

Abstract
This study investigates the predation of P . noctiluca ephyrae on Atlantic Bluefin tuna (ABFT) eggs under different experimental conditions. The specific factors considered in the experimental design were: a) water mix conditions to explore predation under two-dimensional (2D) and three-dimensional (3D) prey distributions, b) prey density to investigate the ingestion rate capacity, and c) incubation time to inspect gut saturation. The eggs and jellyfish ephyrae were collected during the 2012 ABFT spawning survey off Ibiza (Balearic Isl., Western Mediterranean). The results showed that the proportion of feeding ephyrae increased with size. The mean clearance rate of feeding ephyrae, 4.14 L h-1, was the highest ever recorded for ephyrae. Under calm conditions the eggs floated at the surface (2D spatial arrangement) and the clearance rates, at low prey densities, were at least twice those under mixed conditions (3D spatial arrangement). At high prey density, clearance rate did not differ between mix conditions, probably due to the fast gut saturation, which was reached in c.a. 15 min, as revealed by time series observations of gut contents. The fast saturation of ephyrae and their slow digestion time of approximately 18 h suggest the existence of a diel feeding periodicity. We conclude that in the Western Mediterranean, P . noctiluca ephyrae are capable of predating on ABFT eggs, a highly pulsed and spatially restricted resource that potentially switches from a 3D to a 2D configuration in the absence of wind-generated turbulence. The P . noctiluca and Atlantic Bluefin tuna egg system might represent an example of a general mechanism linking pelagic and neustonic food webs.
Introduction
The Atlantic bluefin tuna (hereafter Thunnus thynnus, ABFT) is a highly migratory species targeted on both sides of the Atlantic by different fisheries due to its extraordinary commercial value. This species is composed of two stocks which share Atlantic feeding grounds and migrate to spawn in warmer seas. The Western stock spawns in the Gulf of Mexico [1] and the eastern stock in the Mediterranean [2]. Tuna fishing commenced around 7000 BC in the Mediterranean [3]. Since the 16th century, the early fishing modalities have been replaced gradually by traps [4], and in the late 1990s the increase in market demand and the incorporation of capture-based aquaculture [5] led to a high risk of overfishing. However, the ABFT is certainly exposed to other hazards, since its biogeographic range contracted decades before the rise in fishing intensity. This contraction had been reported since the 1950s [6,7], but the reasons for its disappearance in certain areas are still unclear [8]. In view of the weak condition of the stocks at the beginning of this century, the International Commission for the Conservation of Atlantic Tunas (ICCAT) established a multiannual recovery plan in 2007. However, the rebuilding capacity of fish populations also depends on fishing-independent factors, in particular those responsible for recruitment success or offspring survival during the early life stages of development. Mortality during the egg and larval stages is thought to play a major role in determining survival and latter recruitment into adult stocks [9,10]. There are a wide variety of factors inducing fish egg mortality [11-13], although predation is generally considered the main cause [14-16].
Jellyfishes are top planktonic predators [17], generally considered detrimental to fish populations through competition for zooplankton prey [18] or directly by predation on fish eggs and larvae [19]. Jellyfish predation on fish eggs and larvae has been confirmed from results of experimental studies [20-23]. In addition, a positive selection for ichthyoplankton has been reported [24-27]. In the Mediterranean, P . noctiluca , a warm-water holoplanktonic scyphomedusae [28], is abundant and widespread in various regions [29-33], showing large fluctuations with massive outbreaks correlated with warm, dry conditions [34]. This species represents the most abundant and most venomous jellyfish in the Mediterranean [35]. P . noctiluca is an opportunistic predator [31]; therefore, its impact on fish populations may vary with regions and will depend on its spatial and temporal overlap with the early life stages of fish populations.
The impact of P . noctiluca on ABFT spawning is unknown. However, increasing evidence of their spatio-temporal overlap prompts an evaluation of this predation potential. The abundance of this jellyfish in Balearic waters is increasing and its swarms enter the Mediterranean following the progression of Atlantic surface waters [36]. This is the same migratory displacement process and route known for ABFT spawning [4,37,38]. Balearic waters represent the most important ABFT spawning area in the Mediterranean [39,40], where spawning seems to takes place mostly in offshore mixed waters of the frontal areas in the confluence of Atlantic and Mediterranean water masses [41]. There is evidence from different studies that higher densities of this jellyfish species also occur in the vicinity of frontal areas [27,30]. The results from field studies on spawning behavior, aboard ABFT transport cages in Balearic waters, showed a clear nocturnal concurrence of ABFT spawning and P . noctiluca larvae at the sea surface [42], where jellyfish larvae appeared to exhibit predatory behavior on ABFT eggs.
Solid experimental evidence of predation of P . noctiluca ephyrae on ABFT eggs must be secured before attempting any speculation on the potential impact of this jellyfish on ABFT populations. The diurnal scarcity and nocturnal abundance of P . noctiluca at the sea surface in the ABFT spawning region [42] is indicative of the well-known pattern of diel vertical migration of this species [30,35,43,44], enabling foraging throughout a wide range of water column depths. In contrast, freshly laid ABFT eggs are positively buoyant and rapidly accumulate on a single layer at the surface of experimental beakers (Gordoa, unpublished observation). P . noctiluca captures prey while cruising [30], a feeding mechanism which seems hardly compatible with a two-dimensional prey distribution [45].
ABFT transport cages transiting Balearic waters are used to monitor ABFT spawning patterns and serve as a unique opportunity to concurrently obtain live specimens of ABFT eggs and P . noctiluca for an experimental test of predation. In June 2012, we used this opportunity to conduct predation experiments in a portable laboratory in a coastal aquarium-cave in the island of Ibiza (W Mediterranean). In this study we investigated the predation of P . noctiluca ephyrae on ABFT eggs under different experimental conditions. We explored the role of positive egg buoyancy as a strategy to evade ephyrae predation by measuring predation rates in mixed experimental beakers, where the eggs had a 3D distribution, and still experimental beakers, where eggs accumulated at the surface (2D). We also tested the effect of prey density and incubation time on predation rates and digestion time. Video observations were recorded to provide supplementary information on predatory capture conducts. Our results provide experimental evidence of predation of P . noctiluca ephyrae on ABFT eggs, using highly efficient alternative predation mechanisms for 3D (water column) and 2D (neuston) prey fields.
