

Abstract
Chronic infection of anogenital epithelium with human papillomavirus (HPV) promotes development of cancer. Many pathogens evoke immunosuppressive mechanisms to enable persistent infection. We have previously shown that grafted skin expressing HPV16 E7 oncoprotein from a keratin-14 promoter (K14E7) is not rejected by a syngeneic, immunocompetent host. In this study we show that indoleamine 2, 3-dioxygenase (IDO) 1, an IFN-γ inducible immunoregulatory molecule, is more highly expressed by langerin−ve dermal dendritic cells from K14E7 skin than nontransgenic control skin. Furthermore, inhibiting IDO activity using 1-D/L-methyl tryptophan promotes K14E7 skin graft rejection. Increased IDO1 expression and activity in K14E7 skin requires IFN-γ and iNKT cells, both of which have been shown to negatively regulate T-cell effector function and suppress K14E7 graft rejection. Further, dendritic cells from K14E7 skin express higher level of IFN-γ receptor (IFN-γR) than dendritic cells from control skin. K14E7 transgenic skin recruits significantly higher number of dendritic cells, independent of IFN-γ and IFN-γR expression. Consistent with these observations in a murine model, we found higher expression of IDO1 and IFN-γ but not IDO2 in the cervical epithelium of patients with HPV-associated cervical intraepithelial neoplasia (CIN) 2/3. Our data support a hypothesis that induction of IDO1 in HPV infected skin contributes to evasion of host immunity.
Keywords: human papillomavirus; indoleamine 2,3-dioxygenase; dendritic cells; interferon gamma; natural killer T cells; skin grafting
Introduction
Human papillomavirus (HPV) is an oncogenic virus that induces local epithelial hyperplasia. Persistent HPV infection can eventually lead to cervical cancer development (zur Hausen et al, 1981; Bhat et al, 2011). In most cases, tumor immunotherapy fails in late phase clinical trials despite potent tumor immunity induced by immunotherapy. The mechanisms likely to be responsible for this failure are impaired tumor cell antigen presentation (Matsumoto et al, 2004) and/or the generation of locally immunosuppressive mediators that can induce tumor cell resistance to T-cell effector mechanisms (Wang and Wang, 2007; Pardoll, 2012).
To understand immunosuppressive mechanisms associated with HPV infection, we developed a skin grafting model that mimics hyperplastic precancerous lesions. Unlike skin expressing other non-self antigens, skin expressing HPV16 E7 oncoprotein driven by the Keratin 14 promoter (K14E7) fails to reject when transplanted on a syngeneic immunocompetent host (Dunn et al, 1997; Matsumoto et al, 2004). Resistance of K14E7 grafts to immune-mediated rejection is mainly dependent on interferon (IFN)-γ production from invariant natural killer T (iNKT) cells (Mattarollo et al, 2010). IFN-γ is a cytokine that has both proinflammatory and anti-inflammatory functions. Though IFN-γ is often required by effector T cells for the killing of target cells, IFN-γ can also induce immune mediators which negatively regulate effector T cell function (Minguela et al, 2007).
IFN-γ is an inducer of indoleamine 2,3 dioxygenase 1 (IDO1), which catalyzes the first rate-limiting step of tryptophan metabolism and converts L-tryptophan into metabolites called “kynurenines” (Munn and Mellor, 2012). Tryptophan is an essential amino acid required for the function of effector T cells while kynurenines has multiple effects on T cells that includes inhibition of proliferation, induction of apoptosis and differentiation into a regulatory phenotype (Orabona and Grohmann, 2011). Other enzymes that are involved in tryptophan metabolism and have a possible role in immune resistance include tryptophan 2,3-dioxygenase (TDO) (Platten et al, 2012) and tryptophan hydroxylase-1 (TpH-1) (Nowak et al, 2012). However IDO1 is by far the most studied immunoregulatory enzyme. In addition to classical IDO (IDO1), IDO2 is a recently described isoform of IDO that explains some of the functions not previously explained by IDO1 (Metz et al, 2007). IDO activity controls allograft rejection, protects against autoimmune disease and contributes to inflammatory diseases such as rheumatoid arthritis and colitis (reviewed in (Orabona and Grohmann, 2011)). IDO1 and IDO2 are expressed by a range of human cancers (Lob et al, 2009; Godin-Ethier et al, 2011) and IDO inhibitor 1-D-MT is in clinical trial for the treatment of solid tumors that cannot be removed by surgery.
IDO1-deficient mice are resistant to carcinogen-induced skin tumors (Muller et al, 2008). In preclinical research, both shRNA (Huang et al, 2011) and siRNA-mediated (Yen et al, 2009) targeting of IDO1 and IDO2 has been successfully used for metastatic liver (Huang et al, 2011) and murine bladder tumor models (Yen et al, 2009). Recently it has been shown that the percentage of cells expressing IL-10, Fox-P3, IFN-γ and IDO increases from normal to cervical intraepithelial neoplasia (CIN) suggesting a role of IFN-γ induced IDO1 in a tumor immune escape mechanism (Kobayashi et al, 2008). IDO1 activity in murine dendritic cells (DCs) has been linked with immune tolerance and suppression of anti-tumor immunity (Sharma et al, 2007; Huang et al, 2010; Park et al, 2012).
