

Abstract
The severe degeneration of the germinal epithelium and subsequent male sterility observed in mice null for the retinoic acid receptor α (RARα) gene suggested its critical role in spermatogenesis, although the etiology and progression of these abnormalities remain to be determined. Previous studies have revealed that elongated spermatids in RARα−/− testes were improperly aligned at the tubular lumen and did not undergo spermiation at stage VIII*. We now report a distinctive failure of step 8–9 spermatids to orient properly with regard to the basal aspect of Sertoli cells, resulting in stage VIII*–IX* tubules with randomly oriented spermatids. By in situ terminal deoxynucleo-tidyltransferase-mediated deoxy-UTP nick end labeling (TUNEL), we noted that elongating spermatids frequently underwent apoptosis. Immunohistochemical analysis revealed that while activated caspase-3, the primary effector caspase in the apoptotic cell death machinery, was detected in the nuclei of primary spermatocytes in the first wave of spermatogenesis and occasionally in spermatogonia of both normal and mutant testes, it was not involved in the death of elongating spermatids in RARα−/− testes. Thus, sterility in RARα−/− males was associated with specific defects in spermiogenesis, which may correlate with a failure in both spermatid release and spermatid orientation to the basal aspect of Sertoli cells at stage VIII* in young adult RARα−/− testis. Further, the resulting apoptosis in elongating spermatids appears to involve pathways other than that mediated by activated caspase-3.
Keywords: apoptosis, spermiogenesis, ectoplasmic specializations, acrosome, RARα, caspase-3, retinoid signaling
Introduction
It has been known for decades that dietary retinol (vitamin A) is required by the testis for maintenance of spermatogenesis and by the epididymis for the maturation of spermatozoa reviewed in (Eskild and Hansson, 1994; Packer and Wolgemuth, 1999; Chung and Wolgemuth, 2004). The requirement of vitamin A and its derivatives in the testis has been assessed by examining the effects on spermatogenesis in vitamin A-deficient (VAD) animals, particularly in the rat. The mechanism by which vitamin A affects the development of germ cells remains unknown, while key players in the regulation of spermatogenesis by retinoids have begun to be elucidated through genetic studies on the retinoid receptors (Lufkin et al., 1993; Kastner et al., 1996). There are two known families of retinoid receptors: the retinoic acid receptors (RARs), which recognize either all-trans or 9-cis RA, and the retinoid X receptors (RXRs), which recognize exclusively 9-cis RA (Kastner et al., 1995; Mangelsdorf and Evans, 1995; Mangelsdorf et al., 1995). Each family consists of three types of receptors, α, β and γ, each encoded by distinct genes.
A critical role for RARα signaling in spermatogenesis was evidenced by the severe degeneration of the seminiferous epithelium and sterility observed in RARα-deficient mice. The expression patterns of RARα suggest possible roles in processes as diverse as Sertoli cell functioning, meiotic prophase, and spermiogenesis (Kim and Griswold, 1990; Eskild et al., 1991; van Pelt et al., 1992; Kim and Wang, 1993; Lufkin et al., 1993; Akmal et al., 1997). The testicular abnormalities observed in the adult RARα knockout mice were apparently similar to that seen in mice maintained on a VAD diet (Lufkin et al., 1993). The testes displayed patchy lesions of the seminiferous tubules, with some tubules that were markedly atrophic and/or mostly devoid of spermatogenic cells while some adjacent tubules were relatively normal (Lufkin et al., 1993). However, neither the mechanisms responsible for the degeneration nor the progression of appearance of the testicular abnormalities in the absence of RARα were noted.
In these earlier studies, most of the RARα−/− mice in fact exhibited perinatal lethality, and so spermatoenesis could be evaluated in only a few animals (Lufkin et al., 1993). However, significant improvement in the viability of these animals can be achieved by rearing them in a pathogen-free environment (Chung et al., 2004), rendering possible histological and molecular analysis of the etiology of the testicular degeneration. In particular, we were interested in whether the cells in the degenerating tubules were dying by apoptotic cell death pathways. If so, we were also interested in identifying the types of cells undergoing apoptosis and determining whether this is a primary response to the absence of RARα-mediated signaling by examining testes during the first wave of spermatogenesis. Further, we and others had previously observed defects in spermiation and a failure of elongated spermatids to align properly in stage VIII tubules (Chung et al., 2004). We were thus interested to explore the fate of such improperly aligned spermatids and the cellular basis for this property.
Materials and methods
Source of animals and tissues
RARα−/− mice were obtained from Dr. Pierre Chambon (Lufkin et al., 1993) and were maintained on a mixed 129/C57BL/6 background. Testes of RARα−/− and RARα+/+ mice at 2, 3, 4, 6, 7, 8 and 9 weeks of age were dissected from anesthetized animals that had been perfused through the heart with phosphate-buffered saline (PBS) and then with 4% paraformaldehyde in PBS. Tissues were fixed overnight at 4°C in 4% paraformaldehyde in PBS. Perfused, fixed tissues were embedded in paraffin, sectioned at 5 μm, and mounted on Superfrost slides (Fisher Scientific, Pittsburgh, PA).
TUNEL staining of apoptotic cells
Histological sections were deparaffinized in histoclear, hydrated through a graded alcohol series, and washed with H2O. In situ labeling of apoptotic cells was performed on tissue sections from both RARα−/− and RARα+/+ testes using the in situ cell death detection kit, horseradish peroxidase (POD) (Roche, Indianapolis, IN), according to the manufacturer's instructions. In brief, sections were treated with 20 μg/ml proteinase K in 10 mM Tris/HCl (pH 8.0) and endogenous peroxidase was blocked in 0.3% H2O2 for 30 min. Single- and double-strand breaks were 3′ labeled with fluorescein-labeled nucleotides, using terminal deoxynucleotidyl transferase. Incorporated fluorescein was detected by anti-fluorescein antibody Fab fragments from sheep, conjugated with POD. Bound antibodies were covered with 0.4 μg/μl 3,3′ diaminobenzidine tetrahydrochloride (DAB, Sigma, St. Louis, MO) in 0.1 M Tris (pH 7.2). Only clearly stained cells were considered to be positive and only round-shaped tubules were evaluated. At least 100 tubular cross-sections of each testis from three different animals were counted per developmental time point (Beumer et al., 1997; Rodriguez et al., 1997). The results of individual counts were compared to counts within the same age group as well as between age groups. Significant differences were assessed by statistical analysis using Student's t-test and ANOVA with Scheffe's Post-hoc tests using the StatView Statistical Analysis Software Package (Version 5.0) from SAS Institute Inc. (Cary, NC).
