

Abstract
The study of microbial communities often leads to arguments for the evolution of cooperation due to group benefits. However, multilevel selection models caution against the uncritical assumption that group benefits will lead to the evolution of cooperation. We analyze a microbial social trait to precisely define the conditions favoring cooperation. We combine the multilevel partition of the Price equation with a laboratory model system: swarming in Pseudomonas aeruginosa. We parameterize a population dynamics model using competition experiments where we manipulate expression, and therefore the cost-to-benefit ratio of swarming cooperation. Our analysis shows that multilevel selection can favor costly swarming cooperation because it causes population expansion. However, due to high costs and diminishing returns constitutive cooperation can only be favored by natural selection when relatedness is high. Regulated expression of cooperative genes is a more robust strategy because it provides the benefits of swarming expansion without the high cost or the diminishing returns. Our analysis supports the key prediction that strong group selection does not necessarily mean that microbial cooperation will always emerge.
Keywords: conflict, cooperation, metabolic prudence, Pseudomonas aeruginosa , swarming
Introduction
Over the past decade microbiology has shifted perspective to acknowledge that bacteria are not solitary organisms but rather social organisms that rely on a range of population-level traits, such as biofilms, cell–cell communication and cooperative drug resistance (Kolter and Greenberg, 2006; Greenberg, 2010; Carmona-Fontaine and Xavier, 2012). However, often the existence of microbial social traits is justified by their group-level benefits (Costerton et al, 1999; Monds and O’Toole, 2009; Lee et al, 2010). Social evolution theory predicts that defector phenotypes (i.e., non-cooperative phenotypes, see Table I for our definitions) are favored in mixed populations by individual-level selection (Brannstrom and Dieckmann, 2005; West et al, 2006; Nadell et al, 2009). In fact, experiments with microbes show that a costly cooperative trait may be favored for its group- or species-level benefits but disfavored in populations where different strains and species mix (Strassmann et al, 2000; Griffin et al, 2004; Diggle et al, 2007; Sathe et al, 2010). Understanding how cooperation evolves and remains stable is a key to understanding social traits in bacteria and other microbes (e.g., Crespi, 2001; Fortunato et al, 2003; Rainey and Rainey, 2003; Greig and Travisano, 2004; Griffin et al, 2004; Fiegna et al, 2006; West et al, 2006; Diggle et al, 2007; Ross-Gillespie et al, 2007; Nadell et al, 2009; Ross-Gillespie et al, 2009; Kummerli et al, 2009a, b, c, 47, 48; Sathe et al, 2010; Smith et al, 2010; Foster, 2011; Koschwanez et al, 2011, 2013; Strassmann and Queller, 2011; Xavier, 2011a; Damore and Gore, 2012; Ratcliff et al, 2012; Celiker and Gore, 2012a).
Table 1. Definition of terms used in this paper.
| Cooperative trait | A trait that confers benefits to a recipient individual. The cooperative trait can be costly to the actor (altruistic trait) or beneficial to the actor (mutualism) (West et al, 2006) |
| Cooperator | An individual or a strain carrying the cooperative trait, also called the ‘actor’ |
| Defector | An individual or a strain lacking a cooperative trait but capable of exploiting the cooperation of others |
| Metabolic prudence | A mechanism of gene expression regulation that allows wild-type Pseudomonas aeruginosa to produce biosurfactants at no cost to its fitness (Xavier et al, 2011) |
| Selfishness | A defector strategy where individuals lack a costly cooperative trait but exploit the cooperation of others |
Here, we use a combination of quantitative experiments and mathematical modeling to analyze a model social trait, swarming in Pseudomonas aeruginosa, and to determine conditions favoring cooperation. Swarming is a collective form of migration that allows colonies to expand over soft surfaces and thus provides a group benefit. But swarming also requires that individual bacteria secrete massive amounts of rhamnolipid biosurfactants to lubricate the surface (Deziel et al, 2003; Caiazza et al, 2005). The secreted surfactants are a public good and can be exploited by surfactant-deficient defectors, which benefit from the surfactants secreted by others within the colony without producing surfactants themselves (Xavier et al, 2011). In general, if the production of a public good is costly, then defectors can outcompete cooperators within a population and, in the absence of stabilizing processes such as kin selection or discrimination (West et al, 2006), eventually drive the cooperative trait to extinction. Nonetheless, many natural isolates of P. aeruginosa do secrete rhamnolipids (Deziel et al, 1996), which suggests that there are mechanisms favoring and stabilizing rhamnolipid secretion in the wild. A mechanism found recently (Xavier et al, 2011) explains that P. aeruginosa regulates the expression of the rhamnolipid synthesis operon rhlAB using a combination of quorum sensing (the las/rhl quorum sensing cascade) and nutrient sensing (Figure 1A). Although not all the molecular players in this integration of quorum and nutrient sensing are known (Boyle et al, 2013), it is clear that such a combined regulatory mechanism enables bacteria to delay rhamnolipid production to times when there is excess carbon, and rhamnolipid synthesis becomes affordable (Figure 1B). This mechanism, called metabolic prudence, implements a molecular decision-making circuitry that effectively decreases the fitness cost of rhamnolipid secretion and prevents exploitation by rhamnolipid-deficient defectors (Xavier et al, 2011).
Figure 1.

Pseudomonas aeruginosa synthesizes and secretes rhamnolipid biosurfactants required for swarming motility. (A) The wild-type Pseudomonas aeruginosa regulates the expression of rate-limiting enzyme RhlA using a mechanism called metabolic prudence that integrates nutrient sensing and quorum sensing. (B) Metabolic prudence ensures that rhamnolipids are secreted only when carbon is in excess and population density is high. (C) In this study, we use a genetically engineered inducible strain where RhlA expression is under regulation of the L-arabinose inducible promoter PBAD. (D) The inducible construct circumvents metabolic prudence and provides an experimental handle to modulate biosurfactant synthesis and investigate swarming cooperation thoroughly.
Hamilton’s rule (Hamilton, 1964) explains that cooperation can evolve when br>c where c is the fitness cost to the actor, b is the fitness benefit to the recipient and r is the correlation between the genotypes of actors and recipients, also called relatedness. Metabolic prudence reduces the costs of cooperation by regulating the expression of the cooperative genes (Xavier et al, 2011), but cooperation could also be favored by increasing the benefits or the relatedness (West et al, 2006; Chuang et al, 2010). Furthermore, this mechanism brings up an important question. While some genes are constitutively expressed, many others are conditionally regulated to account for a fluctuating demand for the gene product (Perkins and Swain, 2009); is it better to actively regulate a gene versus constitutively expressing it when the gene regulates a cooperative trait? This question has previously been addressed in other systems (Nadell et al, 2008; Kummerli and Brown, 2010; Geisel, 2011) and, for a fixed but arbitrary level, in P. aeruginosa swarming (Xavier et al, 2011). It remains to be tested whether P. aeruginosa metabolic prudence is still a better strategy when constitutive expression of biosurfactants occurs at an optimum rate, and when selection acts at multiple levels.
