

Abstract
Spatial and temporal variability in cyanobacterial community composition (CCC) within and between eutrophic lakes is not well-described using culture independent molecular methods. We analyzed CCC across twelve locations in four eutrophic lakes and within-lake locations in the Yahara Watershed, WI, on a weekly basis, for 5 months. Taxa were discriminated by length of MspI-digested cpcB/A intergenic spacer gene sequences and identified by comparison to a PCR-based clone library. CCC across all stations was spatially segregated by depth of sampling locations (ANOSIM R = 0.23, p < 0.001). Accordingly, CCC was correlated with thermal stratification, nitrate and soluble reactive phosphorus (SRP, R = 0.2-0.3). Spatial variability in CCC and temporal trends in taxa abundances were rarely correlative between sampling locations in the same lake indicating significant within lake spatiotemporal heterogeneity. Across all stations, a total of 37 bloom events were observed based on distinct increases in phycocyanin. Out of 97 taxa, a single Microcystis , and two different Aphanizomenon taxa were the dominant cyanobacteria detected during bloom events. The Microcystis and Aphanizomenon taxa rarely bloomed together and were significantly anti-correlated with each other at 9 of 12 stations with Pearson R values of -0.6 to -0.9 (p < 0.001). Of all environmental variables measured, nutrients, especially nitrate were significantly greater during periods of Aphanizomenon dominance while the nitrate+nitrite:SRP ratio was lower. This study shows significant spatial variability in CCC within and between lakes structured by depth of the sampling location. Furthermore, our study reveals specific genotypes involved in bloom formation. More in-depth characterization of these genotypes should lead to a better understanding of factors promoting bloom events in these lakes and more reliable bloom prediction models.
Introduction
In many eutrophic lakes, cyanobacteria are responsible for massive accumulations of biomass, otherwise known as "blooms," whether formed through growth, horizontal and vertical migration, or physical forcings (e.g. wind) [1,2]. The incidence and severity of these blooms are thought to be escalating, particularly in the northern hemisphere, due to increased eutrophication of waterways and climate change [3-5]. During summer in northern temperate eutrophic lakes, the most commonly encountered genera include Microcystis , Anabaena , and Aphanizomenon , while Phormidinium, Planktothrix , Gloeotrichia and others occur sporadically [6,7]. In addition, invasive species from tropical/subtropical regions, including Cylindrospermopsis raciborskii have been detected recently in northern temperate eutrophic lakes, including this study [6,8]. While a natural occurrence, these blooms are exacerbated by human impacts and potentially problematic as they may lead to a decline in dissolved oxygen, produce undesirable odors and/or contain toxins harmful to humans, fish and other wildlife. For this reason, lake restoration efforts and mandates by the U.S. Environmental Protection Agency (e.g. total maximum daily loads program) are aimed at reducing the intensity and frequency of cyanobacterial blooms.
The success of freshwater cyanobacteria is attributed to a variety of intersecting circumstances—including warm water temperatures, pH buffering above neutrality, nitrogen fixation, vertical movement via gas vacuoles, carbon/phosphorus/nitrogen storage mechanisms, and colony formation that inhibits predation [9-13]—which may give them a competitive advantage over eukaryotic photoautotrophs [14]. These traits vary whether characterized by species, strain, and/or genotype [15,16]. Species interactions, including competition and synergism, may also be important for bloom development and/or toxin production since changes in specific genotypes of the same genera are observed during bloom formation and decline [17-19]. In addition, new nitrogen input from nitrogen fixing cyanobacteria may be important for growth and/or toxin production by non-nitrogen fixing cyanobacteria such as Microcystis [20].
Variability in cyanobacterial community composition (CCC) has been studied in lakes for decades resulting in a rich understanding of environmental factors involved in promoting their growth (reviewed by [2]). However, it is difficult to predict within any given season the timing and severity of bloom events that may form over the course of days. Furthermore, the majority of information about the ecology of cyanobacteria comes from studies relying upon culturing or microscopic identifications. Recent studies show that such techniques do not reflect the bulk of cyanobacterial diversity in lakes and thus may group functionally distinct taxa as one [21]. This is also indicated by laboratory studies, which show that strains or genotypes of the same species differ in their phenotypic responses to environmental cues (e.g. light, nutrients, colony formation) [15,16,22]. Recent field studies show shifts in genotypes of the dominant taxa of the same genera over spatial, temporal, or chemical/physical gradients [17,19,23,24]. Therefore, an analysis of CCC using culture- independent molecular methods may lead to a better understanding of taxa or genotypes responsible for bloom events and the conditions under which these occur. However, few or no studies have analyzed culture- independent molecular diversity of cyanobacteria across multiple lakes and bloom events at resolved time scales (e.g. weekly).
