

Abstract
The present study examined the dose-dependent effect of vitamin E in reversing bone loss in ovariectomized (Ovx) rats. Sprague-Dawley rats were either Sham-operated (Sham) or Ovx and fed control diet for 120 days to lose bone. Subsequently, rats were divided into 5 groups (n = 12/group): Sham, Ovx-control, low dose (Ovx + 300 mg/kg diet; LD), medium dose (Ovx + 525 mg/kg diet; MD), and high dose (Ovx + 750 mg/kg diet; HD) of vitamin E and sacrificed after 100 days. Animals receiving MD and HD of vitamin E had increased serum alkaline phosphatase compared to the Ovx-control group. Bone histomorphometry analysis indicated a decrease in bone resorption as well as increased bone formation and mineralization in the Ovx groups supplemented with MD and HD of vitamin E. Microcomputed tomography findings indicated no effects of vitamin E on trabecular bone of fifth lumbar vertebrae. Animals receiving HD of vitamin E had enhanced fourth lumbar vertebra quality as evidenced by improved ultimate and yield load and stress when compared to Ovx-control group. These findings demonstrate that vitamin E improves bone quality, attenuates bone resorption, and enhances the rate of bone formation while being unable to restore bone density and trabecular bone structure.
1. Introduction
Ovarian hormone deficiency is a major risk factor for osteoporosis in women [1, 2]. Although there are several FDA-approved medications to either prevent or reverse osteoporosis, women continue to look for safer and more feasible preventative and therapeutic alternatives [3]. In postmenopausal women, the rate of bone turnover increases with the rate of bone resorption exceeding that of bone formation which results in net bone loss [4]. These events, in part, have been linked to the modulation of immune cell mediators (cytokines and prostaglandins) and oxygen-derived free radical (ODFR) formation either in the bone microenvironment or in the cells that serve as osteoclastic precursors such as those of the monocyte-macrophage lineage [5].
A plethora of local cytokines and lipid mediators such as interleukin (IL)-1, IL-6, tumor necrosis factor-alpha (TNF-α), lymphotoxin, leukotrienes, and prostaglandins of the E series (PGE) are involved in regulating bone metabolism [6–8]. Ovarian hormone deficiency is associated with increased activation of certain immune cells leading to an increase in inflammatory mediators, such as IL-1 [9, 10], IL-6 [9, 11], and in particular PGE2 [12–17], which is in part responsible for suppressing osteoblastic activity stimulating osteoclastic differentiation, and activity resulting in an increase in bone resorption [18].
Oxygen-derived free radicals are also formed by a number of activated immune cells, including monocytes, macrophages, and neutrophils [19], and have been reported to be increased in chronic inflammatory diseases [20], aging [21], and osteoporosis [22, 23]. In vivo and in vitro findings [22–24] indicate that excessive free radicals generated in the bone environment increase osteoclastogenesis and bone resorption resulting in reduced bone mineral density (BMD) and osteoporosis [25]. A role of oxidative stress in estrogen-deficiency-induced bone loss has also been demonstrated as ovariectomy significantly reduces two of the primary thiol antioxidants, glutathione and thioredoxin, in osteoclasts [26, 27].
Vitamin E, a fat-soluble antioxidant [26, 28, 29], has been of particular interest due to its ability to suppress the production of certain proinflammatory mediators such as PGE2, TNF-α, IL-1, and IL-6 [30–34] that have been linked to increased bone loss [6–8, 35], and protect against oxidative damage caused by ODFRs [29, 36]. The relationship between vitamin E and bone has been studied to some extent. Xu et al. [37] reported that vitamin E maintains normal bone growth and bone modeling in young animals and protects cartilage against cellular lipid peroxidation. They also found that supplemental vitamin E increased bone mass by lowering the concentrations of free radicals that stimulate bone resorption and suppress bone formation. Vitamin E also reduced the ODFR stimulation of osteoclastic bone resorption [22]. Furthermore, the administration of α-tocopherol was shown to suppress the production of ODFRs in the early stages of fracture healing in rabbits [38], increase the activity of antioxidant enzymes in Ovx rats [39], and enhance bone fracture healing in its early [40] and late phases [41] in Ovx rats and in male rats [42]. In addition, the results of a population study by Melhus et al. [43] indicated the important role of adequate intake of dietary vitamin E in reducing the risk of hip fracture in current smokers. Another cross-sectional study suggests that vitamin E may uncouple bone turnover in postmenopausal women, causing an increase in the rate of bone formation while having no effects on the rate of bone resorption [44]. Collectively, these observations suggest that vitamin E positively influences skeletal health.
Previously, we demonstrated that supplemental levels of vitamin E improve bone quality as well as augment bone matrix protein without altering bone density in old male mice [45] and orchidectomized (Orx) male rats [46], both of which are well-accepted models of male, age-related osteoporosis. Furthermore, we have observed that supplemental doses of vitamin E have protective effects on various bone parameters in hindlimb unloaded male rat model of osteoporosis but not in ambulatory control rats [47]. Additionally, Norazlina et al. [48] reported that vitamin E is able to maintain BMD in Ovx rats. These findings imply that animals under physical stress and hormonal deficiency may benefit from additional vitamin E in the diet. Thus, the purpose of this study was to investigate whether vitamin E dose-dependently reverses bone loss in an Ovx rat model of osteoporosis.
2. Materials and Methods
2.1. Animals and Diets
Seventy-five 12-month-old female Sprague-Dawley rats were housed in an environmentally controlled animal laboratory upon arrival. Guidelines for the ethical care and treatment of animals from the Animal Care and Use Committee were strictly followed. After 5 days of acclimation, rats were either sham-operated (Sham; 1 group; n = 12) or ovariectomized (Ovx; 4 groups). All rats were maintained on a semi-purified casein-based diet (AIN 93M) containing basal levels of vitamin E (75 mg vitamin E/kg diet) for a period of 120 days to induce osteopenia [49]. Ovariectomized rats were then randomly divided into 4 groups (n = 12/group): Ovx control, and Ovx + 300 mg vitamin E/kg diet (low dose, LD), Ovx + 525 mg vitamin E/kg diet (medium dose, MD), Ovx + 750 mg vitamin E/kg diet (high dose, HD). The animals were pair-fed to the average food intake of Sham group and had free access to distilled deionized water. Rats were fed their respective dietary treatments for 100 days and then sacrificed at the end of the treatment period.