Here, we investigate multilevel selection in swarming when the expression of cooperative biosurfactant synthesis is kept constant but at a rate that provides optimal group benefits. We use an engineered strain of P. aeruginosa that allows us to control the expression of biosurfactant synthesis genes, and thus the investment into cooperation by individuals (Figure 1C and D). We find that costly swarming can indeed be favored thanks to the large group benefits of population expansion. Furthermore, because swarming effectively expands the population carrying capacity it is more robust than alternative cooperative strategies that improve growth rates but bring only transient benefits to the population. But we also find, as predicted by theory (West et al, 2006), that costly swarming cooperation can only evolve under stringent conditions of high genetic relatedness. This is the first time that high relatedness is shown to contribute to the evolution of swarming cooperation and compensate for an unnatural cost non-existing in the wild-type strain. Nonetheless, our results show that strong group benefits alone do not necessarily lead to the evolution of cooperative swarming under multilevel selection, whereas the native regulation by metabolic prudence greatly expands the conditions favoring cooperation.
Results and discussion
Swarming is favored by group-level selection but disfavored by individual-level selection
We use an engineered strain of P. aeruginosa that has been genetically altered such that the degree of its cooperative effort can be controlled. This strain (PA14 ΔrhlA:PBADrhlAB) has the biosurfactant synthesis genes placed under the regulation of the promoter PBAD so that their expression can be induced by adding L-arabinose to the medium (Boles et al, 2005). Previous experiments have shown that inducing biosurfactant with L-arabinose at 0.5% (w/v) has a significant cost in liquid cultures and swarming competitions (Xavier et al, 2011). We investigated the induction of swarming cooperation more extensively over a wide range of L-arabinose concentrations (Figure 2A). The results show that inducing surfactant secretion increases fitness of a swarming colony by enabling spreading over the plate (Figure 2B). However, at high induction levels (L-arabinose>0.25%) the metabolic costs of surfactant over-secretion start outweighing the benefits and the swarming colonies spread less. The final colony size, and thus the population fitness, peak at intermediate levels of induction (0.25% L-arabinose).
Figure 2.

Cooperative swarming allows Pseudomonas aeruginosa colonies to expand over large areas. (A) Swarming requires the secretion of biosurfactants. In colonies of the inducible cooperator, fitness peaks at L-arabinose 0.25%, a level at which biosurfactant production is high enough for swarming expansion but low enough that its costs do not overwhelm the benefit of spatial colony expansions. The data point at 0.5% L-arabinose (white bar) comes from a previous study (Xavier, 2011a); all other data (gray bars) were acquired in the present study. CFU = colony forming units. (B) Images of swarming colonies. Dashed line represents the edge of Petri dish with 9 cm diameter. (C) Cooperators (biosurfactant producers induced by 0.25% L-arabinose) expand over the entire Petri dish. Defectors (rhamnolipid-deficient knockouts) are incapable of swarming and the colonies grow less.
Next, we observed that swarming at 0.25% L-arabinose allows a colony of inducible biosurfactant producers to occupy a larger area on the plate and ultimately to grow better compared with colonies of a defector strain lacking biosurfactant secretion. While the swarming colony reaches the edge of a 9-cm wide Petri dish within 24 h, a colony of surfactant-deficient defectors cannot swarm and the colony stays confined to a region of <1 cm wide (Figure 2C). The limiting factors affecting bacterial growth on a plate have been extensively studied (Pirt, 1967; Cooper et al, 1968; Hochberg and Folkman, 1972). Briefly, the prevailing explanation is that a colony of immotile bacteria, such as our defectors, depletes local nutrients and a nutrient gradient is created as nutrients diffuse toward the colony (Nadell et al, 2010). As the region closest to the colony has the lowest concentration of nutrients, the growth of the colony is eventually limited by lack of nutrients. (An alternative mechanism that leads to equivalent outcomes is that toxic waste products accumulate at the immediate surroundings of the colony. Waste products diffuse away from the colony but create a gradient where the concentration is highest closest to the colony where it inhibits growth.) Swarming motility enables a colony to expand beyond the inoculation site, and thus escape the growth-limiting environment.
Comparing the growth of biosurfactant producers (cooperators) and defectors shows that cooperation has a clear benefit in single-strain colonies (Figure 2C). However, natural microbial populations are rarely monoclonal. Processes such as mixing with other strains or species and mutation introduce genetic variation and influence the evolution of microbial social traits (West et al, 2006). Therefore, we investigated whether swarming is still favored when induced cooperators are mixed with defectors within the same colony. For this experiment, we used strains labeled with constitutively expressed fluorescent markers (GFP or DsRed-Express) to allow strain identification (Xavier et al, 2011). In each competition, we mixed the two strains at a desired proportion and inoculated approximately one million cells in a 2-μl droplet onto soft agar and incubated for 24 h. The plates were then imaged (Figure 3A) and the final numbers of each strain were determined. We used the data to calculate the changes in cooperator proportion (measuring individual-level selection) and the final colony size (measuring population fitness). As a control, we compared the relative amounts of rhamnolipid secreted in liquid medium by the inducible strain when alone and mixed with 50% defectors to determine whether the presence of defectors affected biosurfactant production. The amount of rhamnose produced by the inducible strain alone is significantly different compared with the strain mixed with 50% defectors. However, when comparing half of the surfactant production by the inducible strain alone, the results are not significantly different (see Materials and methods for statistical analysis) from the rhamnose levels produced by the inducible strain mixed with 50% defectors, indicating that the surfactant production by the inducible strain is not affected by the presence of non-producing defectors (Supplementary Figure S1).
Figure 3.

Individual-level selection disfavors cooperation: induced cooperators increase population size but lose to defectors. (A) Competitions between cooperators and defectors mixed at varying mixing proportions (L-arabinose 0.25%). Cooperators are labeled with GFP (green) and defectors are labeled with DsRed-Express (red). (B) The change in cooperator proportion due to competition reveals that induced cooperation is strongly selected against. Lines represent best fit for mathematical model (see Materials and methods) and shaded areas represent confidence levels from bootstrapping. Size of data points is proportional to the weight of the data point in the parameter fitting. (C) Colony fitness increases with the proportion of cooperators but shows diminishing returns. (D) The growth yield of cooperators relative to the yield of defectors decreases with increasing levels of biosurfactant induction. Error bars represent minimum and maximum parameter value obtained from bootstrapping.
The mixed-strain competitions show that for every initial mixing proportion the proportion of cooperators decreased after competition, revealing that cooperators are disfavored by individual-level selection (Figure 3B). The change in cooperator proportion as a function of the initial proportion of cooperators exhibits an inverse bell shape that is typical for a trait that is disfavored in competition (Chuang et al, 2009). The population fitness, however, increased with the initial proportion of cooperators, confirming that cooperators benefit the entire colony (Figure 3C). Also, notable was that fitness plateaued for high initial cooperator proportions (above 0.5) indicating diminishing returns (Figure 3C).
Theory for multilevel selection
The swarming competitions using an induced cooperator (Figure 3) reveal that swarming cooperation benefits the population and would thus be favored by group-level selection. However, cooperators are outcompeted within colonies, which means that swarming is disfavored by individual-level selection. In nature, selection can occur simultaneously at multiple levels and the balance between the different levels ultimately determines the evolutionary fate of social traits (Keller, 1999). Could there be situations under which multilevel selection favors costly swarming cooperation?
To answer this question, we consider a theoretical scenario introduced by Hamilton (1975) and more recently applied by Chuang et al (2009) to investigate cooperation in a synthetic microbial system. A population of cooperators and defectors (the global population) is distributed heterogeneously into colonies (or local subpopulations) of varying mixing proportions. Each colony is allowed to compete and, at the end, all colonies are pooled together. The change in the total proportion of cooperators in the global population is given by the average across all subpopulations (Figure 4). It is important to note that this average is a weighted average that takes into account the fitness of each individual colony, because colonies with greater fitness contribute more to the global pool.