In this study, we investigated variability in CCC on a weekly basis over the course of thirty-seven cyanobacterial bloom events occurring at twelve sampling locations in four eutrophic lakes of the Yahara Watershed, Wisconsin, USA. We used a culture-independent molecular method to characterize CCC and asked whether variability in CCC tracked with physical and chemical lake characteristics both spatially (i.e. within and between lakes) and temporally. In addition, we compared CCC and environmental conditions between bloom events to determine the number and identity of taxa responsible for each bloom event, and the prevailing environmental conditions under which blooms occurred by these cyanobacterial taxa. In the year this study was conducted (2008), Southern Wisconsin experienced the highest precipitation on record over a 24-day period in May-June resulting in record high stream flows (500 years) and significant flooding [25].
Materials and Methods
No specific permits were required for the described field study, nor were any specific permissions needed to sample the locations. None of the lakes sampled are privately owned, and our sampling did not involve endangered or protected species.
Description of study stations and sampling
The Yahara River connects a chain of four lakes within the Southern portion of the Yahara watershed in South Central Wisconsin, USA (Figure 1). Lake Mendota at the top of the chain is the largest, followed by Monona, Waubesa (not sampled in this study) and Kegonsa. Lake Wingra is a smaller lake situated apart from the lake chain and is fed by mainly urban runoff and a natural spring [26]. It empties into Lake Monona and is the only lake in this study that does not receive water from the Yahara River or other lakes. All of the lakes are eutrophic and have been heavily impacted by nutrients since the capital city of Madison was settled in the early 1800’s. Lake Mendota has been the subject of scientific studies for nearly a century [27]. No permits were required to sample these lakes as they are in the public domain.
Figure 1. Sampling locations in lakes of the Yahara Watershed.
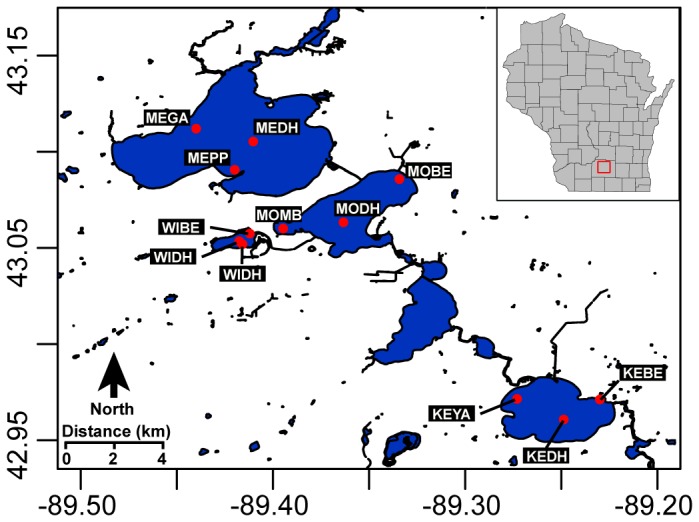
Three locations in each of four lakes were sampled on a weekly basis late May through October.
Integrated samples of the water column receiving photosynthetically active radiation (photic zone depth, PZD, estimated by multiplying secchi disk depth by 1.7 [28]) were taken weekly from three locations in each lake, from late May through October 2008 using a 2" diameter PVC pipe as previously described [29]. On average, 60 samples were taken from each lake on 20 different dates. A buoy was deployed in Lake Mendota as part of the Global Lakes Ecological Observatory Network (www.gleon.org). The buoy was equipped with a phycocyanin fluorometric sensor (Cyclops 7, Turner Designs), data logger (CR-1000, Campbell Scientific) and an RS-232 modem sending data to a shore base station in near real-time. Additional samples were taken from this location when the phycocyanin fluorescent signal was high (steady increase to >2 volts) suggesting that a bloom event may be underway. Water samples were transported back to the laboratory on ice and filtered through a 47 mm, 0.2 µm pore size, polyethersulfone membrane (Supor-200, Pall Corporation). Filters were placed into 2 ml cryogenic tubes containing 1 g of autoclaved 2 mm glass beads and stored at -80 °C for DNA extraction. A 50 ml aliquot of sample water was amended with 0.05 ml of concentrated HCl and stored at 4 °C for measurement of total phosphorus (TP) and 50 ml of water was filtered through a 0.7 µm glass fiber filter (GF/F, Whatman) and stored at -20 °C for measurement of soluble reactive phosphorus (SRP), nitrate, and nitrite.
