

Abstract
Background
Relapsing fever spirochetes are global yet neglected pathogens causing recurrent febrile episodes, chills, nausea, vomiting, and pregnancy complications. Given these nonspecific clinical manifestations, improving diagnostic assays for relapsing fever spirochetes will allow for identification of endemic foci and expedite proper treatment. Previously, an antigen designated the Borrelia immunogenic protein A (BipA) was identified in the North American species Borrelia hermsii. Thus far, BipA appears unique to relapsing fever spirochetes. The antigen remains unidentified outside of these pathogens, while interspecies amino acid identity for BipA in relapsing fever spirochetes is only 24–36%. The current study investigated the immunogenicity of BipA in Borrelia turicatae, a species distributed in the southern United States and Latin America.
Methodology/Principal Findings
bipA was amplified from six isolates of Borrelia turicatae, and sequence analysis demonstrated that the gene is conserved among isolates. A tick transmission system was developed for B. turicatae in mice and a canine, two likely vertebrate hosts, which enabled the evaluation of serological responses against recombinant BipA (rBipA). These studies indicated that BipA is antigenic in both animal systems after infection by tick bite, yet serum antibodies failed to bind to B. hermsii rBipA at a detectable level. Moreover, mice continued to generate an antibody response against BipA one year after the initial infection, further demonstrating the protein's potential toward identifying endemic foci for B. turicatae.
Conclusions/Significance
These initial studies support the hypothesis that BipA is a spirochete antigen unique to a relapsing fever Borrelia species, and could be used to improve efforts for identifying B. turicatae endemic regions.
Author Summary
Undiagnosed febrile illnesses continue to afflict those in resource poor countries. Relapsing fever spirochetes are one such pathogen causing a significant health burden, yet the pathogenesis, ecology, and distribution of B. turicatae is understudied. To address these shortcomings, we analyzed the amino acid sequence of the Borrelia immunogenic protein A (BipA) in isolates of B. turicatae. Mice and a canine were also infected by tick bite and transmission and serological responses were evaluated in these two likely mammalian hosts. B. turicatae was visualized within the blood of both animals and antibody responses generated against recombinant BipA indicated that the antigen that may be unique to infections caused by B. turicatae. Moreover, mice continued to generate antibodies a year after tick bite, suggesting a persistent infection. Our results indicate that the immune responses generated against BipA could identify additional vertebrate hosts, define endemic foci for B. turicatae, and increase the awareness of the disease to improve healthcare.
Introduction
Three causative agents of tick-borne relapsing fever borreliosis in the New World are Borrelia hermsii, Borrelia parkeri, and Borrelia turicatae, with B. hermsii being the most epidemiologically and ecologically characterized species [1]. While B. hermsii is distributed in high elevation coniferous forests and maintained in enzootic cycles with rodents as the primary reservoir, less is known regarding the other two species. Moreover, few epidemiological studies have been performed and little molecular data exists for B. turicatae and its arthropod vectors Ornithodoros turicata. There are endemic foci for B. turicatae in Texas and Florida, where clinical isolates have been obtained from sick dogs [2], [3], which suggests a role for wild canids in the maintenance of the spirochetes in nature. Dr. Oscar Felsenfeld also reported the distribution of O. turicata into Mexico, Central, and South America [4], yet given the absence of Latin American isolates for B. turicatae the identification of endemic foci is unclear.
A limitation in defining the distribution of B. turicatae has been the absence of diagnostic antigens specific for the species. Previously, the Borrelia immunogenic protein A (BipA) of B. hermsii was demonstrated to discriminate infections caused by Lyme and relapsing fever borreliosis. Outside of relapsing fever spirochete spp. a homologue of BipA has not been cataloged in the GenBank database [5]. With 36% amino acid identity between B. hermsii and B. turicatae BipA [5], it is also unclear if the B. turicatae homologue induces a host antibody response during infection. This study investigated the sequence similarity of BipA between B. turicatae isolates, and we developed a tick transmission system for the spirochetes to determine the antigenicity of recombinant BipA (rBipA) during rodent and canine infections. Collectively, these results suggests that BipA can be used as a diagnostic antigen for B. turicatae.