Figure 4.

The theoretical scenario used in multilevel selection analysis. A global population with an initial proportion of cooperators p is sampled to seed several subpopulations each with an initial local cooperator proportion pi (the average across all populations being p). Each subpopulation is allowed to compete for 24 h and at the end of the competition the proportion of cooperators has changed to pi’. In swarming motility, the subpopulations seeded with higher initial proportions of cooperators tend to produce larger colonies (wi increases with pi), and thus will contribute more to the global pool. After competition, all subpopulations are pooled together and the final proportion of cooperators is assessed, 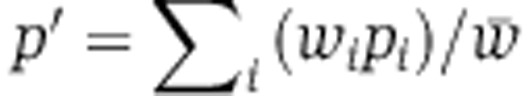 . Cooperation is favored by multilevel selection when Δp=p−p′>0.
. Cooperation is favored by multilevel selection when Δp=p−p′>0.
The approach is based on the multilevel selection framework of the Price equation (Price, 1970). Applied to our system, the Price equation partitions the global change in the proportion of cooperators, Δp, into its group-level and individual-level selection components:
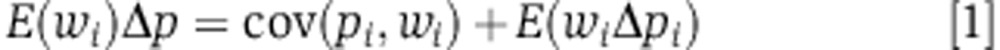 |
Here, wi is the fitness of a colony i and its value is a function of the proportion at which cooperators are initially mixed with defectors in that colony (pi). The change in cooperator proportion within the colony, Δpi, quantifies individual-level selection. E(wi) is the average fitness across all colonies that make up the global population. The Price equation highlights that, assuming that the initial population density is the same in all subpopulations, we need only to know the following three quantities to determine whether cooperators are favored by multilevel selection (Δp>0):
The change in cooperator proportion in a colony as a function of the mixing proportion, Δpi(pi).
The colony fitness as a function of the mixing proportion, wi(pi).
The distribution of cooperators across all colonies.
Points 1 and 2 are addressed using data from our swarming competitions (Figure 3B and C). To interpolate the data for the entire range 0⩽pi⩽1, we developed a population dynamics model (see Materials and methods) that we parameterized by fitting the experimental data (lines in Figure 3B and C). Although simpler statistical regressions could also be used to interpolate such data (Smith et al, 2010), a population dynamics model offers the advantage that the parameters obtained provide mechanistic insight. We tested the model by fitting two additional sets of competition experiments carried out at 0% L-arabinose (low levels of cooperation) and 4% L-arabinose (high levels of cooperation; Supplementary Figure S2). The parameters allowed us to calculate relative growth yields of cooperators and defectors as a function of the level of biosurfactant induction (Figure 3D). Consistent with induced surfactant synthesis carrying a metabolic burden (Xavier et al, 2011), the growth yield of cooperators decreased relative to that of defectors with the increasing level of surfactant induction. When the mathematical model is suitably parameterized (Supplementary Table 1) it provides functions for Δpi(pi) and wi(pi).
Point 3, the distribution of mixing proportions across all colonies, is crucial because it sets the genetic relatedness between the actors and recipients. Relatedness, as defined in Hamilton’s kin selection (Hamilton, 1964), quantifies the similarity of actors and recipients at loci relevant to cooperation (Smith et al, 2010). Relatedness is calculated here from the distribution of cooperator mixing proportions in the subpopulations using the following expression (Grafen, 1985; Damore and Gore, 2012):
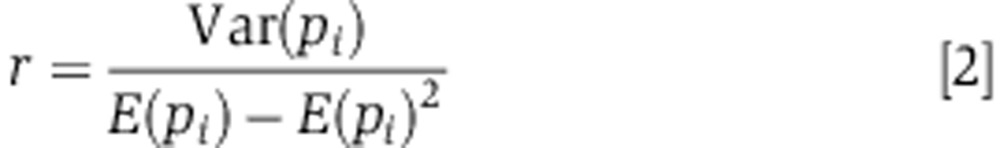 |
Equation 2 highlights the importance of the mean, E(pi), and variance, Var(pi), of the mixing proportions seeding the colonies. In the absence of any process generating variance, all colonies will be founded with the same proportion as the global population, E(pi)=p. In this situation, relatedness is null (r=0), individual-level selection prevails and costly cooperation is disfavored. When variance is extremely high and the two strains segregate entirely, for example if a single cell seeds each colony, relatedness is maximal (r=1) and group-level selection prevails. In real systems, relatedness may have intermediate values and be generated by different processes such as population bottlenecks (Chuang et al, 2009; Nadell et al, 2010), by cooperators physically sticking to each other (Smukalla et al, 2008) or other sources of population viscosity (Queller, 1994). Here, we keep our analysis general by using a statistical model based on a log-normal distribution for the ratio of cooperators to defectors (the logarithm of the cooperator-to-defector ratio follows a normal distribution with mean μ and standard deviation σ; see Materials and methods). Defined this way, the probability density function for mixing proportions, named f(pi), has the convenient feature that it transits gradually from a unimodal distribution (which corresponds to low segregation) to a bimodal distribution (strong segregation) by increasing a single parameter, σ (Figure 5A). The variance is a monotonically increasing function of σ (see Materials and methods) and, consequently, relatedness increases with σ (Equation 2, note that E(pi)=p≈1/(1+10−μ)). This model of population structure allows simulating scenarios where the mean fraction of cooperators varies while σ remains constant. We would expect σ to remain constant whenever there is a fixed population viscosity at small-length scales that is negligible for within-colony cooperation but high enough to influence seeding of the next round of competitions.
Figure 5.

Multilevel selection favors costly swarming only when relatedness is high. (A) Probability density function, f(pi), used to simulate variance in mixing proportions of cooperators. Parameter σ gradually changes the distribution from unimodal (low relatedness) to bimodal (high relatedness). (B) Multilevel selection simulations for a global proportion of cooperators of P=0.5 reveal that σ>1 favors cooperation. Simulations were compared with direct calculations from re-sampled data (blue stars and dashed line with circle, see Materials and methods). (C) Multilevel selection analysis for the entire range of global proportion of cooperators, 0⩽p⩽1, reveals that cooperation can reach fixation when σ>4.2.
With the three tools in hand, Δpi(pi), wi(pi) and f(pi), we investigated multilevel selection computationally in the following way: for a given proportion of cooperators in the global population, p, we generate subpopulations according to f(pi). We then use Δpi(pi) and wi(pi) to calculate the competition outcome in each subpopulation. Finally, we calculate the change in the global proportion of cooperators, Δp, using Price equation. Again, the final cooperator proportion corresponds to a simple calculation of the weighted average of pi across all subpopulations using the normalized colony fitness, wi/E(wi), as the weight parameter (equation 1).
Multilevel selection requires high relatedness to favor costly swarming
To understand whether multilevel selection can favor swarming cooperation, we first investigated a population with a global proportion of cooperators equal to 0.5 (Figure 5B). Multilevel selection simulations showed, as expected, that cooperators are outcompeted when the variance across all colonies is low (low σ, corresponding to a low relatedness). Keeping the global proportion fixed at 0.5 but increasing the variance increases cooperation advantage progressively: with higher variance, it is more likely to have colonies seeded with large proportions of cooperators, which favors cooperation. Cooperators are eventually favored when σ>1 (r>0.53).