Immunohistochemistry
Immunohistochemical analyses were performed using a Vectastain ABC kit (Vector Laboratories, Burlingame, CA) as previously described (Liu et al., 1998). Briefly, histological sections were deparaffinized in histoclear, hydrated through an alcohol series, and washed with H2O. Antigen unmasking was performed by boiling slides in a microwave for 10 min in 0.01 M citrate buffer, pH 6 (Shi et al., 1991). Following treatment with 0.3% H2O2 in water for 10 min, sections were washed with 1 × PBS. Slides were blocked for 1 hr at room temperature with 2.5% rabbit serum in PBS. The primary antibody incubation was carried out overnight at 4°C in a humidified chamber. Polyclonal anti-activated caspase-3 (Cell Signaling Technology, Inc., Beverly, MA; catalog no. 9661; this antibody recognizes the p19 and p17 cleavage products of caspase-3 but not full-length caspase-3) was diluted 1:100 in 1 × PBS. For controls, the slides were incubated with corresponding IgG instead of primary antibody. After incubation, slides were washed for 30min in PBS and processed using the Vectastain ABC kit following the manufacturer's instructions. DAB-stained slides were counterstained with hematoxylin according to standard procedures. They were viewed on a Nikon photomicroscope under bright-field optics. Photomicrographs were taken using Fujicolor 400 film. For staging of the mouse seminiferous epithelium according to Oakberg (1956) and Russell et al. (1990), the testis sections were stained for the periodic acid Schiff (PAS) reaction before hematoxylin counterstaining. For comparison of the relative levels of expression between samples, great care was taken to minimize the variation in fixation and thickness of sections, etc. In addition, tissues from different experimental groups (e.g., mutant versus normal, different ages, etc.) were sectioned onto the same slides to enable relative quantitative comparisons with greater confidence (Mutter et al., 1988).
Assessment of spermatid orientation within tubules
Histological sections were stained for the PAS reaction and stage VIII*–IX* tubules were analyzed. Only those tubules cut in cross-section, as judged by their approximation to a circle and by regularity of cell layers, were used. In normal testes, the orientation of the acrosomes in step 8–9 spermatids was exclusively toward the basal aspect of the Sertoli cell. If the shape of the seminiferous tubules is estimated as a concentric circle, the acrosome of each step 8–9 spermatid is pointed outwards and oriented perpendicular to an individual line drawn through the center point of the circle. In our model, we counted the number of spermatids that corresponded to this normal presentation relative to the number that deviated from this orientation (acrosomes that are not perpendicular to lines drawn through the center).
Results
Failure in both spermatid release and spermatid orientation to the basal aspect of Sertoli cells in young adult RARα −/− testis
It had been reported previously that RARα −/− testes at 4–5 months showed severe degeneration of the germinal epithelium, although no information on the progression of the testicular abnormalities was provided (Lufkin et al., 1993). Our systematic investigation of the progression of the first wave of spermatogenic differentiation revealed that there was a temporary developmental arrest in step 8–9 spermatids in the first wave of spermatogenesis (Chung et al., 2004). Further, we have previously reported a failure of elongated spermatids to align properly in stage VIII tubules (Chung et al., 2004). Normally, step 16 spermatids align at the surface of the tubular lumen prior to spermiation (arrowhead indicating step 16 spermatids in Fig. 1A). Then, in stage IX from control testes, all step 16 spermatids are released and three layers of germ cells including leptotene spermatocytes, late pachytene spermatocytes and step 9 elongating spermatids are observed (Fig. 1B). It is known that step 8–9 spermatids undergo a characteristic morphological transformation during spermiogenesis. Also in stage VIII tubules, the nuclei of the step 8 spermatids move toward their inner cellular membranes. The acrosomal region of the nucleus abuts the spermatid plasma membrane and rotates within the cell to face the basal aspect of the Sertoli cell, with the spermatid tails projecting into the tubule lumen (Oakberg, 1956; Ross, 1976; Russell, 1977b). The orientation of spermatids can be visualized by PAS staining of the acrosomes and determining their position relative to the basal compartment (Fig. 1A). This feature actually differentiates stage VIII from stage VII (Russell et al., 1990). The characteristic orientation of the acrosomes of the very early elongating step 9 spermatids in normal testes was exclusively toward the basal aspect of the Sertoli cell (arrows in Fig. 1B).
Fig. 1.
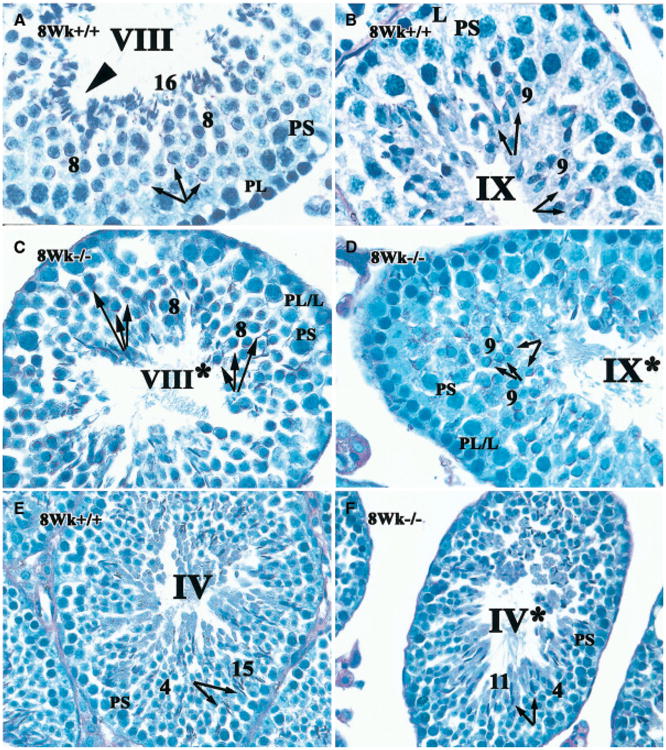
Failure of spermatid release and aberrant orientation of spermatids of RARα−/− testis at stage VIII within the seminiferous epithelium. The acrosome of spermatids in histological sections of testes from RARα+/+(A, B, E) and RARα−/− males (C, D, F) was detected with PAS staining (magenta color). For RARα+/+ testis, early stage VIII (A), stage IX (B), stage IV (E) are shown next to corresponding stages in RARα−/− testis (stage VIII*, C; stage IX*, D; stage IV*, F, respectively). Though the asynchronous cell associations within the seminiferous epithelium of the mutant testes make “staging” difficult, an attempt was made to stage the tubules using the acrosomal system (Russell et al., 1990). PS, pachytene spermatocytes; PL/L, preleptotene or leptotene spermatocytes; PL, preleptotene spermatocytes; L, leptotene spermatocytes; arabic numerals, the step of elongated spermatid. Roman numerals indicate the stage of the seminiferous tubule. A–D, × 100; E, F, × 60. Arrows in A–D indicate the orientation of elongated speramtids while the arrows in E and F show the entrenchment of spermatids (or lack thereof) within a recess of Sertoli cells. Arrowhead in A indicates step 16 spermatids.