Methods
Ethical statement
All animal studies were in accordance with the Mississippi State University Institutional Animal Care and Use Committee (IACUC protocol #'s 11-091 and 12-067). Animal husbandry was provided by veterinary staff and technicians within the Association for Assessment and Accreditation of Laboratory Animal Care and the National Institutes of Health Office of Laboratory Animal Welfare assured program at Mississippi State University. All work was performed in adherence to the United States Public Health Service Policy on Humane Care and Use of Laboratory Animals and the Guide for the Care and Use of Laboratory Animals.
Amplification and sequence analysis of BipA from B. turicatae isolates
B. turicatae was cultivated in mBSK medium containing 12% rabbit serum [6], [7]. Amplification and sequencing primers for bipA were designed using the 91E135 isolate of B. turicatae (Table 1). Additional samples were water (negative control), 95PE-570, 95PE-1807, TCB-1, TCB-2, and FCB-1 isolates [2]. Polymerase chain reaction (PCR) was performed as previously described using the GoTaq Flexi DNA Polymerase (Promega Corporation, Madison, WI, USA). Amplicons were electrophoresed on a 1% agarose gel to visualize the DNA fragment and processed through the QIAquick PCR Purification kit (Qiagen, Germantown, MD, USA), and sequencing performed at Biodesign Institute (Arizona State University, Phoenix, AZ, USA). Nucleotide sequences were analyzed with the Vector NTI software (Life Technologies, Carlsbad, CA, USA), and deposited to GenBank under accession numbers KC845527-KC845531.
Table 1. Oligonucleotides and quantitative PCR probe.
| Primer | Sequence (5′→3′) | Role |
| bipA F D-Topo | CACCATGTGGTTTGTAAGGAGGGTGGATAT | Amplification and sequencing |
| bipA R Topo | ATTTGTAGCTTCAATTTTAGATTTAAACCCTA | Amplification and sequencing |
| bipA F1 | GTTTTCCAAAATTTAGGAGATACTGGTACT | Sequencing |
| bipA F2 | TATGCTTTTTGGTACGGGAGAGAA | Sequencing |
| bipA R1 | GCCTTTACTTGTAACATCAGAACTCAAGA | Sequencing |
| bipA R2 | AATTGAATTTATTGAATTTTCATTTTCTGTT | Sequencing |
| flaB probe | YAK-TGCAGGTGAAGGTGCGCAGGTT-BBQ | Quantitative PCR |
| flaB F | CCAGCATCATTAGCTGGATCAC | Quantitative PCR |
| flaB R | GTTGTGCACCTTCCTGAGC | Quantitative PCR |
Tick colony
The ticks used in the study originated from 12 uninfected adults initially maintained at the Rocky Mountain Laboratories. A cohort of second nymphal stage O. turicata was infected by first needle inoculating a group of 3 three Swiss Webster mice with B. turicatae 91E135 [2], and uninfected ticks were allowed to engorge on the animals. After molting, vector colonization was confirmed by dissecting the midgut and salivary glands from five ticks and performing immunofluorescent assays (IFA) as previously described [8]. Chicken serum generated by Cocalico Biologicals Inc. against B. turicatae recombinant flagellin (rFlaB) was used to detect spirochetes and the secondary antibody was goat anti-chicken IgY Alexa Fluor 568 (Life Technologies, Grand Island, NY, USA).
Transmission of B. turicatae
Prior to the transmission studies, pre-infection serum samples were collected from all animals. Cohorts of five infected ticks per animal were allowed to feed to repletion. The animals used in the study were 10 outbred Swiss Webster mice (Harlan Laboratories Inc., Tampa, FL, USA) and a one-year-old Bluetick hound (Marshall Bioresources, North Rose, NY, USA). For 16 consecutive days, animals were monitored for clinical symptoms and a drop of blood was collected from the mice by tail nick and from the canine's cephalic vein to visualize spirochetes. The day following the first and second febrile episode, 3 ml of blood were collected from the canine for a complete blood count and serum chemistry profile.
Of the mice used in the initial transmission study, four were maintained for one year to assess long-term serological responses generated against BipA. Prior to serum collection for immunoblotting, 2.5 µl of blood was collected for 10 consecutive days and placed into 47.5 µl of SideStep Lysis and Stabilization Buffer (Agilent, Santa Clara, CA, USA). Quantitative PCR (qPCR) was performed as previously described [9] to determine the presence of circulating spirochetes. The probe (flaB probe) and primer set (flaB F and flaB R) used for qPCR were designed for B. turicatae flagellin (Table 1).
