

Abstract
Background
Positron emission tomography research has shown that dopamine D2/3 receptor (D2/3R) availability is negatively correlated with body mass index (BMI) in obese but not in healthy subjects. However, previous positron emission tomography studies have not looked specifically at the ventral striatum (VS), which plays an important role in motivation and feeding. Furthermore, these studies have only used antagonist radiotracers. Normal-weight rats given free access to high-fat diets demonstrate behavioral sensitization to D2/3R agonists but not to antagonists. Sensitization is associated with increased D2/3R affinity, which affects binding of agonists but not antagonists.
Methods
We examined the association between BMI within the nonobese range (18.6–27.8) and D2/3R availability in the VS with the use of the agonist radiotracer [11C]-(+)-PHNO (n = 26) and the antagonist [11C]-raclopride (n = 35) in healthy humans.
Results
In the VS, we found a positive correlation between BMI and [11C]-(+)-PHNO binding but no relationship with [11C]-raclopride binding. Secondary analyses revealed no relationship between BMI and binding in the dorsal striatum with either radiotracer.
Conclusions
We propose that in nonobese individuals, higher BMI may be associated with increased D2R affinity in the VS. This increased affinity may potentiate the incentive salience of food cues and counteract the effects of satiety cues, thereby increasing feeding.
Keywords: Body mass index, dopamine D2 receptor, food addiction, obesity, PET, ventral striatum
Obesity is one of the leading causes of preventable death, reaching pandemic levels in the United States and affecting 35.7% of adults and 17% of youth (1). A growing perspective conceptualizes overeating as a food addiction. Evidence suggests that striatal dopamine, involved in reward, motivation, and food consumption, is altered in obesity (2). Addiction-like dopaminergic dysfunction, specifically reduced striatal dopamine D2/3 receptor (D2/3R) availability, has been observed in rat models of obesity (3,4) and in obese humans in vivo (5–8).
A positron emission tomography (PET) study with the use of the antagonist radiotracer [11C]-raclopride found that lower striatal D2/3R availability predicted higher body mass index (BMI) in severely obese individuals but not in nonobese subjects (5). This is contrary to findings in nonobese rats given free access to regular chow, in which lower [11C]-raclopride binding in the ventral striatum (VS) predicted both greater body weight and preference for cocaine (9).
The VS, including the nucleus accumbens, plays an integral role in processing reward cues and motivating behavior to seek rewards such as palatable foods (2). Thus, changes in D2/3R availability in the VS may alter the rewarding properties and consumption of food, affecting body weight. Left VS activation in response to food cues predicts weight gain in healthy females (10) and correlates with dopamine release in response to reward cues (11). These studies suggest that VS activation and D2/3R availability may show changes related to normal BMI.
Previous PET studies of BMI have not specifically examined D2/3R availability in the VS; instead, region of interest (ROI) analyses of the whole striatum (5), the caudate and putamen (6,7), or a voxel-based approach (7) were used. Furthermore, previous PET studies have only used the D2/3R antagonist radio-tracer [11C]-raclopride. Normal-weight rats given free access to high-fat diets demonstrate behavioral sensitization to direct and indirect D2/3R agonists but not antagonists (12). This sensitization is also observed in rodent models of drug addiction (13) and is associated with increased D2R affinity (14–16).
This suggests that, like cocaine and amphetamine, exposure to high-fat foods may increase the affinity for dopamine at D2Rs. It has been observed in vitro that agonist radiotracers are more sensitive to changes in D2R affinity than are antagonist radio-tracers. Increased D2R affinity, indexed by increased agonist radiotracer binding, has been found to co-occur with no change and even decreases in total D2R binding sites given amphet-amine sensitization (14). Consequently, differences in BMI within the normal range may be related to differences in VS binding of dopamine agonists but not antagonists.
This study investigated the relationship between healthy BMI and D2/3R availability in the VS in humans, with the use of both the agonist radiotracer [11C]-(+)-PHNO and the antagonist [11C]-raclopride. Understanding the dopaminergic correlates of normal BMI will help elucidate the deficits seen in obesity and may inform current models of food addiction as well as the development of novel prevention and treatment strategies.
Methods and Materials
Subjects
All participants were right-handed and free of any major medical or psychiatric disorder as determined by clinical interview, the Mini-International Neuropsychiatric Interview, basic laboratory tests, and electrocardiography. Although obesity was not an exclusion criteria, given our exclusion of major medical conditions (such as diabetes or heart disease), we sampled only persons within a normal BMI range (<30). Participants were required to have a negative urine screen for drugs of abuse and/ or pregnancy at inclusion and before each PET scan. Participants were also asked to abstain from alcohol or caffeine 3 days before the PET scans. Only data collected from nonsmoking participants was analyzed for this study. The sample analyzed for the current study was collected by our laboratory from various PET studies that were approved by the Research Ethics Board of the Centre for Addictions and Mental Health, Toronto. All participants provided written informed consent.
