

Abstract
The increased morbidity, mortality, and ineffective treatment associated with the pathogenesis of chronic inflammatory diseases such as asthma and chronic obstructive pulmonary disease (COPD) have generated much research interest. The key role is played by phospholipases from the A2 superfamily: enzymes which are involved in inflammation through participation in pro- and anti-inflammatory mediators production and have an impact on many immunocompetent cells. The 30 members of the A2 superfamily are divided into 7 groups. Their role in asthma and COPD has been studied in vitro and in vivo (animal models, cell cultures, and patients). This paper contains complete and updated information about the involvement of particular enzymes in the etiology and course of asthma and COPD.
1. Introduction
Both asthma and COPD are airway diseases characterized by impaired airflow in the respiratory tract, chronic airway inflammation, as well as symptoms such as coughing, dyspnea, and wheezing. Intensive studies focused on the pathogenesis of these conditions implicate, among others, the group of phospholipases A2, which possess enzymatic and nonenzymatic properties. This paper presents general information about phospholipases and details the current knowledge about particular phospholipases A2 involved in asthma and COPD in human and animal models. The data regarding interactions between members of this superfamily is summarized, as well as the role of these enzymes in exacerbations of inflammatory diseases.
2. Phospholipases
Phospholipases are enzymes that hydrolyze phospholipids. The main substrates for these enzymes are glycerophospholipids which contain glycerol with a saturated fatty acid in the sn-1 position and an unsaturated fatty acid in the sn-2 position. The phospholipases responsible for hydrolysis of glycerophospholipids are divided into two groups: acylhydrolases and phosphodiesterases. The first group comprises phospholipase A1 (PLA1) and A2 (PLA2), which hydrolyze the ester bond at the sn-1 and sn-2 positions, respectively. The second group comprises phospholipase C (PLC) which cleaves the glycerol-phosphate bond, and phospholipase D (PLD), which liberates phosphatidic acid and alcohol (Figure 1). Phospholipase B shares both the properties of PLA1 and PLA2.
Figure 1.
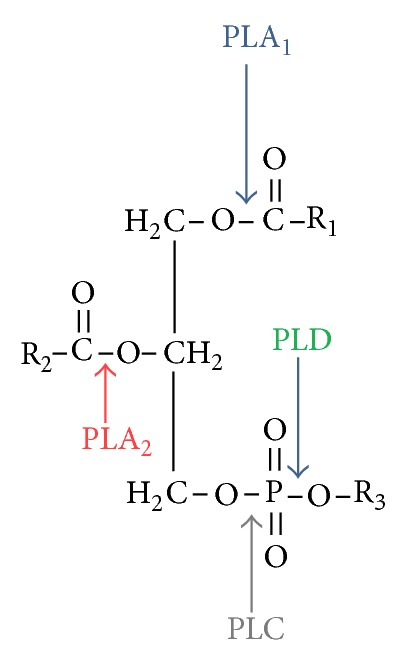
Phospholipases and their role in lipids metabolism.
The structure, function, and catalytic mechanism of the enzyme determine its place within the phospholipase A2 superfamily, be it secretory PLA2 (sPLA2), cytosolic PLA2 (cPLA2), Ca2+-independent phospholipase A2 (iPLA2), PAF acetylhydrolases (PAF-AH), or lysosomal PLA2 (LPLA2). The latest classification, based on genetic structure, divides these enzymes into groups from I to XVI (in each one, the enzyme is represented by a capital letter) [1]. The characteristic features of each group are presented in Table 1. Table 2 includes information about the mechanism of action and function of particular subgroups of PLA2s concerning physiology and pathophysiology.
Table 1.
Characteristics of structure and localization of human phospholipase A2 enzymes. Adapted and modified from [1, 4]. The Roman numeral indicates the group, and the capital letter after the number indicates the subgroup.
| Name | Members (human) | Molecular mass (kDa) | Relationship with Ca2+ | Catalytic site | Localization |
|---|---|---|---|---|---|
|
Secretory phospholipase A2 (sPLA2) |
IB (sPLA2IB) | 13–15 | Secreted | ||
| IIA (sPLA2IIA) | 13–15 | Secreted; membrane; secretory granules | |||
| IID (sPLA2IID) | 14-15 | Secreted | |||
| IIE (sPLA2IIE) | 14-15 | Secreted | |||
| IIF (sPLA2IIF) | 16-17 | Secreted | |||
| III (sPLA2III) | 55 | Dependent | Histidine/Aspartic acid | Secreted | |
| V (sPLA2V) | 14 | Secreted; Golgi apparatus; nuclear envelope; plasma membrane | |||
| X (sPLA2X) | 14 | Secreted | |||
| XIIA (sPLA2XIIA) | 19 | Secreted; cytoplasm | |||
| XIIB (sPLA2XIIB, XIII) | 20 | Secreted | |||
|
| |||||
| IVA (cPLA2 α) | 85 | Dependent | Nucleus; cytoplasmic vesicles | ||
| IVB (cPLA2 β)-three splice variants | 114 | Cytosol | |||
| Cytosolic phospholipase A2 (cPLA2) | IVC (cPLA2 γ) | 61 | Independent | ER; Mitochondrium | |
| IVD (cPLA2 δ) | 92-93 | Serine/Aspartic acid/Arginine | Cytosol; Cytoplasmic vesicle membrane; peripheral membrane protein; cytoplasmic side | ||
| IVE (cPLA2 ε) | 96 | Dependent | Cytosol; lysosome membrane; peripheral membrane protein | ||
| IVF (cPLA2 ζ) | 95 | Cytosol; lysosome membrane; peripheral membrane protein; cytoplasmic side | |||
|
| |||||
| Ca2+-independent phospholipase A2 (iPLA2) | VIA-(iPLA2 β)-five splice variants | 84–90 | Cytosol | ||
| VIB (iPLA2 γ)-four splice variants | 88–91 | ER; peroxisomal and mitochondrial membrane | |||
| VIC (iPLA2 δ, NTE) | 146 | Independent | Serine | ER; single-pass type I membrane protein; cytoplasmic side | |
| VID (iPLA2 ε, adiponutrin) | 53 | Membrane; single-pass type II membrane protein | |||
| VIE (iPLA2 ζ) | 57 | Lipid droplet membrane; single-pass type II membrane protein; cell membrane | |||
| VIF (iPLA2 η) | 28 | Cytoplasm | |||
|
| |||||
| Acidic Ca2+-independent phospholipase A2 | aiPLA2 | 26 | Independent | Serine | Cytoplasm; Lysosome |
|
| |||||
| Lysosomal phospholipase A2 | XV (LPLA2, LLPL, ACS) | 45 | Independent | Serine/Histidine/Aspartic acid | Secreted; Lysosome |
|
| |||||
| PAF acetylhydrolase (PAF-AH) or Lipoprotein-associated phospholipase A2 | VIIA (Lp-PLA2, Plasma PAF-AH) | 45 | Independent | Serine/Histidine/Aspartic acid | Secreted |
| VIIB (PAF-AH II) | 40 | Cytoplasm | |||
| VIIIA (PAF-AH Ib) α1 subunit | 26 | Cytoplasm | |||
| VIIIB (PAF-AH Ib) α2 subunit | 26 | Cytoplasm | |||
|
| |||||
| Adipose-specific phospholipase A2 | XVI (H-Rev107) | 18 | Independent | Cystein/Histidine/Histidine | Cytoplasm, perinuclear region, Single-pass membrane protein |
ER: endoplasmic reticulum; NTE: neuropathy target esterase.
Table 2.
Mechanism of action and function of human phospholipase A2 enzymes. Adapted and modified from [1, 4, 5].