As IDO is essential to the outcome of immune responses against tumors, we questioned if the outcome of K14E7 graft rejection is dependent on IDO activity. In this study we have identified IDO1 as an immunoregulatory molecule downstream of IFN-γ and iNKT cells that suppresses immune responses against HPV16-E7 expressing protein.
Results
IDO inhibition enables rejection of K14E7 skin grafts
To analyze the role of IDO in the rejection of K14E7 grafts, we administered 1-D/L-MT, an inhibitor of IDO activity, at 2mg/ml in the drinking water of the K14E7 and C57BL/6 (C57) donor mice and C57 recipient mice before and after the grafting (Figure 1a). Inhibition of IDO activity significantly increased rejection of K14E7 grafts but not C57 control grafts, suggesting that IDO contributes to suppression of host immune responses against the E7 oncoprotein. K14E7 grafts that were deficient in CD1d expression (NKT-deficient K14E7 grafts) were used as a positive control for graft rejection (Figure 1b). We confirmed that administration of 1-D/L-MT was associated with inhibition of IDO activity by measuring the serum ratios of kynurenine to tryptophan concentrations (Figure 1c).
Figure 1. Inhibition of IDO activity promotes K14E7 graft rejection.

(a) Schematic representation of the administration of 2mg/ml 1-D/L-MT in the drinking water of donor (C57Bl/6 and K14E7) and recipient C57Bl/6 (C57) mice before and after grafting. (b) Kaplan-Meier survival curves of C57, K14E7 and K14E7 grafts deficient in CD1d (CD1d−/−E7) on immunocompetent C57 recipient mice. C57 and K14E7 grafts were treated or not with 1-D/L-MT before and after the grafting. Median graft survival: K14E7 + 1-D/L-MT = 47 d; C57 + 1-D/L-MT = undefined (p = 0.02, log-rank test; n = 8 mice per group for K14E7 and C57 grafts; n=4 for CD1d−/−E7 grafts). (c) IDO activity (kyn/trp) from the serum of C57 mice transplanted with K14E7 grafts treated or not with 1-D/L-MT for 28 days post grafting.
iNKT cells and IFN-γ are required for the IDO1 expression in K14E7 skin
To determine the mechanism by which IDO activity contributes to the suppression of K14E7 graft rejection, we first analyzed the mRNA expression of IDO1 in transgenic K14E7 skin, and found it significantly higher than in nontransgenic C57 skin (Figure 2a). IDO2 is another isoform of IDO that could potentially contribute to an increased kyn/trp ratio and mediates immunosuppression in K14E7 skins (Qian et al, 2012). However we observed no difference in IDO2 mRNA expression in K14E7 and control skin (Figure 2b) suggesting that the increased IDO activity in K14E7 skin is contributed by IDO1. Consistent with these observations, we also found significantly higher IDO activity, as determined by kynurenine/tryptophan (kyn/trp) ratio, in K14E7 skin than in C57 control skin (Figure 2c).
Figure 2. IDO1 expression in K14E7 skin is dependent on IFN-γ and iNKT cells.

Expression of mRNA for (a) IDO1, (b) IDO2 and (d) TDO relative to RPL32 (L32) determined by real-time PCR in the ear skin of K14E7 and nontransgenic C57 mice. (c) IDO activity as measured by HPLC from the ear skin lysates of K14E7 and C57 mice. (n=10 for K14E7; n=7 for C57). Expression of mRNA for (e) IDO1 and (f) IDO2 relative to L32 by real-time PCR in the ear skin of C57, K14E7 and K14E7 mice deficient in Ja18 (Jα18−/−E7), CD1d (CD1d−/−E7), IFN-γ (IFN-γ−/−E7) or IFN-γ receptor (IFN-γR−/−E7). IDO activity as measured by HPLC from (g) ear skin lysates and (h) serum of C57, K14E7, Jα18−/−E7, CD1d−/−E7, IFN-γ−/−E7 and IFN-γR−/−E7. (ns= not significant; n=14 for K14E7 and 10 for C57; n≥4 for others mice).
Tryptophan 2,3 dioxygenase (TDO) is another immunoregulatory enzyme involved in tryptophan metabolism and may potentially contribute to the immunosuppressive environment in K14E7 skins (Platten et al, 2012). However, we did not observe any significant difference in the mRNA expression of TDO between K14E7 skin and control skin (Figure 2d).
We have recently demonstrated that IFN-γ producing iNKT cells contribute to suppressive immune effector mechanisms in E7 transgenic skin (Figure 1b) and protect K14E7 grafts from rejection (Mattarollo et al, 2010). Elevated IDO1 mRNA expression, and not IDO2, in K14E7 skin was not observed if the skin was deficient for IFN-γ expression, if the skin lacked iNKT cells or if the skin lacked the IFN-γ receptor (Figure 2e, 2f). Consistent with the mRNA data, IDO activity in the skin was also dependent on the presence of IFN-γ and of iNKT cells (Figure 2g). To assess whether increased IDO activity in K14E7 skin was a local or systemic phenomenon, we analyzed IDO activity in the serum of IFN-γ and iNKT deficient K14E7 mice. There was no significant difference in IDO activity in the serum of K14E7 and nontransgenic C57 mice suggesting that local IFN-γ in the skin of K14E7 mice contributes to increased IDO (Figure 2h).
