpersistent pathogen (respiratory syncytial virus, RSV) or a chronic persistent pathogen (HIV). We found that cultured antiviral CD8 T cells expressed varying levels of PD-1, TIM-3, and CD244. Upon recognition of cognate antigen, some T cells rapidly downregulated expression of TIM-3 and CD244. We found that CD244 downregulation in cultured or in ex vivo human CD8 T cells required simultaneous signaling both via the TCR and via CD244 itself and involved relocation of CD244 into an acidic intracellular compartment. Consistent with previous findings for other viruses, blockade of CD244-CD48 interaction had a dual effect of either enhancing or inhibiting various CD8 T cell clonotypes. Based on a comparison of CD244 expression on responding T cells in the presence or absence of CD48 blockade, we present evidence that the degree of enhancement or inhibition in the IFNγ response to antigen correlated with the degree of CD244 downmodulation in response to TCR stimulation.
Materials and Methods
Human subjects
Peripheral blood was obtained by venipuncture with acid-citrate dextrose anticoagulant from HIV-infected and uninfected individuals at the Massachusetts General Hospital (MGH), Boston. Written informed consent was obtained from all volunteers prior to enrollment in the study. Buffy coats were obtained from normal human donors via the MGH blood bank. The study was approved by the MGH Institutional Review Board. All subjects were aviremic at the time of blood draw. PBMC from HIV-infected individuals were isolated by Ficoll density gradient centrifugation within six hours of blood draw and cryopreserved in the presence of 10% DMSO in a liquid nitrogen freezer. PBMC from buffy coats were isolated by Ficoll density gradient centrifugation within 24 hours of blood draw and were not frozen.
Antibodies
For flow cytometry the following directly conjugated mAbs were used: CD3-Alexa Fluo 700 (BD; clone UCHT1), CD8-Qdot 605 (Invitrogen; clone 3B5), CD244 –PeCy5.5 (Biolegend; clone C1.7), PD1-V421 (Biolegend; clone EH12.2H7), IFNγ-PeCy7 (BD; clone B27), CD19-APC.Cy7 (BD; clone SJ25C1), CD14-APCCy7 (BD; clone MφP9), CD4-APCCY7 (BD; clone SK3), TIM3-PE (R&D Systems; clone 344823). For blockade anti-human CD48 functional grade purified (eBioscience eBio156-4H9) or mouse IgG1 K isotype control functional grade purified (eBioscience clone P3.6.2.8.1) were used. To prepare avidin-antibody tetramers, biotinylated anti-CD244 (Lifespan Biosciences, clone C1.7) or biotinylated mouse IgG1 control (Invitrogen) were used.
Peptides
Synthetic peptides were purchased from Genscript, or from the MGH peptide core, as follows: RSV-YY9, YLEKESIYY (11); RSV-B*57-RF9, RARRELPRF (12); HIV-YT9, YFPDWQNYT; HIV-B*57-KF11, KAFSPEVIPMF. Epitope identification was facilitated by reference to the Immune Epitope Database (www.iedb.org) (13).
T cell culture
CTL lines represented in Figures 2 and 7 were generated from cryopreserved PBMC from A*01+B*57+ HIV-infected or HIV-uninfected donors. PBMC were thawed and incubated with peptide for 10 days in R10 medium (RPMI supplemented with 10% fetal bovine serum plus hepes buffer, penicillin, streptomycin, and L-glutamine). IL-2 (50 IU/mL) was added on day 3. Specificity for cognate peptide was tested on day 10 by IFNγ ELISA. Lines that produced significant IFNγ in response to cognate peptide were further tested by ICS (Intracellular Cytokine Staining).
Figure 2. Antigen-specific TCR- and CD48-induced downregulation of CD244 in cultured T cell lines.

A. PBMC from an HIV+ donor were stimulated with optimal peptides RSV-YY9 or HIV-YT9 and expanded for ten days in culture. On day ten lines were stimulated with autologous B-LCL loaded with cognate peptide in the presence or absence of CD48 blockade as shown. Cells were stained for surface phenotype and IFNγ expression by ICS. The experiment was repeated seven times with PBMC from the same donor. A representative experiment is shown. B. Fold decrease in expression of CD244 and PD1 among IFNγ+ cells in seven repeated experiments was calculated as (MFI with CD48 blockade/MFI with isotype control), and fold change in IFNγ+ CD8 T cells induced by CD48 blockade was calculated as (IFNγ+ with CD48 blockade/IFNγ+ with isotype control). * indicates Wilcoxon p<0.02.
Figure 7. Enhancement of specific IFNγ response by CD48 blockade correlates with CD244 downmodulation but not with absolute level of CD244 expression.

CD8 T cell lines specific for RSV-epitopes were generated from fifteen HIV+ donors and seven HIVneg donors. After ten days in culture lines were tested for responsiveness to cognate peptide, and twenty four lines were found to be specific. Lines were stimulated overnight with or without cognate peptide in the presence or absence of CD48, and the percent IFNγ+ cells, as well as the MFI of CD244 and PD-1 on IFNγ+ cells was determined by ICS. Lines did not express significant amounts of IFNγ in the absence of cognate peptide (not shown). The fold enhancement of IFNγ response in the presence of cognate peptide was calculated as (% IFNγ+ cells in the presence of CD48 blockade/% IFNγ+ cells in the presence of isotype control). A. Lack of correlation between overall CD244 expression on IFNγ+ cells in the presence of CD48 blockade and enhancement in IFNγ+ response induced by CD48 blockade in HIV+ donors. B. A similar lack of correlation in HIVneg donors. C. Correlation among sixteen RSV-specific CD8 T cell lines from HIV+ donors between fold change in CD244 expression and enhancement in IFNγ+ response induced by CD48 blockade in (p=0.01; rs=0.37). D. Correlation among eight RSV-specific CD8 T cell lines from HIVneg donors between fold change in CD244 expression and enhancement in IFNγ+ response induced by CD48 blockade (p=0.046; rs=0.41) The fold change in CD244 expression on IFNγ+ cells was calculated as (MFI in the presence of CD48 blockade/MFI in the presence of isotype control). rs and p values are based on Spearman’s rank correlation.