| Name | Mechanism of action | Function | Sources | |
|---|---|---|---|---|
| Physiology | Pathophysiology | |||
| Secretory phospholipases A2 (sPLA2s) | (i) Enzymatic (liberation of AA and lysophospholipids) (ii) Autocrine and paracrine action by binding to N-type and M-type receptors or by binding to integrins |
(i) Lipid remodeling for membrane homeostasis (ii) Exocytosis (iii) Phagocytosis (iv) Anticoagulant activity (v) Antibacterial activity (Gram-positive and Gram-negative bacteria) (vi) Antifungal and antiadenoviral activity (vii) Parturition (viii) Spinal processing of nociception |
(i) Inflammatory diseases (rheumatoid arthritis, adult respiratory distress syndrome, inflammatory bowel disease, and pancreatitis) (ii) Sepsis (iii) Atherosclerosis (foam cell formation) (iv) Cancer (v) Surfactant hydrolysis |
Neutrophils, eosinophils, basophils, T-cells, monocytes, macrophages, platelets, mast cells, airway epithelial cells, alveolar type II epithelial cells, |
|
| ||||
| Cytosolic phospholipases A2 (cPLA2s) | (i) enzymatic: lysophospholipase and transacylase activity | (i) AA releasing (ii) Cellular signaling (iii) Parturition (iv) Nociception |
(i) Inflammation (ii) Intestinal ulceration (iii) Psoriasis (iv) Acute lung injury (v) Polyposis (vi) Brain injury (vii) Anaphylaxis |
Every tissue |
|
| ||||
| Ca2+-independent phospholipases A2 (iPLA2s) |
VIA, VIB, VIC, VID, VIEVIF-phospholipase A2 activity VIC-lysophospholipase activity VID-adiponutrin-like activity VIE-triglyceride lipase activity VIF-transacylase activity |
(i) Remodeling of phospholipids (ii) AA releasing (iii) Protein expression (iv) Acetylcholine-mediated endothelium-dependent relaxation of the vasculature (v) Apoptosis (vi) Insulin secretion (vii) Bone formation (viii) Sperm development (ix) Cell proliferation (x) Activation of Ca2+ influx (xi) Axon regeneration in nerve injury (VIA) |
(i) Wallerian degeneration (VIA) (ii) regulation of monocyte migration (VIB) (iii) Oxidant-induced cell injury (VIC) (iv) Ischemia-induced ventricular tachyarrhythmias |
(i) Alveolar cells (ii) Macrophages (iii) Normal and cancer lung tissue (iv) Neurons |
| aiPLA2-phospholipase A2 and peroxiredoxin 6 activity | (i) Degradation and recycling of surfactant phospholipids (remodeling of phosphatidylcholine to dipalmitoyl-phosphatidylcholine (DPPC) (ii) Antioxidative activity |
(i) lung cancer, mesothelioma, sarcoidosis | (i) Alveolar macrophages (ii) Type II epithelial cells (iii) Clara cells |
|
|
| ||||
| Lysosomal phospholipase A2 | (i) Acyl-ceramide synthase (ii) Transacylase activity (iii) Lysophospholipase activity |
(i) may be the crucial enzyme of pulmonary surfactant phospholipid degradation by alveolar macrophages | (i) Phospholipidosis (ii) Complement activation (iii) Induced lung injury |
(i) Alveolar macrophages (ii) Peripheral blood monocytes |
|
| ||||
| PAF acetylhydrolases (PAF-AH) or Lipoprotein-associated phospholipases A2 | (i) Phospholipase A2 activity | (i) Anti-inflammatory properties by hydrolyzing platelet activating factor (PAF) (ii) Protection against oxidative stress (iii) Brain development |
(i) Generation of lysophospholipids and fatty acid hydroperoxides (ROS) (ii) Acute respiratory distress syndrome (iii) Marker of coronary heart disease (iv) Miller-Diker lissencephaly |
(i) Alveolar macrophages (ii) Epithelial type II cells |
|
| ||||
| Adipose-specific phospholipase A2 | (i) Phospholipase A1 and A2 activity | (i) catalyzes the release of fatty acids from phospholipids in adipose tissue | (i) Obesity (ii) Metabolic syndrome |
Adipose tissue |
3. Asthma and COPD
Currently about 300 million people worldwide suffer from asthma, and in 2025, this number is expected to grow by another 100 million. Annually, about 250 000 people die from asthma [2]. Asthma is defined according to the GINA (Global Initiative for Asthma) [3] as a chronic airway inflammatory disease in which many cells and cellular elements are involved. Chronic inflammation is a cause of bronchial hyperresponsiveness, leading to recurrent episodes of wheezing, dyspnea, chest tightness, and coughing, occurring particularly at night or dawn. This is usually accompanied by episodes of diffuse bronchial obstruction of varying severity, which often subside spontaneously or with treatment.
According to GOLD (The Global Initiative for Chronic Obstructive Lung Diseases) [6], COPD is characterized by a progressive and poorly reversible airflow limitation caused by both small airway diseases (airway inflammation and destruction) and parenchymal destruction (loss of alveolar attachment and decrease of elastic recall). Also, other extrapulmonary effects, such as weight loss, nutritional abnormalities, skeletal muscle dysfunction influence the severity of the disease. Apart from the genetic background (hereditary alpha-1 antitrypsin deficiency) [7] cigarette smoke is a crucial environmental factor in COPD development [8]; it is responsible for airway inflammation and further oxidant/antioxidant imbalance (oxidative stress) causing amplification of lung inflammation.
4. Analysis of Phospholipases A2 Involvement in Asthma and COPD
An analysis of studies concerning the profile of PLA2s expression in many experimental systems has revealed ambiguous results. Many different inductors used for cells stimulation cause expression of various types of enzymes in the same cells. Also, the presence of heterogeneous cells in experimental systems influences the expression of PLA2s [9].
Mast cells, Th2 lymphocytes, and eosinophils are the most important cellular components of asthma. It has been established that primary human lung mast cells constitutively express mRNA for the IB, IIA, IID, IIE, IIF, III, V, X, XIIA, and XIIB sPLA2 groups and stimulation with anti-IgE antibodies can induce their secretion [10]. Hence sPLA2 proteins are believed to belong to preformed mediators which are stored in mast cells granules. Cells stimulation by anti-IgE antibodies causes degranulation of mast cells, and sPLA2 appears in the early phase of allergic reaction. Muñoz et al. have shown that sPLA2V is not expressed in eosinophils in detectable amounts. However exogenous hPLA2V can activate eosinophils, inducing the liberation of arachidonic acid (AA) and LTC4 production [11]. Increased cPLA2 α phosphorylation and cPLA2 α activity was observed in eosinophils of asthmatics after allergen challenge [12].
Alveolar macrophages and neutrophils play a crucial role in the pathophysiology of COPD [13, 14]. Human macrophages express cPLA2IVA, iPLA2VIA, and several sPLA2s (IIA, IID, IIE, IIF, V, X, and XIIA, but not group IB and III enzymes). Higher expression of sPLA2IIA is observed after LPS treatment [15]. Neutrophils stimulated in vitro by the tripeptide formyl-Met-Leu-Phe (fMLP) demonstrate mRNA and protein expression of sPLA2V and sPLA2X, where the sPLA2V protein is found in azurophilic and specific granules, and sPLA2X is found only in azurophilic granules. GIB, GIIA, GIID, GIIE, GIIF, GIII, and GXII sPLA2s are undetectable. Cell activation by fMLP or zymosan results in the release of GV but not GX sPLA2 [16].
The BALF of patients with COPD demonstrates a three- to fivefold higher activity of PLA2s in comparison to a control BALF but the protein level shows no difference [17]. No differences in sPLA2IIs serum levels exist between healthy smokers and nonsmokers. However, significantly greater levels of this enzyme are found in the BALF of smokers compared with nonsmokers [18]. Among sPLA2s, sPLA2IID is also considered as a molecule involved in the course of COPD. A change of Gly80Ser in the sPLA2IID protein may be associated with body weight loss in patients suffering from COPD [19]. sPLA2IID can be also involved in control of inflammation by inhibition of CD4+, CD8+ T cells proliferation and induction of regulatory T cell differentiation [20]. Cigarette smoke extract (CSE) can induce the production of cytosolic phospholipase A2 in human pulmonary microvascular endothelial cells [21]. Moreover oxidative stress can increase the activity of cPLA2 by promoting its phosphorylation [22]. cPLA2 also participates in phosphodiesterase 4 signaling, whose inhibition attenuates neutrophilic inflammation in COPD [23]. The increased values of PLA2VII in patients with long-standing pulmonary hypertension (severe complication in COPD) are related to severe endothelial dysfunction [24].
sPLA2V plays a different role in the activation of eosinophils and neutrophils. Hence, its involvement in the pathogenesis of asthma and COPD can vary. Exogenous sPLA2V can activate the production of AA and leukotrienes in both cell types. However, LTB4 is preferentially produced in neutrophils, and LTC4 in eosinophils [11]. The sPLA2V-induced activation of neutrophils in contrast to eosinophils requires the presence and activation of cPLA2 [25]. The inhibition of cPLA2 may be more effective in diseases where neutrophils play a crucial role because they indirectly inhibit also the function of sPLA2.
5. Role of PLA2s in Asthma and COPD
The proposed mechanism of action of phospholipases A2 (PLA2s) in inflammatory diseases includes the liberation of arachidonic acid, generation of lysophospholipids, interaction between enzymes belonging to the A2 superfamily, surfactant degradation, release of cytokines, and the impact on immunological and inflammatory cells (dendritic cells, T-cells, and leukocytes) [26].