Analytical procedures
SRP was measured in filtered samples by the ascorbic acid method 4500 PE [30]. TP was measured as SRP in unfiltered, acidified samples following persulfate digestion in an autoclave for 1 hour at 120°C [30]. Nitrate and nitrite were measured in filtered samples by HPLC as previously described [31]. Phycocyanin was extracted from filters by bead beating in 20 mM sodium acetate (pH 5.5). The extract was centrifuged to remove cell debris and phycocyanin was measured spectrophotometrically at 620 nm with correction at 650 nm [32].
DNA extraction and automated phycocyanin intergenic spacer analysis (APISA)
CCC was determined based on sizing of MspI-digested PC-IGS gene fragments on an automated sequencer, which we call automated phycocyanin intergenic spacer analysis or APISA. It is a variation on terminal restriction fragment length polymorphism analysis, except that the size of fragments is based on the MspI site as well as length of the intergenic spacer. Sizing on an automated sequencer allows for discriminating fragments that differ by 1-4 bp and an estimate of relative abundance. Briefly, DNA was extracted directly from filters using a xanthogenate-phenol-chloroform protocol previously described [6]. The cpcB/A genes plus intergenic spacer region were amplified from total community DNA using primers previously published [33]. Each 50 µl PCR consisted of 2 µl template DNA, 250 µM dNTP’s, 0.4 µM of each primer, 3 mM MgCl2, 0.25 mg/ml BSA, 1 unit of GoTaq Polymerase (Promega), and 5 µl of 10X buffer supplied with Taq. The thermocycler protocol was as previously described [33]. The PCR products were precipitated and desalted with absolute ethanol and ammonium acetate, respectively, and subjected to digestion with MspI restriction endonuclease (Promega) for 2 hours at 37 °C and the reaction stopped at 65°C for 10 min. The digested PCR products were desalted, precipitated, and resuspended in 20 µl of ddH2O. A 3 µl aliquot was mixed with 10 µl of formamide and 0.4 µl of a ROX labeled (Carboxy-X-rhodamine) custom size standard (Bioventures) containing DNA fragments ranging from 30-1250 bp. The APISA DNA fragments were sized by capillary electrophoresis using an ABI 3700 sequencer as previously described [34].
Analysis of APISA data
Electropherograms were aligned and analyzed using Genemarker® v1.75 as previously described [35]. Peak size of unknowns was determined based on mobility of size standards using the local southern method implemented within Genemarker®. Peaks were binned into operational taxonomic units (OTUs) by visually inspecting a trace overlay of all samples. Individual runs were manually checked to ensure correct calling of peaks. An arbitrary cutoff of 100 relative fluorescent units (RFUs) was used to distinguish signal from noise and only runs generating maximum peak heights of at least 3,000 RFUs were accepted. The absolute height of peaks was divided by the sum of peak heights within each run to calculate relative peak height, which was used as an estimate of relative abundance [34].
An attempt was made to assign OTU’s to genera based on comparison to a library of PC-IGS sequences. We previously provided a sequence alignment and phylogenetic analysis of 271 PC-IGS sequences recovered from the same samples as those analyzed in this study [6]. An additional 276 reference PC-IGS sequences from Genbank were included in this alignment in order to assign sequences to genus. Sequences were aligned in the ARB software environment [36] as previously described [6] and classified to genus based on their occurrence in a maximum likelihood tree relative to PC-IGS from well-characterized strains. The TRF Cut tool [37] was implemented within ARB to digest PC-IGS sequences in silico using the MspI restriction site. A list of terminal fragment sizes with their corresponding genus classification was then exported from ARB and used as a custom database in the Phylogenetic Assignment Tool (PAT) [38], which was used to assign a cyanobacterial genus to the OTU’s observed in our samples. For PAT analysis a size tolerance of 2 bp and 4 bp was selected for bins ranging in size from 100-250 bp, and >250 bp, respectively, based on the observation that peak width increases with elution time from the sequencer [38].