Methods
Experiment logistics
Experiments were conducted at the Cap Blanc Aquarium, a coastal cave located in Ibiza (Figure 1). The permission to perform the study on this site was given by the town council of Sant Antoni de Portmany (Ibiza) and the Aquarium Cap Blanc, a private company that holds the concession of the aquarium. This location was chosen for its proximity to the ABFT spawning grounds, in order to minimize transport time of jellyfish and ABFT eggs. The samples were collected during the 2012 spawning survey using a bongo frame fitted with 0.3 mm mesh nets and towed from the rear of an ABFT transport cage at a fixed depth of 3 m [42]. The cage was moved close to the aquarium during 13th to 27th June to facilitate transport of specimens to the field laboratory.
Figure 1. Map showing the location of the field laboratory (●) and of the tuna transport cage (▲).

Sampling was carried out at night during the ABFT spawning window (2: 00 to 5: 00 a.m. local time). Live fish eggs and jellyfish were carefully transferred to transport containers filled with 0.3 mm filtered water for transport to the field laboratory.
Experiment layout
Prior to the experiments, the eggs were kept in two 40-L aerated aquaria and the jellyfish in two 6-L aquaria without aeration. All the aquaria were filled with 3 mm filtered seawater adjusted to the seawater temperature at the time of sampling (23 ± 0.3 °C). Every two days, the aquaria were cleaned and filled with fresh eggs. Due to space limitations, replicated experimental treatments were not done simultaneously but sequentially one at a time, between 10:00 and 18:00, with replicates adequately interspersed in time.
The study considered four treatments arranged in a 2-way orthogonal design with a prey density factor with high (12 eggs L-1) and low (6 eggs L1) density levels and a mixing regime factor with two levels, stirred and still experimental suspension (these treatments led to a homogeneous, 3D distribution of the eggs within the experimental beaker and to a 2D surface accumulation, respectively) (Table 1). The low egg concentration was selected according to other similar experiments with gelatinous predators [14,46] to ensure being in the linear segment of the functional response curve, and doubling the prey density to enable a differential concentration response. Gentle mixing conditions were achieved by using a methacrylate comb with 5 teeth, each 1.5 cm wide and 1.5 cm apart and penetrating 2 cm into the water. The comb was connected to an electric motor rotating at 10 rpm and changing direction every 2 seconds.
Table 1. List of treatments used in the experiments.
| Incubation time |
||||||
|---|---|---|---|---|---|---|
| Mixing regime | Egg concentration | 5’ | 10’ | 15’ | 25’ | 120’ |
| unstirred | low (6 egg L-1) | 2(u) | 2(u) | 4(s) | 3(s) | 4(s) |
| unstirred | high (12 egg L-1) | 3(s) | ||||
| stirred | low (6 egg L-1) | 4(u) | ||||
| stirred | high (12 egg L-1) | 4(s) | ||||
Number of replicates for each treatment and whether the jellyfish guts were under saturated (s) or unsaturated (u) conditions with Atlantic Bluefin Tuna eggs, according to our results.
We used 10 jellyfish per incubation, which lasted 15 minutes. The scarcity of P . noctiluca in 2012 imposed limitations on the availability of ephyrae of different sizes for experimentation. Although the size range of collected jellyfish was 3-13 mm, most of them were smaller than 7 mm (median = 5 mm): consequently, bigger individuals were under-represented in the experiments. A known number of eggs were added to 6-L experimental beakers after being counted on a Petri disc. The eggs were gently spread over the water surface, and under still experimental conditions the ephyrae were introduced immediately afterwards, while under stirred conditions they were introduced once the eggs were fully mixed in the beaker. At the end of each experiment each jellyfish was removed from the tanks and placed alive under a stereo-microscope for measuring and counting the numbers of eggs inside its gut.
Also interspersed with the treatments of the previous experiment, we examined the effect of incubation time at a prey concentration of 6 eggs L1 without stirring (Table 1). For this, we conducted pairs of replicate incubations lasting 5, 10, 25 and 120 minutes, plus the four 15-minute replicates of the previous experiment (Table 1). Randomly and from different treatments, some of the jellyfish which had eggs in their guts after the incubation were kept in sealed 50-mL bottles and examined every 5 hours to monitor egg digestion and/or regurgitation, allowing a crude estimation of digestion time.
Calculation of feeding rates
For each of the experimental beakers, individual ingestion rates were calculated as
where e is the sum of eggs counted inside the guts of all larvae in the incubation, t is the incubation time and N is the total number of ephyrae which were feeding during the incubation.
Population clearance rates were calculated following Moller et al. (2010) [47] as
where V is the volume of water in the incubation container (L) and C i and C f are the initial and final egg concentrations (eggs L-1) in the incubation flask. C i was calculated as the initial concentration of eggs in the incubation flask, while C f was estimated by subtracting the number of eggs observed in the guts of the ephyrae from the initial number of eggs in the container and dividing by V.
Statistical analysis
The response of the proportion of feeding ephyrae (p), the individual ingestion rate (IR) and the individual clearance rate (CR) to mixing conditions and egg concentrations was analyzed with a two-way ANCOVA with mean ephyra size as covariate. Variables were log-transformed to account for allometric power relationships between the feeding rates and body length, and were checked for normality, homogeneity of variances and homogeneity of slopes prior to analysis. In the case of IR, there was a significant difference among regression slopes between treatments (P=0.04). However, the regression lines showed good fits and appeared nearly parallel on visual inspection, so we followed Quinn & Keough [48] and applied ANCOVA under an assumption of homogeneity of slopes. In addition, there were differences in the range of ephyrae size between treatments, so our ANCOVA adjustment for the effect of size involves some degree of extrapolation outside the ephyra size range for each treatment. Finally, one replicate was lost, which led to an unbalanced design, dealt with by using a type III sum of squares.
Results
After the experimental incubations, not all the jellyfish had eggs inside their gastric pouches, indicating that not all were feeding actively on ABFT eggs. The proportion of feeding ephyrae increased with size, with nearly all individuals larger than 8 mm containing ABFT eggs in their guts (Figure 2). In the experiments, the number of feeding ephyrae varied between 0 and 7 out of the ten animals used per incubation. The proportion of feeding ephyrae depended significantly on mean ephyrae size (P=0.030) and on egg concentrations (P=0.020), but was unaffected by mixing conditions (P>0.05, Table 2, Figure 3).
Figure 2. Proportion of feeding ephyrae in all of the experimental beakers for each of 7 different size classes.

The first and the last classes are open. The numbers on top of each bar indicate the number of individuals.