PET Imaging
The radiosynthesis of [11C]-(+)-PHNO and [11C]-raclopride and the acquisition of PET images have been described in detail elsewhere (17–19). Briefly, images were acquired with the use of a high–resolution, head-dedicated PET camera system (CPS-HRRT; Siemens Molecular Imaging, Munich, Germany), which measures radioactivity in 207 brain slices with a thickness of 1.2 mm each. The in-plane resolution was ~2.8 mm full-width at half-maximum. Transmission scans were acquired with the use of a 137Cs (T1/2 = 30.2 years, energy = 662 KeV) single-photon point source to provide attenuation correction, and the emission data were acquired in list mode. The raw data were reconstructed by filtered back-projection. The mean radioactivity dose of [11C]-(+)-PHNO (n = 26) was 8.96 (±1.68) mCi, with a specific activity of 1009.44 (±289.35) mCi/μmoL. The mean radioactivity dose of [11C]-raclopride (n = 35) was 9.22 (±2.49) mCi, with a specific activity of 1133.39 (±433) mCi/μmoL. [11C]-(+)-PHNO scanning data were acquired for 90 min after injection. Once scanning was complete, the data were re-defined into 30 frames (1–15 of 1–min duration and 16–30 of 5–min duration). [11C]-raclopride data were acquired for 60 min and redefined into 28 frames (1–5 of 1-min duration, 6–25 of 2-min duration, and 26–28 of 5-min duration).
Image Analysis
The ROI-based analysis for [11C]-(+)-PHNO and [11C]-raclopride has been described in detail elsewhere (20). Briefly, time-activity curves (TACs) from ROIs were obtained from the dynamic PET images in native space with reference to each subject’s coregistered magnetic resonance image (MRI). The coregistration of each subject’s MRI to PET space was performed with the use of the normalized mutual information algorithm (21) as implemented in SPM2 (SPM2, The Wellcome Department of Cognitive Neurology, London; http://www.fil.ion.ucl.ac.uk/spm). The TACs were analyzed by means of the Simplified Reference Tissue Method (SRTM) (22), with the cerebellum used as the reference region, to derive a quantitative estimate of binding: binding potential nondisplaceable (BPND). The basis function implementation of the SRTM (23) was applied to the dynamic PET images to generate parametric voxelwise BPND maps by means of PMOD (v2.7; PMOD Technologies, Zurich, Switzerland). These images were spatially normalized into Montreal Neurological Institute (MNI) brain space by nearest neighbour interpolation with a voxel size fixed in 2 × 2 × 2 mm3 by means of SPM2. Regional BPND estimates were then derived from ROIs defined in MNI space. The ventral striatum and dorsal striatum (dorsal caudate, hereafter caudate; dorsal putamen, hereafter putamen) were defined according with Mawlawi et al. (24). The definition was made over the participant’s MRI slices oriented in the coronal plane. The VS (inferiorly), caudate, and putamen (superiorly) were defined by a line joining the intersection between the outer edge of the putamen with a vertical line going through the most superior and lateral point of the internal capsule and the center of the portion of the anterior commissure (AC). This line was extended to the internal edge of the caudate. The other boundaries of the VS were visually determined by its dense gray signal and were easily distinguishable from the adjacent structures. The VS was sampled from the anterior boundary of the striatum to the level of the AC coronal plane. The caudate also was sampled from its anterior boundary to the AC coronal plane. Thus, for the VS, the sampled region included the ventral and rostral part of the striatum, with reference to AC having the brain horizontal to the AC-PC line. For the caudate, the sampled region included the dorsal part of the head of the caudate and the anterior third of the body of the caudate. The putamen was sampled from its anterior to posterior boundaries in slices posterior to the AC plane. For [11C]-raclopride scans, BPND in the substantia nigra ROI was not obtainable because binding in this region falls within noise levels (20).
Statistical Analysis
Statistical analyses were conducted with the use of SPSS (v.12.0; SPSS, Chicago, Illinois) and GraphPad (v.5.0; GraphPad Software, La Jolla California). Pearson product-moment correlation coefficients were calculated to examine the relationship between BMI and BPND in the ROIs. Normality of variables was determined by means of the D’Agostino-Pearson test. Student t test and Fisher’s exact test were used where appropriate. The significance level for all testes was set at p < .05 (two-tailed).
Results
Data from 46 healthy volunteers were analyzed, some of which have been reported previously (20,25,26). Twenty-six subjects were scanned with [11C]-(+)-PHNO and 35 subjects were scanned with [11C]-raclopride. A subgroup of these subjects (n = 15) were scanned with both radiotracers in a counterbalanced order, at least 3 hours apart. BMI was calculated as kg/m2 (Table 1). There was no difference in the time of day at which the [11C]-(+)-PHNO and [11C]-raclopride scans were acquired, neither for the full samples (t59 = .16, p = .87) nor for the subsample scanned with both tracers (t28 = .97, p = .34). Within the full sample of persons scanned with [11C]-(+)-PHNO, BMI was not related to age (r = .27, p = .18) nor differed by sex (t24 = .42, p = .66). Within the full sample of persons scanned with [11C]-raclopride, BMI was not related to age (r = .21, p = .23) nor differed by sex (t33 = .21, p = .84).
Table 1.
