

Abstract
Background and Aims
Reciprocal herkogamy, including enantiostyly and heterostyly, involves reciprocity in the relative positions of the sexual elements within the flower. Such systems result in morphologically and, since pollen is deposited on and captured from different parts of the pollinator, functionally distinct floral forms. Deviations from the basic pattern may modify the functionality of these mechanisms. For heterostylous species, such deviations are generally related to environmental disturbances, pollination services and/or reduced numbers of one floral morph. Deviations for enantiostylous species have not yet been reported. This study aims to investigate enantiostyly in Chamaecrista flexuosa, in particular the presence of deviations from the standard form, in an area of coastal vegetation in north-east Brazil.
Methods
Observations and investigations of floral biology, the reproductive system, pollinator behaviour, floral morphology and morphometry were performed.
Key Results
In C. flexuosa flowers, anthers of different size but similar function are grouped. The flowers were self-compatible and set fruits after every treatment, except in the spontaneous self-pollination experiment, thereby indicating their dependence on pollen vectors. The flowers were pollinated by bees, especially Xylocopa cearensis and X. grisencens. Pollen is deposited and captured from the ventral portion of the pollinator's body. Variations in the spatial arrangement of floral elements allowed for the identification of floral morphs based on both morphological and functional criteria. Using morphological criteria, morphologically right (MR) and morphologically left (ML) floral morphs were identified. Three floral morphs were identified using functional criteria: functionally right (FR), functionally central (FC) and functionally left (FL). Combinations of morphologically and functionally defined morphs did not occur in equal proportions. There was a reduced frequency of the MR–FR combination.
Conclusions
The results indicate the occurrence of an atypical enantiostyly in C. flexuosa. This seems to improve reproductive success by increasing the efficiency of pollen deposition and capture.
Keywords: Breeding system, Chamaecrista flexuosa, Fabaceae, Caesalpinioideae, enantiostyly, floral polymorphism, pollen flow, reciprocal herkogamy
INTRODUCTION
Reciprocal herkogamy, represented, for example, by enantiostyly and heterostyly, is a floral polymorphism associated with incompatibility relationships between floral morphs (Barrett, 2002). In enantiostylous and heterostylous species, the floral morphs are reciprocal with respect to the relative positions of the style and anthers (Barrett, 2002). Enantiostylous species have two floral morphs. In one morph, the stigma is on the right hand side of the flower and the anthers on the left: these are called right flowers (R). Left flowers (L) have the reciprocal phenotype and therefore the two morphs represent mirror images (Barrett, 2002). There are two forms of enantiostyly: monomorphic where both floral morphs are present on the same individual; and dimorphic where floral morphs occur on different individuals. In both cases, the two floral morphs will occur at similar frequencies (isoplethy) (Barrett, 2002).
In functional terms, the two floral morphs are incompatible because pollen deposition and capture occur on different sides of the pollinator's body. In addition, enantiostylous species also have inter- and intramorph self-incompatibility mechanisms (Gottsberger and Silberbauer-Gottsberger, 1988; Delgado and Souza, 1997; Westerkamp, 2004; Laporta, 2005; Costa et al., 2007; Bezerra, 2008; Gómez, 2008; Krieck et al., 2008; Dutra et al., 2009; Almeida et al., 2013). The morphological, physiological and functional features described above indicate the existence of a pattern for this group of species.
In some cases of enantiostyly, especially in species of the Cassiinae sub-tribe, the reciprocity between floral morphs is also observed in the petals (Fenster, 1995; Jesson and Barrett, 2002, 2003; Arceo-Gomez et al., 2011). Modifications in the reciprocity of the sterile elements of the flowers apparently do not alter the functionality of floral morphs in the reproductive process and thus may be considered morphological. In the Cassiinae sub-tribe, species bearing pollen flowers are also commonly observed (Vogel, 1978). These flowers have poricidal anthers containing a high number of pollen grains (Faegri and Pijl, 1979). The pollen is a unique floral resource, which is generally collected through vibratory movements (buzz pollination; Buchmann, 1983) made by large bees (Vogel, 1978).
Alterations in heterostylous patterns, such as different proportions of floral morphs, impede incompatibility mechanisms, and partial or total elimination of one morph in the population is interpreted as a consequence of environmental disturbances and the absence or low efficiency of pollinators (Barrett, 2002). In reciprocal herkogamous species, deviations in the positioning of sexual structures may modify the functionality of floral morphs, as with some species presenting flexistyly and others presenting atypical heterostyly (Sobrevila et al., 1983; Hamilton, 1990; Richards and Koptur, 1993; Barrett, 2002; Krieck et al., 2008). For enantiostylous species, especially monomorphic species, these alterations have not yet been studied; therefore, it is difficult to discern the cause of the deviations in the reproductive patterns of these species.