Table 2. ANCOVA table for the ephyrae response to different treatments.
| Source | df | MS | F | P | |
|---|---|---|---|---|---|
| Dependent variable: p | |||||
| egg density | 1 | 0.25 | 7.63 | 0.020* | |
| mixing | 1 | 0.02 | 0.37 | 0.444 | |
| egg density x mixing | 1 | 0.07 | 2.03 | 0.185 | |
| logsize (all ephyrae) | 1 | 0.21 | 6.42 | 0.030* | |
| Dependent variable: IR | |||||
| egg density | 1 | 0.03 | 15.38 | 0.040* | |
| mixing | 1 | 0.07 | 28.67 | 0.004* | |
| egg density x mixing | 1 | 0.03 | 6.83 | 0.026* | |
| logsize (feeding ephyrae) | 1 | 0.39 | 79.70 | 0.000* | |
| Dependent variable: CR | |||||
| egg density | 1 | 0.19 | 16.27 | 0.002* | |
| mixing | 1 | 0.08 | 6.39 | 0.030* | |
| egg density x mixing | 1 | 0.07 | 5.66 | 0.039* | |
| logsize (feeding ephyrae) | 1 | 0.54 | 45.20 | 0.000* |
Dependent variables were the proportion of feeding ephyrae (p , the individual ingestion rate (IR) and the individual clearance rate (CR). It is a two-way, fully orthogonal design with two fixed factors, mixing regime and food concentration, and mean ephyra size as covariate. Asterisk indicates significant (<0.05) P-values.
Figure 3. Effect of egg concentration and mixing regime on the proportion of feeding ephyrae (p), egg ingestion rate (IR) and egg clearance rate (CR) of P . noctiluca ephyrae feeding on Atlantic Bluefin tuna eggs.
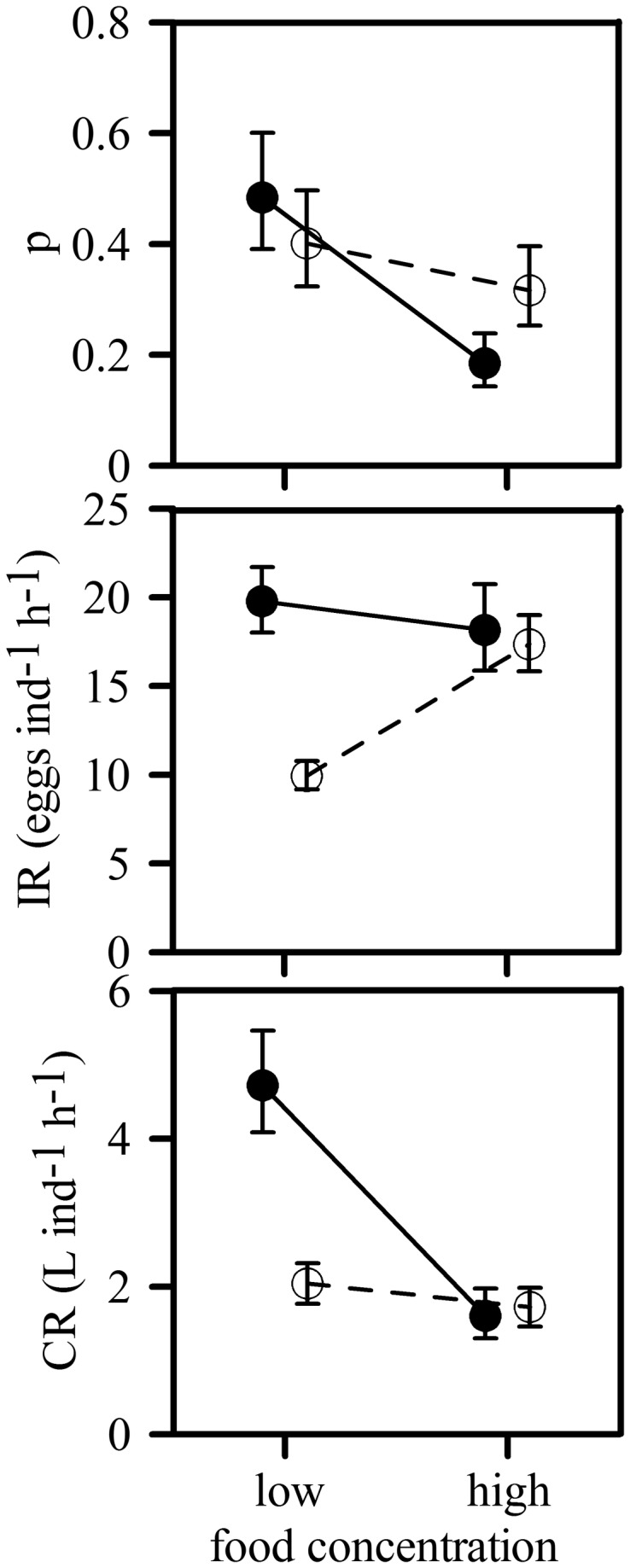
Dots represent geometric means adjusted to the mean ephyra size, which was 0.50 mm for p (includes all ephyrae used in each incubation) and 0.66 mm for IR and CR (includes only feeding ephyrae). Deviations correspond to ±10SE, where SE is the standard error of the log-transformed variables. Filled and empty dots correspond to the unstirred and stirred treatments, respectively.
IR was significantly affected by the average size of the feeding ephyrae (P<0.001) and by a significant interaction between egg concentration and mixing regime (P=0.026). At low egg concentrations, ephyrae in unstirred beakers had almost twice as many eggs in their guts as ephyrae in stirred beakers (Figure 3). Thus, the ephyrae were particularly apt at removing the eggs from the surface, where they accumulated in the absence of stirring, as is clearly shown in video recordings made during the experiments (Movie S1). However, this difference disappeared at the high food concentrations, resulting in similar IR in both the stirred and the unstirred beakers. This is probably due to the saturation of the digestive system of ephyrae (Figure 4A). Video observations revealed differences in predation mechanisms between mixing regimes: the ephyrae used the subumbellar tentacles when the eggs where homogeneously distributed in the water column and the marginal lappets when the eggs were at the surface (Figure 4B-C).
Figure 4. Ephyrae with eggs in different situations: A) ephyra on a Petri disc after feeding, B) ephyra capturing eggs during feeding incubation under stirred conditions, C) ephyra capturing eggs during feeding incubation under unstirred conditions.