In contrast, although nuclei in step 8 and 9 spermatids in RARα−/− testis moved toward their plasma membranes, these spermatids failed to become orientated toward the basal aspect of Sertoli cells (Figs. 1C, 1D, arrows). Rather, they appeared to be randomly oriented with regard to the basal lamina. It should be noted that it is difficult to “stage” the seminiferous tubules in RARα−/− testes, because of the asynchronous cell association within the seminiferous epithelium (Chung et al., 2004). Nonetheless, we attempted to characterize the developmental progression of the cells within the tubules using the acrosomal system and referring to the putative stage with an asterisk. We found that approximately 77% of step 8–9 spermatids were randomly oriented in stage VIII*–IX* tubules in RARα−/− testes, as compared to ∼ 4% of the controls. In addition, four (rather than three) layers of germ cells were observed in stage IX*, where late spermatids were retained in the epithelia (Fig. 1D). As noted previously (Chung et al., 2004), at stage VIII* of RARα−/− testes, no alignment of mature spermatids at the surface of tubular lumen was found (Fig. 1C). Further, while entrenchment of spermatids within more basal aspects of Sertoli cells was observed in RARα+/+ testis at stage IV (arrows in Fig. 1E), it was absent in corresponding RARα−/− stage IV* (arrows in Fig. 1F).
Apoptosis in elongated spermatids occurred in RARα −/− testes
To determine whether the cells in the degenerating testicular tubules and the abnormally oriented elongating spermatids, in particular, in RARα−/− testes, were undergoing apoptosis, Terminal deoxynucleotidyltransf-erase-mediated deoxy-UTP nick end labeling (TUNEL) staining was used in combination with morphological analysis. In normal adult testis, the most frequent TUNEL-positive cells were spermatogonia and early and meiotically dividing spermatocytes (Henriksen et al., 1995; Rodriguez et al., 1997; Sjoblom et al., 1998; Yan et al., 2000). We examined young adult (8–9 weeks of age) control and RARα−/− testes; some tubules with TUNEL-positive germ cells in RARα−/− testes looked very similar to those seen in RARα+/+ testes (Figs. 2B and 2C versus Fig. 2A, respectively). However, the presence of TUNEL-positive spermatids at the periphery of some of the tubules was noted in RARα−/− testes (Figs. 2B, 2D, 2E) but never in RARα+/+ testes (Fig. 2A). These TUNEL-positive spermatids often lay close to the basal lamina. These peripherally located spermatids were at steps 10–11 of spermiogenesis as determined by PAS staining (data not shown) and examining the tubules at high magnification (arrows in insert in Fig. 2D). Tubules exhibiting both TUNEL-positive elongated spermatids as well as occasional TUNEL-positive spermatogonia and spermatocytes were consistently found in the young adult RARα−/− testes (Fig. 2E).
Fig. 2.

Comparison of histology and TUNEL-labeling in young adult testes of RARα+/+ and RARα−/− testis. Histological sections of 8- and 9-week-old (young adult) testes from RARα+/+ (A) and RARα−/− mice (B–E), with corresponding epididymis from RARα+/+ (H) and RARα−/− (I) males, are shown, along with histological sections of 6-week-old testis from RARα−/− mice (F, G). Staining with hematoxylin revealed the appearance of TUNEL-positive spermatids at the basal lamina only in the young adult RARα−/− testis (B, D, E) and not in the RARα+/+ testis (A). Apoptotic elongated spermatids were detected when they were deeply embedded in the seminiferous epithelium of 6-week-old RARα−/− testis (G, inset with high magnification). Panels A and B, × 20; C–E, × 60; H and I, × 60; F and G, × 40. Small panel in D shows magnification at × 100. Small panel in F–G, × 60. Arrowheads in F–G point to the TUNEL-positive spermatogenic cells. Arabic numerals in panel D indicate the step of elongated spermatid.
To further examine the consequences of defective spermiogenesis and spermiation, sections of the corresponding epididymis of young adult RARα+/+ and RARα−/− mice were examined (Fig. 2H and 2I, respectively). As expected, spermatozoa in the epididymis of RARα−/− mice were strikingly diminished in number and most of those that were observed appeared defective (Fig. 2I) when compared to normal control (Fig. 2H).
Germ cell apoptosis during testicular development in RARα+/+ and RARα−/− testes
We then wished to explore whether the apoptotic phenomena observed were a primary effect of the lack of RARα, as opposed to the result of an overall degeneration of the tubules and, hence, an inability to support various stages of spermatogenesis. We therefore used the TUNEL technique to examine histological sections of testes during the first wave of spermatogenesis in both RARα+/+ and RARα−/− mice. The total number of TUNEL-positive germ cells (all TUNEL-positive germ cells except elongated spermatids) per 100 seminiferous tubules in both normal and mutant testes at 2, 3, 4, 5, 6, 7, 8 and 9 weeks of age was quantified. The number of TUNEL-positive germ cells peaked at around 3 weeks in normal mouse testes (Fig. 3A), in agreement with observations of others (Rodriguez et al., 1997; Yan et al., 2000). The highest number of TUNEL-positive germ cells (not including elongated spermatids) was also seen in 3-week-old RARα−/− testes; however, there was a second peak at around 6 weeks, which then dropped at 7, 8 and 9 weeks (Fig. 3A).
Fig. 3.

Germ cell apoptosis during testicular development in RARα+/+ and RARα−/− testes. (A) A bar graph indicating the number of TUNEL-positive germ cells at various intervals after birth in RARα+/+ and RARα−/− mice (100 tubules scored per testis; three mice for each time point). The total number of apoptotic germ cells (excluding elongated spermatids) per 100 seminiferous tubules was counted in testicular sections of 2–9-week-old RARα+/+ and RARα−/− males. The bars represent the mean ± SD of three mice for each time point. (*) indicates significant differences within the same age group as well as between age groups by ANOVA, p<0.05. (**) indicates significant differences within the same age group as well as between age group by ANOVA, p<0.001. (***) indicates significant differences within the same age group as well as between age groups by ANOVA, p< 0.0001. (B) Typical round-shaped tubules with apoptotic heads of elongated spermatids at the periphery of the tubules are designated as T+. × 20. (C) Bar graph indicating the percentage of tubules with TUNEL-positive elongated spermatids at various intervals after birth in RARα−/− mice (100 tubules scored per testis: three mice for each time point). Appearance of TUNEL-positive elongated spermatids was not observed in the tubules until they were 6 weeks of age. These unique TUNEL-positive elongated spermatids were never seen in RARα+/+ testis at various intervals after birth (data not shown).