Two groups of three mice were also needle inoculated intraperitoneally with 1×105 TCB-1 or FCB-1 spirochetes. Infection was confirmed by dark field microscopy and serum samples were collected one month after inoculation to determine reactivity to rBipA expressed from the 91E135 isolate.
Expression of B. turicatae bipA as a recombinant fusion protein, serological and linear regression analysis
To express bipA from B. turicatae 91E135 as a 75 kDa thioredoxin and histidine tagged fusion protein in Escherichia coli, the gene was amplified as previously described [5] using bipA F Topo and bipA R Topo primers (Table 1). The amplicon was cloned into the pET 102/Directional TOPO expression vector following the manufacturer's instructions (Life Technologies). Top10 E. coli were transformed, plasmid DNA isolated, and sequence analysis using bipA F1, bipA F2, bipA R1, bipA R2 primers (Table 1) was performed as previously described [5] to determine if an error had been introduced during amplification. To produce recombinant protein, BL21 E. coli were transformed with the bipA expression vector following the manufacturer's instructions, and induction was performed with 1 mM IPTG. rBipA was purified using the Ni-MAC Purification system (Novagen, Durmstadt, Germany).
Immunoblotting was performed to evaluate the immunogenicity of rBipA during B. turicatae infections. Protein lysates from 1×108 spirochetes, 1 µg of B. hermsii rBipA , and 1 µg B. turicatae rBipA were electrophoresed and transferred to polyvinylidene fluoride (PVDF) membranes using TGX gels, the Mini-PROTEAN Tera cell, and the Mini Trans Blot system (BioRad, Hercules, CA, USA). Pre- and post-infection serum samples were evaluated by immunoblotting at a 1∶500 dilution and the secondary molecule used was Rec-protein G-HRP (Life Technologies) at a 1∶4,000 dilution. Immunoblots were also probed with the Anti-polyHistidine Peroxidase monoclonal antibody (Sigma-Aldrich, St. Louis, MO, USA) at a 1∶4,000 dilution. Titers against rBipA were determined by immunoblotting with serum dilutions ranging from 1∶500 to 1∶125,000.
Linear regression analysis was performed by calculating the density of rBipA protein bands from immunoblots probed with serum samples diluted from 1∶800 to 1∶12,800. Serum samples included Anti-polyHistidine Peroxidase monoclonal antibody (Sigma-Aldrich, St. Louis, MO, USA) and serum samples from mice and the canine infected by tick bite. ImageJ, http://imagej.nih.gov/ij (National Institutes of Health, Bethesda, Maryland, USA), was used to analyze digitally scanned immunoblots and the density of each protein band was calculated. The R software package, www.r-project.org, was used to calculate equations of regression, R2 values, and significance.
Histopathology
One year after tick bite, mice were euthanized by isoflurane inhalation followed by cervical dislocation and tissues were placed in 10% neutral buffered formalin for fixation. Sections of brain and synovial joints from the front and rear leg were processed and embedded in paraffin using standard histologic techniques. Paraffin embedded tissues were cut in 5 µm sections, deparaffinized, adhered to glass slides, and stained with hematoxylin and eosin (H&E), or silver stained using standard histologic techniques. The sections were examined by light microscopy for inflammatory and degenerative lesions in the cerebrum, cerebellum, brainstem, and synovial joints by a board certified veterinary pathologist at Mississippi State University.
Results and Discussion
BipA was originally identified by an immunoproteomic antigen discovery approach [5], [10]. The degree dissimilarity between B. hermsii and B. turicatae homologues prompted our investigation to compare sequences between B. turicatae isolates, and to evaluate the protein's immunogenicity in two mammals likely to be naturally exposed to the spirochetes. PCR amplification of bipA from three canine (FCB-1, TCB-1, and TCB-2) and three tick (91E135, 95PE-570, and 95PE-1807) isolates produced a product of the expected molecular mass, with TCB-1 producing a slightly larger amplicon (Figure 1). Sequence analysis identified an additional 135 nucleotides encoding 45 amino acids for TCB-1 (Figure 2). While there was an overall 89% amino acid identity of BipA between isolates, the sequences flanking the 45 additional amino acids of TCB-1 were 97% identical. A similar observation was reported for B. hermsii BipA [5]. The protein from genomic group II (GG II) isolates of B. hermsii contained five regions of 3–24 amino acid insertions when compared to genomic group I (GG I) isolates. Furthermore, the amino and carboxy terminus of BipA between GGI and GGII isolates shared the highest degree of conservation.