In an area of costal vegetation in Brazil, individuals of the enantiostylous species Chamaecrista flexuosa presenting two floral morphs were observed. By studying the floral biology, pollinator behaviour and floral morphometrics, this study aimed to answer the following questions. (1) Does C. flexuosa have deviations from the enantiostylous pattern? (2) If deviations exist, do they alter the functionality of enantiostyly?
MATERIALS AND METHODS
Study area and species
The fieldwork was conducted at the Nossa Senhora do Oiteiro de Maracaípe Private Natural Heritage Reserve, Ipojuca, Pernambuco, Brazil (08 °31′48′′S, 35 °01′05′′W). The reserve covers 76·2 ha including 60·96 ha of restinga (sandy coastal plain) and 15·24 ha of mangrove vegetation (Almeida et al., 2009). The climate is warm and moist (As', sensu Köppen, 1948), annual average temperatures are 26 °C with annual rainfall average of approx. 2000 mm, which is distributed throughout the year with a higher intensity between May and June (Superintendência do Desenvolvimento do Nordeste, 1990; Inmet, 2005).
Chamaecrista flexuosa is an undershrub species and was found in five patches (hereafter called A, B, C, D and E) in areas of open vegetation within the reserve. Chamaecrista flexuosa flowers throughout the year, although more strongly between February and May. The flower has five petals, including one curved petal called the falcate (Fig. 1). The species is enantiostylous and produces three floral types: flowers with the stigma turned to the right, flowers with the stigma turned to the left and flowers with an intermediate condition. Individuals are monomorphic. Floral morphs were distinguished based on the position of the stigma relative to the centre of the falcate petal. Differences in the arrangement of the sexual elements will have direct functional effects with respect to pollen transfer, while those associated with the petals will be non-functional. Thus, floral morphs can be classified using both functional and non-functional or morphological criteria.
Fig. 1.

Right (R; bottom row) and left (L; top row) floral morphs of the enantiostylous species Chamaecrista flexuosa based on morphological (M) and functional (F) criteria in an area of coastal vegetation in north-east Brazil. (A) ML and FL flower; (B) ML and FC flower; (C) MR and FR flower; (D) MR and FC flower. Dots indicate the centre of the falcate petal, and the arrows indicate the stigma position. Scale bar = 0·3 cm.
The position of the falcate petal relative to the floral axis was used to define morphologically left (ML) and right (MR) flowers. This petal provides a convenient reference since it is opposite the gynoecium. When the falcate petal was to the left of the floral axis, the flower was classified as MR (Fig. 1C, D) and when this petal was to the right, the flower was considered as ML (Fig. 1A, B). The position of the stigma relative to the centre of the falcate petal was used to define functionally left (FL) and right (FR) flowers. When the stigma was to the right relative to the centre of the falcate petal, the flower was FR (Fig. 1C) and the distance to the centre of the falcate petal was considered to be positive. When the stigma was to the left, the flower was FL (Fig. 1A) and the distance was considered to be negative. Flowers with values between –1 and 1 mm were considered to be functionally central (FC) (Fig. 1B, D).
Floral biology
For each floral morph, we counted the number of flower parts and examined the timing of anthesis, stigmatic receptivity (using the peroxidase activity technique; Kearns and Inouye, 1993) and the presence and location of osmophores (using neutral red solution; Vogel, 1990). Data were collected from ten flowers of each morph; flowers were selected from each of the five patches and from distinct individuals within each patch.
Mating system
The mating system of C. flexuosa was investigated by measuring fruit set following a series of contrasting pollination treatments. Self-incompatibility was assessed using a self-pollination treatment in which flowers were hand pollinated with pollen from their own anthers. We also tested for spontaneous self-pollination by bagging pre-anthesis buds. Intra- and intermorph incompatibility were assessed using hand pollinations between flowers of the same (R × R, L × L and C × C) and different morphs (R × L, R × C, L × R, L × C, C × R and C × L; the first letter indicates the pollen donor and the second the pollen recipient). For controlled self- and cross-pollinations, flowers were bagged at the pre-anthesis stage and again after treatment. Fruit set after natural (open) pollination was estimated in tagged flowers. Sample sizes for each treatment are described in Table 1.
Table 1.