2.2. Necropsy and Tissue Processing
One-hundred days after treatment rats were anesthetized with a mixture of ketamine/xylazine (60 and 3 mg/kg body weight, resp.), and whole body BMD as well as bone mineral content (BMC) were measured using dual energy X-ray absorptiometry (DXA; QDR-4500A Elite; Hologic, Waltham, MA, USA). Rats were then bled via the abdominal aorta. Blood samples were collected, and serum was separated by centrifugation at 1500 ×g for 20 min at 4°C. Aliquots of serum were frozen and kept at −20°C for later analyses. The tibiae and the lumbar vertebrae were removed for various analyses. The tibiae were stored in 70% ethanol for histomorphometry while fourth and fifth lumbar vertebrae (L4 and L5, resp.) were frozen at −20°C for biomechanical properties and microstructural analysis, respectively. The uterus of each rat was collected, blotted, and weighed to confirm the success of ovariectomy.
2.3. Serum Vitamin E Analysis
Serum α-tocopherol was determined using the method of Bieri et al. [50]. Briefly, samples and standards were extracted with hexane, evaporated under nitrogen, and reconstituted with diethylether/methanol (1 : 3, v/v). Alpha-tocopherol acetate solution (5 mg/mL) (Sigma Chemical Co., St. Louis, MO, USA) was added to 200 μL of serum or α-tocopherol acetate (Sigma Chemical Co., St. Louis, MO, USA) which was used as an internal standard. Vitamin E was measured in duplicate via HPLC (Waters Corporation, Milford, MA, USA) using a Nova-Pak C18 column (3.9 × 150 mm, 4 μm particle size; Waters Corporation), methanol : water (95 : 5, v/v) as the mobile phase, and absorbance was monitored at 290 nm. The intra- and interassay coefficients of variation were 4.3% and 4.6%, respectively.
2.4. Serum Alkaline Phosphatase (ALP) Activity
Serum ALP activity was determined colorimetrically using a kit from Sigma Diagnostics (St. Louis, MO, USA). This test was performed on a Cobas-Fara II Clinical Analyzer (Montclair, NJ, USA). ALP activity was measured from the time-dependent increase in absorbance at 410 nm due to formation of nitrophenol. The intra- and interassay coefficients of variation of this procedure were 1.9% and 2.8%, respectively.
2.5. Bone Histomorphometry
Quantitative histomorphometry of the right tibia was performed to examine the dynamics of bone changes and bone cell activities in response to the ovariectomy and treatments. Briefly, calcein (8 mg/kg; Sigma, St. Louis, MO, USA) was injected at 8 days and 1 day, before necropsy, in order to prelabel the skeleton of the rats. The tibiae were cut at 1 mm distal to the tibio-ibula junction (TFJ) and 19 mm proximal to the TFJ to obtain a proximal portion for cancellous bone histomorphometry and fixed in 70% ethanol. The proximal tibiae (5 μm thick) were prestained in Villanueva stain, dehydrated in graded ethanol and acetone, and individually embedded in methyl methacrylate. Goldner-stained specimen was used for determining static and structural measurements while the Villanueva-stained specimen was used for assessing fluorochrome labeling and dynamic measurements of bone formation. A saw microtome (Model 1600, Leica, Germany) was used to cut the embedded samples (2–4 μm thick). Sections were analyzed semiautomatically using BIOQUANT software (R&M Biometrics, Nashville, TN, USA). Standard nomenclature was used to express the histomorphometric values. The data collection area contained only trabecular bone and bone marrow; the primary spongiosa was not included.
2.6. Assessment of Trabecular Bone Structures
The microarchitectural trabecular bone structure of L5 vertebra specimens was evaluated using microcomputed tomography (μCT40, Scanco Medical, Switzerland). Lumbar vertebrae were scanned from the caudal to cranial end (530 slices; 16 μm/slice). This region included 530 images obtained from each vertebra using the same isotropic voxel resolution and integration time as described with the tibiae. The volume of interest selected was 25 slices away from the appearance of the growth plate at each end of the vertebral body resulting in approximately 300 slices. The trabecular region was separated from the cortical bone using semiautomated contours. The trabecular bone morphometric parameters assessed with μCT included the BV expressed as a percentage of total volume (BV/TV), trabecular number (Tb.N), trabecular thickness (Tb.Th), and trabecular separation (Tb.Sp). Nonmetric parameters included structure model index (SMI) which is an indicator of plate-rod arrangement of the bone structure and connectivity density (Conn.D).
2.7. Biomechanical Properties
L4 vertebra specimens were cleaned and freed of surrounding soft tissues. BMD was estimated using DXA, equipped with appropriate software for BMD assessment in small laboratory animals.
Biomechanical properties were assessed as described elsewhere [51]. Briefly, vertebral strength was measured at a rate of 3 mm/min using Instron 5543 (Canton, MA, USA). Vertebrae were tested in compression with force applied to the cranial and caudal surfaces. The load-displacement curve was recorded during the test. The ultimate load, yield load, and the stiffness of each specimen were measured from the load-displacement curve. Ultimate stress, yield stress, and flexural modulus of elasticity were calculated from the beam-bending equations.
2.8. Statistical Analysis
Statistical analysis of data involved estimation of means and standard error of the mean (SEM) for each of the groups. Analysis of variance was performed using the mixed model procedure of SAS (version 9.2, SAS Institute, Cary, NC, USA). If significant F-values were reported, a post-hoc test with a Tukey adjustment was performed for multiple comparison purposes. Significance was set at P < 0.05.
3. Results
3.1. Food Intake, Body Weight, and Uterus Weight
Even though rats were pair-fed to the Sham group, all Ovx rats had significantly higher final body weights (P = 0.0003). The vitamin E treatment had no effect on body weight (Table 1). Ovariectomy, as expected, caused atrophy of uterine tissue (P < 0.0001) confirming bilateral ovariectomy (Table 1).
Table 1.
Effects of ovariectomy (Ovx) and three doses of vitamin E: low dose (LD, 300 mg/kg diet), medium dose (MD, 525 mg/kg diet), and high dose (HD, 750 mg/kg diet) on body and tissue weights, serum vitamin E, and serum ALP activity.
| Parameters | Sham | Ovx | LD | MD | HD |
|---|---|---|---|---|---|
| Body Weight (g) | |||||
| Initial | 299.3 ± 11.8 | 307.9 ± 8.9 | 310.4 ± 9.2 | 307.7 ± 8.9 | 308.6 ± 8.9 |
| Final | 328.0 ± 20.19a | 409.8 ± 11.27b | 406.0 ± 10.37b | 392.9 ± 10.35b | 394.9 ± 8.51b |
| Uterine weight (g) | 0.710 ± 0.035a | 0.141 ± 0.027b | 0.209 ± 0.031b | 0.144 ± 0.029b | 0.158 ± 0.028b |
| Vitamin E (mg/mL) | 26.78 ± 2.96ab | 22.47 ± 2.24b | 31.32 ± 2.41a | 29.51 ± 2.24a | 30.98 ± 2.32a |
| ALP (U/L) | 38.63 ± 2.59a | 46.79 ± 2.89a | 56.54 ± 3.68a | 60.29 ± 5.64b | 61.23 ± 4.46b |
Values within a row that share the same superscript letters are not significantly (P ≥ .05) different. Values are means ± S.E.M.