Figure 1. PCR analysis of bipA from six B. turicatae isolates.

Molecular masses are shown in kilobases to the left of the gel.
Figure 2. Amino acid alignments of B. turicatae BipA from six isolates.

Identical amino acids are shaded grey.
Currently, the only known isolates of B. turicatae originate from argasid soft ticks and sick dogs [2]. Furthermore, the mammalian hosts for most species of relapsing fever spirochetes include rodents and insectivores [11]. This knowledge directed us to evaluate the antigenicity of B. turicatae rBipA after tick bite using canine and murine animal models. To establish an infected tick colony, uninfected O. turicata engorged on a Swiss Webster mouse needle-inoculated with B. turicatae 91E135. Spirochete colonization was confirmed by performing IFA on the midgut and salivary glands after the ticks molted (data not shown). The remaining infected ticks fed to repletion on Swiss Webster mice and a Bluetick hound, and within four and eight days after tick bite spirochetes were visualized in murine and canine blood, respectively (Figure 3 A and B). While the mice remained active when B. turicatae were visualized in the blood, the canine became febrile, lethargic, and following a given spirochetemic episode, acutely thrombocytopenic (Table 2). B. turicatae repopulated the blood from both groups of animals within four days after the initial spirochetemia, after which bacteria were undetectable by microscopy.
Figure 3. Visualization of B. turicatae within canine (A) and murine (B) blood after staining thin smears by Wright-Giemsa.
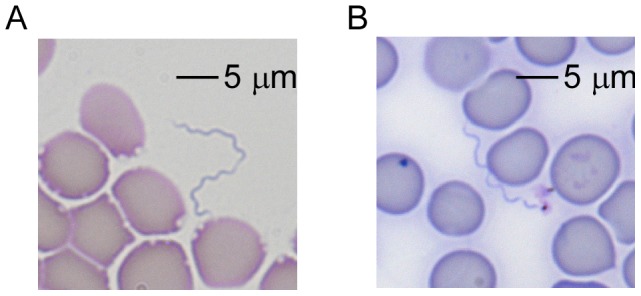
A 5 µm scale is shown.
Table 2. Canine temperature and platelet counts during infection.
| Days post infection | Temperature (Celsius) | Platelet Counts per µlA |
| 0 | 39.0B | 331,000B |
| 5 | 39.0 | - |
| 6 | 39.0 | - |
| 7 | 39.1 | - |
| 8C | 40.4 | - |
| 9 | 38.4 | 6,000 |
| 10 | 38.9 | - |
| 11 | 38.6 | - |
| 12C | 40.1 | - |
| 13 | 38.8 | 8,000 |
| 14 | 38.4 | - |
| 15 | 38.3 | - |
| 16 | 38.1 | 204,000 |
Values are based on automated platelet counts. Reference range was from 160,000 to 650,000. Automated platelet counts were noted on days 9 and 13, when counts were below normal.
Baseline temperature and platelet counts were taken prior to transmission by tick bite.
Spirochetemic episode.
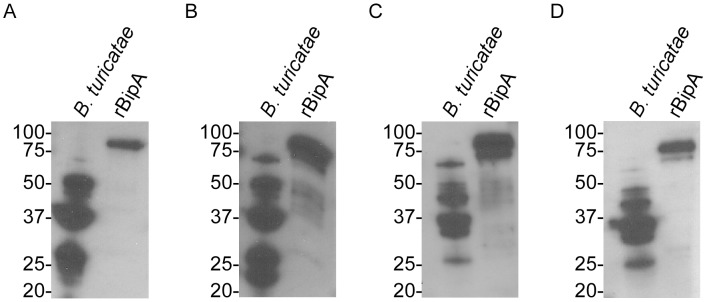
Producing rBipA using the same expression vector as B. hermsii bipA [5] indicated that infected animals generated an immunological response that recognized B. turicatae rBipA, yet antibody binding against the recombinant B. hermsii homologue was undetectable (Figure 4 A and B). Probing the immunoblots with an anti-polyhistidine monoclonal antibody confirmed that similar protein loads of B. turicatae and B. hermsii rBipA were electrophoresed, while pre-infection serum samples failed to produce a detectable antibody response against B. turicatae protein lysates or rBipA (Figure 4 C–E). Canine and murine IgG titers using serum samples collected 8 weeks after tick bite ranged from 1∶12,800 to1∶28,800. Also, regression analysis indicated significant differences (P≤0.05) in slopes and correlation coefficients (R2) when immune serum samples were probed against B. turicatae and B. hermsii rBipA (Figure 5 A–C). These results indicate different affinity characteristics against rBipA from a given species when animals were infected with B. turicatae.