Identification of bloom events
For this study we considered a cyanobacterial bloom to be any accumulation of phycocyanin in the photic zone (i.e. the area of the water column sampled), mathematically defined as an increase in phycocyanin 1.5X above a baseline. Each point along the baseline was calculated as a simple moving average (SMA) using all previous observations, except those identified as occurring within a bloom. The SMA is defined as:
Where S N does not include bloom samples. Hence, the first point on the baseline is equal to the first observed phycocyanin concentration and all subsequent points are calculated as the SMA of all previously observed phycocyanin concentrations that were below 1.5X of the baseline. Excluding bloom samples from the SMA calculation prevents stepwise increases in the baseline after each bloom event, which otherwise would underestimate the number of bloom events occurring later in the year.
Statistics
All chemical and physical data were log transformed prior to all analyses. Correspondence (CA) and canonical correspondence analyses (CCA) were conducted in CANOCO v5.4 [39]. A matrix of relative abundances of each taxa by sample date was imported into CANOCO along with a matrix containing log- transformed chemical and physical data (i.e. environmental data) including TP, SRP, nitrate, nitrite, PZD, photic zone water temperature and DO (i.e. average water temperature and DO over the integrated depths sampled). In addition, the degree to which the water column was stratified at each location was estimated by the difference in water temperature at the surface minus temperature at the lake bottom (ΔT). For the CA and CCA’s, a response model was chosen using indirect or direct gradient analyses, respectively. Biplot scaling was selected with focusing on inter-sample distances. The OTU data was not further transformed and only one outlier was removed. An Analysis of Similarity (ANOSIM) was conducted in Primer v5.0 using a Bray-Curtis dissimilarity matrix. All descriptive statistics, Pearson R correlations, and Mann-Whitney tests were calculated in the R statistical package.
Results
Trends in chemical and physical lake variables
Trends in water temperature were similar across all stations and ranged from 11.5-30 °C over the sampling period. PZD became shallow at all stations early in June and remained shallow averaging 2.7 +/- 2.0 m for the rest of the sampling period (Figure S1). We defined shallow stations as <3.7 m and all others as deep locations, which were all greater than 10 m. At shallow locations PZD extended through nearly the entire water column on most sampling days (Figure S1). Thermal stratification (ΔT) dissipated at all stations in June, due to a larger storm event, and again in October, due to lake mixing in autumn, but remained weak at shallow stations for the entire sampling period (Table 1, Figure S1).
Table 1. Mean (+/- Standard Deviation) of physical and chemical lake characteristics.
| Site | Lake | Depth(m) | T°C | aΔT | bPZD | cNitrate(µg/L) | cNitrite(µg/L) | TP(µg/L) | SRP(µg/L) |
|---|---|---|---|---|---|---|---|---|---|
| MEDH | Mendota | 25.7 | 21.7 (3.8) | 9.7 (3.9) | 3.7 (2.7) | 190 (126) | 69 (46) | 49 (38) | 33 (45) |
| MEGA | Mendota | 18.1 | 20.1 (3.5) | 7.8 (4.0) | 6.0 (3.8) | 243 (208) | 56 (75) | 50 (32) | 63 (54) |
| MEPP | Mendota | 2.3 | 21.3 (2.1) | 0.7 (0.5) | 3.5 (0.9)* | 278 (208) | 94 (106) | 48 (35) | 34 (41) |
| MODH | Monona | 23.1 | 22.8 (3.6) | 7.3 (3.5) | 3.1 (2.1) | 82 (149) | 161 (253) | 35 (28) | 23 (19) |
| MOBE | Monona | 3.6 | 23.3 (3.0) | 0.9 (1.3) | 2.3 (0.6) | 67 (62) | 109 (227) | 50 (41) | 29 (29) |
| MOMB | Monona | 1.8 | 22.4 (4.9) | 0.9 (0.7) | 1.3 (0.7) | 56 (54) | 64 (111) | 61 (33) | 22 (25) |
| KEDH | Kegonsa | 9.6 | 22.8 (3.9) | 1.4 (1.4) | 2.7 (1.8) | 87 (33) | 65 (117) | 94 (45) | 47 (42) |
| KEBE | Kegonsa | 1.2 | 22.2 (4.3) | 0.4 (0.5) | 1.9 (0.6)* | 89 (170) | 70 (118) | 91 (31) | 36 (41) |
| KEYA | Kegonsa | 2.1 | 23.5 (3.6) | 0.7 (0.5) | 2.7 (0.8)* | 41 (37) | 43 (96) | 85 (34) | 40 (44) |
| WIDH | Wingra | 3.7 | 24.8 (3.8) | 1.1 (1.0) | 1.9 (0.6) | BDL | BDL | 20 (10) | 14 (7) |
| WIAR | Wingra | 2.8 | 23.6 (4.3) | 0.6 (0.6) | 1.8 (0.2) | BDL | BDL | 23 (13) | 13 (8) |
| WIBE | Wingra | 2.3 | 24.5 (3.6) | 0.7 (0.6) | 1.7 (0.6) | BDL | BDL | 23 (11) | 13 (5) |
ΔT is the difference in water temperature at the bottom and top of the lake at each site.