Participant Demographics
| [11C]-(+)-PHNO (n = 26) | [11C]-Raclopride (n = 35) | [11C]-(+)-PHNO and [11C]-Raclopride (n = 15) | |
|---|---|---|---|
| Age | |||
| Mean (SD) | 29.88 (7.43) | 31.31a (9.02) | 29.67 (6.81) |
| Range | 20–45 | 20–47 | 20–45 |
| Sex | |||
| M (F) | 19 (7) | 21b (14) | 11 (4) |
| Body Mass Index | |||
| Mean (SD) | 23.66 (2.99) | 23.24c (2.71) | 23.32 (3.13) |
| Range | 18.61–27.77 | 18.61–27.83 | 18.61–27.77 |
Excluding the subjects scanned with both tracers (n = 15), the subjects scanned with [11C]-raclopride only (n = 20) did not differ from those scanned with [11C]-(+)-PHNO only (n = 11) in aage (p = .28), bsex (p = .46), or cbody mass index (p = .89).
The BPND of [11C]-(+)-PHNO in the VS was significantly correlated with BMI (r = .51, p = .008) in the full sample (n = 26) (Figure 1). This corresponded to a large effect size (27), with a shared variance of 26% (r2 = .26). Neither age (r = .14, p = .50) nor sex (r = .02, p = .92) was related to BPND in the VS. Given potential hemisphere differences (10,11), we tested for a hemisphere effect. Whereas BMI was correlated with BPND in the left (r = 40, p = .04) and right (r = .58, p = .002) hemispheres, a dependent-correlations t test revealed that the correlation was stronger in the right hemisphere (t23 = −2.01, p < .05) (Figure 2). Secondary analyses revealed that BMI was not correlated with BPND in the caudate (r = .21, p = .31), putamen (r = .30, p = .14), globus pallidus (r = −.06, p = .79), or substantia nigra (r = .31, p = .13). Although the VS was our a priori ROI, it is noteworthy that the relationship between BMI and BPND in the VS survived correction for multiple comparisons. There are five ROIs in total: the ventral striatum, caudate, putamen, globus pallidus, and substantia nigra. Thus, the Bonferroni-corrected significance threshold for the [11C]-(+)-PHNO–BMI correlations would be p = .01 (.05/5 = .01). Controlling for age or sex did not significantly change our results with [11C]-(+)-PHNO (data not shown).
Figure 1.

Correlation between body mass index (BMI) and [11C]-(+)-PHNO binding potential nondisplaceable (BPND) in the ventral striatum in the full sample of subjects (n = 26).
Figure 2.

The average [11C]-(+)-PHNO binding potential nondisplaceable (BPND) brain maps for persons within the first quartile of body mass index (BMI) (n = 7) and those within the fourth quartile of BMI (n = 7). The range of BMI for the quartiles is as follows: first = 18.6–21.5, fourth = 26.6–28. Persons within the fourth quartile of BMI had significantly greater [11C]-(+)-PHNO BPND in the ventral striatum compared with persons within the first quartile (t12 = 2.213, p = .047).
With [11C]-(+)-PHNO, side effects such as nausea have been observed with an injected mass >3 μg (28). Although all our subjects were scanned with an injected mass <3 μg (2.26 ± .36), we wanted to rule out the possibility that our findings were caused by tracer dose. There was no relationship between the injected mass (μg) and BPND in the VS (r = .14, p = .51; right hemisphere: r = .12, p = .58; left hemisphere: r = .15, p = .48) or with BMI (r = .01, p = .96). Neither the specific activity (mCi/μmol) nor amount injected (mCi) of [11C]-(+)-PHNO was related to BPND in the VS (r = −.11, p = .58 and r = −.14, p = .50, respectively) or BMI (r = −.06, p = .77 and r = −.13, p = .53, respectively). Thus, the observed association between [11C]-(+)-PHNO BPND and BMI is not caused by a confounding effect of tracer dose or mass.
The BPND of [11C]-raclopride in the VS was not correlated with BMI (r = −.09, p = .61) in the full sample (n = 35) (Figure 3). There was no correlation in either hemisphere (left: r = −.22, p = .28; right: r = .28, p = .87). Neither age (r = −.23, p = .19) nor sex (r = −.14, p = .44) was related to BPND in the VS. Secondary analyses revealed no correlation with BMI in the caudate (r = −.04, p = .82), putamen (p = −.06, p = .75), or globus pallidus (r = −.06, p = .75). Controlling for age or sex did not significantly change our results with [11C]-raclopride (data not shown).
Figure 3.
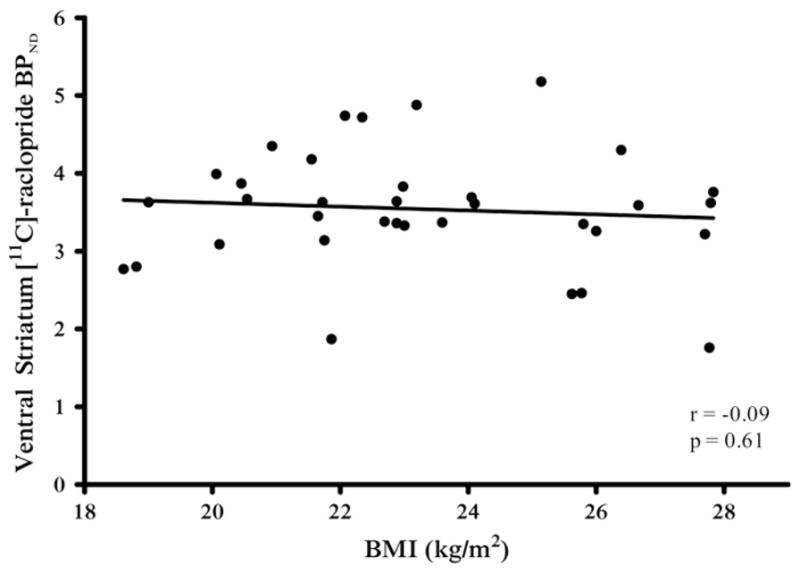
Correlation between body mass index (BMI) and [11C]-raclo-pride binding potential nondisplaceable (BPND) in the ventral striatum in the full sample of subjects (n = 35).