DCs in K14E7 skin express elevated level of IFN-γ receptor
As higher levels of IDO1 expression and IDO activity in K14E7 transgenic skin required IFN-γ, we hypothesized that IDO1 production might be associated with a cell type expressing higher levels of IFN-γR. We did not observe any significant difference in the IFN-γR expression between total K14E7 and C57 skins, and between normal and CIN lesions (data not shown).
However, we observed higher expression of IFN-γR on DCs (CD45+MHCII+CD11c+) from K14E7 skin than DCs from C57 control skin, whereas expression of IFN-γR in non-DCs (CD45+MHCII−CD11c−) and CD45 negative non-hematopoietic population was not significantly different between K14E7 and control skin (Figure 3a, 3b). Interestingly, the higher expression of IFN-γR on K14E7 skin DCs was independent of production of IFN-γ in the skin (Figure 3a, 3b). Together these results suggest that IFN-γR+ DCs in K14E7 skin may be the source of IDO1 and contribute to the suppression of K14E7 graft rejection.
Figure 3. Skin dendritic cells in K14E7 mice express high level of IFN-γR.

(a) Histogram plots showing flow cytometry staining for surface IFN-γ receptor (unfilled black histograms) and isotype controls (filled grey histograms) in the ear skin of C57 (top), K14E7 (middle) and IFN-γ−/−E7 (bottom) mice. (b) Bar graph shows surface levels of IFN-γR expression on DCs from ear skin of C57 (white), K14E7 (black) and IFN-γ−/−E7 (grey) mice shown as δMFI (mean fluorescence intensity) (δMFI = MFI of IFN-γR mAb − MFI of isotype control mAb). IFNγR−/−E7 mice (line bar) were used as a negative control for IFNγR staining. (mean + SEM; n = 4 mice per group for K14E7 and IFN-γ−/−E7; n= 6 for C57 mice, data pooled from 2 independent experiments).
CD11c+ DC in K14E7 skins express IDO
We next tested if DCs in K14E7 skin are the main cells expressing IDO1. Flow cytometry staining on CD45+CD11c+ DCs in K14E7 skin demonstrated IDO1 expression in CD11c+ DCs but not on the CD3+ lymphocyte population (Figure 4a, 4b). In contrast, DCs from nontransgenic C57 skin expressed little or no IDO1. In addition, we observed reduced IDO1 expression in the skin DCs from IFN-γR deficient K14E7 mice (Figure 4a, 4b) confirming that IFN-γR signaling induces increased expression of IDO1 in K14E7 skin DCs. We further show with immunofluorescence staining that IDO1 is expressed by DCs in the dermis of K14E7 mice and injection of IFN-γ induces IDO1 expression in both CD11+c and CD11c− cells in IFN-γ deficient K14E7 mice (Supplementary figure 1a and 1b)
Figure 4. Dendritic cell infiltrates in K14E7 skin express IDO1 that is dependent on IFN-γR on DCs.

(a) Flow cytometry plot showing CD3 and CD11c staining (left) on the skin cells pregated on CD45+ live cells. Histogram plot showing intracellular staining for IDO1 (filled black histogram) and isotype controls (filled grey histogram) in CD3+ lymphocytes and CD11c+ cells from K14E7, C57 and IFN-γR−/−E7 mice. (b) Bar graph shows representative FACS data as the percentage of IDO1 positive DCs in the skin of C57, K14E7 and IFN-γR−/−E7 mice. (mean + SEM; n = 4 mice per group for K14E7 and IFN-γ−/−E7; n= 6 for C57 mice). (c) FACS plot showing gating strategy for sorting resident (CD11c+MHCint) and migratory (CD11c+MHChi) DCs. (d) mRNA expression of IDO1 relative to RPLL32 by real-time PCR in the resident and migratory DCs sorted from the skin draining lymph nodes of K14E7 mice. ND= not detectable. (e) Bar graph shows absolute number of MHCII+CD11c+ DCs (representative of FACS data) in the ear skin of C57, K14E7, IFN-γ−/−E7 and IFN-γR−/−E7 mice.
To further confirm that IDO1 expression by skin DCs is due to local signaling, we isolated migratory (CD11c+MHCIIhigh) and resident DCs (CD11c+MHCIIintermediate) from the skin draining lymph nodes of K14E7 mice (Figure 4c). Resident DCs did not express any detectable IDO1 in contrast to high levels of expression in migratory DCs, suggesting that local IFN-γ in the K14E7 skin induces IDO1 expression (Figure 4d). In addition, we observed a higher proportion of MHCII+CD11c+ DCs in K14E7 skin than in C57 skin, and the increased number of DCs in the K14E7 skin was independent of IFN-γ or IFN-γR expression (Figure 4e). This result suggests that the previously demonstrated rejection of IFN-γ−/−E7 (Mattarollo et al, 2010) and IFN-γR−/−E7 skin grafts (Kluyver R, Ph.D. thesis 2008) is not attributable to the difference in DCs recruitment to the skin.