To generate antigen specific clones, lines were stimulated with cognate peptide and stained for capture using the MACS IFNγ Secretion Assay and Detection Kit (Miltenyi). IFNγ+ cells were sorted at one cell per well into 96 well plates using a FACSAria cell sorter (BD) in Ragon Flow Cytometry Core. Single cells were propagated by stimulation with anti-CD3 (clone 12F6) in the presence of irradiated allogeneic PBMC.
Intracellular Cytokine Staining (ICS)
Cells were incubated with peptide or with antigen-loaded B-LCL for one hour in R10, and then overnight in R10 plus 10 μg/μL brefeldin A. Cells were stained for surface markers (CD3, CD8, CD14, CD4, CD19, CD244, TIM-3, PD-1), fixed, permeablized and stained for internal IFNγ. Flow cytometry data were acquired using a BD LSR II cytometer and were analyzed with FlowJo software (Treestar, Inc.).
Internalization assay
the fluorogenic pH sensor pHrodo-Red-Avidin was purchased from Invitrogen. pHrodo-Red-Avidin was mixed with specific or control antibody at a molar ratio of 1:4 in PBS/1% BSA, and incubated overnight at 4°. Mixture was briefly centrifuged to remove and precipitates and then added to cell suspension at the indicated dose in R10 medium plus 10 μg/mL BFA in the presence or absence of PHA. Cells were incubated at 37° for the indicated time and pHrodo fluorescence was determined by flow cytometry. Flow cytometry data were acquired using a BD Fortessa cytometer with a 561 nm excitation laser, and were analyzed with FlowJo software (Treestar, Inc.).
ELISA
ELISA was used to screening antigen specific CD8 T cell lines and \clones as described. Soluble IFNγ production by T cells lines was quantified using the Human IFNγ ELISA Set (BD OptEIA™San Diego, CA, U.S). CTL lines were stimulated with either medium alone or 5uM of cognate peptide overnight and supernatants were harvested and tested for IFNγ content as described by manufacturer instructions.
Statistical methods
Paired quantitative variables were compared by Wilcoxon signed rank test. Correlation was calculated using the Spearman correlation test. All calculations were performed using GraphPad software version 6 and a difference or a correlation was considered significant when p < 0.05
Results
In light of published reports of activation-induced internalization of CD244 on NK cells, and incorporation of SLAM family receptors into the T cell synapse, we decided to investigate dynamic changes in surface marker expression on human T cells. When ex vivo PBMC were stimulated with the superantigen SEB in an IFNγ ICS assay, we observed that the SEB-responsive cells expressed less CD244 than the nonresponding cells (not shown), consistent with preferential response to SEB by CD244low cells and/or with CD244 downregulation in conjunction with T cell activation.
To investigate further, we endeavored to stimulate T cells under conditions where the TCR stimulus was delivered by APC expressing high levels of the CD244 ligand, CD48. CD48 was highly expressed on B lymphoblastoid cell lines (B-LCL, not shown). Therefore we stimulated PBMC from HIV+ and HIVneg donors in the presence or absence of CD48 blockade, with autologous B-LCL that were loaded or not with SEB, and washed to remove unbound superantigen (Figure 1). After overnight stimulation, we assessed specific IFNγ production and CD244 expression by ICS. The majority of CD8 T cells that produced IFNγ in response to SEB were CD244low in the absence of blockade but CD244hi in the presence of CD48 blockade (Fig 1A), consistent with CD48- dependent downmodulation on SEB-reactive T CD8 cells. We repeated the SEB stimulation using PBMC from six different HIV+ donors, and four HIVneg donors, and determined the level of CD244 expression in the presence or absence of CD48 blockade (Fig 1B). In each case, the level of CD244 expression on SEB-reactive CD8 T cells was higher in the presence of CD48 blockade than without blockade, and the difference was significant (p<0.02, Wilcoxon signed-rank test). No effect of TCR stimulation on CD244 expression was observed for CD4 T cells (not shown). The degree of CD244 downregulation in this assay was independent of the amount of PD-1 expression, and PD-1 expression was not affected by SEB stimulation. Ex vivo TIM-3+ cells failed to produce IFNγ in this assay (not shown).
Figure 1. ex vivo TCR- and CD48-induced downregulation of CD244 but not PD-1.

PBMC from HIV+ and HIVneg donors were cocultured overnight with autologous B-LCL in the presence or absence of SEB and Isotype IgG or CD48 blockade. Cells were stained for surface phenotype and IFNγ expression by ICS. A. Zebra plots show the levels of CD244 and PD-1 on CD8 T cells for a representative subject. B. The SEB stimulation was repeated for six HIV+ and four HIVneg donors the MFI of CD244, PD- in the IFNγ+ gate was determined and the percent of CD8 T cells responding to SEB in the presence or absence of CD48 blockade was determined. Significance was assessed by Wilcoxon signed-rank test; * indicates p<0.02. Data shown represent results of two independent experiments.
We considered that the observed change in median fluorescence intensity (MFI) of CD244 on IFNγ+ T cells in the presence of CD48 blockade might be due to disinhibition of SEB-reactive CD244+ T cells and subsequent recruitment of CD244hi cells into the IFNγ+ population. Therefore we compared the percent IFNγ+ SEB-reactive CD8 T cells in the presence or absence of CD48 blockade. As shown in Fig 1B there was no significant change in the percent of CD8 T cells responding to SEB in the presence or absence of blockade. Overall these results are consistent with CD244 downmodulation being induced by combined signaling via SEB-reactive TCR and CD244 itself.