5.1. The Enzymatic Activity of PLA2s
The enzymatic properties of PLA2s refer to their phospholipase, lysophospholipase, transacylase, adiponutrin-like, triglyceride lipase, peroxiredoxin 6, and acyl-ceramide synthase activities. Phospholipases A2 play a pivotal role in eicosanoid production because they hydrolyze the ester bond at the sn-2 position of the glycerophospholipid membrane, releasing arachidonic acid (AA) and lysophospholipids [27]. Arachidonic acid plays a dual role. It can act as a signaling molecule that regulates the activity of protein kinase C (PKC) and phospholipase Cγ, influences Ca2+ concentration, and acts as an endogenous ligand for PPARγ receptors [28, 29]. AA is also a precursor of lipid inflammatory mediators (eicosanoids). In cyclooxygenase (COX) pathways, it is transformed to prostaglandins and thromboxane while in lipoxygenase (ALOX) pathways, it is converted to leukotrienes. These molecules are responsible for bronchial constriction, increased vessel permeability, and inflammatory cell recruitment [30]. AA is also a substrate for resolvins and lipoxins (LXs) which have anti-inflammatory properties. Lipoxins can block granulocyte chemotaxis, migration, degranulation, oxidative burst, cytokine-mediated signaling in eosinophils, and secretion of cytokines from bronchial epithelial cells [31]. Several independent studies have reported that significantly lower levels of LXs are observed in severe asthmatics compared to patients with nonsevere asthma [32, 33]. Resolvins demonstrate endogenous anti-inflammatory, proresolving, antifibrotic, antiangiogenic, anti-infective, and antihyperalgesic activity [31].
Among cytosolic phospholipases A2, it has been well documented that cPLA2IVA (cPLA2 α) plays an important role in eicosanoid production. In patients with inherited cPLA2 deficiency (loss-of-function mutations in both cPLA2 alleles), a widespread decrease in eicosanoid concentrations has been observed [34]. S111P, R485H, and K651R mutations in PLA2G4A gene are thought to play a crucial role in this condition. The functional consequences of localized mutations concerning cPLA2 catalytic activity, Ca2+ recruitment, and affinity for the phospholipid membrane have been confirmed in vitro and in cell culture [35]. In patients with severe asthma, the microsatellite fragments (T)n and (CA)n in the promoter region of cPLA2 α gene (PLA2G4A) are shorter in comparison to healthy subjects [36]. In addition, asthmatic patients with shorter microsatellite sequences demonstrate greater expression of cPLA2 α mRNA, cPLA2 α protein, PGE2 and 15-HETE, but not LTC4 [37]. cPLA2 participates in intracellular signaling, leading to allergen-induced production of inflammatory cytokines in the PBMC of asthmatics [38]. Hallstrand et al. [39] identified increased expression of three cPLA2s, including cPLA2 α, cPLA2 β, and cPLA2 γ in induced sputum cells from subjects with asthma and exercise-induced bronchoconstriction. Both cPLA2 β and cPLA2 γ enzymes also participate in eicosanoids biosynthesis [40, 41]. Increased cPLA2 expression and subsequent PGE2 production are present in the asthma phenotype. The therapeutic decision to inhibit cPLA2 in asthmatics may be unclear when considering the role of PGE2 in airway inflammation. There is some evidence that PGE2 can act as bronchodilator, as well as an inhibitor of both allergen-induced bronchoconstriction and inflammatory mediators production [42]. It should be noticed that PGE2 acts through four different types of receptors (EP1, EP2, EP3, and EP4). Changes in expression and combination of receptor subtypes actions may affect the action of PGE2 giving it proinflammatory or bronchoprotective outcomes [43–45]. The pleiotropic properties of PGE2 make it difficult to establish the direct impact of PGE2 deficiency which appears as a consequence of cPLA2 inhibition [46]. Moreover, although cPLA2 is a major enzyme, it is not the only one providing substrates for eicosanoids synthesis; hence it cannot be excluded that other existing pathways can also perform this function.
sPLA2s and arachidonic acid accumulate in the BALF of asthmatics after allergen challenge [47, 48]. Despite being specific to the sn-2 bond, sPLA2s play more of a supporting role in AA liberation. Only sPLA2V and sPLA2X can efficiently interact and hydrolyze phospholipids from the outer surface of the cell membrane [9]. In acute and chronic animal asthma models, a deficit of sPLA2X diminishes the features of asthma (eosinophilia, airway hyperresponsiveness to methacholine, airway remodeling, eicosanoids, and Th2 cytokine production) [49].
Hallstrand et al. [50] showed that the expression of sPLA2X predominates in the airway epithelium, and both sPLA2X and sPLA2IIA are the main phospholipases produced by BALF cells. The activity of the sPLA2V protein was found to be greatly lowered and undetectable. They have suggested that sPLA2X is most important among secretory phospholipases. Only sPLA2X, not sPLA2IIA, is correlated with asthma features such as lung function, recruitment of neutrophils in asthmatics [50]. sPLA2X is responsible for production of cysteinyl leukotrienes (cysLTs) which are proinflammatory in asthma and can be responsible for observable features of asthma. Moreover, the level of prostaglandin E2 (PGE2) is also connected with sPLA2X, which can be explained by the fact that sPLA2X increases activity of cPLA2IV which in turn leads to production of PGE2. These results are consistent with earlier studies by the same authors in which gene expression of sPLA2X and sPLA2 XII was demonstrated to be elevated in induced sputum cells of patients with asthma. The level of sPLA2X in induced sputum cells supernatant increased after exercise challenge among asthmatics with exercise-induced bronchoconstriction (EIB) [39]. Lai et al. [51] have confirmed the involvement of sPLA2X. They demonstrated that recombinant sPLA2X caused AA release and rapid onset of cysLT synthesis in human eosinophils.
Limited information suggests a possible anti-inflammatory role of sPLA2X. However in asthma, sPLA2X facilitates the polarization toward proasthmatic M2-macrophage phenotype [52]. It is possible that in a proinflammatory environment, that the sPLA2X propeptide is more rapidly converted to an active form that might influence the Th1/Th2 balance [53]. All these factors may suppress its anti-inflammatory action.
Other sPLA2s (IIA, IID, IIE) contain a heparin-binding domain which allows these enzymes to be taken into the cells and further directed to compartments enriched in AA and enzymes responsible for eicosanoid production [54].
In spite of the fact that several studies have confirmed the participation of iPLA2 β [55] and iPLA2 γ [56] in AA release and eicosanoid production, there is no data indicating that these enzymes play a direct role in asthma. By the induction of Ca2+ influx they can influence the translocation and activity of Ca2+-dependent PLA2s isoforms.
Group VII and VIII PAF-AH hydrolyze the short sn-2 residue of PAF (platelet activating factor). As they lack activity against membrane phospholipids with long-chain sn-2 residues, they are unable to release arachidonic acid from membrane phospholipids [57]. They exhibit pro- and anti-inflammatory properties. On the one hand, they inactivate PAF—the proinflammatory mediator—by hydrolyzing it to inactive acetate and lysolipid but on the other hand, they assist in the generation of lysophospholipids and fatty acid hydroperoxides [4]. Stafforini et al. [58] have established that asthmatics have a decreased level of PAF-AH, and that asthma incidence and severity correlate to PAF-AH deficiency in the Japanese population. Also some PAF-AH gene polymorphisms (Ile198Thr and Ala379Val variants) are known to be a risk factors for developing atopy and asthma [59]. Despite positive effects in animal models [60], administration of human recombinant PAF-AH (rPAF-AH) does not reduce both early and late phase of asthmatic response in mild asthmatics challenged with allergens [61].
The enzymatic activity of PLA2s embraces also lysophospholipid generation. Lysophospholipids are biologically active molecules acting through specific receptors. They are a precursor of platelet activating factor (PAF) and lysophosphatidic acid (LPA). LPA is involved in cell adhesion, motility, and survival. In animal models, lysophospholipid receptors are required for proper development and function of the cardiovascular, immune, respiratory, and reproductive systems [62]. Lysophosphocholine and polyunsaturated fatty acids, including AA, can activate cPLA2 and 5-lipoxygenase by increasing Ca2+ and inducing cPLA2 phosphorylation, which then leads to LTB4 biosynthesis [25]. Lysophospholipid has nonspecific cytotoxic effect that depends on its concentration (critical micelle concentration). At concentration below their unspecific cytotoxic effect lysophospholipids can induce apoptosis by interrupting the synthesis of phosphatidylcholine [63].
Phospholipases A2 activity is also connected with disturbed lipid homeostasis in the lung. Asthma and other inflammatory lung diseases are characterized by impaired surfactant function [64]. Secretory phospholipases degrade phosphatidylcholine (PC), the main component of the surfactant responsible for maintenance of small airway patency. The generation of lysophospholipids and free fatty acids by sPLA2-mediated PC hydrolysis has been implicated in small airway closure in asthm. sPLA2 action is enhanced by eosinophilic lysophospholipases that use lysophospholipids as a substrate [65–68]. The presence of iPLA2 proteins in alveolar macrophages suggests that they might play a role in surfactant degradation [69].
It should be mentioned that some PLA2s are involved in antibacterial defense thanks to their ability to hydrolyze the lipids of the bacterial membrane. sPLA2s IIA, V, X, and IB demonstrate bactericidal activity against gram-positive pathogens but the most effective is sPLA2IIA. Group XII can directly kill E.coli, unlike the other sPLA2s that require cofactors [70]. This property of phospholipases can be important in bacterial exacerbations of asthma and COPD.