Figure 4. Immunoblotting to evaluate serological responses from an infected canine (A) and mice (B) against B. turicatae protein lysates and rBipA.

Immunoblots were also probed with an anti-polyhistidine monoclonal antibody (C), canine pre-infection serum (D), and murine pre-infections serum (E). The (←), (*), and (<) indicate the molecular size of B. turicatae rBipA, native B. turicatae BipA, and B. hermsii rBipA, respectively. Molecular masses are indicated to the left of each immunoblot in kilodaltons.
Figure 5. Linear regression analysis of serological responses generated against B. turicatae rBipA.

Immunoblots containing B. turicatae rBipA (Bt rBipA) and B. hermsii rBipA (Bh rBipA) were probed with an anti-polyhistidine monoclonal antibody (A), and with serum from two mice infected by tick bite (B and C). Serum dilutions, regression slopes, and R2 values are show on the far left and below each immunoblot, respectively. The (*) indicates significant differences between regression slopes and R2 values.
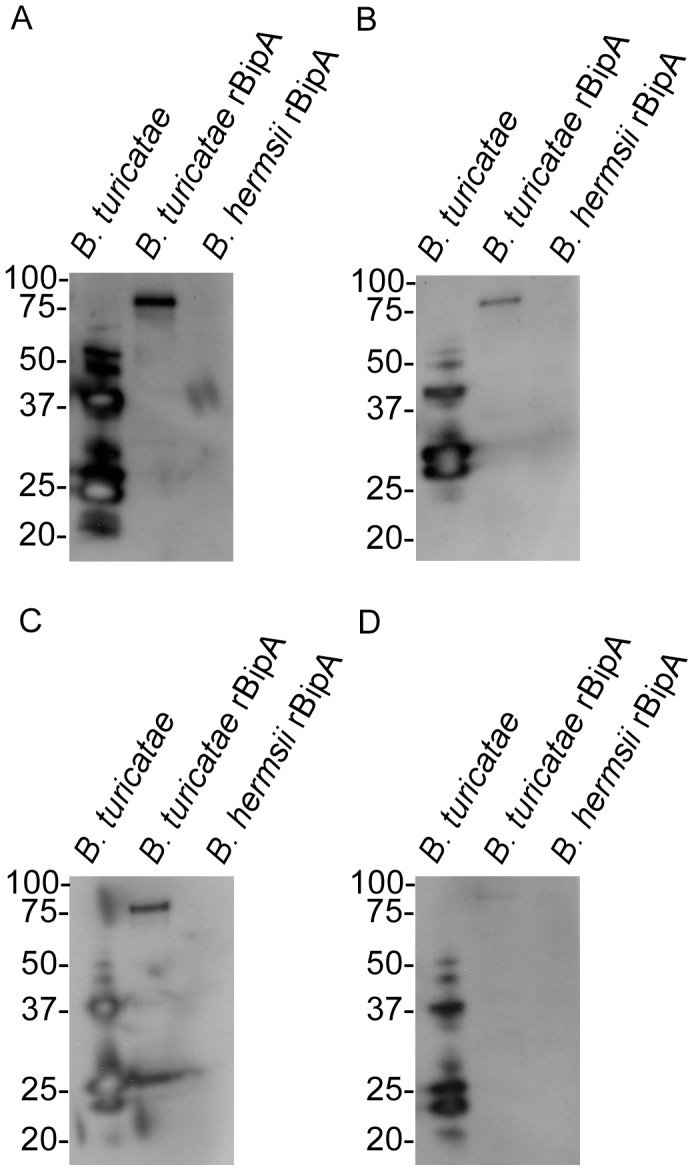
With BipA from TCB-1 and FCB-1 being the most divergent to the 91E135 homologue, we evaluated serological responses against rBipA from animals infected with TCB-1 and FCB-1. Inoculating mice with each isolate determined that antibodies generated against TCB-1 and FCB-1 BipA were cross reactive against rBipA from B. turicatae 91E135 (Figure 6 A–D). Similarly, in B. hermsii there was sufficient amino acid conservation between BipA from B. hermsii GGI and GGII isolates that mice infected with GG II isolates produced a detectable serological response to rBipA that was expressed from a GG I isolate [5]. Collectively, these results suggest that BipA may be a unique antigen for the given species of relapsing fever spirochete causing infection.