To investigate dynamic changes in marker expression on CD8 T cell lines specific for viral peptides, we stimulated PBMC from an HIV-infected subject with synthetic peptides corresponding to either of two epitopes: RSV-YY9 or HIV-YT9. After 10 days in culture, we performed ICS with autologous B-LCL loaded with cognate peptide in the presence or absence of CD48 blockade, and measured the amount of CD244 expressed on specific CTL (Figure 2). In the absence of CD48 blockade, the RSV-YY9-specific CTL expressed very low levels of CD244, whereas in the presence of CD48 blockade they expressed higher levels of CD244, consistent with CD48-mediated CD244 downregulation. In contrast HIV-YT9-specific CTL expressed high levels of CD244 even in the absence of CD48 blockade and CD48 blockade resulted in a small increase CD244 expression compared to that observed in the YY9-specific line. PD-1 expression was not affected by peptide stimulation or by CD48 blockade. To investigate whether T cells of the same clonotype would respond consistently over repeated tests, we conducted seven independent experiments all using PBMC from the same donor. The observed effects on CD244 expression for YY9- and YT9-specific cells were consistent in all experiments (Fig 2B). As for Figure 1 above, we determined that CD48 blockade did not have a consistent or significant effect on the percent of specific IFNγ+ cells for YY9 or YT9-specific lines from this donor. Thus Figure 2 shows that antiviral T cells of different clonotypes/specificities differed in their tendency to downregulate CD244 in response to cognate peptide. Notably this difference could not be attributed to differences in CD48 expression on the APC, because identical autologous B-LCL were used as APC for all stimulations.
To conclusively demonstrate CD48-dependent CD244 downmodulation on antiviral CD8 T cells, we investigated these effects at the level of an individual T cell clone. An RSV-YY9 line comparable to those shown in Fig 2 was generated from the blood of an HIV+ subject. To produce a clone, the line was stimulated with cognate peptide and live-sorted for peptide specificity using IFNγ capture at a concentration of one cell per well, and specific clones were propagated and screened for specificity (not shown). We used the specific clones to investigate the relationship between receptor downregulation and signaling via TCR and CD244.
When a representative clone specific for RSV-YY9 was stimulated with cognate peptide in an overnight ICS assay, expression of CD3 and CD8 were both reduced, as expected for activated T cells. In addition, surface expression of CD244 was almost entirely eliminated. Likewise, expression of TIM-3 was also strongly reduced. In contrast, expression of PD-1 was not reduced (Figure 3A, 3B). We compared these parameters for a panel of autologous CTL clones of varying specificities (not shown), and found that some degree of downregulation of CD244, TIM-3, CD8, CD3 was apparent for all clones, although downregulation of CD244 on the RSV -YY9 clone was the most extreme.
Figure 3. Downregulation of CD244 in a RSV-specific CD8 T cell clone.

A. A CD8 T cell clone specific for RSV-YY9 was incubated overnight in the presence or absence of cognate peptide. The expression of CD244, TIM-3, PD-1, CD8, and CD3 were determined by ICS. B. The experiment shown in A was repeated four times and the MFI of the markers shown was compared in the presence or absence of cognate peptide. * indicates Wilcoxon p<0.02.
In order to determine the respective roles of signaling through TCR and CD244 in receptor downmodulation, we stimulated a RSV-YY9 clone with cognate peptide or with PHA (which both activate the clone via the TCR), or with PMA-ionomycin (which bypasses the TCR), or with PMA-ionomycin plus cognate peptide. As shown in Figure 3B, CD244 and TIM-3 were both strongly downregulated in response to cognate peptide, and weakly downregulated in response to PHA. TIM-3, but not CD244, was also lost from the cell surface in response to PMA-ionomycin treatment. While PMA-ionomycin treatment did not enhance or inhibit CD244 expression; it also did not inhibit downregulation, as shown by the fact that simultaneous stimulation with PMA-ionomycin and cognate peptide did induce CD244 downregulation (Fig 4A). Loss of surface expression of CD244 was observed whether cognate peptide was added directly to CTL clones or if peptide was presented by autologous B-LCL (Data not shown)). As expected, downregulation of CD244, but not TIM-3, was specifically prevented by CD48 blockade (Fig 4B). These results prove that CD244 was downregulated in response CD8 T cell activation, and that simultaneous signals transmitted by TCR and CD244 itself were required.
Figure 4. Downregulation of CD244 is induced by TCR-proximal signaling and CD48.
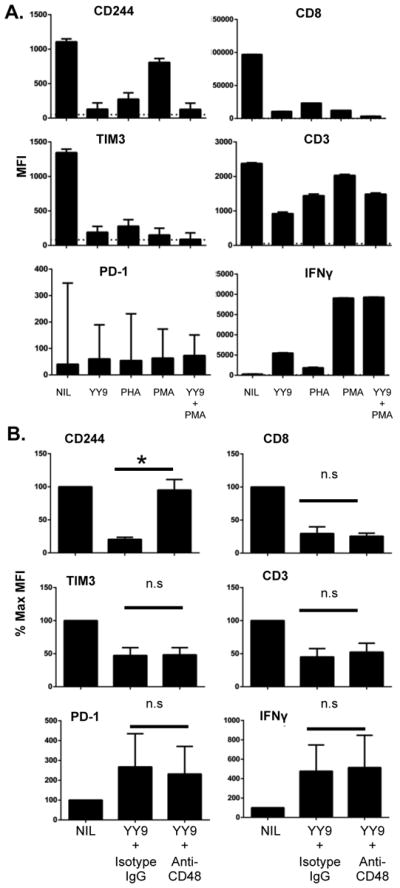
A. To determine the role of TCR signaling, the same clone was stimulated with cognate peptide (YY9), PHA, PMA/ionomycin, or PMA/ionomycing plus cognate peptide as shown. B. To determine whether a signal from CD48 was required for CD244 downregulation, RSV-YY9 clone was incubated overnight in the presence or absence of cognate peptide and CD48 blockade as shown. *p <0.05 paired T test; n.s. = not significant. All results are representative of at least three independent experiments.