5.2. Nonenzymatic Activity of PLA2s
The secretory forms of many PLA2s exert a range of actions in airway inflammation. Apart from their enzymatic activity, they can act as extracellular mediators involved in chemotaxis, cytokine production, and induction of cellular signaling pathways.
Mammalian N-type receptors have been identified for sPLA2IB and IIA, X and M-type receptors for sPLA2IB, IIA, IIE, IIF, V, and X [71]. N-type like receptors are present in lungs whereas M-type receptors have been identified in lung and myeloid cells [72]. The binding of sPLA2s to their M-type receptor deactivates their enzymatic properties [73].
sPLA2s are stored in intrinsic mast cell granulates and are released after cell activation by IgE and non-IgE stimuli [9]. After exocytosis, they can act in both autocrine and paracrine manners. By interacting with heparan sulphate proteoglycans and M-type receptors, they can induce PGD2 and LTC4 production and stimulate the subsequent degranulation of mast cells [74]. Granata et al. [17] delivered an evidence that sPLA2s can act as proinflammatory connections between mast cells and macrophages in the airway. They suggest that the activation of macrophages by sPLA2s leads to production of proinflammatory cytokines which sustain the inflammatory and immune response, chemokines responsible for recruitment of monocytes and neutrophils, as well as destructive lysosomal enzymes, NO, PGE2, and metalloproteinases connected with airway remodeling [17]. The sPLA2s induce β-glucuronidase release and production of IL-6 from human lung macrophages [75]. They influence the migration and adhesion of neutrophils as well as the release of elastase [76, 77]. In eosinophils, sPLA2 IA and IIA stimulate β-glucuronidase release and cytokine production (IL-6, IL-8) by AA and lysophospholipid generation, by interaction with membrane peptidoglycans via their heparin-binding site, and through binding with specific M-type or N-type receptors [78]. The functions of sPLA2s receptors require further studies because there are still some missing or unequivocal information [52].
5.3. Crosstalk between PLA2s
The phospholipases can cooperate in mechanism leading to eicosanoid production. sPLA2 and cPLA2 interaction is quite well documented [79, 80]. The effect of group IIa and V PLA2s on H2O2-induced AA release is dependent upon the presence of cPLA2 and the activation of PKC and ERK1/2 in murine mesangial cells. Offer et al. [81] have described negative feedback between sPLA2 and cPLA2 in eicosanoid production. sPLA2 activation induces production of bronchoconstrictor cysteinyl leukotrienes and suppresses cPLA2 expression and the subsequent production of bronchodilator PGE2. Recently it has been established that in human eosinophils, sPLA2 initiates Ser(505) phosphorylation of cPLA2 α and stimulates leukotriene synthesis through involvement of p38 and JNK MAPK, cPLA2 α, and 5-lipoxygenase activation, which may be an important process also in airways of asthmatics [51]. Also in bone-marrow-derived mast cells, sPLA2 mediates the selective release of AA by binding M-type receptors and then inducing MAPK signaling pathways that lead to cPLA2 activation [82].
5.4. PLA2s in the Exacerbation of Disease
Another aspect of phospholipases and the asthma/COPD relationship is the participation of these enzymes in the pathogenetic mechanisms of disease exacerbation caused by bacterial factors. This role relates to increased expression of selective PLA2s, modulation of their activity and involvement in cellular signaling. Elevated cPLA2 α expression was found in primary human lung macrophages after LPS treatment [15, 83]. LPS stimulates expression of cPLA2 and COX-2 in macrophages, leading to increased production of AA and PGE2 [83]. LPS treatment was also followed by rapid changes in cPLA2 phosphorylation [84, 85]. This is one of the mechanisms of regulating enzyme activity [86]. The LPS-phosphorylated form of cPLA2 is present in induction of iNOS and TNF-α expression [87, 88] and metalloproteinase production [89]. Selective sPLA2 contributes to LPS-intracellular signaling in liver macrophages [84, 90, 91].
In mice with LPS-induced lung inflammation, the expression of sPLA2X remains the same before and after treatment. In this study, increased expression of sPLA2IID and sPLA2V has been observed, as well as decreased sPLA2IIE and sPLA2IIF levels in the lungs. In rats, sPLA2IIA was seen to have the highest expression after LPS administration [92]. In msPLA2X−/− mice with knock-in of human sPLA2X (hsPLA2X), allergen-induced inflammatory cell recruitment into airways (eosinophils) was restored, as well as hyperresponsiveness to methacholine. The application of specific hsPLA2X inhibitor (RO 061606) significantly attenuates airway inflammation symptoms, mucous secretion, and hyperresponsiveness [93]. In sPLA2V−/− knock-out mice, sPLA2V has been proven to play a role in the development of lung injury and neutrophilic inflammation after bacterial stimulus (LPS) [94]. In addition, sPLA2V was seen to be connected with regulation of cell migration and generation of airway hyperresponsiveness after ovalbumin challenge [95]. In a murine allergen-challenged asthma model, administration of rPAF-AH is effective in blocking late-phase pulmonary inflammation [60].
6. The Clinical Significance of Studying the Participation of PLA2s in Airway Inflammatory Diseases
Taking into consideration the severe asthma phenotype, the difficulties related to obtain asthma control utilizing currently available treatments and the progressive character of inflammation in patients with COPD that increases the morbidity, it seems reasonable to study the differences in pathogenesis of the diseases conditions, especially in relation to possible new therapies and drugs. The PLA2s are an interesting object of study for several reasons. The superfamily of these enzymes contains approximately 30 members that have similar and isoform-specific properties. It has been confirmed that they are strictly connected with inflammation. The inhibitors of particular PLA2s show the positive effect in treatment of inflammatory diseases [96] and they inhibit allergic reaction in vitro [38]. The cPLA2 α that evolved together with receptors for eicosanoids, present only in vertebrate, seems to play crucial role in course of inflammation. Its inhibitors such as efipladib [97] and ecopladib [98] successfully inhibit inflammation in rheumatoid arthritis and osteoporosis. The inhaled form of cPLA2 α inhibitor, the PLA-950, is considered as potential new treatment in asthmatic patients as well as other PLA2s can influence the function of cPLA2 α or have similar effects. Recent studies report positive results of a preclinical evaluation of a cPLA2 α inhibitor [99]. The studies and analysis of protein involved in regulation of particular sPLA2 involved in inflammatory diseases could result in finding new target for drugs.
Since 1980, it has been known that glucocorticoids (GCs) can inhibit the activity of PLA2 [100]. The underlying mechanism concerns induction of mRNA and protein expression of lipocortin 1 (annexin 1) and the PLA2 inhibitory protein [101–104]. The structure, function, and mechanism behind the anti-inflammatory action of annexin 1 have been well described elsewhere [105]. Glucocorticoids can also suppress the production of sPLA2IIA by blocking mRNA synthesis and posttranslational expression in rats [106]. It is questionable whether therapeutic doses of glucocorticoids have sufficient power to satisfactorily inhibit the activity of PLA2. Juergens et al. [107] demonstrated that topical GCs at therapeutically relevant concentration (10−8 M) inhibit the spontaneous activity of cPLA2 in the range of 8.6–17.3% depending on the type of GC. They suggest also that this effect may appear as a consequence of a decreased ability to binding the receptors by GCs present in airway in subtherapeutical doses. Although it has been established that treatment with GCs can indirectly inhibit cPLA2 and AA-derivates production resistance to GCs in patients with asthma and COPD could also be problematic. Moreover the GCs have systemic effects and long-term application can cause the side effects. The approach to attack the inflammation process more precisely and downstream (inhibition the eicosanoids production) seems to be rationale.
Another aspect regarding annexin 1 and PLA2s is their cell-specific manner of interactions [105]. Kwon et al. [108] demonstrated that cleavage of annexin 1 causes phosphorylation of cPLA2 during mast-cell activation. Hence it is not clear whether GCs-induced expression of annexin always leads to inhibition of cPLA2 activity. Posttranslational changes can dramatically influence the primary protein function. As previous studies indicate that GCs can stimulate expression of cPLA2 in amnion fibroblast it cannot be excluded that in some specific circumstances GCs may directly induce cPLA2 [109, 110].
7. Conclusions
Previous studies confirm the involvement of phospholipases A2 in asthma and COPD although there are some gaps relating to the roles of specific enzymes. The participation of PLA2 in asthma pathogenesis has been better investigated. The diagnostic problems concerning the overlap syndrome that shares the features of asthma and COPD demand further studies on the pathogenesis of these diseases. The phospholipases A2 through their involvement in the course of inflammation seem to be important aspects of this investigation. As they demonstrate pro- and anti-inflammatory properties, a detailed analysis of their role should act as a focus for further studies intended to bring new insights into the pathogenesis of the diseases and identify targets for new drugs.