We tested the simulation results with direct calculations from experimental data. These calculations did not rely on the mathematical model but were rather carried out by re-sampling the swarming competition data (Figure 3B and C) to re-create global populations with cooperator proportions of 0.5 and four different relatedness values (see Materials and methods). The calculations confirmed our simulation results (Figure 5B, data points).
Finally, we simulated multilevel selection for the full range of global cooperator proportions (Figure 5C). The simulations revealed that, because of the diminishing returns noted earlier (Figure 3C), intermediate values of σ produce an evolutionary equilibrium where cooperators and defectors coexist (Supplementary Figure S3B). The diminishing returns make it particularly difficult for cooperation to spread beyond the fraction at which the diminishing returns set in and, therefore, cooperators only reach fixation (Δp>0 for all values of p) when σ>4.2 (Supplementary Figure S3C). The corresponding value of relatedness is r>0.84 (calculated for p=0.5), which is a very high value. Intuitively, this means that if the process generating relatedness were stochastic sampling of a diluted population (Chuang et al, 2009) then subpopulations would have to be inoculated with an average of 0.65 cells/colony, an extremely low number. Swarming colonies inoculated by extremely low numbers of cells could potentially lead to very different results (a subject that we will address in a future study). The requirement for high relatedness is due to a combination of strong individual-level selection against cooperation (Figure 3B) and diminishing returns in the group-level benefits (Figure 3C; Supplementary Figure S3B). Consequently, costly swarming cooperation can be favored by multilevel selection to the point of reaching fixation but only when the genetic relatedness is very high.
Expansion-driven versus growth-driven cooperation
The calibrated multilevel selection model was also used to test new hypotheses computationally by simulating new competitions beyond the experimental conditions. We asked whether swarming cooperation could still be favored when subpopulations competed for longer periods. Simulations predict that longer competitions would not affect the outcome and that after 48-h competitions cooperators could still reach fixation for σ>4.2 (Supplementary Figure S4). We compared these simulations with a different cooperative scenario inspired by the study by Chuang et al (2009): growth-driven cooperation. In that system, a costly extracellular signal induces antibiotic resistance in its recipients. Such a trait boosts the population growth rate but does not cause population expansion. Therefore, as Chuang et al (2009) noted, the benefits are only felt if the competitions end during the limited time span of exponential growth. Whereas the benefits of swarming cooperation can last beyond the exponential growth phase, the population benefits of growth-driven cooperation should be transient (Figure 6A, see Materials and methods for model implementation).
Figure 6.

Growth-driven cooperation has transient benefits and is disfavored by multilevel selection when competitions are long. (A) Schematic illustrating that while swarming expands population carrying capacity and its benefits persist even after the growth phase, growth-driven cooperation, as demonstrated by Chuang et al (2009) on the other hand, boosts growth rate but its benefits are restricted to the duration of the growth phase. Red line represents defectors and green represents cooperator populations. (B) Growth-driven cooperation can be favored after short competitions lasting only during exponential phase. (C) If the competitions run long enough such that subpopulations reach carrying capacity, then the benefits of cooperation vanish and cooperation is disfavored.
Our simulations of growth-driven cooperation showed indeed that when growth-driven subpopulations are allowed to reach carrying capacity, such as in 48-h long simulations, the benefits of cooperation are lost and the cooperator strain loses globally irrespective of the relatedness value (Figure 6C). In summary, purely growth-driven cooperation, which does not increase carrying capacity, is likely to be disfavored when competitions are long enough. In contrast, swarming cooperation is robust to longer competitions because spatial expansion increases population carrying capacity, thus benefiting the colony in a time-independent way.
Prudent cooperation is widely favored by multilevel selection
Our multilevel selection analysis shows that swarming cooperation by a constitutive cooperator can be favored to fixation even when it is costly, but only when relatedness is high (Figure 5C). In the absence of an active mechanism to increase relatedness such as kin recognition or high population viscosity, the dependence on high relatedness is a stringent constraint that makes costly swarming unlikely in natural populations where strains and species mix (West et al, 2006). We therefore investigated the effect of multilevel selection on metabolic prudence, the native regulatory mechanism of rhamnolipid secretion in P. aeruginosa (Figure 1A). We carried out experimental competitions at a range of mixing proportions between the wild-type and the defector strain and, as we had done before for the inducible cooperator, we measured changes in wild-type proportion and colony fitnesses. The examination of a wide range of mixing proportions revealed that, in contrast to induced cooperators (Figure 3B and C), prudent wild-type cooperators were not disfavored by individual-level selection but actually had a marginal advantage (Figure 7A). Moreover, colony fitness increases steadily with the initial proportion of cooperators, showing comparatively little signs of diminishing returns (Figure 7B). We parameterized our dynamic model of swarming competitions for the wild type (Figure 7A and B, lines). In support of previous results (Xavier et al, 2011), the growth yields obtained from model fitting showed that, unlike induced cooperation, metabolic prudence can increase colony fitness without a detriment to cooperator yields (Figure 7C). We then analyzed multilevel selection. As expected from the marginal advantage in individual-level selection and the absence of diminishing returns, multilevel selection favored wild-type cooperators at all levels of variance in subpopulation mixing proportions. We also investigated whether the small advantage is essential for the success for wild-type cooperators. The results show that even if wild-type cells had no advantage at the individual level and had the same yield as defectors, the wild-type strain would still be favored after multi-level selection thanks to the absence of diminishing returns (Supplementary Figure S5). Furthermore, increasing colony variance (higher relatedness) enhances the advantage of prudent cooperators both in the original scenario where wild-type cells have an individual-level advantage and in the neutral theoretical case (Figure 7D and Supplementary Figure S5, respectively).
Figure 7.

Metabolic prudence is favored more widely than induced cooperation. (A) The change in prudent cooperator (wild-type) proportion due to swarming competitions shows a marginal individual-level advantage of prudent cooperators. (B) Colony fitness increases monotonically with cooperator proportion. (C) Comparison of calculated yields and maximum colony size between prudent (wild-type) and inducible cooperators induced at three levels of L-arabinose. Only the wild-type strain has both high yield and large colony expansion. (D) Multilevel selection of wild-type against the defector strain reveals that prudent cooperators are favored for all values of relatedness.
Conclusion
The evolution of cooperative traits is a central problem in biology (Pennisi, 2005). What are the evolutionary mechanisms favoring cooperation when costly cooperative traits that benefit other individuals cannot be favored by selection acting at the individual level alone? Microbial systems are becoming increasingly popular as models to address this question experimentally and in a quantitative way thanks to large population sizes, short generation times and the ability to manipulate traits genetically (West et al, 2006; Xavier, 2011b; Damore and Gore, 2012; Celiker and Gore, 2012a).