We next determined the number of tubules with TUNEL-positive heads of elongated spermatids, where they populated the periphery of the tubule (Fig. 3B, designated as T+), per 100 seminiferous tubules of developmental ages noted above in mutant mice (Fig. 3C). As mentioned above, TUNEL-positive elongated spermatids were never observed in tubules from normal testes (Fig. 2A). The presence of the TUNEL-positive spermatids was typically not observed in the tubules until they were 6 weeks of age (Figs. 2G, 3C) and reached a maximum at 7 weeks (Fig. 3C). However, the exact timing of the appearance of apoptotic elongated spermatids varied among animals. For example, while detailed examination of 6-week-old RARα−/− testes revealed that apoptotic elongated spermatids were detected by TUNEL staining when they were deeply embedded in the seminiferous epithelium (Fig. 2G), occasionally, no apoptotic elongated spermatids were found in 6-week-old RARα−/− mice (Fig. 2F). By 8 and 9 weeks of age, the frequency of TUNEL-positive spermatids decreased (Fig. 3C).
Activated caspase-3 is involved in the death of spermatogonia and spermatocytes but not elongated spermatids in RARα−/− testes
There is increasing evidence that various animal cells use similar death programs, which depend on one or more members of a family of aspartic-acid-directed cysteine proteases called caspases (cysteinyl aspartate-specific proteinases) to effect the cell death (Alnemri et al., 1996; Nicholson and Thornberry, 1997; Nicholson and Thornberry, 2003). Caspase-3 is the most important effector caspase. Its activation hallmarks the point of no return in programmed cell death signaling (Earnshaw et al., 1999; Said et al., 2004). However, it has been suggested that the death of spermatozoa, which are cells with a transcriptionally inactive nucleus, shows a caspase-independent death program (Weil et al., 1998). To explore whether caspases are involved in the death of elongated spermatids in RARα−/− testes, we followed the post-natal developmental expression of activated caspase-3 in both RARα+/+ and RARα−/− testes.
In 8-day-old testes, where differentiating type A1–4, intermediate, and type B spermatogonia predominate, only a few activated caspase-3-labeled cells were found in either mutant or control testes (data not shown). We were able to detect activated caspase-3 predominantly in early primary spermatocytes in the first wave of spermatogenesis in testes from 2- and 3-week-old mice from both RARα+/+ and RARα−/− strains (Figs. 4B, 4D, 4F; Fig. 5). Activated caspase-3 was occasionally detected in spermatogonia, but not in Sertoli cells or Leydig cells in these testes (Figs. 4B, 4D, 4F). In some tubules, strong expression of activated caspase-3 was found in primary spermatocytes (arrow in Fig. 4D). This expression correlated with the presence of TUNEL-labeled cells in the same area in the next serial section (arrow in Fig. 4C), suggesting it could be the site of cells collectively undergoing apoptosis (arrowhead in Fig. 4C). In contrast, although TUNEL-labeled elongated spermatids at the periphery of the seminiferous tubules were clearly detected in the histological sections of adult RARα−/− testes, no expression of activated caspase-3 was found in the corresponding cell types in RARα−/− testes of the next serial section (Figs. 4G versus 4H, respectively). This indicated that apoptosis observed in elongated spermatids may be effected via a caspase-3-independent pathway.
Fig. 4.

Immunohistochemical localization of caspase-3 in juvenile and young adult testes of RARα+/+ and RARα−/− mice. Histological sections of testes from RARα+/+ (A, B) and RARα−/− males (C–H) were immunostained with anti-activated caspase-3 antibody and were assessed by TUNEL assay. A–H, × 40. P, primary spermatocytes. Left panel shows the section after TUNEL assay while the right panel shows the activated caspase-3-labeled cells in the next serial section of the corresponding age. Arrows in C and D point to the TUNEL-positive and activated caspase-3-labeled spermatogenic cells, respectively. Arrowheads in C point to the TUNEL-positive spermatogenic cells at the site of cells collectively undergoing apoptosis.
Fig. 5.

Activated caspase-3 expression pattern during testicular development in RARα+/+ and RARα−/− testes. A bar graph indicating the number of activated caspase-3-positive germinal cells at various intervals after birth in RARα+/+ and RARα−/− mice (100 tubules scored per testis: three mice for each time point). The total number of activated caspase-3-labeled germ cells per 100 seminiferous tubules was counted in testicular sections of 2-4-, 6-, 8-week-old RARα+/+ and RARα−/− males. The bars represent the mean ± SD of three mice for each time point. (*) indicates significant differences within the same age group as well as between age groups by ANOVA, p<0.05. (**) indicates significant differences within the same age group as well as between age groups by ANOVA, p<0.01. (***) indicates significant differences within the same age group as well as between age groups by ANOVA, p<0.0001.
We then quantified the total number of activated caspase-3-labeled germ cells per 100 seminiferous tubules in both normal and mutant testes at 2–4, 6 and 8 weeks of age. In control testes, the number of activated ca-spase-3-labeled germ cells peaked at 3 weeks (Figs. 4B and 5), in agreement with observations of others using Western immunoblot analysis (Jahnukainen et al., 2004). This suggested that the age-related peak of apoptosis at 3 weeks of age in wild-type testes as revealed by TUNEL assay (Figs. 3A and 4A) was mediated by activated caspase-3 (Figs. 5 and 4B). In RARα−/− testes, the highest amount of activated caspase-3-labeled germ cells was observed at 2-3 weeks (Figs. 4D, 4F and 5), which correlated with the postnatal peaks of apoptosis as detected by TUNEL assay (Figs. 4C, 4E, 4F and 3A). The number of activated caspase-3-labeled cells is significantly diminished in both RARα+/+ and RARα−/− testes after 3 weeks (Fig. 5). Although there is a consistent high number of TUNEL-positive germ cells (mostly primary spermatocytes) observed in 6-week-old mutant testes (Fig. 3A), activated caspase-3 is not obvious in these cells, suggesting that apoptosis of these cells may be effected via a caspase-3 independent pathway.