Ephyrae cleared eggs at rates of up to 6.75 L individual-1 h-1. The CR was significantly affected by the average size of the feeding ephyrae (P<0.001) and by the interaction between egg concentration and mixing regime (P=0.039, Table 2). The CR of ephyrae in the unstirred beakers was more than twice that of ephyrae in stirred beakers at the low egg concentration (Figure 3). Thus, the larvae were more efficient at removing the 36 eggs used per incubation when accumulated in 60 cm2 of water surface than when suspended in the 6 L of water volume. However, this difference disappeared when egg concentration was high, again suggesting some sort of limitation, probably due to gut saturation.
Although our ANCOVA analysis may to some extent be hindered by differences between treatments in the size range of the ephyrae (see methods), it is fully consistent with the results of the experiment testing the effect of incubation time. The proportion of feeding ephyrae was quite constant for the different incubation times (Figure 5). The number of eggs inside the guts of the ephyrae increased during the first 15 minutes of incubation and showed symptoms of saturation afterwards, at a gut content of nearly 3 eggs ind-1 (Figure 5). It should be noted that the conditions during this experiment corresponded to the low egg density/no stirring treatment used in the previous experiment, where we already found symptoms of saturation with an ingestion rate close to 20 eggs ind-1 h-1 (Figure 3). This ingestion rate is equivalent to a gut content of ca. 5 eggs ind-1 after an incubation time of 15 minutes. The ingestion and clearance rates declined after 15 minutes of incubation (Figure 5), basically because the number of eggs inside the guts remained nearly constant, while the incubation time increased.
Figure 5. Effect of incubation time on the proportion of feeding ephyrae (p), the number of eggs inside the gastric pouches after incubation, the ingestion rate (IR) and the Clearance Rate (CR).
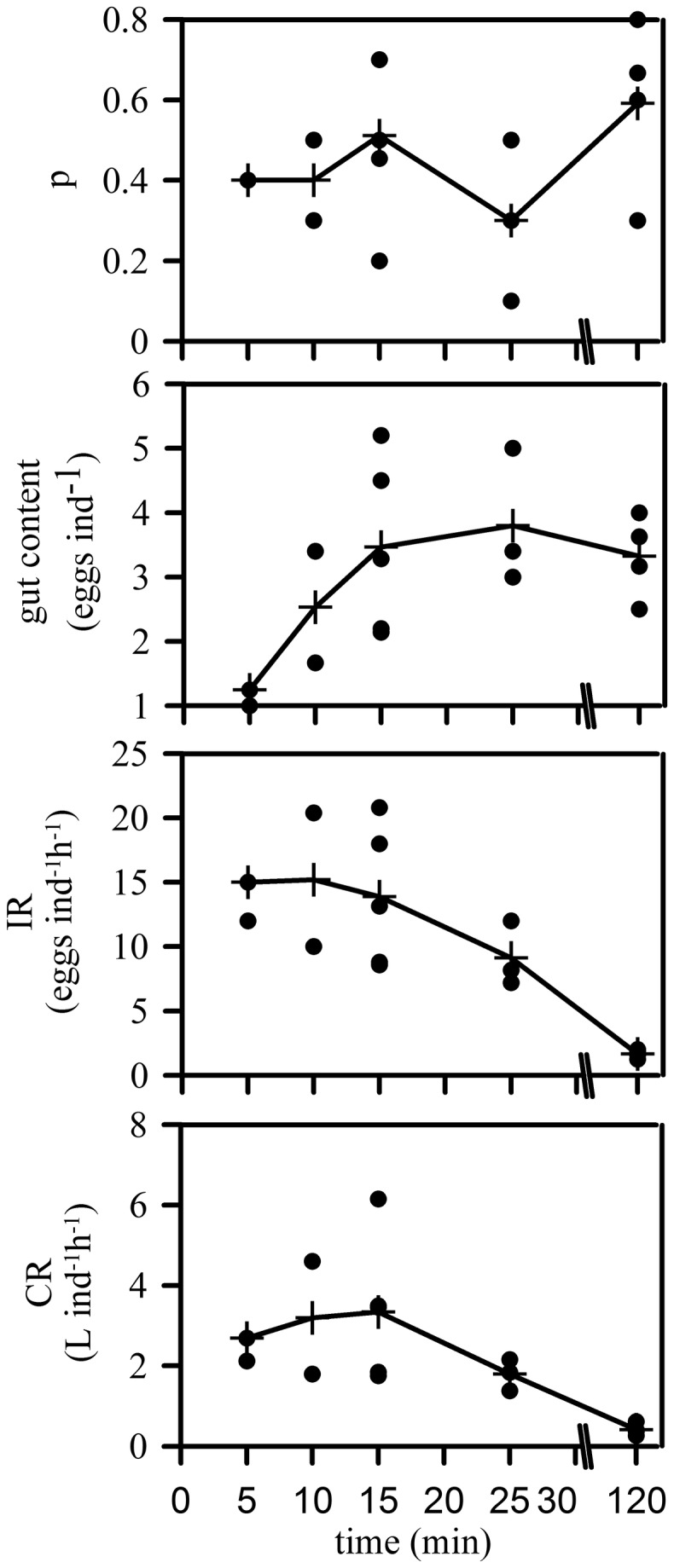
Dots are observations (beakers), and crosses represent means for each incubation time. Crosses have been joined by lines to visualize tendencies.
The digestion time was not significantly related to the size of the ephyra (log-log regression of digestion time on ephyra length, P > 0.05, Figure 6). Therefore, an average digestion time of 18.55±5.26 h can be assumed for all ephyra sizes within the studied size range (Figure 6). This is much longer than the incubation times of our experiments and implies that once full of eggs, the ephyrae must stop ingesting at least until there is sufficient room again for a new egg or, more conservatively, until all the gut contents have been digested. In this scenario, and as a first-order approximation, it follows that the predation rates of P . noctiluca ephyrae on ABFT eggs should be encapsulated between two extremes: saturating and non-saturating conditions. Under saturating conditions, it is the egg volume and the digestion time that limits the ingestion rate. Under non-saturating conditions, it is the clearance potential and the prey concentration that matters. We will make an attempt at calculating the ingestion rate in saturating conditions and the clearance rate in unsaturated conditions. For this purpose, each animal and its gut content is considered separately, rather than in batches as for the preceding calculations.
Figure 6. Digestion time (T), average long-term ingestion rate under saturating conditions () and clearance rate under unsaturating conditions (CRuns) vs ephyra length.