But while social evolution is learning considerably from the interface with microbiology the reverse may not be true. In the microbiology literature, social traits are often explained using group-selection arguments (Costerton et al, 1999; Monds and O’Toole, 2009; Lee et al, 2010). This logic, sometimes referred in the evolutionary literature as the original or ‘old’ type of group selection (West et al, 2007), was introduced by Wynne-Edwards in the 1960s and argues that cooperation is favored because groups of cooperative individuals are fitter than groups of selfish individuals (Wynne-Edwards, 1963). However, the same logic neglects that traits that are costly to individuals would be outcompeted within group selection (West et al, 2006; Foster and Bell, 2012). The solution to this problem comes from recognizing that natural selection acts at multiple levels of biological organization, an idea first introduced in Hamilton’s kin selection (Hamilton, 1964), and ‘modern’ forms of group selection (Traulsen and Nowak, 2006) are equivalent (Lehmann et al, 2007). The Price equation (Equation 1) is one generalization of this idea that, as we illustrate here, is particularly suited for the quantitative analysis of microbial social traits. Recognizing that selection acts at multiple levels is key to analyzing microbial social traits (West et al, 2006) can help reveal novel mechanisms stabilizing cooperation (e.g., Foster et al, 2004; Xavier and Foster, 2007; Ostrowski et al, 2008; Nadell et al, 2010; Nadell and Bassler, 2011; Xavier et al, 2011; Dandekar et al, 2012; Celiker and Gore, 2012b; Koschwanez et al, 2013), and can eventually inspire therapeutic strategies against pathogens (Foster, 2005; Brown et al, 2009; Boyle et al, 2013).
We combined quantitative experiments with mathematical modeling to identify conditions favoring costly cooperative swarming P. aeruginosa (Figure 3B and C). Using an engineered strain enabled us to manipulate cooperation and to show that in spite of individual-level costs, costly swarming can still be favored under specific conditions. However, the conditions allowing the spread of constitutive cooperation are limited. This is in part due to high costs of cooperation but also, to large extent, due to strong diminishing returns (Figure 3C), a feature that is likely to be common in cooperative systems (Foster, 2004). Because population-level benefits of constitutive cooperation level off when the cooperator proportion is p>0.5, it is difficult for cooperation to spread beyond this value (Supplementary Figure S3B). As a consequence, the evolution of constitutive cooperation can require extremely high relatedness, which is unlikely in natural populations where strains and species mix. We also show that metabolic prudence, the native mechanism to regulate rhamnolipid synthesis in P. aeruginosa (Xavier et al, 2011), is particularly advantageous because it provides a small direct benefit to cooperators (Figure 7A) and lacks the strong diminishing returns (Figure 7B). The marginal benefit of metabolic prudence in within-colony competitions is amplified by multilevel selection (Figure 7D), and greatly expands the conditions favoring swarming cooperation.
In our multilevel selection simulations, we assumed that the subdivision of the global population into subpopulations follows a distribution with the same variance at every round. This is a common assumption in social evolution models that can be plausible under certain conditions, for example, limited dispersal. It is also possible, however, that the variance in the formation of subpopulations would change from one competition round to the next, causing the distribution of subpopulations to fluctuate in time. In this scenario, we expect that the evolution of the wild-type strain would be favored even more compared with the inducible cooperator strain, because the wild type is able to win in each competition against defectors irrespective of how the global population is sampled.
A notable conclusion from our study is that the spatial expansion caused by swarming provides persistent benefits to a population (Figure 6C; Supplementary Figure S4). Population growth is always limited by a carrying capacity (e.g., Brown et al, 2004). Cooperative traits that, unlike swarming, boost growth rate without expanding carrying capacity can only have transient benefits, and therefore are less likely to be favored by multilevel selection when competitions are long (Figure 6A). In fact, Chuang et al (2009) had noted that, in their synthetic system, cooperators were favored by multilevel selection only if the subpopulations were sampled during the exponential growth phase; once the populations reached stationary phase the cooperative benefits would disappear. These results also relate to the long-studied differences between r and K selection, where r refers to the maximal intrinsic growth rate and K refers to carrying capacity (Pianka, 1970; MacArthur and Wilson, 2001). An r-strategy depends on traits associated with rapid growth (high fecundity, early maturity and short generation times) and is suitable for primary colonizers of new environments (Loya, 1976). However, in stable environments, quality overcomes quantity and K-strategies that grow slower but have larger carrying capacity have the competitive advantage and can displace r-selected species (Pianka, 1970). In the case of social traits, a cooperative trait that only increases growth rate is comparable to an r-strategy as its benefits are transient. Meanwhile, a cooperative trait that benefits the population by allowing it to grow to a larger carrying capacity is comparable to a K-strategy. Our work is also consistent with theory (Lehmann et al, 2006; Alizon et al, 2008; Houchmandzadeh and Vallade, 2012) and recent experimental studies (Datta et al, 2013; Van Dyken et al, 2013) showing that population range expansion can be a key factor in stabilizing the evolution of cooperation.
There are many examples of microbial social traits that expand carrying capacity even without explicit spatial expansion. Sharing of iron scavenging molecules (West et al, 2006) and digestive enzymes (Greig and Travisano, 2004; Griffin et al, 2004; Gore et al, 2009) can expand the achievable size of a nutrient limited population in shaken test tubes. Similarly, high-yield metabolic pathways allow a population to make more efficient use of nutrients and thus to achieve higher numbers in spite of slower growth (Pfeiffer et al, 2001; Kreft, 2004; Frank, 2010). Expansion of carrying capacity can thus be achieved by many different means in addition to spatial spreading. We therefore expect that cooperation by prudent expansion of carrying capacity, rather than fast growth, is more stable and more commonly found in nature.
Materials and methods
Bacterial strains and swarming assays
The construction of the Pseudomonas aeruginosa strains and their GFP and DsRed-Express varieties was described previously (Xavier et al, 2011). All bacteria were cultured in LB (Lysogeny Broth) liquid medium overnight, followed by triple washing 1 ml with saline buffer. Labeled strains were mixed at different ratios to inoculate in Petri dishes with soft agar (0.5% agar). Minimum medium for soft agar plates was prepared as described previously (Xavier et al, 2011) with the addition of L-arabinose when needed. Serial dilutions were done for each of the cell mixes and CFUs (colony forming units) were also counted out as previously described (Xavier et al, 2011). All incubations were at 37°C. Fluorescently labeled strains were counted from plates imaged with fluorescent scanner Amershan Typhoon 9400 (GE Healthcare). Color pictures of swarming colonies were obtained using the same fluorescent scanner.