We next performed a timecourse experiment to determine the kinetics of CD244 downregulation on a CTL clone specific for RSV1-YY9. As shown in Figure 5A, significant CD244 downregulation occurred during the first two hours of peptide exposure, with the majority of downregulation occurring within the first six hours of stimulation with cognate peptide. CD244 expression was not restored within the first 48 hours of stimulation (Fig 5B), although CD244 expression is high after ten days in culture post stimulation (Figs 2 and 3).
Figure 5. TCR-induced CD244 downregulation is rapid, sensitive and independent of PI3K.

A. RSV-YY9 clone was stimulated with cognate peptide for the indicated time, fixed, and stained for expression of CD244 and CD8. Expression was measured by flow cytometry. B. Cultured CD8 T cells were stimulated with PHA and incubated for the indicated time period. Expression of CD244 was not restored over a 48 hour period. C. RSV-YY9 clone was stimulated with increasing doses of cognate peptide overnight prior to fixation and staining. Expression of the indicated markers was measured by flow cytometry. D. RSV-YY9 clone was incubated overnight with or without cognate peptide in the presence of PI3K inhibitor LY294002, and expression of surface markers and IFNγ was measured by ICS. LY294002 effectively inhibited IFNγ production above 30 μM without affecting downregulation of CD244, TIM-3, CD3, or CD8. Results represent at least two experiments.
When CTL were stimulated by increasing doses of peptide, the markers CD244, CD3, and CD8 were downregulated concomitantly, while IFNγ expression was induced at higher peptide doses (Figure 5C). This is consistent with previously observed differences in the concentration of peptide required to induce CD3 internalization versus IFNγ production (14, 15).
To further examine involvement of TCR signaling pathways we stimulated the same CTL clone in the presence of cognate peptide and increasing doses of the PI3K inhibitor LY294002 (Fig 5D). Downregulation of both CD244 and TIM-3 were independent of PI3K signaling at all doses of LY294002, even though IFNγ expression was strongly inhibited in a dose-depended manner. Overall the results in Figures 4 and 5 demonstrate that downregulation of CD244 is dependent on a TCR-proximal signal in response to cognate peptide. In contrast TIM-3 downregulation is induced by either TCR proximal signals (as observed in the presence of both cognate peptide and LY294002) or TCR distal signals, as induced by the DAG analog PMA.
Downmodulation of CD244 in the above experiments was determined by quantification of surface CD244 expression. Those results do not address the mechanism: in theory loss of CD244 detection might be due to binding of a ligand that competes with the detecting antibody, or to shedding, or to sequestration in membrane ruffles, or to internalization. We hypothesized that CD244 downmodulation was a result of internalization, and developed an assay to measure CD244 internalization using the pH-sensitive indicator dye pHrodo (Fig 6). pHrodo dyes are minimally fluorescent at neutral pH, and fluoresce brightly in acidic environments such as late endosomal/lysosomal compartments (16–18).
Figure 6. Downmodulation of CD244 is mediated by endocytosis. A. Schematic summary of tetramer assay.

pHrodo-Red™produces minimal fluorescence at neutral pH, and fluoresces brightly at low pH. Specific or isotype control antibody was complexed with pHrodo-Red-avidin at a molar ration of 4:1 to form antibody tetramers. Tetramers were allowed to bind to CD8 T cells, and then CD8 T cells were stimulated with PHA or medium-only control. Internalization and transfer of tetramers to an acidic compartment was determined by fluorescent detection of pHrodo-red signal on a flow cytometer. B. pHrodo-red detection in activated CD8 T cells. A cultured CD8 T cell line was stained with anti-CD244 or isotype control tetramers and stimulated with PHA for five hours. Cells were fixed and stained for viability and T cell markers. Plots are gated on live CD8+ T cells. C. Internalization of anti-CD244 tetramers induced by simultaneous signals via CD244 and TCR. A cultured CD8+ T cell line was loaded with specific or isotype tetramer at the indicated concentrations for 30 minutes, washed and then stimulated with PHA or medium control. Cells were incubated at 37° for five hours, and then stained for FACS, fixed, and analyzed by flow cytometry. Results are representative of two similar experiments. The % pHrodo positive cells across multiple treatments was significantly greater for anti-CD244 tetramer compared to isotype control (Wilcoxan p<0.001).
To prepare avidin-antibody tetramers, we complexed biotinylated anti-CD244 antibody (or isotype control) to avidin-pHrodo conjugate at a molar ratio of 4:1. We then added increasing doses of avidin-pHrodo-antibody complexes to cultured CD8 T cells. Cells were washed and stimulated with medium alone or with PHA for six hours. pHrodo fluorescence was determined by flow cytometry. We used pHrodo fluorescence as an indicator of complete internalization and transport to a late endosomal compartment.
For a readout dependent on simple receptor attachment, signal would be expected to follow a monotonic curve and be directly proportional to ligand or antibody dose. In contrast, for a readout that depends on receptor cross-linking, signal is expected to follow a biphasic curve-- increasing to an inflection point, and then decreasing at supraoptimal doses as receptor is saturated and cross-linking is lost. Based on our results shown above, we predicted that receptor internalization as indicated by pHrodo fluorescence would be i) dependent on CD244 crosslinking (biphasic with respect to the dose of antibody-avidin complex), and ii) enhanced by TCR signal (i.e. PHA). Figure 6 shows that observed results were consistent with our predictions: pHrodo-red fluorescence is biphasic with respect to tetramer concentration, and it is enhanced by PHA stimulation. Tetramers formed with nonspecific isotype control antibody did not produce any significant pHrodo signal. Importantly, the fact that pHrodo fluorescence decreases at high tetramer concentration proves that fluorescence is dependent on a cross-linking signal, and is not the product of tetramer loading at the cell surface. These results demonstrate that subsequent to simultaneous signals via TCR and CD244, the CD244 molecule is internalized and enters an acidic endosomal/lysosomal compartment inside the T cell.