Langerin−ve dermal DCs express IDO1 and contributes to increased DC infiltrates in K14E7 skin
In skin, several DC subsets have been identified (Henri et al, 2010). In K14E7 skin, we found that langerin−ve dermal DCs are the main contributors to increased DC numbers (Figure 5a–d). Further, IDO1 expression in the skin of K14E7 mice was restricted to langerin−ve DCs (Figure 5e, 5f). In addition to strengthen the relation of IFN-γ induced IDO1 in immune suppression, we show that splenic DCs from K14E7 mice, when prestimulated with IFN-γ, could induce IDO1 expression (Supplementary figure 2) and also suppress CD8 cell proliferation (Figure 5g).
Figure 5. IDO1 is expressed by langerin−ve dendritic cells.

(a) Dot plot shows classical dermal DCs (CD11b+CD207−), langherans cells (CD207+CD103−) and CD207+CD103+ DCs in the skin of K14E7 mice gated on live CD45+CD11c+MHCII+ cells. Absolute number of (b) classical dermal DC (c) langherans cells and (d) CD103+ dermal DCs subsets in the ear skin of C57 and K14E7 mice (mean + SEM; n = 4 mice per group for K14E7 and n= 6 for C57 mice). (e) Isotype control and (f) real staining for IDO1 and CD207, pregated on CD11c+MHCII+ cells, in the ear skin of K14E7 mice. (g) Proliferation of splenic CD8 cells cultured with splenic DCs (DC:T:irradiated feeder cells = 1:5:5) from K14E7 and IFN-γR−/−E7 mice with or without 1-MT (1mM) and IFN-γ (10ng/ml). Results are expressed as mean cpm ± SEM of triplicate wells (cpm= counts per minute).
IDO expression in CIN patients
Having established that local IDO activity in K14E7 transgenic mouse skin is necessary to protect K14E7 skin from immune mediated rejection, we examined the expression of IDO1, IDO2 and other related genes in HPV-associated CIN-2/3 patients and surrounding cervical epithelial tissues. Consistent with previous studies (Kobayashi et al, 2008), we found higher mRNA expression of IDO1 and IFN-γ, but not of IDO2 in CIN-2/3 cervical biopsy samples (Figure 6).
Figure 6. Elevated IDO1 and IFN-γ mRNA expression in CIN lesions.
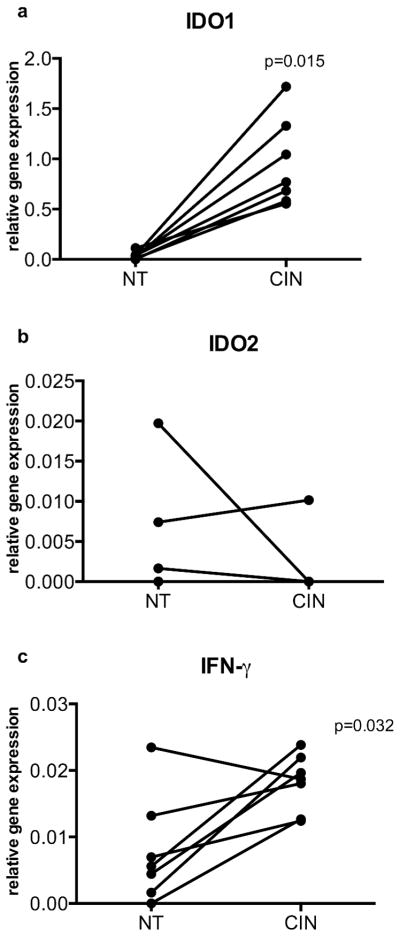
(a) mRNA expression of human IDO1, (b) IDO2 and (c) IFN-γ relative to HPRT by real-time PCR in biopsies of normal (NT) cervical epithelium and CIN 2/3 (CIN) tissue. (mean + SEM; n=7). IDO2 expression was not detectable in most of the samples analyzed.
Discussion
We have previously shown that skin expressing HPV16 E7 oncoprotein is hyperplastic and has increased immune cell infiltrate, relative to skin from syngeneic nontransgenic mice. Immunization along with adoptive transfer of high numbers of E7-specific CD8 T cells is required to successfully reject K14E7 skin grafts (Matsumoto et al, 2004). This generates circulating memory T cells. However they fail to reject second newly placed K14E7 grafts and a further immunization is required (Matsumoto et al, 2004), suggesting that the expression of E7 oncoprotein generates a potent local immunosuppressive environment in the host. Understanding the mechanisms that leads to successful rejection of K14E7 grafts could pave the way for future immunotherapies against HPV-associated cancers.