ALP: alkaline phosphatase.
3.2. Serum Vitamin E Levels and ALP Activity
The mean serum level of vitamin E in rats that received supplemental vitamin E was significantly higher (P = 0.04) than in Ovx control group (Table 1). Also, when compared to Ovx control group, MD and HD of vitamin E significantly (P = 0.0039) increased serum ALP activity by approximately 28 and 30%, respectively.
3.3. Bone Mineral Density and Content
As expected, ovariectomy significantly (P = 0.04) decreased whole-body BMD, but there were no differences (P = 0.88) in BMD among the four Ovx groups due to vitamin E treatment (Table 2). No significant (P = 0.28) differences were observed in whole-body BMC among treatment groups (Table 2).
Table 2.
Effects of ovariectomy (Ovx) and three doses of vitamin E: low dose (LD, 300 mg/kg diet), medium dose (MD, 525 mg/kg diet), and high dose (HD, 750 mg/kg diet) on DXA bone parameters of whole body and fourth lumbar vertebrae (L4).
| Parameters | Sham | Ovx | LD | MD | HD |
|---|---|---|---|---|---|
| Whole body (g) | |||||
| Baseline | |||||
| BMC (g) | 10.3 ± 0.38 | 10.55 ± 0.20 | 10.84 ± 0.18 | 10.57 ± 0.22 | 10.73 ± 0.21 |
| BMD (g/cm2) | 0.1774 ± 0.0029 | 0.1811 ± 0.0030 | 0.1806 ± 0.0013 | 0.1790 ± 0.0020 | 0.1799 ± 0.0018 |
| Final | |||||
| BMC (g) | 11.14 ± 0.40 | 11.72 ± 0.22 | 11.98 ± 0.16 | 11.59 ± 0.26 | 11.8 ± 0.20 |
| BMD (g/cm2) | 0.1734 ± 0.0029a | 0.1657 ± 0.0021a | 0.1635 ± 0.0013ab | 0.1649 ± 0.0020a | 0.1644 ± 0.0018ab |
| L4 vertebra | |||||
| BMD (g/cm2) | 0.240 ± 0.005a | 0.199 ± 0.004b | 0.202 ± 0.004b | 0.199 ± 0.004b | 0.199 ± 0.004b |
Values within a row that share the same superscript letters are not significantly (P ≥ .05) different. Values are means ± S.E.M.
BMC: bone mineral content; BMD: bone mineral density.
3.4. Bone Histomorphometry
Based on bone static histomorphometry Ovx control animals lost approximately 75% of their proximal tibia cancellous bone volume (BV/TV) over the duration of the study (220 days). This loss was not ameliorated by any of the supplemental doses of vitamin E (Table 3). Ovariectomy caused an increase in the values of bone resorption parameters including osteoclast/bone surface (Oc.S/BS) and eroded surface/bone surface (ES/BS) by 76 and 79%, respectively. However, MD as well as HD of vitamin E significantly prevented these increases (Table 3).
Table 3.
Effects of ovariectomy (Ovx) and three doses of vitamin E: low dose (LD, 300 mg/kg diet), medium dose (MD, 525 mg/kg diet), and high dose (HD, 750 mg/kg diet) on the proximal tibial metaphysis cancellous bone cellular activity.
| Parameters | Sham | Ovx | LD | MD | HD |
|---|---|---|---|---|---|
| BV/TV | 12.010 ± 1.526a | 3.020 ± 1.649b | 2.470 ± 1.238b | 1.865 ± 1.238b | 2.022 ± 1.238b |
| Bone resorption parameters | |||||
| Oc.S/BS (%) | 1.276 ± 0.325a | 2.252 ± 0.347b | 1.841 ± 0.347ab | 1.265 ± 0.347a | 1.008 ± 0.347a |
| ES/BS (%) | 1.473 ± 0.375a | 2.651 ± 0.393b | 1.885 ± 0.393a | 1.612 ± 0.393a | 1.218 ± 0.393a |
| Bone formation parameters | |||||
| MS/BS (%) | 4.842 ± 2.144a | 13.97 ± 2.23b | 19.942 ± 2.236c | 21.002 ± 2.236c | 13.137 ± 2.236b |
| BFR/BV (%/yr) | 0.117 ± 0.370a | 1.295 ± 0.380b | 1.909 ± 0.380bc | 2.148 ± 0.380c | 1.456 ± 0.380bc |
Values within a row that share the same superscript letters are not significantly (P ≥ .05) different. Values are means ± S.E.M.
BV/TV: bone volume as percentage of tissue volume; Oc.S/BS: osteoclast/bone surface; ES/BS: eroded surface/bone surface; MS/BS: mineralizing surface/bone surface; BFR/BV: bone formation rate/bone volume.
Bone dynamic histomorphometry indicated that bone formation parameters including mineralizing surface/bone surface (MS/BS) and bone formation rate/bone volume (BFR/BV) were significantly increased due to ovariectomy (Table 3). Supplemental vitamin E at LD and MD increased MS/BS by 42 and 50%, respectively, when compared to the Ovx control group. BFR/BV was significantly increased by 65% in the group receiving the MD of vitamin E when compared to Ovx control group (Table 3).
3.5. Trabecular Microarchitectural Properties of L5 Vertebrae
The effects of Ovx and supplemental doses of vitamin E on trabecular microarchitectural parameters of L5 vertebra bone are presented in Table 4. Ovariectomy decreased BV/TV, Tb.N, and Tb.Th by 37, 11, and 21%, respectively. As expected, the value of Tb.Sp and Conn.D increased by 14 and 29%, respectively, due to ovariectomy. There were no effects of vitamin E treatment observed on BV/TV, Tb.N, Tb.Th, and Tb.Sp indicating that despite the favorable changes in osteoclast and osteoblast activity observed in this study, supplementation with vitamin E at this point in time had not reversed Ovx-induced alterations in trabecular bone microarchitectural parameters. The results also indicate no treatment effect of vitamin E on SMI.
Table 4.