Mathematical modeling
We model swarming competitions as two strains, cooperators (C) and defectors (D), competing for a finite nutrient source (N) in a closed system representing the Petri dish. The model consists of three simple ordinary differential equations:
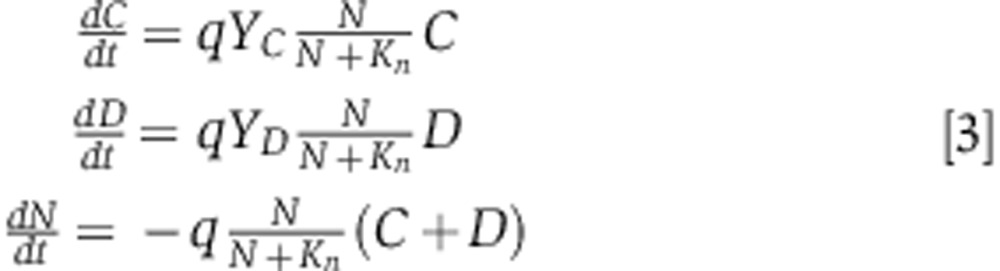 |
which implement Monod growth kinetics with Kn as the half-saturation constant (Monod, 1949). Both competing strains consume nutrients at the same rate, q, but cooperators may have a lower growth yield than defectors, YC<YD, if biosurfactant production is costly. If we now consider that 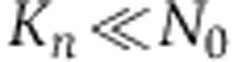 for N≠0:
for N≠0:
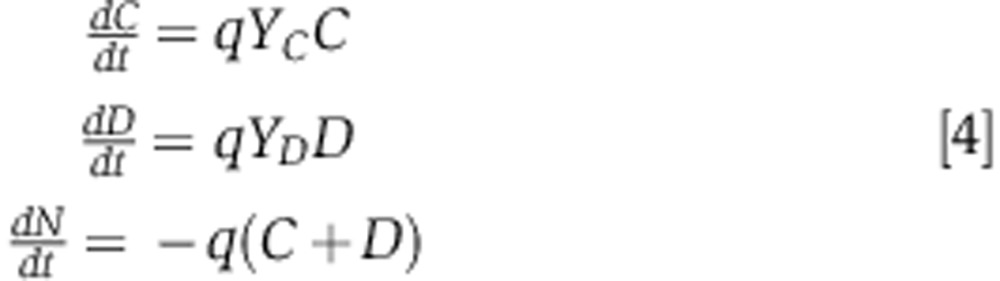 |
if N=0, then growth halts and 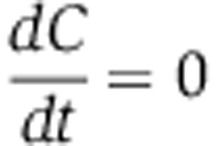 ,
, 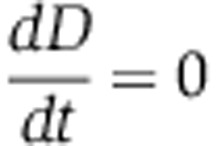 ,
, 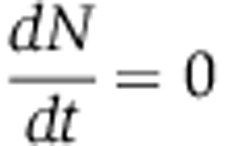 . Therefore, both strains grow exponentially while nutrients are available but stop growing once nutrients run out (zeroth order kinetics). We assume that the maximum amount of nutrients a colony will have access to (N0) is determined at the very beginning of the competition by the proportion of cooperators in the initial mixing of the two competing strains, pi=
. Therefore, both strains grow exponentially while nutrients are available but stop growing once nutrients run out (zeroth order kinetics). We assume that the maximum amount of nutrients a colony will have access to (N0) is determined at the very beginning of the competition by the proportion of cooperators in the initial mixing of the two competing strains, pi=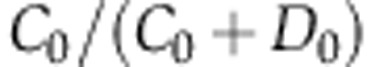 . We observed empirically that when pi⩽0.5, the achieved population size, and therefore N0, increases quickly with small pi increments, however saturates for pi≥0.5 (Figure 3b). Therefore, we describe N0 as a sigmoidal function that captures our empirical observation of diminishing returns:
. We observed empirically that when pi⩽0.5, the achieved population size, and therefore N0, increases quickly with small pi increments, however saturates for pi≥0.5 (Figure 3b). Therefore, we describe N0 as a sigmoidal function that captures our empirical observation of diminishing returns:
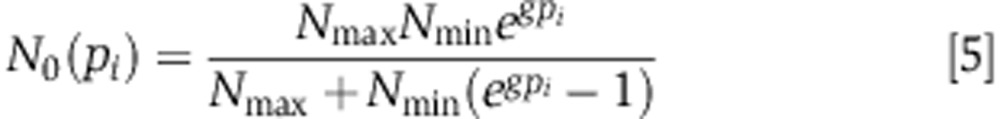 |
Nmin represents the amount of nutrients that a non-swarming population (pi=0) can take up. Nmax represents the maximum amount of nutrients that a swarming colony composed entirely of cooperators (pi=1) can consume. The coefficient g is the rate by which available nutrients increase as a function of pi. To simulate growth-driven cooperation (Figure 4), we used a modified version of our model such that the growth rates, not the available nutrients, increase in the presence of cooperators. N0 is constant and the rate of nutrient uptake q is a function of pi:
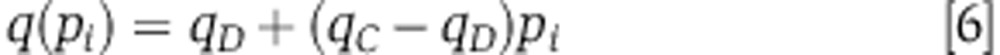 |
qD and qC are defined as the rate at which a population of only defectors or only cooperators takes up nutrients, respectively. In the simulations of growth-driven cooperation, we set qD=0.5 and qC=1 as a population of pure cooperators has a higher growth rate than a population of pure defectors. All simulations and parameter fitting (see supporting Table I) were carried out in Matlab (R2011a, the Mathworks).
Probability density function
The probability density function f(pi) is defined such that that the ratio of cooperators to defectors, C/D, follows a log-normal distribution. The random variable X=log10(C/D) is therefore normally distributed (X∼N(μ,σ)); and random variable pi=C/(C+D), the proportion of cooperators in the population, is a function of X such that pi=g(X)=(1+10−X)−1. The variance and mean of pi were approximated through the Delta method using first-order Taylor expansions of g(x) around X=μ. E(pi)≈(1+10−μ)−1 and Var(pi)≈g′(μ)2σ2. The probability density function of pi has the analytical expression:
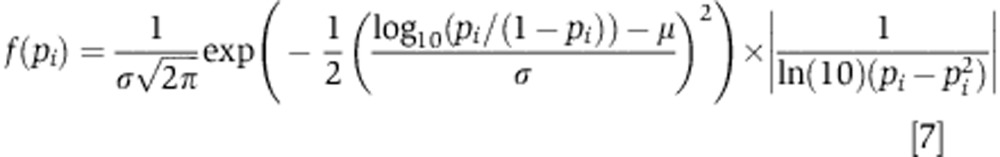 |
Direct calculations of multilevel selection
To confirm the prediction from multilevel selection (Figure 5B), we re-sampled experimental data by selecting equal number of data points with initial cooperator proportion (pi) of 17 and 83% to generate a global population with initial p=0.5. The final proportion of cooperators was used to calculate the global Δp as well and the relatedness value corresponding to σ. The same was done using the data points from initial local cooperator proportions (pi) of 1 and 99%. The data corresponding to initial pi=0 and pi=1 were used to determine the asymptotic value of Δp for r=1.
Surfactant quantification
The relative amount of rhamnolipids produced by the inducible strain in the absence and presence of defectors was quantified using an anthrone assay (Xavier et al, 2011) to assess rhamnose production in the defector, the wild-type and the inducible strains alone, as well as in a mix between the inducible strain+50% defectors in a liquid medium with the same composition of the swarming media. The rhamnose levels were normalized by subtracting the average amount detected from the defector strain (background) and dividing by the average amount produced by the wild-type strain. We used a Wilcoxon rank-sum test to evaluate whether rhamnolipid production by the inducible strain alone was significantly different from production in a mix with 50% defectors. The significance test showed that the inducible strain’s rhamnolipid production is different from the inducible+50% defector strain with P-value=0.028. We also found that when comparing half of the amount of secreted rhamnose by the inducible strain with that of the mix with 50% defectors, there was no significant difference with a P-value of 0.382.
Supplementary Material
Supplementary figures S1-5
Acknowledgments
We thank Kevin Foster, Giles Hooker, Edo Kussel, Silja Heilman, Dave van Ditmarsch, Carey Nadell, Carlos Carmona Fontaine, Vanni Bucci and Will Chang for comments and helpful discussions. This work was supported by a seed grant from the Lucille Castori Center for Microbes, Inflammation, and Cancer and by the Office of the Director, National Institutes of Health of the National Institutes of Health under Award Number DP2OD008440 to JBX. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author contributions: LVR and KEB performed experiments. LVR and JBX designed the study, analyzed the data and created the mathematical model. LVR performed computer simulations. LVR, KEB and JBX wrote manuscript and prepared figures.
Footnotes
The authors declare that they have no conflict of interest.