We have recently shown that iNKT cells are specifically attracted to hyperplastic K14E7 skin and IFN-γ production by iNKT cells is critical for the protection of K14E7 grafts (Mattarollo et al, 2010). We hypothesized that IFN-γ induces immunoregulatory molecules such as IDO1 that is required for the survival of K14E7 grafts on immunocompetent host. In this study we demonstrate that inhibition of IDO activity by 1-D/L-MT successfully rejects K14E7 grafts. We used a mixture of D- and L- stereoisomer of 1-MT that is known to inhibit both IDO1 and IDO2. However we have shown that it is mainly IDO1 and not IDO2 that is overexpressed in K14E7 skins and CIN lesions. This is consistent with previous observations where increased IDO1 has been associated with skin lesions of inflammatory skin disease (Ito et al, 2004). Moreover, IDO1 expression and IDO activity is dependent on IFN-γ and iNKT cells as K14E7 skin deficient in CD1d, Jα18, IFN-γ or IFN-γR had IDO1 expression and IDO activity similar to nontransgenic controls (Figure 3a, 3c). We also observed a transient increase in IDO1 expression day 3 post grafting that was dependent on IFN-γ expression in the K14E7 grafted skin (data not shown). Our data agree with recent studies showing that iNKT ligand a-galactosylceramide (a-galcer) induces IDO1 possibly by releasing IFN-γ (Fallarini et al, 2008; Guillonneau et al, 2009).
TDO is another enzyme of tryptophan metabolism that can potentially contribute to observed local kynurenine concentration in K14E7 skins (Platten et al, 2012). However we did not detect any significant difference in TDO expression between K14E7 and nontransgenic control skins demonstrating that increased IDO1 in K14E7 skins contributes to increased kynurenine production that is likely to be related to observed immunosuppression. We could obtain K14E7 graft rejection in only half of the transplanted grafts. This partial but significant rejection can be explained by incomplete inhibition of IDO by 1-D/L-MT. Indeed we found that graft rejection was correlated to the inhibition of IDO activity. Alternatively, other redundant immunoregulatory compensatory mechanisms including IFN-γ inducible molecules such as programmed cell death 1 ligand 1(PD-L1)/2 or T cell immunoglobulin and mucin-domain (Tim)-3 that may act alone or in association with IDO1 as has been proposed recently (Sharma et al, 2007; Baban et al, 2011), may be induced upon inhibition of IDO activity.
DCs are specialized cell types that acquire process and present antigens to T cells leading to their activation, clonal expansion and differentiation into effector T cells. However some DCs suppress T cell responses by generating T cells with regulatory functions, including CD4+CD25+Foxp3+ regulatory T (Tregs) cells (Huang et al, 2010). IDO1 expression by DCs and plasmacytoid DCs has been shown to induce immune tolerance that is related to the pathogenesis of chronic hepatitis C infection (Higashitani et al, 2012), suppression of antitumor immune responses (Munn et al, 2004; Sharma et al, 2007) and collagen-induced arthritis (Park et al, 2012). DC-SIGN expressing CD1a+ immature DCs in the stroma of high grade CIN lesions express immunosuppressive factors such as IDO1, tumor growth factor β and IL-10 (Kobayashi et al, 2008). However in K14E7 skin, IDO1 expression was restricted to langerin−ve DCs (Figure 5e, 5f). We predict that, like iNKT cells, DCs are specifically recruited to K14E7 hyperplastic skin. DCs recruitment to the hyperplastic skin is independent of IFN-γ or IFN-γR expression (Figure 4e). It is likely that after reaching the skin, these cells produce high levels of IDO1 due to the presence of high quantity of local IFN-γ, as K14E7 skin DCs deficient for IFN-γR did not have IDO1 expression (Figure 4a, 4b). In support of this hypothesis, we observed that unlike migratory DCs, resident DCs did not express IDO1 in the skin draining lymph nodes of K14E7 mice. This is consistent with the recent literature that high level of IFN-γ is anti-inflammatory (Minguela et al, 2007).
The mechanism that IDO1 uses to induce immunosuppression includes generation of regulatory T cells. IDO1 expressing DCs activate FoxP3+ Tregs that are a major regulatory cell population for suppressing effector responses (Huang et al, 2010; Higashitani et al, 2012). However Tregs are not involved in IDO1 mediated immune resistance in K14E7 skin, as we have shown previously that, though high in number in K14E7 skins, depletion of FoxP3+ Tregs did not elicit graft rejection (Mattarollo et al, 2011). IDO enzymatic activity can directly suppress effector responses by depletion of tryptophan, by production of tryptophan metabolites (kynurenines) and by changing redox potentials due to consumption of superoxide radicals (reviewed in (Huang et al, 2010)). T cells are highly sensitive to changes in local tryptophan concentrations, and kynurenines can induce proliferative arrest and apoptosis in T cells that is dependent on GCN-2 kinase (Fallarino et al, 2006). Therefore we hypothesize that in the model of K14E7 skin grafting increased kynurenine and decreased tryptophan concentrations inhibit effector T cells functions and suppress graft rejection.
Overall, our data strongly suggests a critical immune regulatory role of IDO1 in K14E7 hyperplastic skin. In this model, the potent immunosuppressive IDO1 expression in DCs is induced by local IFN-γ produced from iNKT cells in the skin. Consistent with previous studies (Kobayashi et al, 2008), we also observed elevated mRNA expression of IFN-γ and IDO1 in CIN-2/3 samples than normal tissue. Collectively, our observations support the use of 1-MT for immunotherapy against HPV-associated epithelial cancers.