Effects of ovariectomy (Ovx) and three doses of vitamin E: low dose (LD, 300 mg/kg diet), medium dose (MD, 525 mg/kg diet), and high dose (HD, 750 mg/kg diet) on three-dimensional (3D) microcomputed tomography (μCT) structural parameters of fifth lumbar vertebrae.
| Parameters | Sham | Ovx | LD | MD | HD |
|---|---|---|---|---|---|
| BV/TV (%) | 0.382 ± 0.023a | 0.242 ± 0.023b | 0.208 ± 0.023b | 0.217 ± 0.023b | 0.230 ± 0.023b |
| Tb.N (1/mm) | 3.290 ± 0.150a | 2.920 ± 0.150ab | 2.660 ± 0.150b | 2.560 ± 0.150b | 2.900 ± 0.150ab |
| Tb.Th (mcm) | 0.109 ± 0.003a | 0.087 ± 0.003b | 0.084 ± 0.003b | 0.088 ± 0.003b | 0.084 ± 0.003b |
| Tb.Sp (mcm) | 0.295 ± 0.018a | 0.339 ± 0.018ab | 0.378 ± 0.018b | 0.387 ± 0.018b | 0.345 ± 0.018ab |
| Conn.D (1/mm3) | 25.8 ± 2.5a | 33.4 ± 2.5b | 29.5 ± 2.5ab | 25.3 ± 2.5a | 34.1 ± 2.5b |
| SMI | 1.158 ± 0.258a | 0.295 ± 0.258b | 0.640 ± 0.258b | 0.472 ± 0.258b | 0.427 ± 0.258b |
Values within a row that share the same superscript letters are not significantly (P ≥ .05) different. Values are means ± S.E.M.
BV/TV: bone volume as percentage of tissue volume; Tb.Th: trabecular thickness; Tb.N: trabecular number; Tb.Sp: trabecular separation; Conn.D: connective density; SMI: structure model index.
3.6. Bone Density and Biomechanical Properties
Ovariectomy significantly (P < 0.0001) decreased L4 vertebra BMD; however, there were no differences in BMD among the four Ovx groups due to vitamin E treatment (Table 2). Biomechanical properties of the vertebral body such as ultimate and yield load (Figure 1(a)) and ultimate and yield stress (Figure 1(b)) were increased in the group receiving the HD of vitamin E (P = 0.003, P = 0.009, P = 0.003, and P = 0.001, resp.). Ultimately, stiffness and elastic modulus were not affected (P = 0.18, P = 0.06, resp.) by any of the treatments.
Figure 1.
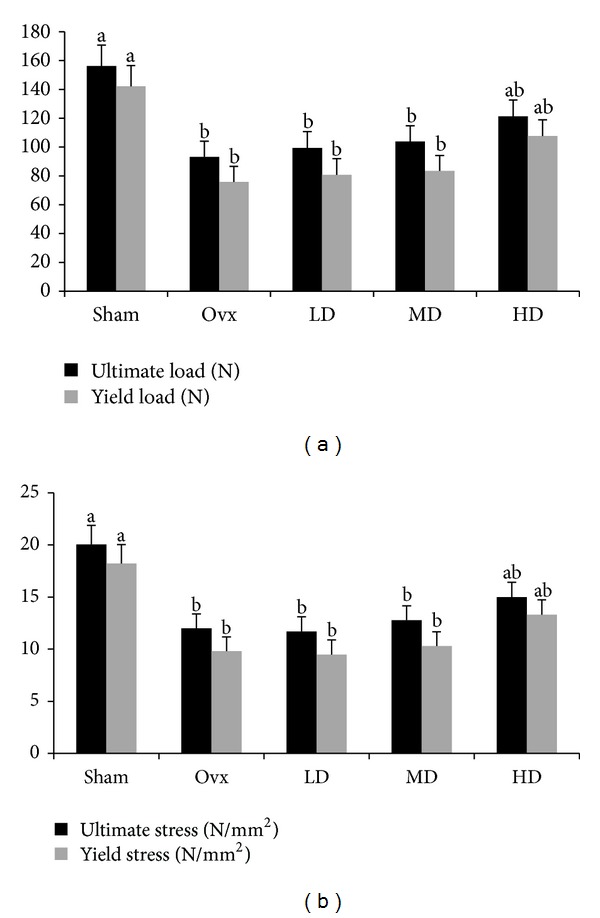
Effects of ovariectomy (Ovx) and three doses of vitamin E: low dose (LD, 300 mg/kg diet), medium dose (MD, 525 mg/kg diet), and high dose (HD, 750 mg/kg diet) on biomechanical parameters, (a) ultimate and yield load and (b) ultimate and yield stress of L4 vertebrae. Values are expressed as means ± S.E.M. Values that share the same superscript letters are not significantly (P ≥ .05) different; n = 12 rats/group.
4. Discussion
The overall findings of our previous studies with vitamin E using rat [46, 47] and mouse [45] models of osteoporosis indicate that vitamin E is able to improve the quality of bone, especially under high-stress conditions such as unloading and aging, while unable to prevent the loss of BMD. Although BMD may at best predict the incidence of fracture by approximately 70% [52–54], other factors such as bone geometry and bone quality are of equal importance. Our data suggest that improvements in bone structural properties may occur in the absence of favorable changes in BMD. Other examples of this phenomenon reported in the literature include the observation that sodium fluoride increases BMD by nearly 10% per year while at the same time increases the rate of fracture [55, 56]. In support of our earlier observations [45, 47, 57], the findings of this study also suggest that vitamin E improves bone biomechanical properties as indicated by higher ultimate and yield load and stress while not altering BMD. Similarly, Shuid et al. [58] have shown that supplementation with vitamin E enhances femoral strength as evidenced by enhanced ultimate and yield load and stress in young male rats. However, the question remains as to why vitamin E is incapable of increasing BMD values after bone loss has occurred. A reasonable explanation for these findings may be tied to the fact that by the end of 120-day pretreatment period, too little trabecular bone remained so that an increase in bone formation induced by vitamin E, due to upregulation of osteoblasts and suppression of bone erosion by osteoclast, was unable to restore the trabecular bone structure.
When the amount of vitamin E in the diet increased from 75 mg/kg diet (control diet) to 300, we observed a significant increase in the mean serum levels of vitamin E. However, when doses of vitamin E were further increased, the effect on circulating vitamin E became indistinguishable which indicates that vitamin E level in the diet may have reached its threshold in terms of absorption or transport. This has been observed even in healthy individuals as reported by Traber and Sies [59] who stated that the efficiency of vitamin E absorption is low and the uptake decreases as the dose increases. In fact, there are studies reporting that increasing doses of vitamin E supplementation (up to 800 mg) fail to raise plasma levels of vitamin E [60, 61]. Despite this lack of an effect on circulating vitamin E, we did see differences in the response in bone. This suggests that tissue levels of vitamin E might have been altered indicating that circulating levels of vitamin E may not reflect tissue vitamin E levels [62]. Furthermore, vitamin E was able to dose-dependently increase serum ALP concentrations, a nonspecific biomarker of bone formation expressed in the osteoblastic plasma membrane. These results are supported by the findings of our previous study in which vitamin E tended to increase the levels of serum ALP [47].