Data from studies focused on role of PLA2s in inflammatory diseases facilitate the understanding of molecular aspects of inflammation. It can be observed that cPLA2 plays a main role in eicosanoid production and other PLA2s may influence their activity thanks to enzymatic properties or act as regulators of inflammation through their nonenzymatic activity. The pleiotropic properties of single phospholipase and their differential expression in many cells confirm that this is well-organized network of interaction, and further studies focused on this aspect may provide more useful knowledge. A comparison of how this network works in different inflammatory diseases, as well as in healthy subjects may indicate a key molecule, whose activity or presence will be a diagnostic parameter or whose activation or inhibition will have therapeutic value.
Asthma and COPD are heterogeneous diseases and current treatment gives only the possibility to obtain the phenotype of well-controlled diseases. Analysis of data regarding the involvement of PLA2s in course of diseases arises the concept to use combined therapy rather than the treatment based on inhibition of one of them. The results from preclinical studies of cPLA2 inhibitors are promising but clinical trials will give concrete knowledge about the effectiveness and possible side effects.
Acknowledgments
This paper was partially funded by Polish National Science Centre Grant no. DEC-2012/05/N/NZ5/02630 and Medical University of Lodz Grants: 502-03/0-149-03/502-04-004 and 503/0-149-03/503-01 and was supported from science budget for years 2010-2013 as research project (N N402 516939).
References
- 1.Schaloske RH, Dennis EA. The phospholipase A2 superfamily and its group numbering system. Biochimica et Biophysica Acta. 2006;1761(11):1246–1259. doi: 10.1016/j.bbalip.2006.07.011. [DOI] [PubMed] [Google Scholar]
- 2.Masoli M, Fabian D, Holt S, Beasley R. The global burden of asthma: executive summary of the GINA Dissemination Committee Report. Allergy. 2004;59(5):469–478. doi: 10.1111/j.1398-9995.2004.00526.x. [DOI] [PubMed] [Google Scholar]
- 3. Global Strategy for Asthma Management and Prevention. Global Initiative for Asthma (GINA), May 2012, http://www.ginasthma.org.
- 4.Kitsiouli E, Nakos G, Lekka ME. Phospholipase A2 subclasses in acute respiratory distress syndrome. Biochimica et Biophysica Acta. 2009;1792(10):941–953. doi: 10.1016/j.bbadis.2009.06.007. [DOI] [PubMed] [Google Scholar]
- 5.Burke JE, Dennis EA. Phospholipase A2 structure/function, mechanism, and signaling. Journal of Lipid Research. 2009;50:S237–S242. doi: 10.1194/jlr.R800033-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. The Global Initiative for Chronic Obstructive Lung Disease (GOLD), May 2012, http://www.goldcopd.com/
- 7.Nakamura H. Genetics of COPD. Allergology International. 2011;60(3):253–258. doi: 10.2332/allergolint.11-RAI-0326. [DOI] [PubMed] [Google Scholar]
- 8.Silverman EK, Speizer FE. Risk factors for the development of chronic obstructive pulmonary disease. Medical Clinics of North America. 1996;80(3):501–522. doi: 10.1016/s0025-7125(05)70451-x. [DOI] [PubMed] [Google Scholar]
- 9.Lambeau G, Gelb MH. Biochemistry and physiology of mammalian secreted phospholipases A2 . Annual Review of Biochemistry. 2008;77:495–520. doi: 10.1146/annurev.biochem.76.062405.154007. [DOI] [PubMed] [Google Scholar]
- 10.Triggiani M, Giannattasio G, Calabrese C, et al. Lung mast cells are a source of secreted phospholipases A2 . Journal of Allergy and Clinical Immunology. 2009;124(3):558.e3–565.e3. doi: 10.1016/j.jaci.2009.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Muñoz NM, Kim YJ, Meliton AY, et al. Human group V phospholipase A2 induces group IVA phospholipase A2-independent cysteinyl leukotriene synthesis in human eosinophils. Journal of Biological Chemistry. 2003;278(40):38813–38820. doi: 10.1074/jbc.M302476200. [DOI] [PubMed] [Google Scholar]
- 12.Seeds MC, Peachman KK, Bowton DL, Sivertson KL, Chilton FH. Regulation of arachidonate remodeling enzymes impacts eosinophil survival during allergic asthma. American Journal of Respiratory Cell and Molecular Biology. 2009;41(3):358–366. doi: 10.1165/rcmb.2008-0192OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Barnes PJ. Alveolar macrophages as orchestrators of COPD. Journal of Chronic Obstructive Pulmonary Disease. 2004;1(1):59–70. doi: 10.1081/COPD-120028701. [DOI] [PubMed] [Google Scholar]
- 14.Ichinose M. Differences of inflammatory mechanisms in asthma and COPD. Allergology International. 2009;58(3):307–313. doi: 10.2332/allergolint.09-RAI-0106. [DOI] [PubMed] [Google Scholar]
- 15.Giannattasio G, Lai Y, Granata F, et al. Expression of phospholipases A2 in primary human lung macrophages. Role of cytosolic phospholipase A2-α in arachidonic acid release and platelet activating factor synthesis. Biochimica et Biophysica Acta. 2009;1791(2):92–102. doi: 10.1016/j.bbalip.2008.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Degousee N, Ghomashchi F, Stefanski E, et al. Groups IV, V, and X phospholipases A2s in human neutrophils. Role in eicosanoid production and gram-negative bacterial phospholipid hydrolysis. Journal of Biological Chemistry. 2002;277(7):5061–5073. doi: 10.1074/jbc.M109083200. [DOI] [PubMed] [Google Scholar]
- 17.Granata F, Nardicchi V, Loffredo S, et al. Secreted phospholipases A2: a proinflammatory connection between macrophages and mast cells in the human lung. Immunobiology. 2009;214(9-10):811–821. doi: 10.1016/j.imbio.2009.06.006. [DOI] [PubMed] [Google Scholar]
- 18.Yamashita JI, Ogawa M, Shirakusa T. Increased expression of membrane-associated phospholipase A2 in the lower respiratory tract of asymptomatic cigarette smokers. Respiratory Medicine. 1996;90(8):479–483. doi: 10.1016/s0954-6111(96)90175-4. [DOI] [PubMed] [Google Scholar]
- 19.Igarashi A, Shibata Y, Yamauchi K, et al. Gly80Ser polymorphism of phospholipase A2-IID is associated with cytokine inducibility in A549 cells. Respiration. 2009;78(3):312–321. doi: 10.1159/000213243. [DOI] [PubMed] [Google Scholar]
- 20.Von Allmen CE, Schmitz N, Bauer M, et al. Secretory phospholipase A2-IID is an effector molecule of CD4+ CD25+ regulatory T cells. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(28):11673–11678. doi: 10.1073/pnas.0812569106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nana SP, Lee JD, Sotto-Santiago S, et al. Prostacyclin prevents pulmonary endothelial cell apoptosis induced by cigarette smoke. American Journal of Respiratory and Critical Care Medicine. 2007;175(7):676–685. doi: 10.1164/rccm.200605-724OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pawliczak R. The role of radical oxygen species in airway inflammation. Polski Merkuriusz Lekarski. 2003;14(84):493–496. [PubMed] [Google Scholar]
- 23.Meliton AY, Muñoz NM, Lambertino A, et al. Phosphodiesterase 4 inhibition of β2-integrin adhesion caused by leukotriene B4 and TNF-α in human neutrophils. European Respiratory Journal. 2006;28(5):920–928. doi: 10.1183/09031936.06.00028406. [DOI] [PubMed] [Google Scholar]
- 24.Tanaseanu C, Tudor S, Tamsulea I, Marta D, Manea G, Moldoveanu E. Vascular endothelial growth factor, lipoporotein-associated phospholipase A2, sP-selectin and antiphospholipid antibodies, biological markers with prognostic value in pulmonary hypertension associated with chronic obstructive pulmonary disease and systemic lupus erithematosus. European Journal of Medical Research. 2007;12(4):145–151. [PubMed] [Google Scholar]
- 25.Kim YJ, Kim KP, Han SK, et al. Group V phospholipase A2 induces leukotriene biosynthesis in human neutrophils through the activation of group IVA phospholipase A2 . Journal of Biological Chemistry. 2002;277(39):36479–36488. doi: 10.1074/jbc.M205399200. [DOI] [PubMed] [Google Scholar]