References
- Alizon S, Taylor P, Kokko H (2008) Empty sites can promote altruistic behavior. Evolution 62: 1335–1344 [DOI] [PubMed] [Google Scholar]
- Boles BR, Thoendel M, Singh PK (2005) Rhamnolipids mediate detachment of Pseudomonas aeruginosa from biofilms. Mol Microbiol 57: 1210–1223 [DOI] [PubMed] [Google Scholar]
- Boyle KE, Heilmann S, van Ditmarsch D, Xavier JB (2013) Exploiting social evolution in biofilms. Curr Opin Microbiol 16: 207–212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brannstrom A, Dieckmann U (2005) Evolutionary dynamics of altruism and cheating among social amoebas. Proc Biol Sci 272: 1609–1616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown JH, Gillooly JF, Allen AP, Savage VM, West GB (2004) Toward a metabolic theory of ecology. Ecology 85: 1771–1789 [Google Scholar]
- Brown SP, West SA, Diggle SP, Griffin AS (2009) Social evolution in micro-organisms and a Trojan horse approach to medical intervention strategies. Philos Trans R Soc Lond B Biol Sci 364: 3157–3168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caiazza NC, Shanks RM, O’Toole GA (2005) Rhamnolipids modulate swarming motility patterns of Pseudomonas aeruginosa. J Bacteriol 187: 7351–7361 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmona-Fontaine C, Xavier JB (2012) Altruistic cell death and collective drug resistance. Mol Syst Biol 8: 627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Celiker H, Gore J (2012a) Cellular cooperation: insights from microbes. Trends Cell Biol 23: 9–15 [DOI] [PubMed] [Google Scholar]
- Celiker H, Gore J (2012b) Competition between species can stabilize public-goods cooperation within a species. Mol Syst Biol 8: 621–621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang JS, Rivoire O, Leibler S (2009) Simpson’s paradox in a synthetic microbial system. Science 323: 272–275 [DOI] [PubMed] [Google Scholar]
- Chuang JS, Rivoire O, Leibler S (2010) Cooperation and Hamilton’s rule in a simple synthetic microbial system. Mol Syst Biol 6: 398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper AL, Dean ACR, Hinshelwood C (1968) Factors affecting the growth of bacterial colonies on agar plates. Proc Biol Sci 171: 175–199 [DOI] [PubMed] [Google Scholar]
- Costerton JW, Stewart PS, Greenberg EP (1999) Bacterial biofilms: A common cause of persistent infections. Science 284: 1318–1322 [DOI] [PubMed] [Google Scholar]
- Crespi BJ (2001) The evolution of social behavior in microorganisms. Trends Ecol Evol 16: 178–183 [DOI] [PubMed] [Google Scholar]
- Damore JA, Gore J (2012) Understanding microbial cooperation. J Theor Biol 299: 31–41 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dandekar AA, Chugani S, Greenberg EP (2012) Bacterial quorum sensing and metabolic incentives to cooperate. Science 338: 264–266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Datta MS, Korolev KS, Cvijovic I, Dudley C, Gore J (2013) Range expansion promotes cooperation in an experimental microbial metapopulation. Proc Natl Acad Sci USA 110: 7354–7359 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deziel E, Lepine F, Milot S, Villemur R (2003) rhlA is required for the production of a novel biosurfactant promoting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs), the precursors of rhamnolipids. Microbiology 149: 2005–2013 [DOI] [PubMed] [Google Scholar]
- Deziel E, Paquette G, Villemur R, Lepine F, Bisaillon J (1996) Biosurfactant production by a soil pseudomonas strain growing on polycyclic aromatic hydrocarbons. Appl Environ Microbiol 62: 1908–1912 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diggle SP, Griffin AS, Campbell GS, West SA (2007) Cooperation and conflict in quorum-sensing bacterial populations. Nature 450: 411–U417 [DOI] [PubMed] [Google Scholar]
- Fiegna F, Yu YT, Kadam SV, Velicer GJ (2006) Evolution of an obligate social cheater to a superior cooperator. Nature 441: 310–314 [DOI] [PubMed] [Google Scholar]
- Fortunato A, Strassmann JE, Santorelli L, Queller DC (2003) Co-occurrence in nature of different clones of the social amoeba, Dictyostelium discoideum. Mol Ecol 12: 1031–1038 [DOI] [PubMed] [Google Scholar]
- Foster KR (2004) Diminishing returns in social evolution: the not-so-tragic commons. J Evol Biol 17: 1058–1072 [DOI] [PubMed] [Google Scholar]
- Foster KR (2005) Biomedicine. Hamiltonian medicine: why the social lives of pathogens matter. Science 308: 1269–1270 [DOI] [PubMed] [Google Scholar]
- Foster KR (2011) The sociobiology of molecular systems. Nat Rev Genet 12: 193–203 [DOI] [PubMed] [Google Scholar]
- Foster KR, Bell T (2012) Competition, not cooperation, dominates interactions among culturable microbial species. Curr Biol 22: 1845–1850 [DOI] [PubMed] [Google Scholar]
- Foster KR, Shaulsky G, Strassmann JE, Queller DC, Thompson CR (2004) Pleiotropy as a mechanism to stabilize cooperation. Nature 431: 693–696 [DOI] [PubMed] [Google Scholar]
- Frank SA (2010) The trade-off between rate and yield in the design of microbial metabolism. J Evol Biol 23: 609–613 [DOI] [PubMed] [Google Scholar]
- Geisel N (2011) Constitutive versus responsive gene expression strategies for growth in changing environments. PLoS One 6: e27033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gore J, Youk H, van Oudenaarden A (2009) Snowdrift game dynamics and facultative cheating in yeast. Nature 459: 253–256 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grafen A (1985) A geometric view of relatedness. InOxford Surveys in Evolutionary Biology Dawkins R, Ridley M (eds)New York, USA: Oxford University Press, [Google Scholar]
- Greenberg EP (2010) Sociomicrobiology: a personal perspective on an emerging research area. Microbe Magazine 5: 206–211 [Google Scholar]
- Greig D, Travisano M (2004) The Prisoner’s Dilemma and polymorphism in yeast SUC genes. Proc Biol Sci 271: S25–S26 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffin AS, West SA, Buckling A (2004) Cooperation and competition in pathogenic bacteria. Nature 430: 1024–1027 [DOI] [PubMed] [Google Scholar]
- Hamilton WD (1964) The genetical evolution of social behaviour I & II. J Theor Biol 7: 1–52 [DOI] [PubMed] [Google Scholar]
- Hamilton WD (1975) Innate social aptitudes of man: an approach from evolutionary genetics. InBiosocial Anthropology Fox R (ed), pp133–153London UK: Malaby Press, [Google Scholar]
- Hochberg MS, Folkman J (1972) Mechanism of size limitation of bacterial colonies. J Infect Dis 126: 629–635 [DOI] [PubMed] [Google Scholar]
- Houchmandzadeh B, Vallade M (2012) Selection for altruism through random drift in variable size populations. BMC Evol Biol 12: 61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller L (1999) Levels of Selection in Evolution Princeton University Press, : Princeton, NJ, [Google Scholar]
- Kolter R, Greenberg EP (2006) Microbial sciences: the superficial life of microbes. Nature 441: 300–302 [DOI] [PubMed] [Google Scholar]
- Koschwanez JH, Foster KR, Murray AW (2011) Sucrose utilization in budding yeast as a model for the origin of undifferentiated multicellularity. PLoS Biol 9: e1001122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koschwanez JH, Foster KR, Murray AW (2013) Improved use of a public good selects for the evolution of undifferentiated multicellularity. Elife 2: e00367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreft JU (2004) Biofilms promote altruism. Microbiology 150: 2751–2760 [DOI] [PubMed] [Google Scholar]