Discussion
In the VAD rat testis, at least three major defects in spermatogenesis that are believed to be involved in severe testicular degeneration have been reported (Ismail et al., 1990; van Pelt and de Rooij, 1990; reviewed in de Rooij et al., 1994; Packer and Wolgemuth, 1999; Chung and Wolgemuth, 2004). They include failure of A1 spermatogonia to differentiate into A2 spermatogonia, a delay in the onset of and an abnormality in the progression of meiotic prophase, and spermatid degeneration. Spermatid degeneration has further been associated with a delay in spermiation and an up-regulation of the transcript of all-trans-RA-responsive gene, sulfated glycoprotein 2/clusterin at stage VIII (Huang and Marshall, 1983; Morales and Griswold, 1991) and a disruption of Sertoli cell tight junctions (Huang et al., 1988). We have observed that elongated spermatids do not properly align at the surface of tubular lumen for spermiation in RARα−/− testes (Chung et al., 2004). We now note both defects in spermatid release and abnormalities in orientation of spermatids relative to the basal aspect of Sertoli cells at stage VIII* RARα−/− testes. We speculate that the failure in orientation of the spermatids relative to the Sertoli cells affected the subsequent entrenchment of spermatid heads, which normally occurs in stage II–IV seminiferous epithelium (Oakberg, 1956; Ross, 1976; Russell, 1977b, 1993; Russell et al., 1983b). We further propose that the failure of this crucial alignment within the seminiferous epithelium may contribute to the abnormal spermiation observed in RARα−/− testes. An analogous failure in spermatid orientation and entrenchment of spermatids was observed in procarbazine-treated rat testes, which involved specific defects in the acrosomes of the cap phase of developing spermatids (Russell et al., 1983a, 1983b).
As the developing spermatids become situated in apical invaginations or crypts of the Sertoli cells, ectoplasmic specializations (ES) are formed in Sertoli cells adjacent to spermatid heads. ES are specialized types of adhesion junctions, composed of distinct regions of the plasma membrane, elements of the cytoskeletal system, and components of the endoplasmic reticulum (Russell, 1993; Vogl et al., 2000). The appearance and/or disappearance of ES have been correlated temporally with three events of spermatogenesis, wherein changes in intercellular adhesion occur: (1) the positioning of newly elongating spermatids in Sertoli cell crypts during stage VIII; (2) the release of morphologically mature sperm cells from the apices of Sertoli cells during stage VIII and (3) the movement of preleptotene spermatocytes from basal to adluminal compartments of the epithelium during stage VII/VIII (Russell, 1977a, 1993; Vogl et al., 2000). The presence of an acrosome has been proposed to be involved in the alignment of the spermatid head with the ES, which in turn may influence the orientation and positioning of the late spermatids within the seminiferous epithelium and their position within recesses of the Sertoli cell (Russell et al., 1983b). Rajguru et al. (1982) used subcapsular injection of [3H]retinol and showed that vitamin A is in spermatids in association with the Golgi. As the spermatids mature, the Golgi develops into the acrosome. In bovine and rabbit sperm, high levels of vitamin A have been demonstrated in the acrosome and acrosomal membrane (Gambhir and Ahluwalia, 1975; Velazquez et al., 1975). Taking these observations collectively, we propose that there is an intimate relationship between RARα-mediated signaling pathway and the proper functioning of ES, possibly involving the acrosome.
In normal testis, spermatogonia and meiotic spermatocytes are the primary cells that undergo apoptosis and apoptotic haploid germ cells are rarely detected (Billig et al., 1995; Callard et al., 1995; Hikim et al., 1995; Shetty et al., 1996). Surprisingly, we observed spermatids undergoing apoptosis, at least as assessed by TUNEL-staining, apparently engulfed by Sertoli cells situated at the basal lamina. Huang and Marshall (1983) also observed the presence of late spermatids and residual bodies at the luminal edge of epithelia at stages later than stage VIII of the spermatogenic cycle in VAD rat testis, concomitant with defective spermiation of these spermatids. The similar findings of defects in spermiation in both models suggested that mechanisms leading to completion of spermiation are extremely sensitive to change in the status in vitamin A, most likely through RA and an RARα receptor-mediated pathway. Further, the decrease in the number of total elongated spermatids in older age may be due to the presence of vacuolar-like spaces and depletion of germ cells (Chung et al., 2004).
The chief effectors of the apoptotic cell death pathway are the caspase family of cysteine proteases (Earnshaw et al., 1999; Said et al., 2004). The initiator caspases (caspase-2, -8, and -9) cleave and activate the executor caspases (caspase-3, -6, and -7), which cause cellular collapse by cleaving a specific set of protein substrates. Activated caspase-3 has been reported to be absent or present in insignificant levels in the normal adult testes (Kim et al., 2001; Omezzine et al., 2003). However, TUNEL-positive, activated caspase-3 positive spermatocytes were also observed in adult rat testes following an LH-suppressive treatment (Kim et al., 2001). Further, in utero exposure to flutamide has been reported to induce in the adult rat testes a chronic apoptotic germ cell death process associated with a long-term increase in the expression and activation of caspase-3 in round spermatids at stages VII, VIII and IX (Omezzine et al., 2003).
Our systematic analysis of activated caspase-3 expression during the first wave of spermatogenesis in both normal and mutant testes at different postnatal ages surprisingly revealed the presence of activated caspase-3 in the normal testes in the first wave of spermatogenesis in juvenile mice. Activated caspase-3 protein may therefore be involved in the age-related peak of apoptosis in the first wave of spermatogenesis in both wild-type and mutant testes. In addition, the detection of significantly higher number of activated ca-spase-3 positive germ cells in 2-week-old mutant testes may be due to the fact that caspase-3 is involved in the loss of primary spermatocytes in the first wave of spermatogenesis that we observed previously as early as at 2 weeks of age in RARα−/− mice (Chung et al., 2004).
Induction of apoptosis and DNA degradation as evidenced by TUNEL-positive staining, in mouse sperm and avian erythroid cells, two cell types that contain nuclei that are transcriptionally inactive, appears to be caspase independent (Weil et al., 1998). That is, the death of spermatozoa, which have little cytoplasm, may not involve the classical apoptotic machinery, since caspases do not appear to be used (Weil et al., 1998). Our observation of a lack of activated caspase-3 in the TUNEL-positive elongated spermatids in the mutant mice suggested that the apparent apoptosis observed in elongated spermatids may be via a caspase-independent pathway. Caspase-independent cell death can nonetheless involve mitochondria, which play an important role via the release of proapoptotic proteins (Cregan et al., 2004; Lorenzo and Susin, 2004). The intermembrane space of mitochondria contains proteins such as apotosis-inducing factor and endonuclease G, both of which provoke program cell death in a caspase-independent manner (Wang et al., 2002). Accordingly, if vertebrate cells have a caspase-independent death program(s), it may serve as a backup suicide mechanism. If this is the case, RARα−/− testes could provide an attractive system in which to identify the components of such a program.