We wanted to investigate how CD244 downregulation in response to antigen might relate to antiviral CD8 T cell function. Because most of our subjects did not have significant ex vivo RSV-specific CD8 responses (not shown), we generated antiviral CD8 T cell lines as for Figure 2 above, from HIV+ and healthy HIVneg donors (Fig. 7). PBMC from each donor were stimulated with optimal viral peptides matched to the donor HLA type and expanded in culture for 10 days. Each antigen-responsive line was tested by ICS in the presence or absence of CD48 blockade. Consistent with the reported “dual functions” of CD244, we found that CD48 blockade sometimes produced an increase in the percent of IFNγ+ CD8 T cells, sometimes produced a decrease, and sometimes had no effect. We quantified the CD244-dependent change in antigen response as the ratio of the percent IFNγ+ CD8 T cells in the presence versus the absence of CD48 blockade.
Previous studies show that activating and inhibitory functions of CD244 are partly explained by the absolute level of CD244 expressed on the T cell surface, and partly explained by co-expression of signaling molecules and strength of signal from CD244 ligands (19). For our T cell lines we determined the total level of CD244 expression on IFNγ+ cells in the presence of CD48 blockade; RSV-specific lines from HIVneg donors expressed somewhat lower CD244 levels compared to HIV+ donors, although the difference was not significant (p=0.06). Furthermore, when we compared CD244 MFI to the degree of enhancement in IFNγ production produced by blockade there was no significant correlation for HIV+ or HIVneg donors (Fig 7A and B respectively).
Because our results suggest that CD244 downmodulation may be related to involvement of CD244 in the immune synapse we wondered whether the change in CD244 expression induced by TCR activation would correlate with enhancement of IFNγ production in response to CD48 blockade. Consistent with our observations above, we found that antigen stimulation produced a variable decrease (but never an increase) in surface expression of CD244. When we compared the degree of enhancement in IFNγ production versus the change in CD244 expression on responding cells in response to CD48 blockade, we found a significant positive correlation among RSV-specific CD8 lines from HIV+ donors (Fig 6C; slope=+0.38, rs=0.37, p=0.01). Intriguingly, for HIVneg donors we found the opposite relationship: a significant negative correlation (Fig 7D; slope= −0.35, rs=0.41, p=0.046). These results suggest a close connection between activation-induced CD244 downregulation and CD48-mediated modulation of CD8 T cell function.
Discussion
T cells are controlled by complex networks of signals. As one of many “switches” in these networks, CD244 may be of special interest because it has been demonstrated to have dual functions, with the ability to enhance or inhibit antiviral human CD8 T cell functions in different contexts (5). For example CD244 is one of a suite of markers progressively upregulated on exhausted T cells (1, 20–26). Consistent with a role in exhaustion, in subjects chronically infected with hepatitis B virus, blockade of CD244 or CD48 resulted in enhancement of antiviral cytotoxicity and cytokine production (27). In contrast, for subjects with HTLV-I associated neurologic disease, blockade of CD244, or knockdown of the signaling molecule SAP, reduced HTLV-specific antiviral functions (28). In subjects with HIV infection, blockade of CD244 was found to enhance HIV-specific CD8 T cell proliferation from PBMC (22). For subjects infected with HCV, blockade of CD48 was found to variously enhance or inhibit different CD8 clonotypes, with opposite effects correlating with the degree of SAP expression (7). In the present report we also find variable and opposing effects of CD48 blockade on different antiviral clonotypes. We also identify a mechanistic correlate of enhancement and inhibition: rapid dynamic downregulation of CD244 on the surface of the responding cell.
Recent studies have elucidated the molecular basis for dual action of CD244 (29). Human subjects with XLP disease, associated with mutations in SAP gene SH2D1A, are at high risk of lethal EBV infection due to an inability to clear EBV-infected B cells (30, 31). Many aspects of XLP disease are recapitulated in SH2D1A knockout mice (32). Zhao et al (10) demonstrated that SH2D1A -deficient murine CD8 T cells were competent to kill CD48-negative targets, but specifically defective in forming cytolytic immune synapses with B cell targets expressing high levels of CD48 and Ly108; these defects were associated with recruitment of SHP-1 phosphatase and decreased activation of Src kinases at the immune synapse. It is interesting to note that in addition to severe disease observed in XLP subjects with loss-of-function mutations in SH2D1A, subtle effects on SAP expression, for example due to a promoter-linked polymorphism, may also modulate human disease (33, 34).
Previous studies showed that CD244 can localize to the immune synapse in NK cells (8, 35), that SLAMf6 can localize to the synapse in T cells (36), and that CD244 relocates to the T cell-B-LCL interface in a SAP-dependent manner (9). CD244 downregulation was previously demonstrated in response to activation of NK cells (37). CD229 (SLAMf3) was found to be downregulated on Jurkat T cells in response to artificial coligation of CD229 plus TCR (38), similar to our finding that CD244 is downregulated on specific T cells by combined signals from CD48 and cognate peptide-MHC. To our knowledge the present report is the first to describe activation-induced downregulation of CD244 on T cells.
In this study we observed CD244 downregulation on primary ex vivo CD8 T cells and in cultured CD8 T cell lines and clones in response to activation via the TCR (by cognate ligand, SEB or PHA). Notably, downregulation was neither induced nor prevented by PMA/ionomycin, which activate T cells by bypassing TCR signaling, strongly suggesting that downregulation is mediated by signals proximal to the TCR and the immune synapse. TCR-dependent downregulation of CD244 also required signaling via CD244 itself, such that downregulation was prevented by CD48 blockade.