Methods
Mice
C57BL/6 mice and HPV16-E7 transgenic mice, in which E7 oncoprotein is expressed in basal keratinocytes under the control of K14 promoter (designated K14E7), were obtained from the Animal Resources Center (Perth, Australia). IFN-γ−/− and IFN-γR−/− mice were purchased from The Jackson Laboratory (Bar Harbor, ME) and NKT cell-deficient CD1d−/− and Jα18−/− mice were obtained from M. Smyth (Melbourne, Australia). To generate K14E7 transgenic crosses with CD1d−/−, Jα18−/−, IFN-γ−/− and IFN-γR−/− knockout (KO) mice, hemizygous K14E7 mice were crossed and then backcrossed with homozygous CD1d−/−, Jα18−/−, IFN-γ−/− or IFN-γR−/− mice to an F2 generation. All mice were on C57Bl/6 background and K14E7 negative littermate controls were used for several experiments. All mice were housed under specific pathogen-free conditions at the Princess Alexandra Hospital Biological Research Facility (Brisbane, Australia), sex-matched and used at 6–10 weeks of age for all experiments. All animal procedures were approved by the University of Queensland Animal Ethics Committee.
Skin transplantation
Skin from donor mice was transplanted onto the flank of recipient mice as previously described (Dunn et al, 1997; Matsumoto et al, 2004). Briefly, donor skin was taken from the dorsal and ventral surfaces of the ear (~1cm2) and placed onto the thoracic flank region of anaesthetized recipients. Grafts were held in place with antibiotic-permeated gauze (Bactigras; Smith and Nephew, London, UK) and bandaged with micropore tape and Flex-wrap (Lyppard, Queensland, Australia). Bandages were removed 7 days after grafting, and grafts were monitored daily for loss of distinct border and signs of ulceration or necrosis to >80% of the graft, which was used to define graft rejection.
Inhibition of IDO with 1-methyl-dl-tryptophan
1-methyl-DL-tryptophan (1-D/L-MT) was obtained from Sigma-Aldrich and prepared as a 20-mmol/l stock in 0.1 N NaOH, adjusted to pH 7.4 and stored at 4 ° C protected from light. Mice were given 2mg/ml of 1-D/L-MT in the drinking water, supplemented with 2g/L of aspartame (Equal), before and after grafting as shown in Figure 1a. Mice drank an average of 5ml/day and water was replaced every 5 days during the grafting. Mice administrated with 1-D/L-MT in the drinking water did not show change in weight and the amount of water intake was similar to control groups.
Immunofluorescence
Excised ear skin samples were fixed in PLP buffer, dehydrated in 20% sucrose and stained with fluorochrome conjugated antibodies as described previously (Mittal et al, 2010).
Quantitative real time PCR
Mouse ear samples were homogenized in trizol and RNA was extracted using a RNeasy kit (Qiagen). All human subjects provided written informed consent and the experiments were approved by the institution ethics committee (#2009001060) and conducted in compliance with the Helsinki guidelines.
High performance liquid chromatography
Serum and ear skin tissues from mice were collected and analyzed for kynurenine and tryptophan concentrations (TM) on HPLC (Agilent Technologies Series 1200) as described previously (Laich et al, 2002). In brief, tissues were homogenized in PBS buffer containing protease inhibitors (Roche Applied Sciences). Tissue supernatants and serum were deproteinated in TCA and 50Tl was used for HPLC (C18 reverse-phase column). Peak area values for kynurenine and tryptophan were determined and the results for IDO activity were plotted as a Kynurenine/tryptophan (kyn/trp) ratio.
Flow cytometry
Flow cytometry staining was performed as previously described (Mattarollo et al, 2010).
T cell stimulation assay
DCs were purified by CD11c+ positive selection and splenic CD8+ cells were stained with anti-CD8 PE. Anti-PE beads (Miltenyi Biotech) were used to purify the cells [magnetic cell sorter (MACS), Miltenyi Biotech]. Purified DCs from spleen of K14E7 mice were incubated with splenic CD8 (2×105 cells/well) from K14E7 mice (DC: T cell = 1:10) in the presence irradiated splenocytes in RPMI complete medium for 4 days in a 96-well U-bottom plate. For some experiments, purified splenic DCs from K14E7 and IFN-γR- deficient K14E7 mice were preincubated with recombinant mouse IFN-γ (R&D systems, USA) for 24 h before culturing with T cells. The cultures were then pulsed with 1 μTCi [3H] thymidine for 18 h and the radioactivity incorporated was determined with a Beta Scintillation Counter (Microbeta Trilux, Perkin Elmer, Wellesley, MA).
Statistical analysis
Kaplan–Meier plots were used to analyze skin graft survival, and a log-rank test was performed to assess the statistical significance of differences between survival curves. For human CIN samples, a two-tailed paired nonparametric Wilcoxon test was performed. For all other data in which statistics were performed, a two-tailed, nonparametric Mann–Whitney test was used for assessment of differences between groups. All experiments were repeated two to three times with similar results. Differences with a P-value <0.05 were considered to be significant. Prism (Graph pad Software, La Jolla, CA) was used for graphs and statistical analysis.