Given the divergent relationship between BMI and BPND in the VS with the two radiotracers, we analyzed a subsample of participants (n = 15) who were scanned with both. This was done to explicitly control for individual differences that may exist between the full samples. Again, we observed a positive correlation between BMI and BPND in the VS with [11C]-(+)-PHNO (r = .55, p = .03) but no correlation with [11C]-raclopride (r = −.16, p = .56). A dependent-correlations t test revealed that the correlation between BMI and [11C]-(+)-PHNO BPND was significantly stronger than the correlation between BMI and [11C]-raclopride BPND (t12 = 2.95, p < .05). This supported our results with the full samples (Figure 4).
Figure 4.
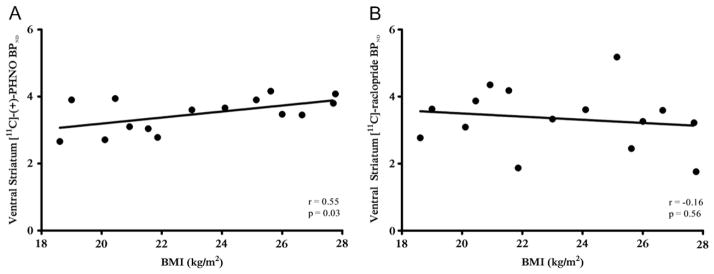
Correlation between body mass index (BMI) and binding potential nondisplaceable (BPND) in the ventral striatum in the subgroup of subjects (n = 15) scanned with both (A) [11C]-(+)-PHNO and (B) [11C]-raclopride.
Discussion
In the present PET study, we investigated how variation in normal BMI relates to D2/3R availability in the VS in humans, with the use of both an agonist and antagonist radiotracer, [11C]-(+)-PHNO and [11C]-raclopride, respectively. Supporting previous findings (5,6), BMI within the normal range was not correlated with [11C]-raclopride binding in the VS. However, normal BMI was positively correlated with [11C]-(+)-PHNO binding in the VS. These differential results were confirmed in a subsample of subjects scanned with both radiotracers, ruling out the influence of participant differences.
Differences in radioligand binding in vivo are usually explained by changes in at least one of three parameters: number of available receptors (Bmax), endogenous dopamine levels (binding competition), or receptor affinity for the ligand (Kd). With the use of the D3R antagonist GSK598809, it has been estimated that ~74% of the [11C]-(+)-PHNO signal in the human VS is attributed to binding at D2R, whereas ~26% is attributed to D3R (29). Similarly, it has been estimated in nonhuman primates that ~19% of the [11C]-raclopride signal in the VS can be occupied by the D3R-preferential antagonist BP897 (30). If our results were caused by changes in D2R expression, it would be unlikely that [11C]-(+)-PHNO would detect the change but [11C]-raclopride would not, especially because [11C]-raclopride labels a greater number of D2Rs in vitro (31). It is also unlikely that our results with [11C]-(+)-PHNO represent altered expression of D3Rs because the contribution of D3Rs to the VS signal for both radiotracers is small, although this possibility cannot be ruled out entirely. Furthermore, we observed no relationship between BMI and BPND in those ROIs in which the majority of the [11C]-(+)-PHNO signal is attributable to D3R binding: the sub-stantia nigra (100%) and globus pallidus (65%) (29). Although D3R function has been suggested to affect susceptibility for obesity in rodents (30), evidence has been mixed (32). In accordance with our findings, recent evidence in overweight and obese individuals suggests that D3Rs do not mediate brain responses to food cues (33).
Another possibility is that our findings with [11C]-(+)-PHNO could be explained by decreases in endogenous dopamine levels with higher BMI. Both [11C]-(+)-PHNO and [11C]-raclopride are sensitive to changes in endogenous dopamine levels (34,35). With the use of an amphetamine challenge in healthy subjects, it has been estimated that [11C]-(+)-PHNO is 1.65 times more sensitive to changes in endogenous dopamine in the VS compared with [11C]-raclopride (36). Taking into account this difference in sensitivity, if our findings with [11C]-(+)-PHNO were driven solely by decreases in endogenous dopamine, we would expect the correlation coefficient between BMI and [11C]-raclopride BPND in the VS to be .30. The observed correlation coefficient was –.089. Furthermore, the percent increase in the average [11C]-(+)-PHNO BPND from the lightest to heaviest persons in our sample (those in the first and fourth quartiles, respectively) was 17.87%. If this change was solely due to endogenous dopamine, we could have expected a 10.83% increase in [11C]-raclopride BPND from the first to the fourth quartile. Instead, we observed a percent change of −9.38%. Thus, we propose that if the relationship between BMI and [11C]-(+)-PHNO BPND was being driven solely by changes in endogenous dopamine, there would have been at least a trend for a positive correlation with [11C]-raclopride. Given that D3Rs have a >20-fold higher affinity for dopamine than D2Rs in vitro (15,16), any reductions in endogenous dopamine levels would affect [11C]-(+)-PHNO BPND at D2Rs before D3Rs (36). Therefore, it is unlikely that the effect observed with [11C]-(+)-PHNO is caused by a difference in its ability to detect changes in endogenous dopamine at D3Rs versus D2Rs compared with [11C]-raclopride.