The solid line indicates: in the upper panel, the mean T (18.55±5.26 h); in the middle panel, the least-squares power regression (log (, eggs ind-1 h-1)=(-0.305) +(2.05) log (ephyra length, mm), n=70, R2=0.65, P<0.001); in the lower panel, the least-squares power regression (log (CRuns, L ind-1 h-1)=(0.529) +(0.934) log (ephyra length, mm), n=35, R2=0.176, P=0.006). Animals used for calculation of the and the CRuns belong to experimental treatments labeled "(s)" and "(u)" in Table 1.
An average long-term ingestion rate in saturating conditions (, eggs ind-1 h-1) can be calculated by assuming that complete digestion is necessary to resume predation. Under these conditions:
where e sat is the number of eggs inside the full gut of an individual and T is the digestion time. increased rapidly with ephyra length, according to the power relationship; (eggs in-1 h-1) = 0.5[ephyra length (mm)]2.05 (Figure 6).
In unsaturated conditions, the animals would be clearing prey with maximum rates, which can be easily calculated from the number of eggs inside their guts and the egg concentrations in the beakers at the beginning and end of the incubations. Firstly, an instantaneous individual ingestion rate in unsaturated conditions (IRuns, eggs h-1 individual-1) can be calculated as:
where e uns is the number of eggs inside the ephyra after incubation under non-saturating conditions and t is incubation time. Methods in Møller et al. (2010) [47] can then be adapted to estimate an individual clearance rate (L h-1 individual-1) as:
where C m is the mean egg concentration experienced by the ephyrae inside the experimental container during the course of the incubation [47]. C m can be estimated as:
CRuns calculated using these equations grew almost isometrically with ephyra length, according to the power law CRuns (L ind-1 h-1) = 3.4[ephyra length (mm)]0.93 (Figure 6).
Discussion
The results of this study clearly demonstrate the predatory capacity of P . noctiluca on ABFT eggs. Overall, we found four specific experimental results to be discussed in the context of the coupled spatio-temporal distribution patterns of both species in the study area. Firstly, in the lab, large ephyrae are voracious predators of ABFT tuna eggs, with remarkably high clearance rates. Secondly, those clearance rates are sustained or even increased when the eggs are accumulated on the surface of the incubation containers rather than suspended in the water. Thirdly, the ingestion rate of P . noctiluca ephyrae rapidly saturates, leading to cessation of their prey capture behavior. Finally, P . noctiluca ephyrae are likely to resume predation after a long digestion time of approximately 18 h.
To our knowledge, P . noctiluca exhibited among the highest clearance rates reported for jellyfish of a similar size (Table 3). This difference may be in part methodological, as most previous studies measured predation rates by counting the number of prey inside the experimental container before and after incubation (but see 49). However, non-feeding individuals are not detected by this approach, thereby underestimating actual individual clearance rates. Thus, in the present study, the gut contents of all individual ephyrae were inspected right after the incubation, to readily detect non-feeding individuals.
Table 3. Experimentally measured clearance rates of small scyphomedusae.
| Reference | Predator Species | Predator size | Prey | Prey size/life stage | Clearance rate |
|---|---|---|---|---|---|
| Hansson & Kiørboe 2006 | Sarsiatubulosa | - | copepods | - | *0.04-0.10 L h-1 |
| Tilves et al. 2013 | Pelagianoctiluca | 3.8-4.2 mm | Mnemiopsisleidy | 1-3 mm | 0.4 L h-1 |
| 20-25 mm | 8-12 mm | 62 L h-1 | |||
| Morand et al. 1987 | Pelagianoctiluca | 8 mm | Artemia | nauplii | *0.13 L h-1 |
| Riisgård & Madsen | Aurelia aurita | 5.1-5.9 mm | Artemia | 0.9 mm | 0.05-0.09 L h-1 |
| Bailey & Batty 1984 | Aurelia aurita | 20-22 mm | Clupeaarengus | Yolksac | 2.2-5.5 L h-1 |
| Fancett &Jenkins 1988 | Cyanea capillata | 25 mm | Paracalunus indicus | - | 0-2.5 L h-1 |
| This study | Pelagianoctiluca | 5 mm | Thunnus thynnus | 1 mm | 0.73-7.54 L h-1 |
Converted from original units L d- 1
In this study, the individual ephyrae cleared actively during rapid ca. 15 min bursts, but halted feeding afterwards. Accordingly, an assumption of steady-state ingestion rate has to be rejected for P . noctiluca at the prey concentrations and conditions of our experiments, a situation which is similar to that observed by Hanson & Kiørboe ( [50]) in the jellyfish Sarsiatubulosa . Our study lends support to their idea that jellyfish use their large gastric pouches as a buffering, food-accumulating organ allowing bursts of high food intake when prey is available in short-lived pulses, as is the case of ABFT eggs during spawning events in the field. During the spawning season in 2012, the average weight of the captured ABFT shoals was 30 t, with a maximum of around 120 t [51]. Spawning takes place at night, with the exact timing varying between ABFT groups, but always taking place within a small temporal window (0: 00 to 3: 00 GMT). Spawning events last about ten minutes, during which the shoals stop horizontal displacement and stay at the surface. According to previous calculations [52] based on average fecundities [53], a 30 t shoal should produce around 840 million eggs within a diameter of approximately 40 m (personal observation). As a result, local egg densities of ca. 668 449 eggs m-2 can be reached in the natural environment, 263 times higher than the highest density used in this study. Thus, potential predators present at the right time and location will suddenly experience an extraordinarily abundant prey source before it is dispersed by ocean currents. Clearly, high capture efficiencies and large guts should be highly beneficial under these circumstances.
Short, intense predation episodes by the ephyrae on this pulsed food source might be relevant in their life cycle, in particular in the oligotrophic waters of the Balearic archipelago [54]. This could be especially important for a species like P . noctiluca , which has no benthic stage and therefore depends exclusively on surface-dwelling prey. It is worth noting that the digestion time of the ephyrae is close to the duration of one day. This suggests that ephyrae digestion times and feeding rates are metabolic adaptations to a diel feeding cycle. This supposition is consistent with the pattern of diel vertical migration in P . noctiluca [35,43], most probably related to feeding [55].