- 26.Bowton DL, Dmitrienko AA, Israel E, Zeiher BG, Sides GD. Impact of a soluble phospholipase A2 inhibitor on inhaled allergen challenge in subjects with asthma. Journal of Asthma. 2005;42(1):65–71. doi: 10.1081/jas-200044748. [DOI] [PubMed] [Google Scholar]
- 27.Dennis EA. Phospholipase A2 in eicosanoid generation. American Journal of Respiratory and Critical Care Medicine. 2000;161(2, part 2):S32–S35. doi: 10.1164/ajrccm.161.supplement_1.ltta-7. [DOI] [PubMed] [Google Scholar]
- 28.Shuttleworth TJ. Arachidonic acid activates the noncapacitative entry of Ca2+ during [Ca2+]i oscillations. Journal of Biological Chemistry. 1996;271(36):21720–21725. doi: 10.1074/jbc.271.36.21720. [DOI] [PubMed] [Google Scholar]
- 29.Graber MN, Alfonso A, Gill DL. Ca2+ pools and cell growth: arachidonic acid induces recovery of cells growth-arrested by Ca2+ pool depletion. Journal of Biological Chemistry. 1996;271(2):883–888. doi: 10.1074/jbc.271.2.883. [DOI] [PubMed] [Google Scholar]
- 30.Funk CD. Prostaglandins and leukotrienes: advances in eicosanoid biology. Science. 2001;294(5548):1871–1875. doi: 10.1126/science.294.5548.1871. [DOI] [PubMed] [Google Scholar]
- 31.Uddin M, Levy BD. Resolvins: natural agonists for resolution of pulmonary inflammation. Progress in Lipid Research. 2011;50(1):75–88. doi: 10.1016/j.plipres.2010.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Planagumà A, Kazani S, Marigowda G, et al. Airway lipoxin A4 generation and lipoxin A4 receptor expression are decreased in severe asthma. American Journal of Respiratory and Critical Care Medicine. 2008;178(6):574–582. doi: 10.1164/rccm.200801-061OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Levy BD, Bonnans C, Silverman ES, Palmer LJ, Marigowda C, Israel E. Diminished lipoxin biosynthesis in severe asthma. American Journal of Respiratory and Critical Care Medicine. 2005;172(7):824–830. doi: 10.1164/rccm.200410-1413OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Adler DH, Cogan JD, Phillips JA, et al. Inherited human cPLA2 α deficiency is associated with impaired eicosanoid biosynthesis, small intestinal ulceration, and platelet dysfunction. Journal of Clinical Investigation. 2008;118(6):2121–2131. doi: 10.1172/JCI30473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Reed KA, Tucker DE, Aloulou A, et al. Functional characterization of mutations in inherited human cPLA2 deficiency. Biochemistry. 2011;50(10):1731–1738. doi: 10.1021/bi101877n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sokolowska M, Borowiec M, Ptasinska A, et al. 85-kDa cytosolic phospholipase A2 group IVα gene promoter polymorphisms in patients with severe asthma: a gene expression and case-control study. Clinical and Experimental Immunology. 2007;150(1):124–131. doi: 10.1111/j.1365-2249.2007.03459.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sokolowska M, Stefanska J, Wodz-Naskiewicz K, Pawliczak R. Cytosolic phospholipase A2 group IVA influence on GM-CSF expression in human lung cells: a pilot study. Medical Science Monitor. 2010;16(9):BR300–BR306. [PubMed] [Google Scholar]
- 38.Whalen KA, Legault H, Hang C, et al. In vitro allergen challenge of peripheral blood induces differential gene expression in mononuclear cells of asthmatic patients: inhibition of cytosolic phospholipase A2 α overcomes the asthma-associated response. Clinical and Experimental Allergy. 2008;38(10):1590–1605. doi: 10.1111/j.1365-2222.2008.03059.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hallstrand TS, Chi EY, Singer AG, Gelb MH, Henderson WR. Secreted phospholipase A2 group X overexpression in asthma and bronchial hyperresponsiveness. American Journal of Respiratory and Critical Care Medicine. 2007;176(11):1072–1078. doi: 10.1164/rccm.200707-1088OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Asai K, Hirabayashi T, Houjou T, Uozumi N, Taguchi R, Shimizu T. Human group IVC phospholipase A2 (cPLA2 γ): roles in the membrane remodeling and activation induced by oxidative stress. Journal of Biological Chemistry. 2003;278(10):8809–8814. doi: 10.1074/jbc.M212117200. [DOI] [PubMed] [Google Scholar]
- 41.Mancuso DJ, Abendschein DR, Jenkins CM, et al. Cardiac ischemia activates calcium-independent phospholipase A2 β, precipitating ventricular tachyarrhythmias in transgenic mice: rescue of the lethal electrophysiologic phenotype by mechanism-based inhibition. Journal of Biological Chemistry. 2003;278(25):22231–22236. doi: 10.1074/jbc.C300033200. [DOI] [PubMed] [Google Scholar]
- 42.Gauvreau GM, Watson RM, O’Byrne PM. Protective effects of inhaled PGE2 on allergen-induced airway responses and airway inflammation. American Journal of Respiratory and Critical Care Medicine. 1999;159(1):31–36. doi: 10.1164/ajrccm.159.1.9804030. [DOI] [PubMed] [Google Scholar]
- 43.Chung KF. Evaluation of selective prostaglandin E2 (PGE2) receptor agonists as therapeutic agents for the treatment of asthma. Science’s STKE. 2005;2005(303):p. pe47. doi: 10.1126/stke.3032005pe47. [DOI] [PubMed] [Google Scholar]
- 44.Sastre B, del Pozo V. Role of PGE2 in asthma and nonasthmatic eosinophilic bronchitis. Mediators of Inflammation. 2012;2012:9 pages. doi: 10.1155/2012/645383.645383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Phipps RP, Stein SH, Roper RL. A new view of prostaglandin E regulation of the immune response. Immunology Today. 1991;12(10):349–352. doi: 10.1016/0167-5699(91)90064-Z. [DOI] [PubMed] [Google Scholar]
- 46.Yedgar S, Krimsky M, Cohen Y, Flower RJ. Treatment of inflammatory diseases by selective eicosanoid inhibition: a double-edged sword? Trends in Pharmacological Sciences. 2007;28(9):459–464. doi: 10.1016/j.tips.2007.07.005. [DOI] [PubMed] [Google Scholar]
- 47.Chilton FH, Averill FJ, Hubbard WC, Fonteh AN, Triggiani M, Liu MC. Antigen-induced generation of lyso-phospholipids in human airways. Journal of Experimental Medicine. 1996;183(5):2235–2245. doi: 10.1084/jem.183.5.2235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Bowton DL, Seeds MC, Fasano MB, Goldsmith B, Bass DA. Phospholipase A2 and arachidonate increase in bronchoalveolar lavage fluid after inhaled antigen challenge in asthmatics. American Journal of Respiratory and Critical Care Medicine. 1997;155(2):421–425. doi: 10.1164/ajrccm.155.2.9032172. [DOI] [PubMed] [Google Scholar]
- 49.Henderson WR, Chi EY, Bollinger JG, et al. Importance of group X-secreted phospholipase A2 in allergen-induced airway inflammation and remodeling in a mouse asthma model. Journal of Experimental Medicine. 2007;204(4):865–877. doi: 10.1084/jem.20070029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hallstrand TS, Lai Y, Ni Z, et al. Relationship between levels of secreted phospholipase A2 groups IIA and X in the airways and asthma severity. Clinical and Experimental Allergy. 2011;41(6):801–810. doi: 10.1111/j.1365-2222.2010.03676.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lai Y, Oslund RC, Bollinger JG, et al. Eosinophil cysteinyl leukotriene synthesis mediated by exogenous secreted phospholipase A2 group X. Journal of Biological Chemistry. 2010;285(53):41491–41500. doi: 10.1074/jbc.M110.153338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Murakami M, Taketomi Y, Sato H, Yamamoto K. Secreted phospholipase A2 revisited. Journal of Biochemistry. 2011;150(3):233–255. doi: 10.1093/jb/mvr088. [DOI] [PubMed] [Google Scholar]
- 53.Murakami M, Taketomi Y, Miki Y, Sato H, Hirabayashi T, Yamamoto K. Recent progress in phospholipase A2 research: from cells to animals to humans. Progress in Lipid Research. 2011;50(2):152–192. doi: 10.1016/j.plipres.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 54.Kudo I, Murakami M. Phospholipase A2 enzymes. Prostaglandins and Other Lipid Mediators. 2002;68-69:3–58. doi: 10.1016/s0090-6980(02)00020-5. [DOI] [PubMed] [Google Scholar]
- 55.Tay HK, Melendez AJ. FcγRI-triggered generation of arachidonic acid and eicosanoids requires iPLA2 but not cPLA2 in human monocytic cells. Journal of Biological Chemistry. 2004;279(21):22505–22513. doi: 10.1074/jbc.M308788200. [DOI] [PubMed] [Google Scholar]