In terms of bone resorption, static histomorphometry data of proximal tibiae indicate that supplementation with vitamin E reduces osteoclast activity as indicated by decreases in Oc.S/BS and ES/BS in the vitamin E-supplemented groups when compared to Ovx-control group. Thus, these findings suggest that vitamin E supplementation decreases bone resorption in the Ovx rat model. A recent report by Fujita et al. [63] suggests that vitamin E reduces bone mass in nonstressed wildtype mice or rats that had their diets supplemented with α-tocopherol by enhancing bone resorption and osteoclast size as seen in wildtype bone-marrow cells stimulated with receptor activator of nuclear factor kappa-B and treated with α-tocopherol; however, our findings are in agreement with the findings of several other studies which report the beneficial effects of vitamin E on bone. For instance, Hermizi et al. [64] have demonstrated that supplementation with vitamin E lowered Oc.S/BS and ES/BS in male rats after receiving nicotine in comparison to control group. Additionally, Ahmad et al. [65] reported that vitamin E is also able to decrease ES/BS in male rats receiving ferric nitriloacetate. Based on the aforementioned findings, one could speculate that vitamin E supplementation is only beneficial under stress conditions. However, Mehat et al. [66] have shown that supplementation with vitamin E reduces ES/BS in young male rats when compared to control group suggesting that vitamin E is also beneficial under normal conditions.
Rates of bone turnover were evaluated using dynamic histomorphometry of the proximal tibiae. MS/BS was higher in the groups supplemented with vitamin E than in the Ovx control group. In addition, supplementation with the MD of vitamin E increased bone formation (BFR/BS) by 65% when compared to Ovx control group. These results indicate that supplementation with vitamin E increases mineralization and bone formation in the Ovx rat model. These findings are supported by Mehat et al. [66] that have found higher MS/BS and BFR/BS in young male rats following supplementation with vitamin E when compared to control group.
Supplementation with vitamin E was not able to reverse the unfavorable alterations of microarchitectural parameters in the L5 vertebrae due to ovariectomy. These findings were similar to our previous findings [46] in which vitamin E did not alter BV/TV, Tb.Th, and Tb.N in Orx male rats; however, it slightly differs from one of our earlier findings [47] in which BV/TV as well as Tb.N was increased by adequate and high doses of vitamin E in hindlimb unloaded male rats when interactions between diet and unloading were evaluated. The slight difference in the findings from our laboratory may be explained by the use of histomorphometry in the latter study versus μCT in the former and present study; yet, it is important to note that in this study, histomorphometry as well as μCT data are in agreement. Contrary to our findings, Shuid et al. [58] have shown that supplementation with vitamin E significantly increases BV/TV, Tb.Th, and Tb.N and decreases Tb.Sp. Since supplemental vitamin E bioavailability is greatly dependent on the form it is presented to the intestine absorptive cells, this difference may be, in part, attributed to the distinct methods of supplementation utilized in each study. While Shuid et al. [58] administered 60 mg/kg body weight/day by oral gavage, we fed the animals a diet containing 300 (LD), 525 (MD), and 750 (HD) mg of vitamin E/kg diet. In addition, Shuid et al. [58] were investigating the ability of vitamin E prevent bone loss using normal male rats whereas our study was examining the role of vitamin E in reversing bone loss due to ovarian hormone deficiency using Ovx rats.
One of the mechanisms by which vitamin E affects bone health is through its anti-inflammatory and antioxidative properties. The role of vitamin E in these processes has been clearly identified and is significant in the prevention of chronic diseases including atherosclerosis [67, 68] and cancer [69]. Macrophages are involved not only in the initiation but also in the regulation of inflammatory and immune responses [18, 70–72]. The increased secretion of proinflammatory molecules from macrophages such as PGE2, IL-6, TNF-α [73], and IL-1 [9] may lead to an increase in the rate of bone resorption, hence, accelerating bone loss. For instance, vitamin E did not suppress the synthesis of PGE2 and IL-6 in lipopolysaccharide-stimulated macrophages [74]. These findings suggest that vitamin E may act differently under various stress conditions. In terms of bone, this may partially explain the contrasting findings in Ovx and Orx rats compared to hindlimb unloaded rats. Although a few studies have reported that vitamin E positively influences bone quality [47, 58, 64], these studies examined the role of vitamin E in preventing bone loss but not its reversal. In conclusion, our observations as well as those of other investigators suggest that vitamin E supplementation is able to prevent bone loss and improve bone biomechanical properties even though it is unable to reverse the loss of bone due to ovarian hormone deficiency.
Acknowledgments
This work was funded by the United States Department of Agriculture (USDA) NRI grant. Vitamin E was generously provided by Archer Daniels Midland Company.
References
- 1.Dixon JM. Hormone replacement therapy and the breast. British Medical Journal. 2001;323(7326):1381–1382. doi: 10.1136/bmj.323.7326.1381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Marcus R. Post-menopausal osteoporosis. Best Practice and Research. 2002;16(3):309–327. doi: 10.1053/beog.2002.0284. [DOI] [PubMed] [Google Scholar]