- Kummerli R, Brown SP (2010) Molecular and regulatory properties of a public good shape the evolution of cooperation. Proc Natl Acad Sci USA 107: 18921–18926 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kummerli R, Gardner A, West SA, Griffin AS (2009a) Limited dispersal, budding dispersal, and cooperation: an experimental study. Evolution 63: 939–949 [DOI] [PubMed] [Google Scholar]
- Kummerli R, Griffin AS, West SA, Buckling A, Harrison F (2009b) Viscous medium promotes cooperation in the pathogenic bacterium Pseudomonas aeruginosa. Proc Biol Sci 276: 3531–3538 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kummerli R, Jiricny N, Clarke LS, West SA, Griffin AS (2009c) Phenotypic plasticity of a cooperative behaviour in bacteria. J Evol Biol 22: 589–598 [DOI] [PubMed] [Google Scholar]
- Lee HH, Molla MN, Cantor CR, Collins JJ (2010) Bacterial charity work leads to population-wide resistance. Nature 467: 82–85 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehmann L, Keller L, West S, Roze D (2007) Group selection and kin selection: Two concepts but one process. Proc Natl Acad Sci USA 104: 6736–6739 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehmann L, Perrin N, Rousset F (2006) Population demography and the evolution of helping behaviors. Evolution 60: 1137–1151 [PubMed] [Google Scholar]
- Loya Y (1976) The Red Sea coral Stylophora pistillata is an r strategist. Nature 259: 478–480 [Google Scholar]
- MacArthur R, Wilson E (2001) The Theory of Island Biogeography (Princeton Landmarks in Biology) Princeton NJ: Princeton University Press, [Google Scholar]
- Monds RD, O’Toole GA (2009) The developmental model of microbial biofilms: ten years of a paradigm up for review. Trends Microbiol 17: 73–87 [DOI] [PubMed] [Google Scholar]
- Monod J (1949) The growth of bacterial cultures. Annu Rev Microbiol 3: 371–394 [Google Scholar]
- Nadell CD, Bassler BL (2011) A fitness trade-off between local competition and dispersal in Vibrio cholerae biofilms. Proc Natl Acad Sci USA 108: 14181–14185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadell CD, Foster KR, Xavier JB (2010) Emergence of spatial structure in cell groups and the evolution of cooperation. PLoS Comput Biol 6: e1000716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadell CD, Xavier JB, Foster KR (2009) The sociobiology of biofilms. FEMS Microbiol Rev 33: 206–224 [DOI] [PubMed] [Google Scholar]
- Nadell CD, Xavier JB, Levin SA, Foster KR (2008) The evolution of quorum sensing in bacterial biofilms. PLoS Biol 6: e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostrowski EA, Katoh M, Shaulsky G, Queller DC, Strassmann JE (2008) Kin discrimination increases with genetic distance in a social amoeba. PLoS Biol 6: e287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pennisi E (2005) How did cooperative behavior evolve? Science 309: 93. [DOI] [PubMed] [Google Scholar]
- Perkins TJ, Swain PS (2009) Strategies for cellular decision-making. Mol Syst Biol 5: 326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeiffer T, Schuster S, Bonhoeffer S (2001) Cooperation and competition in the evolution of ATP-producing pathways. Science 292: 504–507 [DOI] [PubMed] [Google Scholar]
- Pianka ER (1970) On r- and K-Selection. Am Nat 104: 592–597 [Google Scholar]
- Pirt SJ (1967) A kinetic study of the mode of growth of surface colonies of bacteria and fungi. J Gen Microbiol 47: 181–197 [DOI] [PubMed] [Google Scholar]
- Price GR (1970) Selection and covariance. Nature 227: 520–521 [DOI] [PubMed] [Google Scholar]
- Queller D (1994) Genetic relatedness in viscous populations. Evol Ecol 8: 70–73 [Google Scholar]
- Rainey PB, Rainey K (2003) Evolution of cooperation and conflict in experimental bacterial populations. Nature 425: 72–74 [DOI] [PubMed] [Google Scholar]
- Ratcliff WC, Denison RF, Borrello M, Travisano M (2012) Experimental evolution of multicellularity. Proc Natl Acad Sci USA 109: 1595–1600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross-Gillespie A, Gardner A, Buckling A, West SA, Griffin AS (2009) Density dependence and cooperation: theory and a test with bacteria. Evolution 63: 2315–2325 [DOI] [PubMed] [Google Scholar]
- Ross-Gillespie A, Gardner A, West SA, Griffin AS (2007) Frequency dependence and cooperation: theory and a test with bacteria. Am Nat 170: 331–342 [DOI] [PubMed] [Google Scholar]
- Sathe S, Kaushik S, Lalremruata A, Aggarwal RK, Cavender JC, Nanjundiah V (2010) Genetic heterogeneity in wild isolates of cellular slime mold social groups. Microb Ecol 60: 137–148 [DOI] [PubMed] [Google Scholar]
- Smith J, Van Dyken JD, Zee PC (2010) A generalization of Hamilton’s rule for the evolution of microbial cooperation. Science 328: 1700–1703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smukalla S, Caldara M, Pochet N, Beauvais A, Guadagnini S, Yan C, Vinces MD, Jansen A, Prevost MC, LatgÈ J-P, Fink GR, Foster KR, Verstrepen KJ (2008) FLO1 is a variable green beard gene that drives biofilm-like cooperation in budding yeast. Cell 135: 726–737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strassmann JE, Queller DC (2011) Evolution of cooperation and control of cheating in a social microbe. Proc Natl Acad Sci 108: 10855–10862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strassmann JE, Zhu Y, Queller DC (2000) Altruism and social cheating in the social amoeba Dictyostelium discoideum. Nature 408: 965–967 [DOI] [PubMed] [Google Scholar]
- Traulsen A, Nowak MA (2006) Evolution of cooperation by multilevel selection. Proc Natl Acad Sci USA 103: 10952–10955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Dyken JD, Muller MJ, Mack KM, Desai MM (2013) Spatial population expansion promotes the evolution of cooperation in an experimental Prisoner’s Dilemma. Curr Biol 23: 919–923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- West SA, Griffin AS, Gardner A (2007) Social semantics: altruism, cooperation, mutualism, strong reciprocity and group selection. J Evol Biol 20: 415–432 [DOI] [PubMed] [Google Scholar]
- West SA, Griffin AS, Gardner A, Diggle SP (2006) Social evolution theory for microorganisms. Nat Rev Microbiol 4: 597–607 [DOI] [PubMed] [Google Scholar]
- Wynne-Edwards VC (1963) Animal dispersion in relation to social behaviour. Can Med Assoc J 88: 1255–1256 [PMC free article] [PubMed] [Google Scholar]
- Xavier JB (2011a) Social interaction in synthetic and natural microbial communities. Mol Syst Biol 7: 483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xavier JB (2011b) Social interaction in synthetic and natural microbial communities. Mol Syst Biol 7: 483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xavier JB, Foster KR (2007) Cooperation and conflict in microbial biofilms. Proc Natl Acad Sci USA 104: 876–881 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xavier JB, Kim W, Foster KR (2011) A molecular mechanism that stabilizes cooperative secretions in Pseudomonas aeruginosa. Mol Microbiol 79: 166–179 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary figures S1-5