Figure 6. Serological responses from mice infected with TCB-1 (A and B) and FCB-1 (C and D).
B. turicatae protein lysates and rBipA produced from the 91E135 isolate are shown. Molecular masses are indicated to the left of each immunoblot in kilodaltons.
Previous studies by Cadavid et al. reported that BALB/c and SCID mice needle inoculated with the Ozona isolate of B. turicatae developed long-term infections of the brain and joints [12]. Given the persistent nature of the spirochetes within rodents, IgG responses in mice were evaluated one year after transmission by tick bite. Prior to serological analyses, qPCR performed on murine blood samples collected for 10 consecutive days indicated that the mice were no longer spirochetemic (data not shown). Immunoblotting demonstrated that three of four mice continued to generate an IgG response against rBipA one year after the initial exposure, while one animal produced a weakly detectable response (Figure 7 A–D). Interestingly, B. turicatae is no longer detected within the blood of Swiss Webster mice after approximately 14 days after tick bite (data not shown), and with a serum half-life of 20–30 days for IgG, these results suggest a persistent infection and antigen exposure to the host immune response.
Figure 7. Long-term serological responses from four mice one year after tick bite (A–D).
B. turicatae protein lysates and rBipA produced from the 91E135 isolate and B. hermsii are shown. Molecular masses are indicated to the left of each immunoblot in kilodaltons.
Central nervous system (CNS) infections by relapsing fever spirochetes vary between species and genetic variants. Borrelia duttonii can reemerge in the blood from the brain after a period of quiescence [13]. Serotype A of B. turicatae Ozona were neurotropic in mice, while animals infected with serotype B spirochetes colonize the joints and heart [12], [14], [15]. Interestingly, CNS infection caused by B. duttonii and serotype A of B. turicatae Ozona failed to produce noticeable tissue damage [12], [13]. In our study, postmortem necropsies of mice one year after infection by tick bite did not identify inflammatory or degenerative changes within the cerebrum, cerebellum, brain stem, or diarthrodial joints of the fore or hind limbs (data not shown). Spirochetes were also undetectable in tissue sections, and it was unclear if the animals were still infected at the time of euthanasia. However, the persistent antibody responses generated against rBipA can be targeted to increase the likelihood of determining if an animal has been exposed to the spirochetes.
With results suggesting that BipA may be a species-specific antigen, additional studies should evaluate homologues from less characterized yet closely related species to B. turicatae. For example multilocus sequencing indicated that Borrelia johnsonii, a novel species of relapsing fever spirochete that colonizes Carios kelleyi [16], was closely related to B. turicatae and Borrelia parkeri. As additional sequence information is obtained from B. johnsonii and B. parkeri and animal models developed, the diagnostic potential of BipA can be further evaluated as an antigen unique to a given species of relapsing fever spirochete.
The maintenance and ecology of B. turicatae in the southern United States and Latin America is poorly understood, and given the nonspecific clinical symptoms, the disease is likely under reported. Historically, mapping endemic foci has been associated with capturing ticks at sites where human infection occurred and evaluating the arthropods for spirochete colonization, or by obtaining clinical isolates from sick dogs [2], [3], [17]. Pathogen surveillance based on identifying infected ticks can be difficult because O. turicata are nest-, den-, and cave-dwelling with a 5–60 minute bloodmeal [3], [11], [18], and the ticks are rarely identified on the host. We are also unaware of serological surveys for B. turicatae probably due to the degree of antibody cross-reactivity that occurs during spirochete infections [5], [19], [20]. Given the characterization of BipA, serological analyses to identify endemic foci for B. turicatae where rodents and wild canids are monitored as sentinels are possible.
Acknowledgments
We thank Dr. Antonio Ibarra for providing critical comments for this manuscript and Jamie Allison for technical assistance.