We posit that our findings are probably explained by changes in D2R affinity for [11C]-(+)-PHNO in the VS. It has been demonstrated in vitro that agonist and antagonist radioligands label different populations of D2Rs. Specifically, D2R agonists but not antagonists are sensitive to changes in the number of active or “high-affinity states” of the receptor (i.e., those coupled to intracellular G-proteins) (14). Although this phenomenon remains to be tested in vivo, the positive association between [11C]-(+)-PHNO binding and BMI within the nonobese range may be due to increased affinity for dopamine at D2Rs in the VS with greater BMI. This increased D2R affinity may be related to increased motivation to consume palatable foods (37,38). This is supported by a recent study in rodents that found that the amount of sucrose intake during the dark phase is positively correlated with D2R sensitivity in the nucleus accumbens, such that the D2R of rodents that consume more sucrose have greater sensitivity to and activation by dopamine (39).
Within the normal range, higher BMI may be driven by an increase in the motivational properties of food. Food cues release dopamine in the VS of rodents (40) and can elicit feeding in satiated rats (41) and humans (42). Furthermore, VS activation in response to food cues is predictive of weight gain in healthy females (10) and is positively correlated with dopamine release during anticipation of rewards (11). Increased D2R affinity in the VS may potentiate the motivating effects of food cues, thereby increasing the number of meals. Conversely, leptin and insulin, hormones that signal energy abundance, reduce dopamine signaling in the nucleus accumbens and suppress feeding (43). Thus, increased D2R affinity may counter-act satiety signaled by reduced dopamine levels, thereby potentiating “not knowing when to stop” eating.
Our findings in conjunction with previous research suggest a dissociable relationship between D2R function and BMI in obesity versus health. Higher weight within the normal range may be driven by increases in D2R affinity (incentive sensitization), whereas higher weight in obesity may be driven by a reduction in D2R expression (reward deficiency). Obesity is related to a reduction in total D2R expression (3,5), mirroring the reduced D2R expression seen in drug addiction (44). This suggests that whereas feeding behavior may exist on a continuum, the state of obesity, much like drug addiction, may be categorically distinct. This is supported by the fact that less [11C]-raclopride binding in the striatum is correlated with greater BMI in obese individuals but not in healthy control subjects (5). Consistently, obese individuals are more likely to carry the Taq1 A1 allele of the D2R gene (45), which is associated with reduced D2R expression and [11C]-raclopride binding (46). This further supports that reduced [11C]-raclopride binding in obesity reflects reduced D2R expression, leading to a “reward deficiency syndrome,” whereby obese individuals overeat to compensate for hypoactivation of reward circuits (5). Future research is needed to examine the role of D2R affinity in obesity.
Because this was a retrospective study, we did not have a direct measure of reward sensitivity in our subjects. However, our interpretation is consistent with recent findings of a nonlinear relationship between sensitivity to reward (SR) and BMI (31), which has been replicated in children (33). These studies demonstrate that within the nonobese BMI range, there is a positive association between self-reported SR and BMI, such that higher BMI is associated with increased SR. Thus, within the normal range, higher BMI may be associated with increased appetitive drive for rewards such as food. We propose that increased D2R affinity may be a contributing neurobiological mechanism. These studies also observe that within the obese range there is a negative relationship between BMI and SR, such that higher BMI was associated with reduced SR. This is consistent with obesity being associated with reward deficiency leading to compensatory overeating, with reduced D2R expression being a contributing neurobiological factor.
Our group, along with others, did not find normal weight to be related to D2R function in the dorsal striatum. Abnormal functioning of the dorsal striatum may relate specifically to obesity and/or food addiction. Reduced D2R expression is seen in the dorsal striatum of obese humans (6) and in animal models of obesity (3). Youth at risk for obesity show greater activation in the right caudate on receipt of palatable food and monetary reward (47). Similarly, obese individuals show increased glucose metabolism and activation in response to food cues in the right caudate during euglycemic hyperinsulinemia (induced satiety) (48). Interestingly, we found that the relationship between normal BMI and [11C]-(+)-PHNO binding was strongest in the right VS. Future research should clarify the role of the dorsal and ventral striatum and each hemisphere in BMI.
There are a number of limitations to the current study. First, this study was retrospective. Second, we did not directly measure eating behaviors or adiposity in participants. Third, although the majority of the [11C]-(+)-PHNO signal in the VS is caused by D2R binding, we could not parse out the contribution of D3Rs; thus, changes in D3R expression cannot be fully ruled out. Finally, we did not examine endogenous dopamine levels; thus, its contribution cannot be fully ruled out. This study sets the groundwork for exploring the role of D2R agonist binding sites in the etiology, treatment, and prevention of obesity.