The mating system of Chamaecrista flexuosa at five sites (approx. 200 individuals) located in an area of coastal vegetation in noth-east Brazil. Values are expressed as the number of flowers/fruit set (%)
| Treatment | R | L | C |
|---|---|---|---|
| Control | 36/44·4 | 43/48·8 | 28/64·2 |
| Spontaneous self-pollination | 43/0 | 38/0 | 47/0 |
| Self-pollination | 22/45·4 | 24/66·6 | 20/60·0 |
| Cross-pollination | ||||||||
| R × R | R × L | R × C | L × R | L × L | L × C | C × R | C × L | C × C |
| 24/83·3 | 31/51·6 | 27/48·1 | 23/43·4 | 30/60·0 | 28/50·0 | 32/37·5 | 24/37·5 | 40/45·0 |
R, right flowers; L, left flowers; C, central flowers.
In cross-pollination experiments, the first letter represents the pollen donor and the second letter represents the pollen recipient.
Floral visitor behaviour
The frequency of floral visits was recorded during focal observations over four non-consecutive days between March and May 2009. Total observation time was 28 h. Focal observations (between March 2009 and August 2010) were also used to assess visitor behaviour, including means of arrival, contact with sexual elements, reward collection strategy and result of the visit (i.e. pollination if the stigma and anthers were both contacted or pillage if no contact was made or only the anthers were contacted).
Floral visitors were collected and identified by the first author as described by Silveira et al. (2002) and were deposited in the collection of the Laboratório de Ecologia Reprodutiva de Angiospermas of the Universidade Federal Rural de Pernambuco. The ventral portion width (thorax and abdomen) of the most frequent floral visitors was measured in the field, using a digital caliper (error: 0·01 mm), for comparison with the width of the falcate petal.
Occurrence and proportion of floral morphs
The proportion of morphological and functional morphs within individual patches was estimated using a total of 355 open flowers (patch A, 15 flowers; B, 98 flowers; C, 87 flowers; D, 43 flowers; and E, 112 flowers). The width of the falcate petal was also measured for each flower.
Statistical analysis
Fruit set between experiments concerning the reproductive system were compared using the χ2 test, and the morph ratio was assessed using a G-test; both were performed with BioEstat 5·0 software (Ayres et al., 2007).
RESULTS
Floral biology and breeding system
Chamaecrista flexuosa is an enantiostylous species with pentamerous flowers that are asymmetric with a dialisepalous, green calyx and a dialipetalous, yellow corolla (Fig. 1A–D). The androecium includes three groups of stamens with different anther sizes, which are positioned in the flower centre: small (five stamens), medium (two stamens) and large (four stamens). The gynoecium has a superior ovary with a curved style and a stigma that is positioned in the centre of the stamens and does not have any variability in its position within an individual flower. The neutral red solution did not indicate any areas containing odour glands. Anthesis was initiated at approx. 0430 h, when the stigma was receptive and the pollen was available. Anthesis ended at approx. 0830 h; when petal senescence began, the petal colours changed to orange, the stigma was not receptive and pollen was not available.
In pollination experiments, fruits were set under all treatments except spontaneous self-pollination (Table 1). There were no significant differences in fruit and seed for intramorph (R × R–L × L, P = 0·344; R × R–C × C, P = 0·263; and L × L–C × C, P = 0·528) or intermorph crosses (R × L–L × R, P = 0·315; R × C–C × R, P = 0·617; and L × C–C × L, P = 0·475). In the control group, the C morph exhibited higher fruit set than either the R or L morphs, although this difference was not statistically significant (Table 1).
Floral visitors
Chamaecrista flexuosa flowers were visited by six Apidae bee species: Xylocopa (Neoxylocopa) cearensis, X. (Neoxylocopa) carbonaria Smith, 1854, X. (Neoxylocopa) grisescens, Centris sp., Florilegus (Euflorilegus) sp. and Eufriesea sp. Xylocopa cearensis was the most frequent visitor (Fig. 2) representing 30 % of the total. However, all six visitors appear to be pollinators since all contacted both the stigma and the anthers, from which pollen was collected.
Fig. 2.
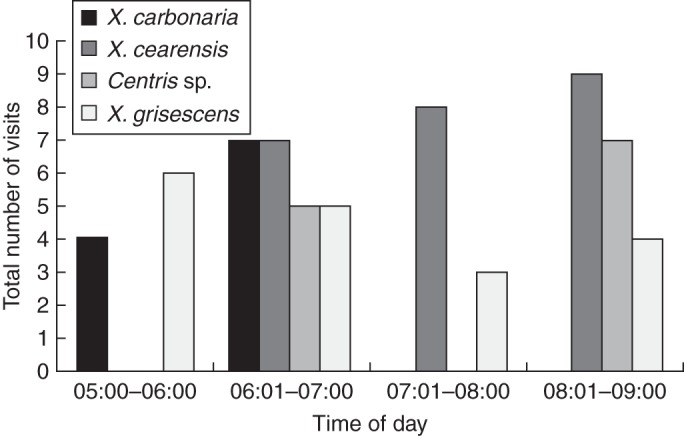
Frequency of visits of the main pollinators to flowers of Chamaecrista flexuosa during four non-consecutive days between March and May 2009 at five sites located in an area of coastal vegetation in north-east Brazil. X. = Xylocopa (Neoxylocopa).