A distinct feature of the appearance of TUNEL-positive germ cells in the first wave of spermatogenesis of RARα-deficient testes was the presence of TUNEL-positive elongating spermatids suggesting that their demise is a primary effect of the lack of expression of the RARα gene. This revealed a checkpoint of RARα function at the transition of round spermatids to elongating spermatids during spermiogenesis, when marked apoptosis was observed. The decision by elongating spermatids to undergo apoptosis, as assessed by TUNEL-staining, is unique to RARα-deficiency because the only TUNEL-positive cells detected in VAD testes were early meiotic germ cells and pachytene spermatocytes, but not elongated spermatids (Akmal et al., 1998). Although various and specific points of arrest of spermatogenesis in genetically modified and mutant mice have been reported (de Rooij and de Boer, 2003), this is, to our knowledge, the first suggestion that apoptosis may serve as a mechanism for eliminating these aberrant, abnormally positioned elongating spermatids in vivo.
Our current studies have revealed the crucial role of RARα in spermiogenesis, which involves intricate interactions between germ cells and Sertoli cells as spermatids attach to the epithelium and orient prior to release into the lumen. The observations raise the possibility that there may be differences in the spermatogenic phenotypes observed in animals in which RARα is disrupted only in the germ line or only in somatic cells as compared to the present model wherein it is mutated in both the germ line and somatic cells since the onset of gonadal development. We anticipate that this approach will be very powerful in highlighting specific functions and pathways in which the receptor acts, pathways that are obscured when function is blocked in a ubiquitous manner, as in total null animals.
Acknowledgments
We thank Dr. Weng Kong Sung for expert technical advice and the Animal Core of the P01 grant under the direction of Dr. Cathy Mendelsohn for breeding some of the RARα mutant mice used in initial stages of this study. We thank Dr. Pierre Chambon for the generous gift of RARα mutant mice and Drs. P. Chambon, M. Mark and N.B. Ghyselinck for helpful discussion. This work was supported by P01 DK54057, Project 5 and Core B, to D.J.W. and fellowships from the Croucher Foundation, Hong Kong to S.S.W.C.
Contributor Information
Sanny S.W. Chung, Department of Genetics and Development, Columbia University Medical Center, 630W, 168th Street, New York, NY 10032, USA, Fax: (212) 305-6084
Xiangyuan Wang, Department of Genetics and Development, Columbia \University Medical Center, 630W, 168th Street, New York, NY 10032, USA, Fax: (212) 305-6084.
Debra J. Wolgemuth, Email: djw3@columbia.edu, Department of Genetics and Development, Columbia University Medical Center, 630W, 168th Street, New York, NY 10032, USA, Fax: (212) 305-6084; Department of Obstetrics and Gynecology, Columbia University Medical Center, New York, NY 10032, USA; The Center for Reproductive Sciences, Columbia University Medical Center, New York, NY 10032, USA; The Institute of Human Nutrition, Columbia University Medical Center, New York, NY 10032, USA; The Herbert Irving Comprehensive Cancer Center, Columbia University Medical Center, New York, NY 10032, USA.
References
- Akmal KM, Dufour JM, Kim KH. Retinoic acid receptor alpha gene expression in the rat testis: potential role during the prophase of meiosis and in the transition from round to elongating spermatids. Biol Reprod. 1997;56:549–556. doi: 10.1095/biolreprod56.2.549. [DOI] [PubMed] [Google Scholar]
- Akmal KM, Dufour JM, Vo M, Higginson S, Kim KH. Ligand-dependent regulation of retinoic acid receptor alpha in rat testis: in vivo response to depletion and repletion of vitamin A. Endocrinology. 1998;139:1239–1248. doi: 10.1210/endo.139.3.5775. [DOI] [PubMed] [Google Scholar]
- Alnemri ES, Livingston DJ, Nicholson DW, Salvesen G, Thornberry NA, Wong WW, Yuan J. Human ICE/CED-3 protease nomenclature. Cell. 1996;87:171. doi: 10.1016/s0092-8674(00)81334-3. [DOI] [PubMed] [Google Scholar]
- Beumer TL, Roepers-Gajadien HL, Gademan LS, Rutgers DH, de Rooij DG. P21(Cip1/WAF1) expression in the mouse testis before and after X irradiation. Mol Reprod Dev. 1997;47:240–247. doi: 10.1002/(SICI)1098-2795(199707)47:3<240::AID-MRD2>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- Billig H, Furuta I, Rivier C, Tapanainen J, Parvinen M, Hsueh AJ. Apoptosis in testis germ cells: developmental changes in gonadotropin dependence and localization to selective tubule stages. Endocrinology. 1995;136:5–12. doi: 10.1210/endo.136.1.7828558. [DOI] [PubMed] [Google Scholar]
- Callard GV, Jorgensen JC, Redding JM. Biochemical analysis of programmed cell death during premeiotic stages of spermatogenesis in vivo and in vitro. Dev Genet. 1995;16:140–147. doi: 10.1002/dvg.1020160207. [DOI] [PubMed] [Google Scholar]
- Chung SS, Sung W, Wang X, Wolgemuth DJ. Retinoic acid receptor alpha is required for synchronization of spermatogenic cycles and its absence results in progressive breakdown of the spermatogenic process. Dev Dyn. 2004;230:754–766. doi: 10.1002/dvdy.20083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung SS, Wolgemuth DJ. Role of retinoid signaling in the regulation of spermatogenesis. Cytogenet Genome Res. 2004;105:189–202. doi: 10.1159/000078189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cregan SP, Dawson VL, Slack RS. Role of AIF in caspase-dependent and caspase-independent cell death. Oncogene. 2004;23:2785–2796. doi: 10.1038/sj.onc.1207517. [DOI] [PubMed] [Google Scholar]