Acknowledgments
The authors thank the PET Centre staff at the Centre for Addiction and Mental Health, including Alvina Ng and Laura Nguyen, for technical assistance in data collection. They also thank Wanna Mar, Carol Borlido, and Kathryn Kalahani-Bargis for assistance in participant recruitment.
This study was partially funded by Canadian Institutes of Health Research (MOP-114989) and U.S. National Institute of Health (RO1MH084886-01A2).
Footnotes
Dr. Nakajima reports having received grants from Japan Society for the Promotion of Science and Inokashira Hospital Research Fund and speaker’s honoraria from GlaxoSmith Kline, Janssen Pharmaceutical, Pfizer, and Yoshitomiyakuhin within the past 3 years. Dr. Graff-Guerrerro currently receives research support from the following external funding agencies: Canadian Institutes of Health Research, the U.S. National Institute of Health, and the Mexico Instituto de Ciencia y Tecnología para la Capital del Conocimiento en el Distrito Federal (ICyTDF). He has also received professional services compensation from Abbott Laboratories, Gedeon-Richter Plc, and Lundbeck; grant support from Janssen; and speaker compensation from Eli Lilly. Mr. Caravaggio, Ms. Raitsin, Dr. Gerretsen, and Dr. Wilson reported no biomedical financial interests or potential conflicts of interest.
References
- 1.Ogden CLCM, Kit BK, Flegal KM. Prevalence of Obesity in the United States, 2009–2010. NCHS Data Brief, No 82. Hyattsville, MD: National Center for Health Statistics; 2012. [PubMed] [Google Scholar]
- 2.Volkow ND, Wang G-J, Baler RD. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn Sciences. 2011;15:37–46. doi: 10.1016/j.tics.2010.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Johnson PM, Kenny PJ. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci. 2010;13:635–641. doi: 10.1038/nn.2519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Huang XF, Zavitsanou K, Huang X, Yu Y, Wang H, Chen F, et al. Dopamine transporter and D2 receptor binding densities in mice prone or resistant to chronic high fat diet-induced obesity. Behav Brain Res. 2006;175:415–419. doi: 10.1016/j.bbr.2006.08.034. [DOI] [PubMed] [Google Scholar]
- 5.Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, et al. Brain dopamine and obesity. Lancet. 2001;357:354–357. doi: 10.1016/s0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- 6.Volkow ND, Wang GJ, Telang F, Fowler JS, Thanos PK, Logan J, et al. Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: possible contributing factors. Neuroimage. 2008;42:1537–1543. doi: 10.1016/j.neuroimage.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Haltia LT, Rinne JO, Merisaari H, Maguire RP, Savontaus E, Helin S, et al. Effects of intravenous glucose on dopaminergic function in the human brain in vivo. Synapse. 2007;61:748–756. doi: 10.1002/syn.20418. [DOI] [PubMed] [Google Scholar]
- 8.de Weijer B, van de Giessen E, van Amelsvoort T, Boot E, Braak B, Janssen I, et al. Lower striatal dopamine D2/3 receptor availability in obese compared with non-obese subjects. Eur J Nucl Med Mol Imaging. 2011;1:37. doi: 10.1186/2191-219X-1-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Michaelides M, Thanos PK, Kim R, Cho J, Ananth M, Wang G-J, et al. PET imaging predicts future body weight and cocaine preference. Neuroimage. 2012;59:1508–1513. doi: 10.1016/j.neuroimage.2011.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Demos KE, Heatherton TF, Kelley WM. Individual differences in nucleus accumbens activity to food and sexual images predict weight gain and sexual behavior. J Neurosci. 2012;32:5549–5552. doi: 10.1523/JNEUROSCI.5958-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schott BH, Minuzzi L, Krebs RM, Elmenhorst D, Lang M, Winz OH, et al. Mesolimbic functional magnetic resonance imaging activations during reward anticipation correlate with reward-related ventral striatal dopamine release. J Neurosci. 2008;28:14311–14319. doi: 10.1523/JNEUROSCI.2058-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Baladi MG, Daws LC, France CP. You are what you eat: Influence of type and amount of food consumed on central dopamine systems and the behavioral effects of direct- and indirect-acting dopamine receptor agonists. Neuropharmacology. 2012;63:76–86. doi: 10.1016/j.neuropharm.2012.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Robinson TE, Berridge KC. Review: The incentive sensitization theory of addiction: Some current issues. Philos Trans R Soc Lond B Biol Sci. 2008;363:3137–3146. doi: 10.1098/rstb.2008.0093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Seeman P, McCormick PN, Kapur S. Increased dopamine D2(High) receptors in amphetamine-sensitized rats, measured by the agonist [(3)H](+)PHNO. Synapse. 2007;61:263–267. doi: 10.1002/syn.20367. [DOI] [PubMed] [Google Scholar]