- 56.Murakami M, Masuda S, Ueda-Semmyo K, et al. Group VIB Ca2+-independent phospholipase A2 γ promotes cellular membrane hydrolysis and prostaglandin production in a manner distinct from other intracellular phospholipases A2 . Journal of Biological Chemistry. 2005;280(14):14028–14041. doi: 10.1074/jbc.M413766200. [DOI] [PubMed] [Google Scholar]
- 57.McIntyre TM, Prescott SM, Stafforini DM. The emerging roles of PAF acetylhydrolase. Journal of Lipid Research. 2009;50:S255–S259. doi: 10.1194/jlr.R800024-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Stafforini DM, Numao T, Tsodikov A, et al. Deficiency of platelet-activating factor acetylhydrolase is a severity factor for asthma. Journal of Clinical Investigation. 1999;103(7):989–997. doi: 10.1172/JCI5574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kruse S, Mao XQ, Heinzmann A, et al. The Ile198Thr and Ala379Val variants of plasmatic Paf-acetylhydrolase impair catalytical activities and are associated with atopy and asthma. American Journal of Human Genetics. 2000;66(5):1522–1530. doi: 10.1086/302901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Henderson WR, Lu J, Poole KM, Dietsch GN, Chi EY. Recombinant human platelet-activating factor-acetylhydrolase inhibits airway inflammation and hyperreactivity in mouse asthma model. Journal of Immunology. 2000;164(6):3360–3367. doi: 10.4049/jimmunol.164.6.3360. [DOI] [PubMed] [Google Scholar]
- 61.Henig NR, Aitken ML, Liu MC, Yu AS, Henderson WR. Effect of recombinant human platelet-activating factor-acetylhydrolase on allergen-induced asthmatic responses. American Journal of Respiratory and Critical Care Medicine. 2000;162(2, part 1):523–527. doi: 10.1164/ajrccm.162.2.9911084. [DOI] [PubMed] [Google Scholar]
- 62.Rivera R, Chun J. Biological effects of lysophospholipids. Reviews of Physiology, Biochemistry and Pharmacology. 2008;160:25–46. doi: 10.1007/112_0507. [DOI] [PubMed] [Google Scholar]
- 63.Urbina JA. Mechanisms of action of lysophospholipid analogues against trypanosomatid parasites. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2006;100(supplement 1):S9–S16. doi: 10.1016/j.trstmh.2006.03.010. [DOI] [PubMed] [Google Scholar]
- 64.Hurley BP, McCormick BA. Multiple roles of phospholipase A2 during lung infection and inflammation. Infection and Immunity. 2008;76(6):2259–2272. doi: 10.1128/IAI.00059-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Hite RD, Seeds MC, Jacinto RB, Grier BL, Waite BM, Bass DA. Lysophospholipid and fatty acid inhibition of pulmonary surfactant: non-enzymatic models of phospholipase A2 surfactant hydrolysis. Biochimica et Biophysica Acta. 2005;1720(1-2):14–21. doi: 10.1016/j.bbamem.2005.10.014. [DOI] [PubMed] [Google Scholar]
- 66.Kwatia MA, Doyle CB, Cho W, Enhorning G, Ackerman SJ. Combined activities of secretory phospholipases and eosinophil lysophospholipases induce pulmonary surfactant dysfunction by phospholipid hydrolysis. Journal of Allergy and Clinical Immunology. 2007;119(4):838–847. doi: 10.1016/j.jaci.2006.12.614. [DOI] [PubMed] [Google Scholar]
- 67.Ackerman SJ, Kwatia MA, Doyle CB, Enhorning G. Hydrolysis of surfactant phospholipids catalyzed by phospholipase A2 and eosinophil lysophospholipases causes surfactant dysfunction: a mechanism for small airway closure in asthma. Chest. 2003;123(3):p. 355S. doi: 10.1378/chest.123.3_suppl.355s. [DOI] [PubMed] [Google Scholar]
- 68.Enhorning G. Surfactant in airway disease. Chest. 2008;133(4):975–980. doi: 10.1378/chest.07-2404. [DOI] [PubMed] [Google Scholar]
- 69.Hiraoka M, Abe A, Lu Y, et al. Lysosomal phospholipase A2 and phospholipidosis. Molecular and Cellular Biology. 2006;26(16):6139–6148. doi: 10.1128/MCB.00627-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Koduri RS, Grönroos JO, Laine VJO, et al. Bactericidal properties of human and murine groups I, II, V, X, and XII secreted phospholipases A2 . Journal of Biological Chemistry. 2002;277(8):5849–5857. doi: 10.1074/jbc.M109699200. [DOI] [PubMed] [Google Scholar]
- 71.Rouault M, Le Calvez C, Boilard E, et al. Recombinant production and properties of binding of the full set of mouse secreted phospholipases A2 to the mouse M-type receptor. Biochemistry. 2007;46(6):1647–1662. doi: 10.1021/bi062119b. [DOI] [PubMed] [Google Scholar]
- 72.Triggiani M, Granata F, Frattini A, Marone G. Activation of human inflammatory cells by secreted phospholipases A2 . Biochimica et Biophysica Acta. 2006;1761(11):1289–1300. doi: 10.1016/j.bbalip.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 73.Ancian P, Lambeau G, Lazdunski M. Multifunctional activity of the extracellular domain of the M-type (180 kDa) membrane receptor for secretory phospholipases A2 . Biochemistry. 1995;34(40):13146–13151. doi: 10.1021/bi00040a028. [DOI] [PubMed] [Google Scholar]
- 74.Granata F, Staiano RI, Loffredo S, et al. The role of mast cell-derived secreted phospholipases A2 in respiratory allergy. Biochimie. 2010;92(6):588–593. doi: 10.1016/j.biochi.2010.02.030. [DOI] [PubMed] [Google Scholar]
- 75.Triggiani M, Granata F, Oriente A, et al. Secretory phospholipases A2 induce β-glucuronidase release and IL-6 production from human lung macrophages. Journal of Immunology. 2000;164(9):4908–4915. doi: 10.4049/jimmunol.164.9.4908. [DOI] [PubMed] [Google Scholar]
- 76.Silliman CC, Moore EE, Zallen G, et al. Presence of the M-type sPLA2 receptor on neutrophils and its role in elastase release and adhesion. American Journal of Physiology. 2002;283(4):C1102–C1113. doi: 10.1152/ajpcell.00608.2001. [DOI] [PubMed] [Google Scholar]
- 77.Gambero A, Landucci ECT, Toyama MH, et al. Human neutrophil migration in vitro induced by secretory phospholipases A2: a role for cell surface glycosaminoglycans. Biochemical Pharmacology. 2002;63(1):65–72. doi: 10.1016/s0006-2952(01)00841-3. [DOI] [PubMed] [Google Scholar]
- 78.Triggiani M, Granata F, Balestrieri B, et al. Secretory phospholipases A2 activate selective functions in human eosinophils. Journal of Immunology. 2003;170(6):3279–3288. doi: 10.4049/jimmunol.170.6.3279. [DOI] [PubMed] [Google Scholar]
- 79.Ni Z, Okeley NM, Smart BP, Gelb MH. Intracellular actions of group IIA secreted phospholipase A2 and group IVA cytosolic phospholipase A2 contribute to arachidonic acid release and prostaglandin production in rat gastric mucosal cells and transfected human embryonic kidney cells. Journal of Biological Chemistry. 2006;281(24):16245–16255. doi: 10.1074/jbc.M513874200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Balboa MA, Pérez R, Balsinde J. Amplification mechanisms of inflammation: paracrine stimulation of arachidonic acid mobilization by secreted phospholipase A2 is regulated by cytosolic phospholipase A2-derived hydroperoxyeicosatetraenoic acid. Journal of Immunology. 2003;171(2):989–994. doi: 10.4049/jimmunol.171.2.989. [DOI] [PubMed] [Google Scholar]
- 81.Offer S, Yedgar S, Schwob O, et al. Negative feedback between secretory and cytosolic phospholipase A2 and their opposing roles in ovalbumin-induced bronchoconstriction in rats. American Journal of Physiology. 2005;288(3):L523–L529. doi: 10.1152/ajplung.00199.2004. [DOI] [PubMed] [Google Scholar]
- 82.Fonteh AN, Atsumi GI, LaPorte T, Chilton FH. Secretory phospholipase A2 receptor-mediated activation of cytosolic phospholipase A2 in murine bone marrow-derived mast cells. Journal of Immunology. 2000;165(5):2773–2782. doi: 10.4049/jimmunol.165.5.2773. [DOI] [PubMed] [Google Scholar]
- 83.Jiang YJ, Lu B, Choy PC, Hatch GM. Regulation of cytosolic phospholipase A2, cyclooxygenase-1 and -2 expression by PMA, TNFα, LPS and M-CSF in human monocytes and macrophages. Molecular and Cellular Biochemistry. 2003;246(1-2):31–38. [PubMed] [Google Scholar]