- 3.Barnes PM, Powell-Griner E, McFann K, Nahin RL. Complementary and alternative medicine use among adults: United States, 2002. Advance Data. 2004;(343):1–19. [PubMed] [Google Scholar]
- 4.Stepan JJ, Pospichal J, Presl J, Pacovsky V. Bone loss and biochemical indices of bone remodeling in surgically induced postmenopausal women. Bone. 1987;8(5):279–284. doi: 10.1016/8756-3282(87)90002-0. [DOI] [PubMed] [Google Scholar]
- 5.Raisz LG. Bone cell biology: new approaches and unanswered questions. Journal of Bone and Mineral Research. 1993;8(supplement 2):S457–S465. doi: 10.1002/jbmr.5650081306. [DOI] [PubMed] [Google Scholar]
- 6.Tam VK, Schotland S, Green J. Inflammatory cytokines (IL-1α, TNF-α) and LPS modulate the Ca2+ signaling pathway in osteoblasts. American Journal of Physiology—Cell Physiology. 1998;274(6):C1686–C1698. doi: 10.1152/ajpcell.1998.274.6.C1686. [DOI] [PubMed] [Google Scholar]
- 7.Traianedes K, Dallas MR, Garrett IR, Mundy GR, Bonewald LF. 5-lipoxygenase metabolites inhibit bone formation in vitro. Endocrinology. 1998;139(7):3178–3184. doi: 10.1210/endo.139.7.6115. [DOI] [PubMed] [Google Scholar]
- 8.Manolagas SC. Birth and death of bone cells: basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocrine Reviews. 2000;21(2):115–137. doi: 10.1210/edrv.21.2.0395. [DOI] [PubMed] [Google Scholar]
- 9.Cheleuitte D, Mizuno S, Glowacki J. In vitro secretion of cytokines by human bone marrow: effects of age and estrogen status. Journal of Clinical Endocrinology and Metabolism. 1998;83(6):2043–2051. doi: 10.1210/jcem.83.6.4848. [DOI] [PubMed] [Google Scholar]
- 10.Walston J, Arking DE, Fallin D, et al. IL-6 gene variation is not associated with increased serum levels of IL-6, muscle, weakness, or frailty in older women. Experimental Gerontology. 2005;40(4):344–352. doi: 10.1016/j.exger.2005.01.012. [DOI] [PubMed] [Google Scholar]
- 11.Dinarello CA. Interleukin 1 and interleukin 18 as mediators of inflammation and the aging process. The American Journal of Clinical Nutrition. 2006;83(2):447s–455s. doi: 10.1093/ajcn/83.2.447S. [DOI] [PubMed] [Google Scholar]
- 12.Bartocci A, Papademetriou V, Schlick E, Nisula BC, Chirigos MA. Effect of crude and purified human chorionic gonadotropin on murine delayed-type hypersensitivity: a role for prostaglandins. Cellular Immunology. 1982;71(2):326–333. doi: 10.1016/0008-8749(82)90266-0. [DOI] [PubMed] [Google Scholar]
- 13.Meydani SN, Meydani M, Verdon CP, Shapiro AA, Blumberg JB, Hayes KC. Vitamin E supplementation suppresses prostaglandin E1(2) synthesis and enhances the immune response of aged mice. Mechanisms of Ageing and Development. 1986;34(2):191–201. doi: 10.1016/0047-6374(86)90034-5. [DOI] [PubMed] [Google Scholar]
- 14.Meydani SN, Barklund MP, Liu S, et al. Vitamin E supplementation enhances cell-mediated immunity in healthy elderly subjects. The American Journal of Clinical Nutrition. 1990;52(3):557–563. doi: 10.1093/ajcn/52.3.557. [DOI] [PubMed] [Google Scholar]
- 15.Banfi G, Iorio EL, Corsi MM. Oxidative stress, free radicals and bone remodeling. Clinical Chemistry and Laboratory Medicine. 2008;46(11):1550–1555. doi: 10.1515/CCLM.2008.302. [DOI] [PubMed] [Google Scholar]
- 16.Hayek MG, Meydani SN, Blumberg JB. Age differences in eicosanoid production of mouse splenocytes: effects on mitogen-induced T-cell proliferation. Journals of Gerontology. 1994;49(5):B197–B207. doi: 10.1093/geronj/49.5.b197. [DOI] [PubMed] [Google Scholar]
- 17.Hayek MG, Mura C, Wu D, et al. Enhanced expression of inducible cyclooxygenase with age in murine macrophages. Journal of Immunology. 1997;159(5):2445–2451. [PubMed] [Google Scholar]
- 18.Wu D, Beharka AA, Han SN, et al. Age-associated increase in PGE2 synthesis and COX activity in murine macrophages is reversed by vitamin E. American Journal of Physiology—Cell Physiology. 1998;275(3):C661–C668. doi: 10.1152/ajpcell.1998.275.3.C661. [DOI] [PubMed] [Google Scholar]
- 19.Nathan CF. Secretory products of macrophages. The Journal of Clinical Investigation. 1987;79(2):319–326. doi: 10.1172/JCI112815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mundy GR. Cytokines and growth factors in the regulation of bone remodeling. Journal of Bone and Mineral Research. 1993;8(supplement 2):S505–S510. doi: 10.1002/jbmr.5650081315. [DOI] [PubMed] [Google Scholar]
- 21.Harman D. Aging and oxidative stress. The Journal of the International Federation of Clinical Chemistry. 1998;10(1):24–27. [PubMed] [Google Scholar]
- 22.Garrett IR, Boyce BF, Oreffo ROC, Bonewald L, Poser J, Mundy GR. Oxygen-derived free radicals stimulate osteoclastic bone resorption in rodent bone in vitro and in vivo. The Journal of Clinical Investigation. 1990;85(3):632–639. doi: 10.1172/JCI114485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.van’t Hof RJ, Ralston SH. Nitric oxide and bone. Immunology. 2001;103(3):255–261. doi: 10.1046/j.1365-2567.2001.01261.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gokturk E, Turgut A, Baycu C, Gunal I, Seber S, Gulbas Z. Oxygen-free radicals impair fracture healing in rats. Acta Orthopaedica Scandinavica. 1995;66(5):473–475. doi: 10.3109/17453679508995590. [DOI] [PubMed] [Google Scholar]
- 25.Basu S, Michaëlsson K, Olofsson H, Johansson S, Melhus H. Association between oxidative stress and bone mineral density. Biochemical and Biophysical Research Communications. 2001;288(1):275–279. doi: 10.1006/bbrc.2001.5747. [DOI] [PubMed] [Google Scholar]
- 26.Lean JM, Davies JT, Fuller K, et al. A crucial role for thiol antioxidants in estrogen-deficiency bone loss. The Journal of Clinical Investigation. 2003;112(6):915–923. doi: 10.1172/JCI18859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Muthusami S, Ramachandran I, Muthusamy B, et al. Ovariectomy induces oxidative stress and impairs bone antioxidant system in adult rats. Clinica Chimica Acta. 2005;360(1-2):81–86. doi: 10.1016/j.cccn.2005.04.014. [DOI] [PubMed] [Google Scholar]