- 15.Bailey A, Metaxas A, Yoo JH, McGee T, Kitchen I. Decrease of D2 receptor binding but increase in D2-stimulated G-protein activation, dopamine transporter binding and behavioural sensitization in brains of mice treated with a chronic escalating dose ’binge’ cocaine administration paradigm. Eur J Neurosci. 2008;28:759–770. doi: 10.1111/j.1460-9568.2008.06369.x. [DOI] [PubMed] [Google Scholar]
- 16.Lee JM, DeLeon-Jones F, Fields JZ, Ritzmann RF. Cyclo (Leu-Gly) attenuates the striatal dopaminergic supersensitivity induced by chronic morphine. Alcohol Drug Res. 1987;7:1–10. [PubMed] [Google Scholar]
- 17.Wilson AA, Garcia A, Jin L, Houle S. Radiotracer synthesis from [(11)C]-iodomethane: a remarkably simple captive solvent method. Nucl Med Biol. 2000;27:529–532. doi: 10.1016/s0969-8051(00)00132-3. [DOI] [PubMed] [Google Scholar]
- 18.Wilson AA, McCormick P, Kapur S, Willeit M, Garcia A, Hussey D, et al. Radiosynthesis and evaluation of [11C]-(+)-4-propyl-3,4,4a,5,6,10b-hexahydro-2H-naphtho[1,2-b][1,4]oxazin-9-ol as a potential radiotracer for in vivo imaging of the dopamine D2 high-affinity state with positron emission tomography. J Med Chem. 2005;48:4153–4160. doi: 10.1021/jm050155n. [DOI] [PubMed] [Google Scholar]
- 19.Graff-Guerrero A, Redden L, Abi-Saab W, Katz DA, Houle S, Barsoum P, et al. Blockade of [11C](+)-PHNO binding in human subjects by the dopamine D3 receptor antagonist ABT-925. Int J Neuropsychopharmacol. 2010;13:273–287. doi: 10.1017/S1461145709990642. [DOI] [PubMed] [Google Scholar]
- 20.Graff-Guerrero A, Willeit M, Ginovart N, Mamo D, Mizrahi R, Rusjan P, et al. Brain region binding of the D2/3 agonist [11C]-(+)-PHNO and the D2/3 antagonist [11C]raclopride in healthy humans. Hum Brain Mapp. 2008;29:400–410. doi: 10.1002/hbm.20392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Studholme C, Hill DL, Hawkes DJ. Automated three-dimensional registration of magnetic resonance and positron emission tomography brain images by multiresolution optimization of voxel similarity measures. Med Phys. 1997;24:25–35. doi: 10.1118/1.598130. [DOI] [PubMed] [Google Scholar]
- 22.Lammertsma AA, Hume SP. Simplified reference tissue model for PET receptor studies. Neuroimage. 1996;4:153–158. doi: 10.1006/nimg.1996.0066. [DOI] [PubMed] [Google Scholar]
- 23.Gunn RN, Lammertsma AA, Hume SP, Cunningham VJ. Parametric imaging of ligand-receptor binding in PET using a simplified reference region model. Neuroimage. 1997;6:279–287. doi: 10.1006/nimg.1997.0303. [DOI] [PubMed] [Google Scholar]
- 24.Mawlawi O, Martinez D, Slifstein M, Broft A, Chatterjee R, Hwang DR, et al. Imaging human mesolimbic dopamine transmission with positron emission tomography, I: Accuracy and precision of D(2) receptor parameter measurements in ventral striatum. J Cereb Blood Flow Metab. 2001;21:1034–1057. doi: 10.1097/00004647-200109000-00002. [DOI] [PubMed] [Google Scholar]
- 25.Mamo D, Graff A, Mizrahi R, Shammi CM, Romeyer F, Kapur S. Differential effects of aripiprazole on D(2), 5-HT(2), and 5-HT(1A) receptor occupancy in patients with schizophrenia: A triple tracer PET study. Am J Psychiatry. 2007;164:1411–1417. doi: 10.1176/appi.ajp.2007.06091479. [DOI] [PubMed] [Google Scholar]
- 26.Graff-Guerrero A, Mizrahi R, Agid O, Marcon H, Barsoum P, Rusjan P, et al. The dopamine D2 receptors in high-affinity state and D3 receptors in schizophrenia: A clinical [11C]-(+)-PHNO PET study. Neuropsychopharmacology. 2009;34:1078–1086. doi: 10.1038/npp.2008.199. [DOI] [PubMed] [Google Scholar]
- 27.Cohen J. A power primer. Psychol Bull. 1992;112:155–159. doi: 10.1037//0033-2909.112.1.155. [DOI] [PubMed] [Google Scholar]
- 28.Rabiner EA, Laruelle M. Imaging the D3 receptor in humans in vivo using [11C](+)-PHNO positron emission tomography (PET) Int J Neuropsychopharmacol. 2010;13:289–290. doi: 10.1017/S1461145710000088. [DOI] [PubMed] [Google Scholar]
- 29.Tziortzi AC, Searle GE, Tzimopoulou S, Salinas C, Beaver JD, Jenkinson M, et al. Imaging dopamine receptors in humans with [11C]-(+)-PHNO: dissection of D3 signal and anatomy. Neuroimage. 2011;54:264–277. doi: 10.1016/j.neuroimage.2010.06.044. [DOI] [PubMed] [Google Scholar]
- 30.Davis C, Fox J. Sensitivity to reward and body mass index (BMI): Evidence for a non-linear relationship. Appetite. 2008;50:43–49. doi: 10.1016/j.appet.2007.05.007. [DOI] [PubMed] [Google Scholar]