Before devoping the tetramer assay shown in Figure 6, we attempted several other approaches to monitor CD244 dynamics at the cell surface (not shown). Labeling CD244 at the cell surface with monomeric fluorescent antibody prior to activation interfered with receptor downmodulation, presumably by blocking interaction with CD48 (not shown). To prove that CD244 was being internalized, we developed a strategy based on two features: the ability of an avidin molecule to crosslink four biotinylated antibodies, and the pH-dependent fluorescence of the pHrodo-Red reagent (16–18). By mixing avidin and biotinylated antibody at a molar ratio of 1:4, we enhanced antibody uptake compared to a ratio of 1:1 or 1:2 (not shown); we presume that antibody tetramers are able to crosslink CD244 in a similar manner to an APC expressing CD48. As shown in Figure 6, pHrodo-avidin uptake increased with increasing dose of avidin, up to an inflection point, after which signal decreased. This is a classic dose-response curve for a crosslinking-dependent signal, such that supraoptimal doses of avidin lead to saturation of the receptor and reduced crosslinking. Receptor internalization is a feature of many lymphocyte signaling pathways; pH-sensitive antibody/ligand tetramers may be a useful tool for monitoring dynamic relocation of receptors in live primary T cells in multiple contexts.
In order to investigate whether the observed dynamic changes in CD244 expression were connected with T cell function, we compared the degree of enhancement or inhibition by CD48 blockade with the degree of CD244 downregulation in response to cognate peptide in cultured antiviral (RSV-specific) CD8 T cell lines. The fact that we observed a significant positive correlation in HIV+ donors suggests that differential CD244 incorporation into the synapse is intimately connected with modulation of TCR signaling. This fine-tuning function is consistent with the finding that CD244 modulation of antiviral T cell function correlates with the expression of a specific signaling molecule (SAP) (7). Compared to HIV+ subjects, T cell lines derived from HIVneg subjects presented an opposite, negative correlation between IFNγ production and CD244 downregulation. This suggests a model in which the context of signaling-molecule expression is determined by systemic factors such as chronic infection, which in turn drive some CTL toward positive or negative responses to CD244 ligation. Clearly, many key questions about the role CD244 in T cell signaling and response remain to be answered in future studies.
The effect of TCR stimulation on TIM-3 downregulation on T cell clones requires further investigation. Whereas CD48 blockade facilitated our ability to observe ex vivo IFNγ production by CD244+ cells in response to antigen and superantigen, we were unable to make a similar observation for TIM-3+ CD8 T cells ex vivo as antibody against the TIM-3 ligand Gal-9 had no effect (not shown). Short-term lines propagated ex vivo did express varying levels of TIM-3, even on IFNγ+ antigen specific cells, but there was no effect of TCR stimulation on TIM-3 expression (not shown). In contrast to PBMC and lines our antiviral clones did usually express high levels of TIM-3, which was downregulated in response to antigen or PMA (Fig. 4A). The mechanism of this modulation remains to be determined.
In conclusion, our results provide evidence that simultaneous ligation of CD244 by CD48, and of TCR by cognate peptide-MHC on an antigen-presenting cell, results in variable downmodulation of CD244, presumably via incorporation into the synapse. For a particular clonotype, the degree of downregulation correlates with the degree of enhancement or inhibition in antiviral functions induced by CD48 blockade. Our results help to explain the variable results seen for modulation of antiviral responses by CD244 blockade in human subjects, and point the way to possible interventions that might enhance the effects of therapeutic and prophylactic vaccines for infectious disease and cancer.
Acknowledgments
We wish to thank Sarah Allan and Oliver Davis for technical assistance, and Dr. Bruce Walker for scientific help and discussions. We also gratefully acknowledge the participation of volunteer blood donors.
Footnotes
This work was supported by the Terry and Susan Ragon Foundation, and by NIH/NIAID:1R56AI095088 and 5R01AI30914 (DGK), P01 AI-080192 (DEK and DGK), andP30 AI060354 (Harvard University Center for AIDS Research). DEK is supported by a Research Scholar Career Award of the Quebec Health Research Fund (FRQS).
Abbreviations: HCV, Hepatitis C Virus; B-LCL, B lymphoblastoid cell line; n.s., not significant; RSV, respiratory syncytial virus; SLAM, surface receptor signaling lymphocytic activation molecule; SHP-1, SRC homology region 2 domain-containing phosphatase-1; MFI, Median Fluorescence Intensity; XLP, X-linked Lymphoproliferative Disease.