- 28.Crary EJ, McCarty MF. Potential clinical applications for high-dose nutritional antioxidants. Medical Hypotheses. 1984;13(1):77–98. doi: 10.1016/0306-9877(84)90132-4. [DOI] [PubMed] [Google Scholar]
- 29.Azzi A, Breyer I, Feher M, et al. Specific cellular responses to α-tocopherol. Journal of Nutrition. 2000;130(7):1649–1652. doi: 10.1093/jn/130.7.1649. [DOI] [PubMed] [Google Scholar]
- 30.Wang Y, Huang DS, Liang B, Watson RR. Nutritional status and immune responses in mice with murine AIDS are normalized by vitamin E supplementation. Journal of Nutrition. 1994;124(10):2024–2032. doi: 10.1093/jn/124.10.2024. [DOI] [PubMed] [Google Scholar]
- 31.van Tits LJ, Demacker PN, de Graaf J, Hak-Lemmers HL, Stalenhoef AF. α-Tocopherol supplementation decreases production of superoxide and cytokines by leukocytes ex vivo in both normolipidemic and hypertriglyceridemic individuals. The American Journal of Clinical Nutrition. 2000;71(2):458–464. doi: 10.1093/ajcn/71.2.458. [DOI] [PubMed] [Google Scholar]
- 32.Jialal I, Devaraj S, Kaul N. The effect of α-Tocopherol on monocyte proatherogenic activity. Journal of Nutrition. 2001;131(2):389s–394s. doi: 10.1093/jn/131.2.389S. [DOI] [PubMed] [Google Scholar]
- 33.Wu DY, Hayek MG, Meydani SN. Vitamin E and macrophage cyclooxygenase regulation in the aged. Journal of Nutrition. 2001;131(2):382S–288S. doi: 10.1093/jn/131.2.382S. [DOI] [PubMed] [Google Scholar]
- 34.Asehnoune K, Strassheim D, Mitra S, Kim JY, Abraham E. Involvement of reactive oxygen species in toll-like receptor 4-dependent activation of NF-κB. Journal of Immunology. 2004;172(4):2522–2529. doi: 10.4049/jimmunol.172.4.2522. [DOI] [PubMed] [Google Scholar]
- 35.Manolagas SC, Jilka RL. Mechanisms of disease: bone marrow, cytokines, and bone remodeling—emerging insights into the pathophysiology of osteoporosis. The New England Journal of Medicine. 1995;332(5):305–311. doi: 10.1056/NEJM199502023320506. [DOI] [PubMed] [Google Scholar]
- 36.Ricciarelli R, Zingg J-M, Azzi A. Vitamin E: protective role of a Janus molecule. The FASEB Journal. 2001;15(13):2314–2325. doi: 10.1096/fj.01-0258rev. [DOI] [PubMed] [Google Scholar]
- 37.Xu H, Watkins BA, Seifert MF. Vitamin E stimulates trabecular bone formation and alters epiphyseal cartilage morphometry. Calcified Tissue International. 1995;57(4):293–300. doi: 10.1007/BF00298885. [DOI] [PubMed] [Google Scholar]
- 38.Durak K, Bilgen ÖF, Kaleli T, Tuncel P, Özbek R, Turan K. Antioxidant effect of α-tocopherol on fracture haematoma in rabbits. Journal of International Medical Research. 1996;24(5):419–424. doi: 10.1177/030006059602400503. [DOI] [PubMed] [Google Scholar]
- 39.Shuid AN, Mohamad S, Muhammad N, Afian Mokhtar S, Muhammad N, Nirwana Soelaiman I. Bone oxidative changes during early fracture healing of postmenopausal osteoporosis rat model. Asian Journal of Animal and Veterinary Advances. 2011;6(12):1193–1203. [Google Scholar]
- 40.Shuid AN, Mohamad S, Muhammad N, et al. Effects of α-tocopherol on the early phase of osteoporotic fracture healing. Journal of Orthopaedic Research. 2011;29(11):1732–1738. doi: 10.1002/jor.21452. [DOI] [PubMed] [Google Scholar]
- 41.Mohamad S, Shuid A, Mohamed N, Fadzilah F, Mokhtar S, Abdullah S, et al. The effects of alpha-tocopherol supplementation on fracture healing in a postmenopausal osteoporotic rat model. Clinics. 2012;67(9):1077–1085. doi: 10.6061/clinics/2012(09)16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Turk CY, Halici M, Guney A, Akgun H, Sahin V, Muhtaroglu S. Promotion of fracture healing by vitamin E in rats. Journal of International Medical Research. 2004;32(5):507–512. doi: 10.1177/147323000403200508. [DOI] [PubMed] [Google Scholar]
- 43.Melhus H, Michaëlsson K, Holmberg L, Wolk A, Ljunghall S. Smoking, antioxidant vitamins, and the risk of hip fracture. Journal of Bone and Mineral Research. 1999;14(1):129–135. doi: 10.1359/jbmr.1999.14.1.129. [DOI] [PubMed] [Google Scholar]
- 44.Hamidi MS, Corey PN, Cheung AM. Effects of vitamin E on bone turnover markers among US postmenopausal women. Journal of Bone and Mineral Research. 2012;27(6):1368–1380. doi: 10.1002/jbmr.1566. [DOI] [PubMed] [Google Scholar]
- 45.Arjmandi BH, Juma S, Beharka A, Bapna MS, Akhter M, Meydani SN. Vitamin E improves bone quality in the aged but not in young adult male mice. The Journal of Nutritional Biochemistry. 2002;13(9):543–549. doi: 10.1016/s0955-2863(02)00199-7. [DOI] [PubMed] [Google Scholar]
- 46.Chai SC, Wei C-I, Brummel-Smith K, Arjmandi BH. The role of vitamin E in reversing bone loss. Aging. 2008;20(6):521–527. doi: 10.1007/BF03324879. [DOI] [PubMed] [Google Scholar]
- 47.Smith BJ, Lucas EA, Turner RT, et al. Vitamin E provides protection for bone in mature hindlimb unloaded male rats. Calcified Tissue International. 2005;76(4):272–279. doi: 10.1007/s00223-004-0269-8. [DOI] [PubMed] [Google Scholar]
- 48.Norazlina M, Ima-Nirwana S, Gapor MT, Khalid BAK. Palm vitamin E is comparable to α-tocopherol in maintaining bone mineral density in ovariectomised female rats. Experimental and Clinical Endocrinology and Diabetes. 2000;108(4):305–310. doi: 10.1055/s-2000-7758. [DOI] [PubMed] [Google Scholar]
- 49.Arjmandi BH, Getlinger MJ, Goyal NV, et al. Role of soy protein with normal or reduced isoflavone content in reversing bone loss induced by ovarian hormone deficiency in rats. The American Journal of Clinical Nutrition. 1998;68(supplement 6):1358S–1363S. doi: 10.1093/ajcn/68.6.1358S. [DOI] [PubMed] [Google Scholar]
- 50.Bieri JG, Tolliver TJ, Catignani GL. Simultaneous determination of α-tocopherol and retinol in plasma or red cells by high pressure liquid chromatography. The American Journal of Clinical Nutrition. 1979;32(10):2143–2149. doi: 10.1093/ajcn/32.10.2143. [DOI] [PubMed] [Google Scholar]