- 84.Dieter P, Kolada A, Kamionka S, Schadow A, Kaszkin M. Lipopolysaccharide-induced release of arachidonic acid and prostaglandins in liver macrophages: regulation by Group IV cytosolic phospholipase A2, but not by Group V and Group IIA secretory phospholipase A2 . Cellular Signalling. 2002;14(3):199–204. doi: 10.1016/s0898-6568(01)00243-1. [DOI] [PubMed] [Google Scholar]
- 85.Qi HY, Shelhamer JH. Toll-like receptor 4 signaling regulates cytosolic phospholipase A2 activation and lipid generation in lipopolysaccharide-stimulated macrophages. Journal of Biological Chemistry. 2005;280(47):38969–38975. doi: 10.1074/jbc.M509352200. [DOI] [PubMed] [Google Scholar]
- 86.Tian W, Wijewickrama GT, Kim JH, et al. Mechanism of regulation of group IVA phospholipase A2 activity by Ser727 phosphorylation. Journal of Biological Chemistry. 2008;283(7):3960–3971. doi: 10.1074/jbc.M707345200. [DOI] [PubMed] [Google Scholar]
- 87.Zhou X, Yang W, Li J. Ca2+- and protein kinase C-dependent signaling pathway for nuclear factor-κB activation, inducible nitric-oxide synthase expression, and tumor necrosis factor-α production in lipopolysaccharide-stimulated rat peritoneal macrophages. Journal of Biological Chemistry. 2006;281(42):31337–31347. doi: 10.1074/jbc.M602739200. [DOI] [PubMed] [Google Scholar]
- 88.Coffey MJ, Phare SM, Peters-Golden M. Induction of inducible nitric oxide synthase by lipopolysaccharide/ interferon gamma and sepsis down-regulates 5-lipoxygenase metabolism in murine alveolar macrophages. Experimental Lung Research. 2004;30(7):615–633. doi: 10.1080/01902140490476391. [DOI] [PubMed] [Google Scholar]
- 89.Shankavaram UT, DeWitt DL, Wahl LM. Lipopolysaccharide induction of monocyte matrix metalloproteinases is regulated by the tyrosine phosphorylation of cytosolic phospholipase A2 . Journal of Leukocyte Biology. 1998;64(2):221–227. doi: 10.1002/jlb.64.2.221. [DOI] [PubMed] [Google Scholar]
- 90.Kessen UA, Schaloske RH, Stephens DL, Lucas KK, Dennis EA. PGE2 release is independent of upregulation of Group V phospholipase A2 during long-term stimulation of P388D1 cells with LPS. Journal of Lipid Research. 2005;46(11):2488–2496. doi: 10.1194/jlr.M500325-JLR200. [DOI] [PubMed] [Google Scholar]
- 91.Shridas P, Bailey WM, Talbott KR, Oslund RC, Gelb MH, Webb NR. Group X secretory phospholipase A2 enhances TLR4 signaling in macrophages. Journal of Immunology. 2011;187(1):482–489. doi: 10.4049/jimmunol.1003552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hamaguchi K, Kuwata H, Yoshihara K, et al. Induction of distinct sets of secretory phospholipase A2 in rodents during inflammation. Biochimica et Biophysica Acta. 2003;1635(1):37–47. doi: 10.1016/j.bbalip.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 93.Hendersen WR, Oslund RC, Bollinger JG, Ye X, Tien YT, Xue J, et al. Blockade of human group X secreted phospholipase A2-induced airway inflammation and hyperresponsiveness in a mouse asthma model by a selective group X secreted phospholipase A2 inhibitor. doi: 10.1074/jbc.M111.235812. The Journal of Biological Chemistry. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Muñoz NM, Meliton AY, Meliton LN, Dudek SM, Leff AR. Secretory group V phospholipase A2 regulates acute lung injury and neutrophilic inflammation caused by LPS in mice. American Journal of Physiology. 2009;296(6):L879–L887. doi: 10.1152/ajplung.90580.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Muñoz NM, Meliton AY, Arm JP, Bonventre JV, Cho W, Leff AR. Deletion of secretory group V phospholipase A2 attenuates cell migration and airway hyperresponsiveness in immunosensitized mice. The Journal of Immunology. 2007;179(7):4800–4807. doi: 10.4049/jimmunol.179.7.4800. [DOI] [PubMed] [Google Scholar]
- 96.Malaviya R, Ansell J, Hall L, et al. Targeting cytosolic phospholipase A2 by arachidonyl trifluoromethyl ketone prevents chronic inflammation in mice. European Journal of Pharmacology. 2006;539(3):195–204. doi: 10.1016/j.ejphar.2006.03.018. [DOI] [PubMed] [Google Scholar]
- 97.McKew JC, Lee KL, Shen MWH, et al. Indole cytosolic phospholipase A2 α inhibitors: discovery and in vitro and in vivo characterization of 4-{3-[5-chloro-2-(2-{[(3,4- dichlorobenzyl)sulfonyl]amino}ethyl)-1-(diphenylmethyl)-1H-indol-3-yl]propyl} benzoic acid, efipladib. Journal of Medicinal Chemistry. 2008;51(12):3388–3413. doi: 10.1021/jm701467e. [DOI] [PubMed] [Google Scholar]
- 98.Lee KL, Foley MA, Chen L, et al. Discovery of ecopladib, an indole inhibitor of cytosolic phospholipase A2 α . Journal of Medicinal Chemistry. 2007;50(6):1380–1400. doi: 10.1021/jm061131z. [DOI] [PubMed] [Google Scholar]
- 99.Hewson CA, Patel S, Calzetta L, et al. Preclinical evaluation of an inhibitorof cytosolic phospholipase A2 α for the treatment of asthma. Journal of Pharmacology and Experimental Therapeutics. 2012;340(3):656–665. doi: 10.1124/jpet.111.186379. [DOI] [PubMed] [Google Scholar]
- 100.Hirata F, Schiffmann E, Venkatasubramanian K. A phospholipase A2 inhibitory protein in rabbit neutrophils induced by glucocorticoids. Proceedings of the National Academy of Sciences of the United States of America. 1980;77(5):2533–2536. doi: 10.1073/pnas.77.5.2533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.De Caterina R, Sicari R, Giannessi D, et al. Macrophage-specific eicosanoid synthesis inhibition and lipocortin-1 induction by glucocorticoids. Journal of Applied Physiology. 1993;75(6):2368–2375. doi: 10.1152/jappl.1993.75.6.2368. [DOI] [PubMed] [Google Scholar]
- 102.Huang X, Pawliczak R, Yao XL, et al. Characterization of the human p11 promoter sequence. Gene. 2003;310(1-2):133–142. doi: 10.1016/s0378-1119(03)00529-8. [DOI] [PubMed] [Google Scholar]
- 103.Huang XL, Pawliczak R, Yao XL, et al. Interferon-γ induces p11 gene and protein expression in human epithelial cells through interferon-γ-activated sequences in the p11 promoter. Journal of Biological Chemistry. 2003;278(11):9298–9308. doi: 10.1074/jbc.m212704200. [DOI] [PubMed] [Google Scholar]
- 104.Yao XL, Cowan MJ, Gladwin MT, Lawrence MM, Angus CW, Shelhamer JH. Dexamethasone alters arachidonate release from human epithelial cells by induction of p11 protein synthesis and inhibition of phospholipase A2 activity. Journal of Biological Chemistry. 1999;274(24):17202–17208. doi: 10.1074/jbc.274.24.17202. [DOI] [PubMed] [Google Scholar]
- 105.Lim LHK, Pervaiz S. Annexin 1: the new face of an old molecule. The FASEB Journal. 2007;21(4):968–975. doi: 10.1096/fj.06-7464rev. [DOI] [PubMed] [Google Scholar]
- 106.Nakano T, Ohara O, Teraoka H, Arita H. Glucocorticoids suppress group II phospholipase A2 production by blocking mRNA synthesis and post-transcriptional expression. Journal of Biological Chemistry. 1990;265(21):12745–12748. [PubMed] [Google Scholar]
- 107.Juergens UR, Jäger F, Darlath W, Stöber M, Vetter H, Gillissen A. Comparison of in vitro-activity of common used topical glucocorticoids on cytokine- and phospholipase inhibition. European Journal of Medical Research. 2004;9(8):383–390. [PubMed] [Google Scholar]
- 108.Kwon JH, Lee JH, Kim KS, Chung YW, Kim IY. Regulation of cytosolic phospholipase A2 phosphorylation by proteolytic cleavage of annexin A1 in activated mast cells. The Journal of Immunology. 2012;188(11):5665–5673. doi: 10.4049/jimmunol.1102306. [DOI] [PubMed] [Google Scholar]
- 109.Guo C, Li J, Myatt L, Zhu X, Sun K. Induction of Gαs contributes to the paradoxical stimulation of cytosolic phospholipase A2 α expression by cortisol in human amnion fibroblasts. Molecular Endocrinology. 2010;24(5):1052–1061. doi: 10.1210/me.2009-0488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Guo C, Yang Z, Li W, Zhu P, Myatt L, Sun K. Paradox of glucocorticoid-induced cytosolic phospholipase A2 group IVA messenger RNA expression involves glucocorticoid receptor binding to the promoter in human amnion fibroblasts. Biology of Reproduction. 2008;78(1):193–197. doi: 10.1095/biolreprod.107.063990. [DOI] [PubMed] [Google Scholar]