All visitor species used the falcate petal to land on the flower and to gain access to the stamens. Once on the stamens, most visitors oriented their bodies so that the posterior portion (i.e. thorax and abdomen) was in close proximity to the anther pores. Visitors would then make vibratory movements that resulted in the deposition of a belt of pollen on the anterior portion of the thorax (Fig. 3). Given the stigma position and the mode of arrival on flowers, contact with and pollen capture by the stigma occurred prior to the vibratory movements. Unlike other visitors, the smaller Florilegus sp. collected pollen directly from the anthers, storing it on the posterior legs.
Fig. 3.
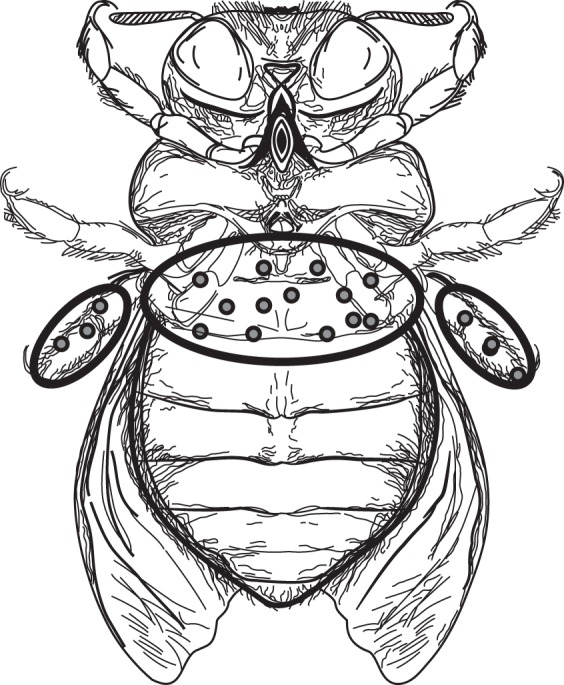
Areas of pollen deposition (dots) on bees that visited Chamaecrista flexuosa flowers in an area of coastal vegetation in north-east Brazil. The species represented is Xylocopa (Neoxylocopa) cearensis.
With the exception of Florilegus sp., which is a small species, the thorax and abdomen width of visiting species ranged from 0·8 to 1·2 cm. This size range falls within that of the width of the falcate petal (1·0–1·2 cm) and is close to the maximum distance of the stigma from the centre of this petal (1·0 cm).
Occurrence and proportion of floral morphs
Similar patterns regarding the proportion of floral morphs were found using the morphological and functional criteria. A higher proportion of ML flowers was observed when all of the patches were considered, but similar proportions of ML and MR flowers were found when the patches were considered separately (Table 2; Fig. 4). Similar patterns were found using the functional criterion: a higher proportion of FL floral morphs was observed (46·48 % of the total) when all the patches were considered (followed by FC, 34·93 %; and FR, 18·59 %, Table 3; Fig. 5), and similar proportions of the three floral morphs were observed when the patches were analysed separately (Table 3). The following P-values were found after performing comparisons between floral morphs (considering all patches): FL and FR, <0·0001; FL and FC, 0·019; and FR and FC, <0·0001.
Table 2.
The occurrence of floral morphs (using the morphological criterion) of Chamaecrista flexuosa, collected during one day at five sites (approx. 200 monomorphic individuals) located in an area of coastal vegetation in north-east Brazil
| Patch and floral morph |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | Total | |||||||
| ML | MR | ML | MR | ML | MR | ML | MR | ML | MR | ML | MR | |
| Number of flowers | 7 | 8 | 56 | 42 | 48 | 39 | 22 | 21 | 65 | 47 | 198 | 157 |
| P-value | 0·796 | 0·189 | 0·391 | 0·879 | 0·108 | 0·034 | ||||||
The P-value is for the comparison between morphs within each site
ML, morphologically left flower; MR, morphologically right flower.
Fig. 4.