Supplementary Material
Acknowledgments
This work was supported by grants from the NIH (5U01CA141583), National Health and Medical Research Council of Australia (569938), Australian Research Council and the Cancer Council Queensland also ACRF. We thank Dr. Lewis Perrin from Mater Medical Centre, Brisbane for providing human CIN and normal tissues, Prof Ranjeny Thomas’ group for technical assistance with HPLC, and staff of the Biological Research Facility at Princess Alexandra Hospital for excellent technical assistance with grafting and animal care.
Abbreviations
- CIN
cervical intraepithelial neoplasia
- DCs
dendritic cells
- HPV
human papillomavirus
- IDO
Indoleamine 2, 3-dioxygenase
- IFN-γ
interferon gamma
- 1-D/L-MT
1-methyl-d/l-tryptophan
- iNKT
invariant natural killer T cells
Footnotes
Conflict of interest
The authors state no conflict of interest.
References
- Baban B, Chandler PR, Johnson BA, 3rd, et al. Physiologic control of IDO competence in splenic dendritic cells. J Immunol. 2011;187(5):2329–2335. doi: 10.4049/jimmunol.1100276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhat P, Mattarollo SR, Gosmann C, et al. Regulation of immune responses to HPV infection and during HPV-directed immunotherapy. Immunol Rev. 2011;239(1):85–98. doi: 10.1111/j.1600-065X.2010.00966.x. [DOI] [PubMed] [Google Scholar]
- Dunn LA, Evander M, Tindle RW, et al. Presentation of the HPV16E7 protein by skin grafts is insufficient to allow graft rejection in an E7-primed animal. Virology. 1997;235(1):94–103. doi: 10.1006/viro.1997.8650. [DOI] [PubMed] [Google Scholar]
- Fallarini S, Paoletti T, Panza L, et al. Alpha-galactosylceramide modulates the induction of indoleamine 2,3-dioxygenase in antigen presenting cells. Biochem Pharmacol. 2008;76(6):738–750. doi: 10.1016/j.bcp.2008.07.001. [DOI] [PubMed] [Google Scholar]
- Fallarino F, Grohmann U, You S, et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory phenotype in naive T cells. J Immunol. 2006;176(11):6752–6761. doi: 10.4049/jimmunol.176.11.6752. [DOI] [PubMed] [Google Scholar]
- Godin-Ethier J, Hanafi LA, Piccirillo CA, et al. Indoleamine 2,3-dioxygenase expression in human cancers: clinical and immunologic perspectives. Clin Cancer Res. 2011;17(22):6985–6991. doi: 10.1158/1078-0432.CCR-11-1331. [DOI] [PubMed] [Google Scholar]
- Guillonneau C, Mintern JD, Hubert FX, et al. Combined NKT cell activation and influenza virus vaccination boosts memory CTL generation and protective immunity. Proc Natl Acad Sci U S A. 2009;106(9):3330–3335. doi: 10.1073/pnas.0813309106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henri S, Guilliams M, Poulin LF, et al. Disentangling the complexity of the skin dendritic cell network. Immunol Cell Biol. 2010;88(4):366–375. doi: 10.1038/icb.2010.34. [DOI] [PubMed] [Google Scholar]
- Higashitani K, Kanto T, Kuroda S, et al. Association of enhanced activity of indoleamine 2,3-dioxygenase in dendritic cells with the induction of regulatory T cells in chronic hepatitis C infection. J Gastroenterol. 2012 doi: 10.1007/s00535-012-0667-z. [DOI] [PubMed] [Google Scholar]
- Huang L, Baban B, Johnson BA, 3rd, et al. Dendritic cells, indoleamine 2,3 dioxygenase and acquired immune privilege. Int Rev Immunol. 2010;29(2):133–155. doi: 10.3109/08830180903349669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang TT, Yen MC, Lin CC, et al. Skin delivery of short hairpin RNA of indoleamine 2,3 dioxygenase induces antitumor immunity against orthotopic and metastatic liver cancer. Cancer Sci. 2011;102(12):2214–2220. doi: 10.1111/j.1349-7006.2011.02094.x. [DOI] [PubMed] [Google Scholar]
- Ito M, Ogawa K, Takeuchi K, et al. Gene expression of enzymes for tryptophan degradation pathway is upregulated in the skin lesions of patients with atopic dermatitis or psoriasis. J Dermatol Sci. 2004;36(3):157–164. doi: 10.1016/j.jdermsci.2004.08.012. [DOI] [PubMed] [Google Scholar]
- Kobayashi A, Weinberg V, Darragh T, et al. Evolving immunosuppressive microenvironment during human cervical carcinogenesis. Mucosal Immunol. 2008;1(5):412–420. doi: 10.1038/mi.2008.33. [DOI] [PubMed] [Google Scholar]
- Laich A, Neurauter G, Widner B, et al. More rapid method for simultaneous measurement of tryptophan and kynurenine by HPLC. Clin Chem. 2002;48(3):579–581. [PubMed] [Google Scholar]