The highly positive buoyancy of ABFT eggs does not represent a strategy of floating on the sea surface and evading predation by jellyfish and other pelagic predators. In particular, P . noctiluca is a typical cruising predator that screens the water following a sinusoidal search path [56], generating feeding currents around its umbrella and feeding tentacles [57]. A priori, this mechanism seems suited to operating in a 3D, not 2D, prey distribution. However, our experiments show that P . noctiluca ephyrae can sustain higher apparent clearance rates when ABFT eggs are accumulated at the water surface, allowing for rejection of our initial hypothesis. Nevertheless, our measured feeding rates in a 2D prey distribution might overestimate feeding rates in the field, because water motions generated by the feeding ephyrae may well have altered the uniform distribution of eggs towards the edges of the beakers, enhancing their catchability. Our own visual observations during the incubations do not suggest that this may have been a major source of bias. Video observations suggest that this species switches from cruising tactile predation with capture at the subumbellar tentacles (Movie S2) to some kind of directed prey-detection mechanism with capture at the marginal lappets (Figure 3B-C, Movie S1). Interestingly, adult P . noctiluca exhibit a different capture system for non-motile prey at the water surface, based on positioning their bells upside down and moving their oral arms towards the surface film [56]. Video observations illustrated ephyrae feeding behavior, exhibiting controlled and directed movements between ephyrae to displace the competitor in the pursuit of a prey (Movie S1).
Theory indicates that predators should have shorter searching times and higher consumption rates in a 3D than in a 2D prey distribution [45]. This does not seem to apply for P . noctiluca ephyrae, which seem to be capable of easily switching from a 3D to a 2D configuration. Neustonic 2D predation may increase predation success on an egg population that has already been dispersed but that floats at the surface in the absence of wind. On more general grounds, the P . noctiluca -ABFT egg system might represent an example of a general mechanism linking pelagic and neustonic food webs. In the absence of wind forcing, planktonic organisms that swim upwards or that have positive buoyancy will accumulate on the surface, thus becoming a valuable food source for both obligate (e.g. Physalia Physalis , Velellavelella or Porpitaporpita ) and facultative neustonic predators (e.g. P . noctiluca ). The onset of wind-generated turbulence on the sea surface will bring many of those organisms back into suspension, where pelagic predators will be at an advantage.
We conclude that in the Western Mediterranean, P . noctiluca ephyrae are functionally well suited to predating on ABFT eggs, a highly-pulsed, spatially-restricted resource that potentially switches from a 3D to a 2D configuration in the absence of wind-generated turbulence. However, the potential impact of P . noctiluca on ABFT early life stages must be verified against field observations in future studies. Moreover, the possibility that the annual cyclic fluctuations of ABFT [4] could be related to those observed in P . noctiluca [34,58,59] should also be investigated.
Supporting Information
A video showing the interaction between two ephyrae of Pelagianoctiluca at the time of catching Atlantic Bluefin tuna eggs used as prey under unstirred experimental conditions.
(WMV)
Video shots illustrating: 3D distribution of eggs under mixing condition, ephyra catching eggs with subumbellar tentacles under mixing conditions and ephyra under a stereo-microscope after feeding experiment.
(WMV)
Acknowledgments
We thank the Balfegó Group S.L. for their continued support and cooperation with the Centro de Estudios Avanzados de Blanes (CEAB), Spanish National Research Council (CSIC), facilitating the tuna transport fleet and tuna spawners to carry out this study. We want to thank the captains and crews of the tuna transport vessels “Estela Nova” and “Coyote” for their assistance and commitment during the sampling. Thanks to Gustavo Carreras for his essential and permanent technical support. We are especially grateful to the town council of Sant Antoni de Portmany (Ibiza) and the Aquarium Cap Blanc for offering their facilities and we highly appreciate the assistance of the staff of the aquarium. Finally, we thank Paul Martin for his assistance in the final review of the text of this article.
Funding Statement
This study is part of the project “Spatio-temporal overlap and trophic interactions between Atlantic Bluefin Tuna spawning and jellyfish in the Balearic spawning grounds” funded by the Spanish Ministry of the Economy and Competitiveness. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. McGowan MF, Richards WJ (1989) Bluefin tuna, Thunnus thynnus, larvae in the Gulf Stream off the southeastern United States: Satellite and shipboard observations of their environment. Fish Bull 87: 615-631. [Google Scholar]
- 2. Richards WJ (1976) Spawning of bluefin tuna (Thunnus thynnus) in the Atlantic Ocean and adjacent seas. ICCAT Col Vol Sci Pap 5: 267-278
- 3. Desse J, Desse-Berset N (1994) Osteometry and fishing strategies at Cape Andreas Kastros (Cyprus, 8th millennium BP). Annalen-Koninklijk Museum voor Midden-Afrika-Zoologische Wetenschappen 274: 69-79
- 4. Ravier C, Fromentin J (2001) Long-term fluctuations in the eastern Atlantic and Mediterranean bluefin tuna population. ICES J Mar Sci 58: 1299-1317. doi:10.1006/jmsc.2001.1119. [Google Scholar]
- 5. Miyake P, De la Serna J, Di Natale A, Farrugia A, Katavic I et al. (2003) General review of bluefin tuna farming in the Mediterranean area. Col: ICCAT Vol Sci Pap 55: 114-124. [Google Scholar]
- 6. Collette BB, Carpenter KE, Polidoro BA, Juan-Jordá MJ, Boustany A et al. (2011) High value and long life-double jeopardy for tunas and billfishes. Science 333: 291-292. doi:10.1126/science.1208730. PubMed: 21737699. [DOI] [PubMed] [Google Scholar]
- 7. Worm B, Tittensor DP (2011) Range contraction in large pelagic predators. Proc Natl Acad Sci USA 108: 11942-11947. doi:10.1073/pnas.1102353108. PubMed: 21693644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. MacKenzie BR, Mariani P (2012) Spawning of bluefin tuna in the Black sea: historical evidence, environmental constraints and population plasticity. PLOS ONE 7(7): e39998. doi:10.1371/journal.pone.0039998. PubMed: 22848367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Hunter JR (1976) Culture and growth of northern anchovy, Engraulis mordax, larvae. Fish Bull 74: 81-88. [Google Scholar]