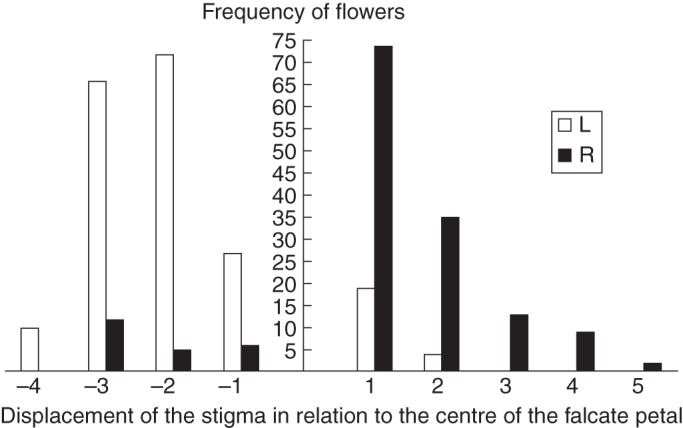
Number of floral morphs (using the morphological criterion) and the positioning of the stigma in relation to the falcate petal of flowers of Chamaecrista flexuosa in an area of coastal vegetation in north-east Brazil. L, R, left and right flowers, respectively.
Table 3.
The occurrence of floral morphs (using the stigma positioning; functional criterion) in Chamaecrista flexuosa at five sites (A–E) in an area of coastal vegetation in north-east Brazil.
| Patch and floral morph |
||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | Total | |||||||||||||
| FL | FR | FC | FL | FR | FC | FL | FR | FC | FL | FR | FC | FL | FR | FC | FL | FR | FC | |
| Number of flowers | 4 | 8 | 3 | 47 | 15 | 36 | 2 | 13 | 32 | 19 | 10 | 14 | 53 | 20 | 39 | 165 | 66 | 124 |
| FL × FR | 0·3865 | <0·0001 | 0·0006 | 0·1374 | 0·0002 | <0·0001 | ||||||||||||
| FR × FC | 0·2278 | 0·0051 | 0·0196 | 0·5403 | 0·0769 | <0·0001 | ||||||||||||
| FL × FC | 0·4876 | 0·2724 | 0·2955 | 0·4862 | 0·0536 | 0·0186 | ||||||||||||
The table presents P-values for the comparison between morphs within each site. FL, functionally left flower; FR, functionally right flower; FC, functionally central flower.
Fig. 5.
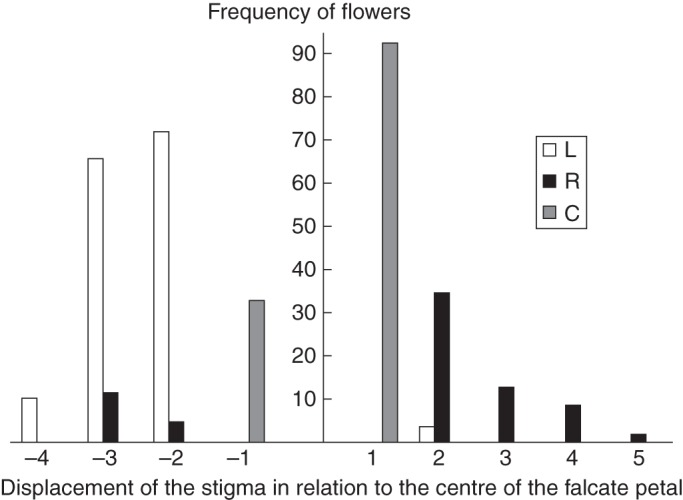
Occurrence of the floral morphs based on stigma positioning (functional criterion) and the positioning of the stigma in relation to the falcate petal of Chamaecrista flexuosa in an area of coastal vegetation in north-east Brazil. L, R, C: left, right and central flowers, respectively.
DISCUSSION
Features such as yellow floral parts, poricidal anthers, the absence of nectaries and the use of pollen as the floral resource are common in the sub-tribe Cassiinae (Gottsberger and Silberbauer-Gottsberger, 1988). These features suggest that C. flexuosa produces mellitophilous, pollen flowers (Vogel, 1978; Faegri and van der Pijl, 1979; Buchmann, 1983).
Stamen heteromorphism is a feature observed in some species with pollen flowers (Carvalho and Oliveira, 2003; Hrycan and Davis, 2005). Chamaecrista flexuosa has stamens of different length. However, all appear to be functionally similar since all the pollen is deposited in the same region of the visitor's body. In other studies of Caesalpinioideae that exhibit stamen heteromorphism, the pollen was deposited in distinct regions, suggesting that stamens of different length are functionally different (Vogel, 1978; Westerkamp, 2004; Laporta, 2005; Costa et al, 2007; Gómez, 2008; Dutra et al., 2009).