References
- 1.Youngblood B, Wherry EJ, Ahmed R. Acquired transcriptional programming in functional and exhausted virus-specific CD8 T cells. Current opinion in HIV and AIDS. 2012;7:50–57. doi: 10.1097/COH.0b013e32834ddcf2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Porichis F, Kaufmann D. Role of PD-1 in HIV Pathogenesis and as Target for Therapy. Current HIV/AIDS Reports. 2012;9:81–90. doi: 10.1007/s11904-011-0106-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Detre C, Keszei M, Romero X, Tsokos GC, Terhorst C. SLAM family receptors and the SLAM-associated protein (SAP) modulate T cell functions. Seminars in immunopathology. 2010;32:157–171. doi: 10.1007/s00281-009-0193-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mathew SO, Rao KK, Kim JR, Bambard ND, Mathew PA. Functional role of human NK cell receptor 2B4 (CD244) isoforms. European journal of immunology. 2009;39:1632–1641. doi: 10.1002/eji.200838733. [DOI] [PubMed] [Google Scholar]
- 5.Waggoner SN, Kumar V. Evolving role of 2B4/CD244 in T and NK cell responses during virus infection. Front Immunol. 2012;3:377. doi: 10.3389/fimmu.2012.00377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cannons JL, Tangye SG, Schwartzberg PL. SLAM family receptors and SAP adaptors in immunity. Annual review of immunology. 2011;29:665–705. doi: 10.1146/annurev-immunol-030409-101302. [DOI] [PubMed] [Google Scholar]
- 7.Schlaphoff V, Lunemann S, Suneetha PV, Jaroszewicz J, Grabowski J, Dietz J, Helfritz F, Bektas H, Sarrazin C, Manns MP, Cornberg M, Wedemeyer H. Dual function of the NK cell receptor 2B4 (CD244) in the regulation of HCV-specific CD8+ T cells. PLoS pathogens. 2011;7:e1002045. doi: 10.1371/journal.ppat.1002045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Liu D, Bryceson YT, Meckel T, Vasiliver-Shamis G, Dustin ML, Long EO. Integrin-dependent organization and bidirectional vesicular traffic at cytotoxic immune synapses. Immunity. 2009;31:99–109. doi: 10.1016/j.immuni.2009.05.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dupre L, Andolfi G, Tangye SG, Clementi R, Locatelli F, Arico M, Aiuti A, Roncarolo MG. SAP controls the cytolytic activity of CD8+ T cells against EBV-infected cells. Blood. 2005;105:4383–4389. doi: 10.1182/blood-2004-08-3269. [DOI] [PubMed] [Google Scholar]
- 10.Zhao F, Cannons JL, Dutta M, Griffiths GM, Schwartzberg PL. Positive and negative signaling through SLAM receptors regulate synapse organization and thresholds of cytolysis. Immunity. 2012;36:1003–1016. doi: 10.1016/j.immuni.2012.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Heidema J, de Bree GJ, de Graaff PMA, van Maren WWC, Hoogerhout P, Out TA, Kimpen JLL, van Bleek GM. Human CD8+ T cell responses against five newly identified respiratory syncytial virus-derived epitopes. Journal of General Virology. 2004;85:2365–2374. doi: 10.1099/vir.0.80131-0. [DOI] [PubMed] [Google Scholar]
- 12.Brandenburg AH, de Waal L, Timmerman HH, Hoogerhout P, de Swart RL, Osterhaus AD. HLA class I-restricted cytotoxic T-cell epitopes of the respiratory syncytial virus fusion protein. Journal of virology. 2000;74:10240–10244. doi: 10.1128/jvi.74.21.10240-10244.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vita R, Zarebski L, Greenbaum JA, Emami H, Hoof I, Salimi N, Damle R, Sette A, Peters B. The immune epitope database 2.0. Nucleic acids research. 2010;38:D854–862. doi: 10.1093/nar/gkp1004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Valitutti S, Muller S, Dessing M, Lanzavecchia A. Different responses are elicited in cytotoxic T lymphocytes by different levels of T cell receptor occupancy. The Journal of experimental medicine. 1996;183:1917–1921. doi: 10.1084/jem.183.4.1917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hemmer B, Stefanova I, Vergelli M, Germain RN, Martin R. Relationships Among TCR Ligand Potency, Thresholds for Effector Function Elicitation, and the Quality of Early Signaling Events in Human T Cells. The Journal of Immunology. 1998;160:5807–5814. [PubMed] [Google Scholar]
- 16.Miksa M, Komura H, Wu R, Shah KG, Wang P. A novel method to determine the engulfment of apoptotic cells by macrophages using pHrodo succinimidyl ester. J Immunol Methods. 2009;342:71–77. doi: 10.1016/j.jim.2008.11.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Deriy LV, Gomez EA, Zhang G, Beacham DW, Hopson JA, Gallan AJ, Shevchenko PD, Bindokas VP, Nelson DJ. Disease-causing mutations in the cystic fibrosis transmembrane conductance regulator determine the functional responses of alveolar macrophages. J Biol Chem. 2009;284:35926–35938. doi: 10.1074/jbc.M109.057372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Han J, Burgess K. Fluorescent Indicators for Intracellular pH. Chemical Reviews. 2009;110:2709–2728. doi: 10.1021/cr900249z. [DOI] [PubMed] [Google Scholar]
- 19.Chlewicki LK, Velikovsky CA, Balakrishnan V, Mariuzza RA, Kumar V. Molecular basis of the dual functions of 2B4 (CD244) J Immunol. 2008;180:8159–8167. doi: 10.4049/jimmunol.180.12.8159. [DOI] [PubMed] [Google Scholar]