- 31.Kiss B, Horti F, Bobok A. In vitro and in vivo comparison of [(3)H](+)-PHNO and [(3)H]raclopride binding to rat striatum and lobes 9 and 10 of the cerebellum: A method to distinguish dopamine D(3) from D(2) receptor sites. Synapse. 2011;65:467–478. doi: 10.1002/syn.20867. [DOI] [PubMed] [Google Scholar]
- 32.Verbeken S, Braet C, Lammertyn J, Goossens L, Moens E. How is reward sensitivity related to bodyweight in children? Appetite. 2012;58:478–483. doi: 10.1016/j.appet.2011.11.018. [DOI] [PubMed] [Google Scholar]
- 33.Dodds CM, O’Neill B, Beaver J, Makwana A, Bani M, Merlo-Pich E, et al. Effect of the dopamine D3 receptor antagonist GSK598809 on brain responses to rewarding food images in overweight and obese binge eaters. Appetite. 2012;59:27–33. doi: 10.1016/j.appet.2012.03.007. [DOI] [PubMed] [Google Scholar]
- 34.Shotbolt P, Tziortzi AC, Searle GE, Colasanti A, van der Aart J, Abanades S, et al. Within-subject comparison of [(11)C]-(+)-PHNO and [(11)C]raclopride sensitivity to acute amphetamine challenge in healthy humans. J Cereb Blood Flow Metab. 2012;32:127–136. doi: 10.1038/jcbfm.2011.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Willeit M, Ginovart N, Graff A, Rusjan P, Vitcu I, Houle S, et al. First human evidence of d-amphetamine induced displacement of a D2/3 agonist radioligand: A [11C]-(+)-PHNO positron emission tomography study. Neuropsychopharmacology. 2008;33:279–289. doi: 10.1038/sj.npp.1301400. [DOI] [PubMed] [Google Scholar]
- 36.Caravaggio F, Mamo D, Menon M, Borlido C, Gerretsen P, Wilson A, et al. Imaging D3 receptor occupancy by endogenous dopamine in humans: A [11C]-(+)-PHNO PET study. Poster presented at: Annual Meeting of Society for Neuroscience; October 12–17; New Orleans, Louisiana. 2012. [Google Scholar]
- 37.Egecioglu E, Skibicka KP, Hansson C, Alvarez-Crespo M, Friberg PA, Jerlhag E, et al. Hedonic and incentive signals for body weight control. Rev Endocr Metab Disord. 2011;12:141–151. doi: 10.1007/s11154-011-9166-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Berridge KC. Liking’ and ’wanting’ food rewards: Brain substrates and roles in eating disorders. Physiol Behav. 2009;97:537–550. doi: 10.1016/j.physbeh.2009.02.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tonissaar M, Herm L, Rinken A, Harro J. Individual differences in sucrose intake and preference in the rat: circadian variation and association with dopamine D2 receptor function in striatum and nucleus accumbens. Neurosci Lett. 2006;403:119–124. doi: 10.1016/j.neulet.2006.04.023. [DOI] [PubMed] [Google Scholar]
- 40.Phillips AG, Vacca G, Ahn S. A top-down perspective on dopamine, motivation and memory. Pharmacol Biochem Behav. 2008;90:236–249. doi: 10.1016/j.pbb.2007.10.014. [DOI] [PubMed] [Google Scholar]
- 41.Weingarten HP. Conditioned cues elicit feeding in sated rats: A role for learning in meal initiation. Science. 1983;220:431–433. doi: 10.1126/science.6836286. [DOI] [PubMed] [Google Scholar]
- 42.Cornell CE, Rodin J, Weingarten H. Stimulus-induced eating when satiated. Physiol Behav. 1989;45:695–704. doi: 10.1016/0031-9384(89)90281-3. [DOI] [PubMed] [Google Scholar]
- 43.Palmiter RD. Is dopamine a physiologically relevant mediator of feeding behavior? Trends Neurosci. 2007;30:375–381. doi: 10.1016/j.tins.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 44.Martinez D, Greene K, Broft A, Kumar D, Liu F, Narendran R, et al. Lower level of endogenous dopamine in patients with cocaine dependence: Findings from PET imaging of D(2)/D(3) receptors following acute dopamine depletion. Am J Psychiatry. 2009;166:1170–1177. doi: 10.1176/appi.ajp.2009.08121801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Chen AL, Blum K, Chen TJ, Giordano J, Downs BW, Han D, et al. Correlation of the Taq1 dopamine D2 receptor gene and percent body fat in obese and screened control subjects: a preliminary report. Food Funct. 2012;3:40–48. doi: 10.1039/c1fo10089k. [DOI] [PubMed] [Google Scholar]
- 46.Comings DE, Blum K. Reward deficiency syndrome: Genetic aspects of behavioral disorders. Prog Brain Res. 2000;126:325–341. doi: 10.1016/S0079-6123(00)26022-6. [DOI] [PubMed] [Google Scholar]
- 47.Stice E, Yokum S, Burger KS, Epstein LH, Small DM. Youth at risk for obesity show greater activation of striatal and somatosensory regions to food. J Neurosci. 2011;31:4360–4366. doi: 10.1523/JNEUROSCI.6604-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nummenmaa L, Hirvonen J, Hannukainen JC, Immonen H, Lindroos MM, Salminen P, et al. Dorsal striatum and its limbic connectivity mediate abnormal anticipatory reward processing in obesity. PLoS One. 2012;7:3. doi: 10.1371/journal.pone.0031089. [DOI] [PMC free article] [PubMed] [Google Scholar]