A few studies have suggested that incompatibility mechanisms are present in Caesalpinioideae (Arroyo, 1981; Owens and Lewis, 1989; Carvalho and Oliveira, 2003; Leite and Machado, 2010). However, incompatibility seems to have been lost from the herbaceous and sub-shrub of the group (Arroyo, 1981). Consistent with this observation and studies on other Cassiinae (Dulberger, 1981; Liu and Koptur, 2003; Ribeiro and Lovato, 2004; Laporta, 2005; Costa et al., 2007; Gómez, 2008), C. flexuosa exhibits both self- and intramorph compatibility. Based on these observations, C. flexuosa appears to have a pattern of enantiostyly similar to that previously described for the group.
Monomorphic, enantiostylous species commonly exhibit morphological and functional dimorphism (Jesson and Barrett, 2002, 2003) and isoplethy (Jesson and Barrett, 2003; Laporta, 2005). However, C. flexuosa represents an exception to this basic pattern because there are three recognizable stigma positions and the proportions of each floral morph differ. In addition to the basic pattern of left and right functional forms, C. flexuosa also has a central morph in which the stigma is located close to the anthers. This stigma position would appear to favour spontaneous self-pollination in the FC morph. Consistent with this expectation, the FC morph exhibits higher fruit set relative to the other functional morphs (FR and FL). However, our experiments suggest that this species does not self in the absence of pollinators. Instead, selfing requires the vibratory movements of visitors to release the pollen (Buchmann, 1983).
Visitors to C. flexuosa flowers are predominantly large bee species. This appears to be a common trend for restinga species (Oliveira-Rebouças and Gimenes, 2004). Strong winds are typical of this ecosystem and would tend to favour larger species (Gottsberger and Silbebauer-Gottsberger, 1988; Viana and Pinheiro, 1998). Beyond this, the close correspondence between the width of the principal pollinators (X. cearensis, X. carbonaria, X. grisescens and Centris sp.) and that of the falcate petal, which is used as a landing platform, suggests a direct relationship between these large bees and C. flexuosa.
The influence of the falcate petal on pollen transfer has been described in several studies of Cassiinae (Gottsberger Silbebauer-Gottsberger, 1988; Delgado and Souza, 1997; Westerkamp, 2004; Laporta, 2005; Costa et al., 2007; Dutra et al., 2009). However, in contrast to other enantiostylous species, C. flexuosa floral morphs deposit pollen in the same broad region of the pollinator's body rather than distinct zones. In this case, the close size relationship between the falcate petal and floral visitors may help maximize the overall efficiency of pollination because the falcate petal will direct the stamens to areas of the visitor's body that will later have contact with the stigma. Small bee species will be less effective pollinators because they fail to contact both the anthers and the stigma. In addition, small species may be poor pollinators because they fail to generate strong enough vibratory movements to release pollen.
There are several similarities between enantiostyly and heterostyly, a strategy in which floral morphs are primarily distinguished by differences in the height of the sexual elements (Ganders, 1979; Barrett, 2002). For example, as in enantiostylous species, heterostylous plants commonly exhibit isoplethy (Ganders, 1979; Barrett and Richards, 1990). In heterostyly, deviations from the basic pattern are well described and these species are considered to be variants or atypical (Sobrevila et al., 1983; Hamilton, 1990; Richards and Koptur, 1993; Barrett, 2002). Barrett (2002) has suggested that such deviations may reflect environmental disturbances, low pollinator efficiency or absence and the loss of a floral morph from the population. As far as we know, no studies have described morpho-functional deviations for enantiostylous species or suggested a relationship between such deviations and the environment.
We consider this report to be the first description of atypical enantiostyly. In the case of C. flexuosa, atypical enantiostyly may increase the efficiency of pollen transfer between the anthers and the stigma. The juxtaposition of the anthers and falcate petal results in pollen deposition across a belt-like region to the pollinator's body, while the polymorphic stigma position favours pollen capture from different parts of this region. This system seems likely to maximize the reproductive fitness of the species.
ACKNOWLEDGEMENTS
We thank the RPPN Nossa Senhora do Outeiro de Maracaípe for logistic support. This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil) for the grants awarded to N.M.deA. and a Research Productivity grant to I.C.M., and by the Programa de Pós-graduação em Botânica (Universidade Federal Rural de Pernambuco).