- de Rooij DG, de Boer P. Specific arrests of spermatogenesis in genetically modified and mutant mice. Cytogenet Genome Res. 2003;103:267–276. doi: 10.1159/000076812. [DOI] [PubMed] [Google Scholar]
- de Rooij DG, van Pelt AMM, Van de Kant HJG, van der Saag PT, Peters AHFM, Heyting C, de Boer P. Role of retinoids in spermatogonial proliferation and differentiation and the meiotic prophase. In: Bartke A, editor. Function of somatic cells in the testis. Springer; Berlin: 1994. p. 345. [Google Scholar]
- Earnshaw WC, Martins LM, Kaufmann SH. Mammalian caspases: structure, activation, substrates, and functions during apoptosis. Annu Rev Biochem. 1999;68:383–424. doi: 10.1146/annurev.biochem.68.1.383. [DOI] [PubMed] [Google Scholar]
- Eskild W, Hansson V. Vitamin A functions in the reproductive organs. In: Blomhoff R, editor. Vitamin A in health and disease. Dekker; New York: 1994. pp. 531–559. [Google Scholar]
- Eskild W, Ree AH, Levy FO, Jahnsen T, Hansson V. Cellular localization of mRNAs for retinoic acid receptor-alpha, cellular retinol-binding protein, and cellular retinoic acid-binding protein in rat testis: evidence for germ cell-specific mRNAs. Biol Reprod. 1991;44:53–61. doi: 10.1095/biolreprod44.1.53. [DOI] [PubMed] [Google Scholar]
- Gambhir KK, Ahluwalia BS. Vitamin A in bovine sperm acrosomes. J Reprod Fertil. 1975;43:129–132. doi: 10.1530/jrf.0.0430129. [DOI] [PubMed] [Google Scholar]
- Henriksen K, Hakovirta H, Parvinen M. In-situ quantification of stage-specific apoptosis in the rat seminiferous epithelium: effects of short-term experimental cryptorchidism. Int J Androl. 1995;18:256–262. [PubMed] [Google Scholar]
- Hikim AP, Wang C, Leung A, Swerdloff RS. Involvement of apoptosis in the induction of germ cell degeneration in adult rats after gonadotropin-releasing hormone antagonist treatment. Endocrinology. 1995;136:2770–2775. doi: 10.1210/endo.136.6.7750502. [DOI] [PubMed] [Google Scholar]
- Huang HF, Marshall GR. Failure of spermatid release under various vitamin A states—an indication of delayed spermiation. Biol Reprod. 1983;28:1163–1172. doi: 10.1095/biolreprod28.5.1163. [DOI] [PubMed] [Google Scholar]
- Huang HF, Yang CS, Meyenhofer M, Gould S, Boccabella AV. Disruption of sustentacular (Sertoli) cell tight junctions and regression of spermatogenesis in vitamin-A-deficient rats. Acta Anat (Basel) 1988;133:10–15. doi: 10.1159/000146606. [DOI] [PubMed] [Google Scholar]
- Ismail N, Morales C, Clermont Y. Role of spermatogonia in the stage-synchronization of the seminiferous epithelium in vitamin-A-deficient rats. Am J Anat. 1990;188:57–63. doi: 10.1002/aja.1001880107. [DOI] [PubMed] [Google Scholar]
- Jahnukainen K, Chrysis D, Hou M, Parvinen M, Eksborg S, Soder O. Increased apoptosis occurring during the first wave of spermatogenesis is stage-specific and primarily affects midpachytene spermatocytes in the rat testis. Biol Reprod. 2004;70:290–296. doi: 10.1095/biolreprod.103.018390. [DOI] [PubMed] [Google Scholar]
- Kastner P, Mark M, Chambon P. Nonsteroid nuclear receptors: what are genetic studies telling us about their role in real life? Cell. 1995;83:859–869. doi: 10.1016/0092-8674(95)90202-3. [DOI] [PubMed] [Google Scholar]
- Kastner P, Mark M, Leid M, Gansmuller A, Chin W, Grondona JM, Decimo D, Krezel W, Dierich A, Chambon P. Abnormal spermatogenesis in RXR beta mutant mice. Genes Dev. 1996;10:80–92. doi: 10.1101/gad.10.1.80. [DOI] [PubMed] [Google Scholar]
- Kim JM, Ghosh SR, Weil AC, Zirkin BR. Caspase-3 and caspase-activated deoxyribonuclease are associated with testicular germ cell apoptosis resulting from reduced intratesticular testosterone. Endocrinology. 2001;142:3809–3816. doi: 10.1210/endo.142.9.8375. [DOI] [PubMed] [Google Scholar]
- Kim KH, Griswold MD. The regulation of retinoic acid receptor mRNA levels during spermatogenesis. Mol Endocrinol. 1990;4:1679–1688. doi: 10.1210/mend-4-11-1679. [DOI] [PubMed] [Google Scholar]
- Kim KH, Wang ZQ. Action of vitamin A on testis: role of the Sertoli cell. In: Griswold MD, Rusell LD, editors. The sertoli cell. Cache River Press; Clearwater, FL: 1993. pp. 514–535. [Google Scholar]
- Liu D, Matzuk MM, Sung WK, Guo Q, Wang P, Wolgemuth DJ. Cyclin A1 is required for meiosis in the male mouse. Nat Genet. 1998;20:377–380. doi: 10.1038/3855. [DOI] [PubMed] [Google Scholar]
- Lorenzo HK, Susin SA. Mitochondrial effectors in caspase-independent cell death. FEBS Lett. 2004;557:14–20. doi: 10.1016/s0014-5793(03)01464-9. [DOI] [PubMed] [Google Scholar]
- Lufkin T, Lohnes D, Mark M, Dierich A, Gorry P, Gaub MP, LeMeur M, Chambon P. High postnatal lethality and testis degeneration in retinoic acid receptor alpha mutant mice. Proc Natl Acad Sci USA. 1993;90:7225–7229. doi: 10.1073/pnas.90.15.7225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mangelsdorf DJ, Evans RM. The RXR heterodimers and orphan receptors. Cell. 1995;83:841–850. doi: 10.1016/0092-8674(95)90200-7. [DOI] [PubMed] [Google Scholar]
- Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, Blumberg B, Kastner P, Mark M, Chambon P, Evans RM, et al. The nuclear receptor superfamily: the second decade. Cell. 1995;83:835–839. doi: 10.1016/0092-8674(95)90199-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morales CR, Griswold MD. Variations in the level of transferrin and SGP-2 mRNAs in Sertoli cells of vitamin A-deficient rats. Cell Tissue Res. 1991;263:125–130. doi: 10.1007/BF00318407. [DOI] [PubMed] [Google Scholar]
- Mutter GL, Grills GS, Wolgemuth DJ. Evidence for the involvement of the proto-oncogene c-mos in mammalian meiotic maturation and possibly very early embryogenesis. EMBO J. 1988;7:683–689. doi: 10.1002/j.1460-2075.1988.tb02863.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicholson DW, Thornberry NA. Caspases: killer proteases. Trends Biochem Sci. 1997;22:299–306. doi: 10.1016/s0968-0004(97)01085-2. [DOI] [PubMed] [Google Scholar]