- 10. Theilacker GH (1986) Starvation-induced mortality of young sea-caught jack mackerel, Trachurus symmetricus, determined with histological and morphological methods. Fish Bull 84: 1-18. [Google Scholar]
- 11. Browman HI (2003) Assessing the impacts of solar ultraviolet radiation on the early life stages of crustacean zooplankton and ichthyoplankton in Marine coastal systems. Estuaries 26: 30-39. doi:10.1007/BF02691691. [Google Scholar]
- 12. Cameron P, von Westernhagen H (1997) Malformation rates in embryos of North Sea fishes in 1991 and 1992. Mar Pollut Bull 34: 129-134. doi:10.1016/S0025-326X(96)00069-0. [Google Scholar]
- 13. Meneses I, Vendrell C, Stratoudakis Y (2003) Mackerel (Scomber scombrus) eggs parasitized by Ichthyodinium chabelardi in the north-east Atlantic: an overlooked source of mortality. J Plankton Res 25: 1177-1181. doi:10.1093/plankt/25.9.1177. [Google Scholar]
- 14. Hunter JR (1976) Report of a colloquium on larval fish mortality studies and their relation to fishery research, January 1975. Department of Commerce, NOAA, National Marine Fisheries Service; pp. 1-5. [Google Scholar]
- 15. Lasker R (1985) What limits clupeoid production? Can J Fish Aquat Sci 42: 31-38. doi:10.1139/f85-260. [Google Scholar]
- 16. Sissenwine MP (1984) Why do fish populations vary? Berlin Heidelberg: Springer Verlag; pp. 59-94. [Google Scholar]
- 17. Compte J, Gascon S, Quintana XD, Boix D (2010) Top-predator effects of jellyfish Odessia maeotica in Mediterranean salt marshes. Mar Ecol Prog Ser 402: 147-159. doi:10.3354/meps08453. [Google Scholar]
- 18. Purcell JE, Sturdevant MV (2001) Prey selection and dietary overlap among zooplanktivorous jellyfish and juvenile fishes in Prince William Sound, Alaska. Mar Ecol Prog Ser 210: 67-83. doi:10.3354/meps210067. [Google Scholar]
- 19. Brodeur RD, Sugisaki H, Hunt GL Jr (2002) Increases in jellyfish biomass in the Bering Sea: implications for the ecosystem. Mar Ecol Prog Ser 233: 89-103. doi:10.3354/meps233089. [Google Scholar]
- 20. Bailey KM (1984) Comparison of laboratory rates of predation on five species of marine fish larvae by three planktonic invertebrates: Effects of larval size on vulnerability. Mar Biol 79: 303-309. doi:10.1007/BF00393262. [Google Scholar]
- 21. Duffy JT, Epifanio CE, Fuiman LA (1997) Mortality rates imposed by three scyphozoans on red drum (Sciaenops ocellatus Linnaeus) larvae in field enclosures. J Exp Mar Biol Ecol 212: 123-131. doi:10.1016/S0022-0981(96)02741-4. [Google Scholar]
- 22. Lynam CP, Heath MR, Hay SJ, Brierley AS (2005) Evidence for impacts by jellyfish on North Sea herring recruitment. Mar Ecol Prog Ser 298: 157-167. doi:10.3354/meps298157. [Google Scholar]
- 23. Titelman J, Hansson LJ (2006) Feeding rates of the jellyfish Aurelia aurita on fish larvae. Mar Biol 149: 297-306. doi:10.1007/s00227-005-0200-5. [Google Scholar]
- 24. Fancett, Jenkins GP (1988) Predatory impact of scyphomedusae on ichthyoplankton and other zooplankton in Port Phillip Bay. J Exp Mar Biol Ecol 116: 63-77. doi:10.1016/0022-0981(88)90246-8. [Google Scholar]
- 25. Graham WM, Kroutil RM (2001) Size-based prey selectivity and dietary shifts in the jellyfish, Aurelia aurita . J Plankton Res 23: 67-74. doi:10.1093/plankt/23.1.67. [Google Scholar]
- 26. Purcell JE, Nemazie DA, Dorsey SE, Houde ED, Gamble JC (1994) Predation mortality of bay anchovy Anchoa mitchilli eggs and larvae due to scyphomedusae and ctenophores in Chesapeake Bay. Mar Ecol Prog Ser 114: 47-58. doi:10.3354/meps114047. [Google Scholar]
- 27. Sabatés A, Pagés F, Atienza D, Fuentes V, Purcell JE et al. (2010) Planktonic cnidarian distribution and feeding of Pelagia noctiluca in the NW Mediterranean Sea. Hydrobiologia 645: 153-165. doi:10.1007/s10750-010-0221-z. [Google Scholar]
- 28. Russell FS (1970) The Medusae of the British Isles II: Pelagic Scyphozoa with a supplement to the first volume on hydromedusae. London: Cambridge University Press. 283pp. [Google Scholar]
- 29. Bernard P, Berline L, Gorsky G (2011) Long term (1981-2008) monitoring of the jellyfish Pelagia noctiluca (Cnidaria, Scyphozoa) on the French Mediterranean Coasts. J Oceanography Res Data 4: 1-10. [Google Scholar]
- 30. Ferraris M, Berline L, Lombard F, Guidi L, Elineau A et al. (2012) Distribution of Pelagia noctiluca (Cnidaria, Scyphozoa) in the Ligurian Sea (NW Mediterranean Sea). J Plankton Res 34: 874-885. doi:10.1093/plankt/fbs049. [Google Scholar]
- 31. Malej A (1989) Shifts in size classes and structure of net zooplankton in the presence of the jellyfish Pelagia noctiluca (Scyphozoa). Nova Thalassia 10: 89-97. [Google Scholar]
- 32. Morand P, Goy J, Dallot S (1992) Recrutement et fluctuations à long-terme de Pelagia noctiluca (Cnidaria, Scyphozoa). Ann Inst Oceanogr. 68: 151-158. [Google Scholar]
- 33. Vucetic T (1983) Some causes of the blooms and unusual distribution of the jellyfish Pelagia noctiluca in the Mediterranean (Adriatic). Proceedings of the Workshop on Jellyfish Blooms in the Mediterranean. Athens. pp. 167-176. [Google Scholar]
- 34. Goy J, Morand P, Etienne M (1989) Long-term fluctuations of Pelagia noctiluca (Cnidaria, Scyphomedusa) in the western Mediterranean Sea. Prediction by climatic variables. Deep Sea Res 36: 269-279. doi:10.1016/0198-0149(89)90138-6. [Google Scholar]
- 35. Mariottini GL, Giacco E, Pane L (2008) The mauve stinger Pelagia noctiluca (Forsskal, 1775). Distribution, ecology, toxicity and epidemiology of stings. Mar Drugs 6: 496-513. doi:10.3390/md6030496. PubMed: 19005582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Licandro P, Conway DVP, Daly Yahia MN, Fernandez de Puelles ML, Gasparini S et al. (2010) A blooming jellyfish in the northeast Atlantic and Mediterranean. Biol Lett 6: 688-691. doi:10.1098/rsbl.2010.0150. PubMed: 20375044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Block BA, Dewar H, Blackwell SB, Williams TD, Prince ED et al. (2001) Migratory movements, depth preferences, and thermal biology of Atlantic bluefin tuna. Science 293: 1310-1314. doi:10.1126/science.1061197. PubMed: 11509729. [DOI] [PubMed] [Google Scholar]
- 38. Mather FJ, Mason JM, Jones AC (1995) Historical document: life history and fisheries of Atlantic bluefin tuna: NOAA Tech. Memo. NMFS-SEFSC-370.