LITERATURE CITED
- Almeida EB, Jr, Olivo MA, Araújo EL, Zickel CS. Caracterização da vegetação de restinga da RPPN de Maracaípe, PE, Brasil, com base na fisionomia, flora, nutrientes do solo e lençol freático. Acta Botanica Brasilica. 2009;23:36–48. [Google Scholar]
- Almeida NM, Castro CC, Leite AV, Novo RR, Machado IC. Enantiostyly in Chamaecrista ramosa (Fabaceae-Caesalpinioideae): floral morphology, pollen transfer dynamics and breeding system. Plant Biology. 2013;15:369–375. doi: 10.1111/j.1438-8677.2012.00651.x. [DOI] [PubMed] [Google Scholar]
- Arceo-Gomez G, Martínez ML, Parra-Tabla V, Garcia-Franco JG. Anther and stigma morphology in mirror-image flowers of Chamaecrista chamaecristoides (Fabaceae): implications for buzz pollination. Plant Biology. 2011;13:19–24. doi: 10.1111/j.1438-8677.2010.00324.x. [DOI] [PubMed] [Google Scholar]
- Ayres M, Ayres M, Jr, Ayres DL, Santos ASS. BioEstat: aplicações estatísticas nas áreas das ciências biomédicas. Belém, Brazil: Sociedade Civil Mamirauá; 2007. [Google Scholar]
- Arroyo MTK. Breeding systems and pollination biology in Leguminosae. In: Polhill RM, Raven PH, editors. Advances in legume systematics (Part 2). Kew, London: Royal Botanic Gardens; 1981. pp. 723–769. [Google Scholar]
- Barrett SCH. The evolution of plant sexual diversity. Nature Reviews Genetics. 2002;3:274–284. doi: 10.1038/nrg776. [DOI] [PubMed] [Google Scholar]
- Barrett SCH, Richards JH. Heterostyly in tropical plants. Memoirs of the New York Botanical Garden. 1990;55:35–61. [Google Scholar]
- Bezerra ELS. Guilda de flores de óleo do Parque Nacional do Catimbau: fenologia, polinização e sistema reprodutivo. PhD Thesis: Universidade Federal de Pernambuco, Brazil; 2008. [Google Scholar]
- Buchmann SL. Buzz pollination in angiosperms. In: Jones CE, Little RJ, editors. Handbook of experimental pollination biology. New York: Van Nostrand Reinhold; 1983. pp. 73–113. [Google Scholar]
- Carvalho DA, Oliveira PE. Biologia reprodutiva e polinização de Senna sylvestris (Vell.) H. S. Irwin and Barneby (Leguminosae, Caesalpinioideae) Revista Brasileira de Botânica. 2003;26:319–328. [Google Scholar]
- Costa CBN, Lambert SM, Borba EL, Queiroz LP. Post zygotic reproductive isolation between sympatric taxa in the Chamaecrista desvauxii complex (Leguminosae-Caesalpinioideae) Annals of Botany. 2007;99:625–635. doi: 10.1093/aob/mcm012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado SA, Souza SM. Biologia floral do gênero Cassia em la region de Los Tuxtlas, Vera Cruz. Boletín de la Sociedad Botánica de México. 1997;37:5–45. [Google Scholar]
- Dulberger R. The floral biology of Cassia didymobotrya and C. auriculata (Caesalpiniaceae) American Journal of Botany. 1981;68:1350–1360. [Google Scholar]
- Dutra VF, Vieira MF, Garcia FCP, Lima HC. Fenologia reprodutiva, síndromes de polinização e dispersão em espécies de Leguminosae dos campos rupestres do Parque Estadual do Itacolomi, Minas Gerais, Brasil. Rodriguesia. 2009;60:371–387. [Google Scholar]
- Faegri K, van der Pijl L. The principles of pollination ecology, 3rd edn. London: Pergamon Press; 1979. [Google Scholar]
- Fenster CB. Mirror image and their effect on outcrossing rate in Chamaecrista fasciculata (Leguminosae) American Journal of Botany. 1995;82:46–50. [Google Scholar]
- Ganders FR. The biology of heterostyly. New Zealand Journal of Botany. 1979;17:607–635. [Google Scholar]
- Gómez GA. Biología reproductiva de Chamaecrista chamaecristoides (Fabaceae), especie enantiostilica de las dunas costeras. Masters thesis, Instituto de Ecologia, México. 2008 [Google Scholar]
- Gottsberger G, Silberbauer-Gottsberger I. Evolution of flower structures and pollination in Neotropical Cassiinae (Caesalpiniaceae) species. Phyton. 1988;28:293–320. [Google Scholar]
- Hamilton CW. Variations on a distylous theme in Mesoamerican Psychotria subgenus Psychotria (Rubiaceae) Memoirs of the New York Botanical Garden. 1990;55:65–75. [Google Scholar]