- Lob S, Konigsrainer A, Zieker D, et al. IDO1 and IDO2 are expressed in human tumors: levo- but not dextro-1-methyl tryptophan inhibits tryptophan catabolism. Cancer Immunol Immunother. 2009;58(1):153–157. doi: 10.1007/s00262-008-0513-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto K, Leggatt GR, Zhong J, et al. Impaired antigen presentation and effectiveness of combined active/passive immunotherapy for epithelial tumors. J Natl Cancer Inst. 2004;96(21):1611–1619. doi: 10.1093/jnci/djh301. [DOI] [PubMed] [Google Scholar]
- Mattarollo SR, Rahimpour A, Choyce A, et al. Invariant NKT cells in hyperplastic skin induce a local immune suppressive environment by IFN-gamma production. J Immunol. 2010;184(3):1242–1250. doi: 10.4049/jimmunol.0902191. [DOI] [PubMed] [Google Scholar]
- Mattarollo SR, Yong M, Gosmann C, et al. NKT cells inhibit antigen-specific effector CD8 T cell induction to skin viral proteins. J Immunol. 2011;187(4):1601–1608. doi: 10.4049/jimmunol.1100756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metz R, Duhadaway JB, Kamasani U, et al. Novel tryptophan catabolic enzyme IDO2 is the preferred biochemical target of the antitumor indoleamine 2,3-dioxygenase inhibitory compound D-1-methyl-tryptophan. Cancer Res. 2007;67(15):7082–7087. doi: 10.1158/0008-5472.CAN-07-1872. [DOI] [PubMed] [Google Scholar]
- Minguela A, Pastor S, Mi W, et al. Feedback regulation of murine autoimmunity via dominant anti-inflammatory effects of interferon gamma. J Immunol. 2007;178(1):134–144. doi: 10.4049/jimmunol.178.1.134. [DOI] [PubMed] [Google Scholar]
- Mittal D, Saccheri F, Venereau E, et al. TLR4-mediated skin carcinogenesis is dependent on immune and radioresistant cells. EMBO J. 2010;29(13):2242–2252. doi: 10.1038/emboj.2010.94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muller AJ, Sharma MD, Chandler PR, et al. Chronic inflammation that facilitates tumor progression creates local immune suppression by inducing indoleamine 2,3 dioxygenase. Proc Natl Acad Sci U S A. 2008;105(44):17073–17078. doi: 10.1073/pnas.0806173105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munn DH, Mellor AL. Indoleamine 2,3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2012 doi: 10.1016/j.it.2012.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munn DH, Sharma MD, Hou D, et al. Expression of indoleamine 2,3-dioxygenase by plasmacytoid dendritic cells in tumor-draining lymph nodes. J Clin Invest. 2004;114(2):280–290. doi: 10.1172/JCI21583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowak EC, de Vries VC, Wasiuk A, et al. Tryptophan hydroxylase-1 regulates immune tolerance and inflammation. J Exp Med. 2012;209(11):2127–2135. doi: 10.1084/jem.20120408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orabona C, Grohmann U. Indoleamine 2,3-dioxygenase and regulatory function: tryptophan starvation and beyond. Methods Mol Biol. 2011;677:269–280. doi: 10.1007/978-1-60761-869-0_19. [DOI] [PubMed] [Google Scholar]
- Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12(4):252–264. doi: 10.1038/nrc3239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park MJ, Park KS, Park HS, et al. A distinct tolerogenic subset of splenic IDO(+)CD11b(+) dendritic cells from orally tolerized mice is responsible for induction of systemic immune tolerance and suppression of collagen-induced arthritis. Cell Immunol. 2012;278(1–2):45–54. doi: 10.1016/j.cellimm.2012.06.009. [DOI] [PubMed] [Google Scholar]
- Platten M, Wick W, Van den Eynde BJ. Tryptophan Catabolism in Cancer: Beyond IDO and Tryptophan Depletion. Cancer Res. 2012;72(21):5435–5440. doi: 10.1158/0008-5472.CAN-12-0569. [DOI] [PubMed] [Google Scholar]
- Qian F, Liao J, Villella J, et al. Effects of 1-methyltryptophan stereoisomers on IDO2 enzyme activity and IDO2-mediated arrest of human T cell proliferation. Cancer Immunol Immunother. 2012;61(11):2013–2020. doi: 10.1007/s00262-012-1265-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma MD, Baban B, Chandler P, et al. Plasmacytoid dendritic cells from mouse tumor-draining lymph nodes directly activate mature Tregs via indoleamine 2,3- dioxygenase. J Clin Invest. 2007;117(9):2570–2582. doi: 10.1172/JCI31911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang HY, Wang RF. Regulatory T cells and cancer. Curr Opin Immunol. 2007;19(2):217–223. doi: 10.1016/j.coi.2007.02.004. [DOI] [PubMed] [Google Scholar]
- Yen MC, Lin CC, Chen YL, et al. A novel cancer therapy by skin delivery of indoleamine 2,3-dioxygenase siRNA. Clin Cancer Res. 2009;15(2):641–649. doi: 10.1158/1078-0432.CCR-08-1988. [DOI] [PubMed] [Google Scholar]
- zur Hausen H, de Villiers EM, Gissmann L. Papillomavirus infections and human genital cancer. Gynecol Oncol. 1981;12(2 Pt 2):S124–128. doi: 10.1016/0090-8258(81)90067-6. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.