- 39. Duclerc J, Sacchi J, Piccinetti C, Piccinetti-Manfrin G, Dicenta A et al. (1973) New information on reproduction of bluefin tuna (Thunnus thynnus L.) and other species of Thunnidae in the Mediterranean. Rev Trav Institut Peches Maritimes Nantes 37: 163-176. [Google Scholar]
- 40. Rodriguez-Roda J, Dicenta A (1981) Áreas de puesta del Atún, Melva y Bonito en las costas de España y Marruecos. Col: ICCAT Vol Sci Pap 15: 278–283. [Google Scholar]
- 41. Alemany F, Quintanilla L, Velez-Belchi P, Garcia A, Cortés D et al. (2010) Characterization of the spawning habitat of Atlantic bluefin tuna and related species in the Balearic Sea (western Mediterranean). Prog Oceanogr 86: 21-38. doi:10.1016/j.pocean.2010.04.014. [Google Scholar]
- 42. Gordoa A, Olivar MP, Arévalo R, Viñas J, Molí B et al. (2009) Determination of Atlantic bluefin tuna (Thunnus thynnus) spawning time within a transport cage in the western Mediterranean. ICES J Mar Sci 66: 2205-2210. doi:10.1093/icesjms/fsp211. [Google Scholar]
- 43. Franqueville C (1971) Macroplancton profond (invertébrés) de la Méditerranée nord-occidentale. Tethys 3: 11-56. [Google Scholar]
- 44. Larson R, Mills C, Harbison G (1991) Western Atlantic midwater hydrozoan and scyphozoan medusae: in situ studies using manned submersibles. Hydrobiologia 216: 311-317. [Google Scholar]
- 45. Pawar S, Dell AI, Savage VM (2012) Dimensionality of consumer search space drives trophic interaction strengths. Nature 486: 485-489. PubMed: 22722834. [DOI] [PubMed] [Google Scholar]
- 46. Riisgard HU, Madsen CV (2011) Clearance rates of ephyrae and small medusae of the common jellyfish Aurelia aurita offered different types of prey. J Sea Res 65: 51-57. doi:10.1016/j.seares.2010.07.002. [Google Scholar]
- 47. Møller LF, Canon JM, Tiselius P (2010) Bioenergetics and growth in the ctenophore Pleurobrachia pileus . Hydrobiologia 645: 167-178. doi:10.1007/s10750-010-0219-6. [Google Scholar]
- 48. Quinn GGP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge University Press. [Google Scholar]
- 49. Bailey K, Batty R (1984) Laboratory study of predation by Aurelia aurita on larvae of cod, flounder, plaice and herring: development and vulnerability to capture. Mar Biol 83: 287-291. doi:10.1007/BF00397461. [Google Scholar]
- 50. Hansson LJ, Kiørboe T (2006) Effects of large gut volume in gelatinous zooplankton: Ingestion rate, bolus production and food patch utilization by the jellyfish Sarsia Tubulosa. J Plankton Res 28: 937-942. doi:10.1093/plankt/fbl030. [Google Scholar]
- 51. Gordoa A (2013) Analyses of connections between Atlantic Bluefin tuna fisheries at both sites of the Atlantic comprising Balfegó catch rates in Balearic spawning ground. Col: ICCAT Vol Sci Pap 69: In press [Google Scholar]
- 52. Gordoa A (2010) Temporal pattern of daily CPUE on the bluefin tuna (Thunnus thynnus) in the western Mediterranean spawning area. ICCAT Col Vol Sci Pap 65: 828-836. [Google Scholar]
- 53. Medina A, Abascal FJ, Aragón L, Mourente G, Aranda G et al. (2007) Influence of sampling gear in assessment of reproductive parameters for bluefin tuna in the western Mediterranean. Mar Ecol Prog Ser 337: 221-230. doi:10.3354/meps337221. [Google Scholar]
- 54. Fernández de Puelles ML, Alemany F, Jansa J (2007) Zooplankton time-series in the Balearic Sea (Western Mediterranean): variability during the decade 1994–2003. Prog Oceanogr 74: 329-354. doi:10.1016/j.pocean.2007.04.009. [Google Scholar]
- 55. Giorgi R, Avian M, De Olazábal S, Sandrini LR (1991) Feeding of Pelagia noctiluca in open sea. Jellyfish Blooms Mediterr Proc II Workshop Jellyfish Mediterr Sea: 102-111. [Google Scholar]
- 56. Sandrini LR, Avian M (1989) Feeding mechanism of Pelagia noctiluca (Scyphozoa: Semaeostomeae); laboratory and open sea observations. Mar Biol 102: 49-55. doi:10.1007/BF00391322. [Google Scholar]
- 57. Acuña JL, López-Urrutia Á, Colin S (2011) Faking giants: the evolution of high prey clearance rates in jellyfishes. Science 333: 1627-1629. doi:10.1126/science.1205134. PubMed: 21921197. [DOI] [PubMed] [Google Scholar]
- 58. Condon RH, Duarte CM, Pitt KA, Robinson KL, Lucas CH et al. (2013) Recurrent jellyfish blooms are a consequence of global oscillations. Proc Natl Acad Sci U S A 110: 1000-1005. doi:10.1073/pnas.1210920110. PubMed: 23277544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Mills CE (2001) Jellyfish blooms: are populations increasing globally in response to changing ocean conditions? In: Jellyfish Blooms: Ecological and Societal Importance. Springer Verlag; pp. 55-68. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A video showing the interaction between two ephyrae of Pelagianoctiluca at the time of catching Atlantic Bluefin tuna eggs used as prey under unstirred experimental conditions.
(WMV)
Video shots illustrating: 3D distribution of eggs under mixing condition, ephyra catching eggs with subumbellar tentacles under mixing conditions and ephyra under a stereo-microscope after feeding experiment.
(WMV)