- 51.Akhter MP, Iwaniec UT, Haynatzki GR, Fung YK, Cullen DM, Recker RR. Effects of nicotine on bone mass and strength in aged female rats. Journal of Orthopaedic Research. 2003;21(1):14–19. doi: 10.1016/S0736-0266(02)00107-9. [DOI] [PubMed] [Google Scholar]
- 52.Wasnich RD, Ross PD, Heilbrun LK, Vogel JM. Prediction of postmenopausal fracture risk with use of bone mineral measurements. The American Journal of Obstetrics and Gynecology. 1985;153(7):745–751. doi: 10.1016/0002-9378(85)90338-2. [DOI] [PubMed] [Google Scholar]
- 53.Kanis JA. Osteoporosis III: diagnosis of osteoporosis and assessment of fracture risk. The Lancet. 2002;359(9321):1929–1936. doi: 10.1016/S0140-6736(02)08761-5. [DOI] [PubMed] [Google Scholar]
- 54.Taaffe DR, Lang TF, Harris TB. Poor correlation of mid-femoral measurements by CT and hip measurements by DXA in the elderly. Aging. 2003;15(2):131–135. doi: 10.1007/BF03324490. [DOI] [PubMed] [Google Scholar]
- 55.Wardlaw GM. Putting osteoporosis in perspective. Journal of the American Dietetic Association. 1993;93(9):1000–1006. doi: 10.1016/0002-8223(93)92038-y. [DOI] [PubMed] [Google Scholar]
- 56.Delmas PD. Treatment of postmenopausal osteoporosis. The Lancet. 2002;359(9322):2018–2026. doi: 10.1016/S0140-6736(02)08827-X. [DOI] [PubMed] [Google Scholar]
- 57.Arjmandi BH, Akhter M, Chakkalakal D, et al. Effects of isoflavones, vitamin E, and their combination on bone in an aged rat model of osteopenia. Journal of Bone and Mineral Research. 2001;16( supplement 1):p. s533. [Google Scholar]
- 58.Shuid AN, Mehat Z, Mohamed N, Muhammad N, Soelaiman IN. Vitamin E exhibits bone anabolic actions in normal male rats. Journal of Bone and Mineral Metabolism. 2010;28(2):149–156. doi: 10.1007/s00774-009-0122-2. [DOI] [PubMed] [Google Scholar]
- 59.Traber MG, Sies H. Vitamin E in humans: demand and delivery. Annual Review of Nutrition. 1996;16:321–347. doi: 10.1146/annurev.nu.16.070196.001541. [DOI] [PubMed] [Google Scholar]
- 60.Schultz M, Leist M, Petrzika M, Gassmann B, Brigelius-Flohe R. Novel urinary metabolite of α-tocopherol, 2,5,7,8-tetramethyl-2(2′- carboxyethyl)-6-hydroxychroman, as an indicator of an adequate vitamin E supply? The American Journal of Clinical Nutrition. 1995;62(6):S1527–S1534. doi: 10.1093/ajcn/62.6.1527S. [DOI] [PubMed] [Google Scholar]
- 61.Jialal I, Fuller CJ, Huet BA. The effect of α-tocopherol supplementation on LDL oxidation. A dose-response study. Arteriosclerosis, Thrombosis, and Vascular Biology. 1995;15(2):190–198. doi: 10.1161/01.atv.15.2.190. [DOI] [PubMed] [Google Scholar]
- 62.Blatt DH, Leonard SW, Traber MG. Vitamin E kinetics and the function of tocopherol regulatory proteins. Nutrition. 2001;17(10):799–805. doi: 10.1016/s0899-9007(01)00637-2. [DOI] [PubMed] [Google Scholar]
- 63.Fujita K, Iwasaki M, Ochi H, et al. Vitamin E decreases bone mass by stimulating osteoclast fusion. Nature Medicine. 2012;18(4):589–594. doi: 10.1038/nm.2659. [DOI] [PubMed] [Google Scholar]
- 64.Hermizi H, Faizah O, Ima-Nirwana S, Ahmad Nazrun S, Norazlina M. Beneficial effects of tocotrienol and tocopherol on bone histomorphometric parameters in Sprague-Dawley male rats after nicotine cessation. Calcified Tissue International. 2009;84(1):65–74. doi: 10.1007/s00223-008-9190-x. [DOI] [PubMed] [Google Scholar]
- 65.Ahmad NS, Khalid BAK, Luke DA, Nirwana SI. Tocotrienol offers better protection than tocopherol from free radical-induced damage of rat bone. Clinical and Experimental Pharmacology and Physiology. 2005;32(9):761–770. doi: 10.1111/j.1440-1681.2005.04264.x. [DOI] [PubMed] [Google Scholar]
- 66.Mehat MZ, Shuid AN, Mohamed N, Muhammad N, Soelaiman IN. Beneficial effects of vitamin E isomer supplementation on static and dynamic bone histomorphometry parameters in normal male rats. Journal of Bone and Mineral Metabolism. 2010;28(5):503–509. doi: 10.1007/s00774-010-0159-2. [DOI] [PubMed] [Google Scholar]
- 67.Meydani M. Vitamin E and atherosclerosis: beyond prevention of LDL oxidation. Journal of Nutrition. 2001;131(2):366S–368S. doi: 10.1093/jn/131.2.366S. [DOI] [PubMed] [Google Scholar]
- 68.Chai SC, Arjmandi BH. Vitamin E dose-dependently reduces aortic fatty lesion formation in orchidectomized aged rats. Aging. 2011;23(1):11–16. doi: 10.1007/BF03337742. [DOI] [PubMed] [Google Scholar]
- 69.Dutta A, Dutta SK. Vitamin E and its role in the prevention of atherosclerosis and carcinogenesis: a review. Journal of the American College of Nutrition. 2003;22(4):258–268. doi: 10.1080/07315724.2003.10719302. [DOI] [PubMed] [Google Scholar]
- 70.Wu D, Hayek MG, Meydani S. Vitamin E and macrophage cyclooxygenase regulation in the aged. Journal of Nutrition. 2001;131(2):382S–388S. doi: 10.1093/jn/131.2.382S. [DOI] [PubMed] [Google Scholar]
- 71.Beharka AA, Wu D, Han SN, Meydani SN. Macrophage prostaglandin production contributes to the age-associated decrease in T cell function which is reversed by the dietary antioxidant vitamin E. Mechanisms of Ageing and Development. 1997;93(1–3):59–77. doi: 10.1016/s0047-6374(96)01819-2. [DOI] [PubMed] [Google Scholar]
- 72.Wu DY, Meydani M, Beharka AA, Serafini M, Martin KR, Meydani SN. In vitro supplementation with different tocopherol homologues can affect the function of immune cells in old mice. Free Radical Biology and Medicine. 2000;28(4):643–651. doi: 10.1016/s0891-5849(99)00276-2. [DOI] [PubMed] [Google Scholar]
- 73.Pfeilschifter J, Köditz R, Pfohl M, Schatz H. Changes in proinflammatory cytokine activity after menopause. Endocrine Reviews. 2002;23(1):90–119. doi: 10.1210/edrv.23.1.0456. [DOI] [PubMed] [Google Scholar]
- 74.Beharka AA, Han SN, Adolfsson O, et al. Long-term dietary antioxidant supplementation reduces production of selected inflammatory mediators by murine macrophages. Nutrition Research. 2000;20(2):281–296. [Google Scholar]