- 20.Wherry EJ, Ha SJ, Kaech SM, Haining WN, Sarkar S, Kalia V, Subramaniam S, Blattman JN, Barber DL, Ahmed R. Molecular Signature of CD8+ T Cell Exhaustion during Chronic Viral Infection. Immunity. 2007;27:670–684. doi: 10.1016/j.immuni.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 21.Doering T, Crawford A, Angelosanto J, Paley M, Ziegler C, Wherry J. Network Analysis Reveals Centrally Connected Genes and Pathways Involved in CD8+ T Cell Exhaustion versus Memory. Immunity. 2012;37:1130–1144. doi: 10.1016/j.immuni.2012.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yamamoto T, Price DA, Casazza JP, Ferrari G, Nason M, Chattopadhyay PK, Roederer M, Gostick E, Katsikis PD, Douek DC, Haubrich R, Petrovas C, Koup RA. Surface expression patterns of negative regulatory molecules identify determinants of virus-specific CD8+ T-cell exhaustion in HIV infection. Blood. 2011;117:4805–4815. doi: 10.1182/blood-2010-11-317297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bengsch B, Seigel B, Ruhl M, Timm J, Kuntz M, Blum HE, Pircher H, Thimme R. Coexpression of PD-1, 2B4, CD160 and KLRG1 on exhausted HCV-specific CD8+ T cells is linked to antigen recognition and T cell differentiation. PLoS pathogens. 2010;6:e1000947. doi: 10.1371/journal.ppat.1000947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shin H, Blackburn SD, Intlekofer AM, Kao C, Angelosanto JM, Reiner SL, Wherry EJ. A role for the transcriptional repressor Blimp-1 in CD8(+) T cell exhaustion during chronic viral infection. Immunity. 2009;31:309–320. doi: 10.1016/j.immuni.2009.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Porichis F, Kwon DS, Zupkosky J, Tighe DP, McMullen A, Brockman MA, Pavlik DF, Rodriguez-Garcia M, Pereyra F, Freeman GJ, Kavanagh DG, Kaufmann DE. Responsiveness of HIV-specific CD4 T cells to PD-1 blockade. Blood. 2011;118:965–974. doi: 10.1182/blood-2010-12-328070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Peretz Y, He Z, Shi Y, Yassine-Diab B, Goulet JP, Bordi R, Filali-Mouhim A, Loubert JB, El-Far M, Dupuy FP, Boulassel MR, Tremblay C, Routy JP, Bernard N, Balderas R, Haddad EK, Sekaly RP. CD160 and PD-1 co-expression on HIV-specific CD8 T cells defines a subset with advanced dysfunction. PLoS pathogens. 2012;8:e1002840. doi: 10.1371/journal.ppat.1002840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Raziorrouh B, Schraut W, Gerlach T, Nowack D, Gruner NH, Ulsenheimer A, Zachoval R, Wachtler M, Spannagl M, Haas J, Diepolder HM, Jung MC. The immunoregulatory role of CD244 in chronic hepatitis B infection and its inhibitory potential on virus-specific CD8+ T-cell function. Hepatology. 2010;52:1934–1947. doi: 10.1002/hep.23936. [DOI] [PubMed] [Google Scholar]
- 28.Enose-Akahata Y, Matsuura E, Oh U, Jacobson S. High expression of CD244 and SAP regulated CD8 T cell responses of patients with HTLV-I associated neurologic disease. PLoS pathogens. 2009;5:e1000682. doi: 10.1371/journal.ppat.1000682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Le Borgne M, Shaw AS. SAP signaling: a dual mechanism of action. Immunity. 2012;36:899–901. doi: 10.1016/j.immuni.2012.06.002. [DOI] [PubMed] [Google Scholar]
- 30.Palendira U, Low C, Chan A, Hislop AD, Ho E, Phan TG, Deenick E, Cook MC, Riminton DS, Choo S, Loh R, Alvaro F, Booth C, Gaspar HB, Moretta A, Khanna R, Rickinson AB, Tangye SG. Molecular pathogenesis of EBV susceptibility in XLP as revealed by analysis of female carriers with heterozygous expression of SAP. PLoS biology. 2011;9:e1001187. doi: 10.1371/journal.pbio.1001187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hislop AD, Palendira U, Leese AM, Arkwright PD, Rohrlich PS, Tangye SG, Gaspar HB, Lankester AC, Moretta A, Rickinson AB. Impaired Epstein-Barr virus-specific CD8+ T-cell function in X-linked lymphoproliferative disease is restricted to SLAM family-positive B-cell targets. Blood. 2010;116:3249–3257. doi: 10.1182/blood-2009-09-238832. [DOI] [PubMed] [Google Scholar]
- 32.Crotty S, McCausland MM, Aubert RD, Wherry EJ, Ahmed R. Hypogammaglobulinemia and exacerbated CD8 T-cell-mediated immunopathology in SAP-deficient mice with chronic LCMV infection mimics human XLP disease. Blood. 2006;108:3085–3093. doi: 10.1182/blood-2006-04-018929. [DOI] [PubMed] [Google Scholar]
- 33.Boggio E, Melensi M, Bocca S, Chiocchetti A, Comi C, Clemente N, Orilieri E, Soluri MF, D’Alfonso S, Mechelli R, Gentile G, Poggi A, Salvetti M, Ramenghi U, Dianzani U. The -346T polymorphism of the SH2D1A gene is a risk factor for development of autoimmunity/lymphoproliferation in males with defective Fas function. Human immunology. 2012;73:585–592. doi: 10.1016/j.humimm.2012.02.025. [DOI] [PubMed] [Google Scholar]
- 34.Sawada S. Slam-associated protein plays a key role in development of autoimmunity. Autoimmunity Reviews. 2012;11:804–805. doi: 10.1016/j.autrev.2012.02.010. [DOI] [PubMed] [Google Scholar]
- 35.Sinha SK, Gao N, Guo Y, Yuan D. Mechanism of Induction of NK Activation by 2B4 (CD244) via Its Cognate Ligand. The Journal of Immunology. 2010;185:5205–5210. doi: 10.4049/jimmunol.1002518. [DOI] [PubMed] [Google Scholar]
- 36.Snow AL, Marsh RA, Krummey SM, Roehrs P, Young LR, Zhang K, van Hoff J, Dhar D, Nichols KE, Filipovich AH, Su HC, Bleesing JJ, Lenardo MJ. Restimulation-induced apoptosis of T cells is impaired in patients with X-linked lymphoproliferative disease caused by SAP deficiency. The Journal of clinical investigation. 2009;119:2976–2989. doi: 10.1172/JCI39518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sandusky MM, Messmer B, Watzl C. Regulation of 2B4 (CD244)-mediated NK cell activation by ligand-induced receptor modulation. European journal of immunology. 2006;36:3268–3276. doi: 10.1002/eji.200636146. [DOI] [PubMed] [Google Scholar]
- 38.Del Valle JM, Engel P, Martin M. The cell surface expression of SAP-binding receptor CD229 is regulated via its interaction with clathrin-associated adaptor complex 2 (AP-2) J Biol Chem. 2003;278:17430–17437. doi: 10.1074/jbc.M301569200. [DOI] [PubMed] [Google Scholar]