- Hrycan WC, Davis AR. Comparative structure and pollen production of the stamens and pollinator-deceptive staminodes of Commelina coelestis and C. dianthifolia (Commelinaceae) Annals of Botany. 2005;95:1113–1130. doi: 10.1093/aob/mci134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- INMET (Instituto Nacional de Meteorologia) 2005 Available at www.inmet.gov.br. (accessed 20 November 2009) [Google Scholar]
- Jesson LK, Barrett SCH. Solving the puzzle of mirror-image flowers. Nature. 2002;417:707. doi: 10.1038/417707a. [DOI] [PubMed] [Google Scholar]
- Jesson LK, Barrett SCH. The comparative biology of mirror-image flowers. International Journal of Plant Sciences. 2003;164:237–249. [Google Scholar]
- Kearns CA, Inouye DW. Techniques for pollination biologists. Niwot: University of Colorado Press; 1993. [Google Scholar]
- Köppen W. Climatologia: un estudio de los climas de la tierra. Ciudad del Mexico: Fondo de Cultura Económica; 1948. [Google Scholar]
- Krieck C, Finatto T, Müller TS, Guerra MP, Orth AI. Biologia reprodutiva de Alpinia zerumbet (Pers.) B.L.Burtt and R.M.Sm. (Zingiberaceae) em Florianópolis, Santa Catarina. Revista Brasileira de Plantas Medicinais. 2008;10:103–110. [Google Scholar]
- Laporta C. Floral biology and reproductive system of enantiostylous Senna corymbosa (Caesalpiniaceae) Revista de Biologia Tropical. 2005;53:49–61. [PubMed] [Google Scholar]
- Leite AV, Machado IC. Reproductive biology of woody species in caatinga, a dry forest of northeastern Brazil. Journal of Arid Environments. 2010;74:1374–1380. [Google Scholar]
- Liu H, Koptur S. Breeding system and pollination of a narrowly endemic herb of the Lower Florida Keys: impacts of the urban–wildland interface. American Journal of Botany. 2003;90:1180–1187. doi: 10.3732/ajb.90.8.1180. [DOI] [PubMed] [Google Scholar]
- Oliveira-Rebouças P, Gimenes M. Abelhas (Apoidea) visitantes de flores de Comolia ovalifolia DC Triana (Melastomataceae) em uma área de restinga na Bahia. Neotropical Entomology. 2004;33:315–320. [Google Scholar]
- Owens SJ, Lewis GP. Taxonomic and functional implications of stigma morphology in species of Cassia, >Chamaecrista, and Senna (Leguminosae: Caesalpinioideae) Plant Systematics and Evolution. 1989;163:93–105. [Google Scholar]
- Ribeiro RA, Lovato MB. Mating system in a neotropical tree species, Senna multijuga (Fabaceae) Genetic Molecular Biology. 2004;27:418–424. [Google Scholar]
- Richards JH, Koptur S. Floral variation and distyly in Guettarda scabra (Rubiaceae) American Journal of Botany. 1993;80:31–40. [Google Scholar]
- Silveira FA, Melo GAR, Almeida EAB. Abelhas Brasileiras: sistemática e identificação. Belo Horizonte, Brazil: Fundação Araucária; 2002. [Google Scholar]
- Sobrevila C, Ramirez N, Enrech NX. Reproductive biology of Palicourea fendleri and P. petiolares (Rubiaceae), heterostylous shrubs of a tropical cloud forest in Venezuela. Biotropica. 1983;15:161–169. [Google Scholar]
- Superintendência do Desenvolvimento do Nordeste. Dados pluviométricos do Nordeste – Pernambuco. Série pluviométrica. Recife, Brazil: Superintendência do Desenvolvimento do Nordeste; 1990. [Google Scholar]
- Viana VM, Pinheiro LAFV. Conservação da biodiversidade em fragmentos florestais. Série Técnica IPEF. 1998;12:25–42. [Google Scholar]
- Westerkamp C. Ricochet pollination in Cassias – and how bees explain enantiostyly. In: Magalhães FB, Pereira JO, editors. Solitary bees: conservation, rearing and management for pollination. Fortaleza, Brazil: Universidade Federal do Ceará; 2004. pp. 225–230. [Google Scholar]
- Vogel S. Evolutionary shifts from reward to deception in pollen flowers. In: Richards AJ, editor. The pollination of flowers by insects. London: Academic Press; 1978. pp. 89–96. [Google Scholar]
- Vogel S. The role of scent glands in pollination. Rotterdam: A.A. Balkema; 1990. [Google Scholar]