Funding Statement
This study was accomplished using startup funds provided by Mississippi State University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Schwan TG, Raffel SJ, Schrumpf ME, Porcella SF (2007) Diversity and distribution of Borrelia hermsii . Emerg Infect Dis 13: 436–442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Schwan TG, Raffel SJ, Schrumpf ME, Policastro PF, Rawlings JA, et al. (2005) Phylogenetic analysis of the spirochetes Borrelia parkeri and Borrelia turicatae and the potential for tick-borne relapsing fever in Florida. J Clin Microbiol 43: 3851–3859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Rawlings JA (1995) An overview of tick-borne relapsing fever with emphasis on outbreaks in Texas. Tex Med 91: 56–59. [PubMed] [Google Scholar]
- 4. Felsenfeld O (1973) The problem of relapsing fever in the Americas. IMS Ind Med Surg 42: 7–10. [PubMed] [Google Scholar]
- 5. Lopez JE, Schrumpf ME, Nagarajan V, Raffel SJ, McCoy BN, et al. (2010) A novel surface antigen of relapsing fever spirochetes can discriminate between relapsing fever and Lyme borreliosis. Clin Vaccine Immunol 17: 564–571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Barbour AG (1984) Isolation and cultivation of Lyme disease spirochetes. Yale J Biol Med 57: 521–525. [PMC free article] [PubMed] [Google Scholar]
- 7. Battisti JM, Raffel SJ, Schwan TG (2008) A system for site-specific genetic manipulation of the relapsing fever spirochete Borrelia hermsii . Methods Mol Biol 431: 69–84. [DOI] [PubMed] [Google Scholar]
- 8. Schwan TG, Hinnebusch BJ (1998) Bloodstream- versus tick-associated variants of a relapsing fever bacterium. Science 280: 1938–1940. [DOI] [PubMed] [Google Scholar]
- 9. McCoy BN, Raffel SJ, Lopez JE, Schwan TG (2010) Bloodmeal size and spirochete acquisition of Ornithodoros hermsi (Acari: Argasidae) during feeding. J Med Entomol 47: 1164–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Lopez JE, Porcella SF, Schrumpf ME, Raffel SJ, Hammer CH, et al. (2009) Identification of conserved antigens for early serodiagnosis of relapsing fever Borrelia. Microbiology 155: 2641–2651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Balashov YS (1972) Bloodsucking ticks (Ixodoidea)- vectors of diseases of man and animals. Misc Publ Entomol Soc Am 8: 161–376. [Google Scholar]
- 12. Cadavid D, Thomas DD, Crawley R, Barbour AG (1994) Variability of a bacterial surface protein and disease expression in a possible mouse model of systemic Lyme borreliosis. J Exp Med 179: 631–642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Larsson C, Andersson M, Pelkonen J, Guo BP, Nordstrand A, et al. (2006) Persistent brain infection and disease reactivation in relapsing fever borreliosis. Microbes Infect 8: 2213–2219. [DOI] [PubMed] [Google Scholar]
- 14. Pennington PM, Allred CD, West CS, Alvarez R, Barbour AG (1997) Arthritis severity and spirochete burden are determined by serotype in the Borrelia turicatae-mouse model of Lyme disease. Infect Immun 65: 285–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Pennington PM, Cadavid D, Barbour AG (1999) Characterization of VspB of Borrelia turicatae, a major outer membrane protein expressed in blood and tissues of mice. Infect Immun 67: 4637–4645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Schwan TG, Raffel SJ, Schrumpf ME, Gill JS, Piesman J (2009) Characterization of a novel relapsing fever spirochete in the midgut, coxal fluid, and salivary glands of the bat tick Carios kelleyi . Vector Borne Zoonotic Dis 9: 643–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Whitney MS, Schwan TG, Sultemeier KB, McDonald PS, Brillhart MN (2007) Spirochetemia caused by Borrelia turicatae infection in 3 dogs in Texas. Vet Clin Pathol 36: 212–216. [DOI] [PubMed] [Google Scholar]
- 18. Adeyeye OA, Butler JF (1989) Population structure and seasonal intra-burrow movement of Ornithodoros turicata (Acari: Argasidae) in gopher tortoise burrows. J Med Entomol 26: 279–283. [DOI] [PubMed] [Google Scholar]
- 19. Magnarelli LA, Anderson JF, Johnson RC (1987) Cross-reactivity in serological tests for Lyme disease and other spirochetal infections. J Infect Dis 156: 183–187. [DOI] [PubMed] [Google Scholar]
- 20. Schwan TG, Schrumpf ME, Hinnebusch BJ, Anderson DE, Konkel ME (1996) GlpQ: an antigen for serological discrimination between relapsing fever and Lyme borreliosis. J Clin Microbiol 34: 2483–2492. [DOI] [PMC free article] [PubMed] [Google Scholar]