- Nicholson DW, Thornberry NA. Apoptosis. Life and death decisions. Science. 2003;299:214–215. doi: 10.1126/science.1081274. [DOI] [PubMed] [Google Scholar]
- Oakberg EF. A description of spermiogenesis in the mouse and its use in an analysis of the cycle of the seminiferous epithelium and germ cell renewal. Am J Anat. 1956;99:391–414. doi: 10.1002/aja.1000990303. [DOI] [PubMed] [Google Scholar]
- Omezzine A, Chater S, Mauduit C, Florin A, Tabone E, Chuzel F, Bars R, Benahmed M. Long-term apoptotic cell death process with increased expression and activation of caspase-3 and -6 in adult rat germ cells exposed in utero to flutamide. Endocrinology. 2003;144:648–661. doi: 10.1210/en.2002-220692. [DOI] [PubMed] [Google Scholar]
- Packer AI, Wolgemuth DJ. Genetic and molecular approaches to understanding the role of retinoids in mammalian spermatogenesis. In: Nau H, Blaner WS, editors. Retinoids: the biochemical and molecular basis of vitamin A and retinoid action. Springer-Verlag; Berlin: 1999. pp. 347–368. [Google Scholar]
- Rajguru SU, Kang YH, Ahluwalia BS. Localization of retinol (vitamin A) in rat testes. J Nutr. 1982;112:1881–1891. doi: 10.1093/jn/112.10.1881. [DOI] [PubMed] [Google Scholar]
- Rodriguez I, Ody C, Araki K, Garcia I, Vassalli P. An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. EMBO J. 1997;16:2262–2270. doi: 10.1093/emboj/16.9.2262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross MH. The Sertoli cell junctional specialization during spermiogenesis and at spermiation. Anat Rec. 1976;186:79–104. [Google Scholar]
- Russell L. Movement of spermatocytes from the basal to the adluminal compartment of the rat testis. Am J Anat. 1977a;148:313–328. doi: 10.1002/aja.1001480303. [DOI] [PubMed] [Google Scholar]
- Russell L. Observations on rat Sertoli ectoplasmic (‘junctional’) specializations in their association with germ cells of the rat testis. Tissue Cell. 1977b;9:475–498. doi: 10.1016/0040-8166(77)90007-6. [DOI] [PubMed] [Google Scholar]
- Russell LD. Role in spermiation. In: Griswold MD, Russell LD, editors. The sertoli cell. Cache River Press; Clear-water, FL: 1993. pp. 269–303. [Google Scholar]
- Russell LD, Ettlin RA, SinhaHikim AP, Clegg ED. Histological and histopathological evaluation of the testis. Cache River Press; Clearwater, FL: 1990. [Google Scholar]
- Russell LD, Lee IP, Ettlin R, Malone JP. Morphological pattern of response after administration of procarbazine: alteration of specific cell associations during the cycle of the seminiferous epithelium of the rat. Tissue Cell. 1983a;15:391–404. doi: 10.1016/0040-8166(83)90071-x. [DOI] [PubMed] [Google Scholar]
- Russell LD, Lee IP, Ettlin R, Peterson RN. Development of the acrosome and alignment, elongation and entrenchment of spermatids in procarbazine-treated rats. Tissue Cell. 1983b;15:615–626. doi: 10.1016/0040-8166(83)90011-3. [DOI] [PubMed] [Google Scholar]
- Said TM, Paasch U, Glander HJ, Agarwal A. Role of caspases in male infertility. Hum Reprod Update. 2004;10:39–51. doi: 10.1093/humupd/dmh003. [DOI] [PubMed] [Google Scholar]
- Shetty J, Marathe GK, Dighe RR. Specific immunoneutralization of FSH leads to apoptotic cell death of the pachy-tene spermatocytes and spermatogonial cells in the rat. Endocrinology. 1996;137:2179–2182. doi: 10.1210/endo.137.5.8612566. [DOI] [PubMed] [Google Scholar]
- Shi SR, Key ME, Kalra KL. Antigen retrieval in formalin-fixed, paraffin-embedded tissues: an enhancement method for immunohistochemical staining based on microwave oven heating of tissue sections. J Histochem Cytochem. 1991;39:741–748. doi: 10.1177/39.6.1709656. [DOI] [PubMed] [Google Scholar]
- Sjoblom T, West A, Lahdetie J. Apoptotic response of spermatogenic cells to the germ cell mutagens etoposide, ad-riamycin, and diepoxybutane. Environ Mol Mutagen. 1998;31:133–148. doi: 10.1002/(sici)1098-2280(1998)31:2<133::aid-em5>3.0.co;2-n. [DOI] [PubMed] [Google Scholar]
- van Pelt AM, de Rooij DG. Synchronization of the seminiferous epithelium after vitamin A replacement in vitamin A-deficient mice. Biol Reprod. 1990;43:363–367. doi: 10.1095/biolreprod43.3.363. [DOI] [PubMed] [Google Scholar]
- van Pelt AM, van den Brink CE, de Rooij DG, van der Saag PT. Changes in retinoic acid receptor messenger ribonucleic acid levels in the vitamin A-deficient rat testis after administration of retinoids. Endocrinology. 1992;131:344–350. doi: 10.1210/endo.131.1.1319320. [DOI] [PubMed] [Google Scholar]
- Velazquez A, Rosado A, Jr, Rosado A. Participation of vitamin A in the maturation of rabbit spermatozoa. Int J Fertil. 1975;20:151–151. [PubMed] [Google Scholar]
- Vogl AW, Pfeiffer DC, Mulholland D, Kimel G, Guttman J. Unique and multifunctional adhesion junctions in the testis: ectoplasmic specializations. Arch Histol Cytol. 2000;63:1–15. doi: 10.1679/aohc.63.1. [DOI] [PubMed] [Google Scholar]
- Wang X, Yang C, Chai J, Shi Y, Xue D. Mechanisms of AIF-mediated apoptotic DNA degradation in Caenorhabditis elegans. Science. 2002;298:1587–1592. doi: 10.1126/science.1076194. [DOI] [PubMed] [Google Scholar]
- Weil M, Jacobson MD, Raff MC. Are caspases involved in the death of cells with a transcriptionally inactive nucleus?. Sperm and chicken erythrocytes. J Cell Sci. 1998;111:2707–2715. doi: 10.1242/jcs.111.18.2707. [DOI] [PubMed] [Google Scholar]
- Yan W, Suominen J, Samson M, Jegou B, Toppari J. Involvement of Bcl-2 family proteins in germ cell apoptosis during testicular development in the rat and pro-survival effect of stem cell factor on germ cells in vitro. Mol Cell Endocrinol. 2000;165:115–129. doi: 10.1016/s0303-7207(00)00257-4. [DOI] [PubMed] [Google Scholar]