b PZD was estimated by multiplying secchi disk depth by 1.7. As such, average PZD calculated at some locations (*) extends beyond the maximum depth.
Nitrate and nitrite were below detection limits (BDL) in Lake Wingra on the majority of sample dates.
At all locations, dissolved nutrients were highest in June, well after spring mixing, possibly due to heavy rain events, and again in October due to lake mixing in autumn (Figure S1). Nutrients also followed similar trends across all lake locations with two exceptions. Nitrate and nitrite were below detection limits (0.01 mg/L) at all sampling locations in Lake Wingra for nearly the entire sampling period. In both Lake Kegonsa and Wingra, TP followed an opposite trend compared to the other lakes with highest concentrations in August and/or September, depending on lake location (Figure S1). Increases in TP in these lakes were presumably due to sediment resuspension in these shallower, polymictic lakes. At the north end of the lake chain, average concentrations of nitrate in Lake Mendota were 2-7 times higher than all other lakes (Table 1) while SRP was similar across Lakes Mendota, Monona, and Kegonsa, but significantly lower in Lake Wingra. The SRP may have been lower in Lake Wingra because this lake is not subject to agricultural runoff and a larger percentage of the lake supports macrophyte growth of Eurasian milfoil ( Myriophyllum spicatum ).
Identification of cyanobacteria
To identify cyanobacteria, the APISA OTUs were matched to phylogenetically characterized PC-IGS sequences digested with MspI in silico. There were 94 total OTUs detected, and nine of these could be matched to the PC-IGS sequences digested in silico with MspI (Figure 2). The nine OTUs identified were from Microcystis , Aphanizomenon , Chroococcus , and Cylindrospermopsis taxa, which agrees with our previous identification of cyanobacteria in these lakes based on PCR-based clone libraries [6]. One APISA OTU (i.e. AnaAphChr690) represented sequences from three different genera ( Anabaena , Aphanizomenon , and Chroococcus ). Therefore, the identity of this OTU is ambiguous among these three genera and was not included when summing relative abundances of genera.
Figure 2. Identification of cyanobacteria represented by APISA OTU’s.

Nine of the 94 unique PC-IGS terminal fragments (TRFs) detected among 197 sample could be matched by size to an in silico MspI digestion of PC-IGS sequences previously retrieved from the same samples. These were the most abundant TRFs detected representing 87.1 ± 13.5% of the total peak heights in any given sample indicating that these are the most abundant taxa in our samples.
The nine OTUs that could be identified accounted for on average 87 ± 14% of the total peak heights detected in any given sample. None of the unidentified OTUs were significantly abundant in any one sample and were rarely detected in more than three samples. However, the sum of relative abundances of all unidentified OTUs was on average the second highest at each sampling station. This suggests that a diverse community of less abundant cyanobacterial taxa exists in these lakes that are not represented by known PC-IGS sequences. The Microcystis (Mcy) OTU designated Mcy215 was on average the most abundant in the entire dataset followed by unidentified taxa> AnaAphChr690> Aphanizomenon (Aph) 680> Mcy661> Mcy506> Aph700> Chroococcus (Chr) 352> Chr132, and Cylindrospermopsis (Cyl) 605. Collectively, the Microcystis taxa made up, on average, about half of the relative abundance at each sampling location (51 +/- 9%).
Spatial and temporal variability in cyanobacterial community composition
A total of 228 samples were collected across the four lakes and twelve stations (17-26 samples per station). Of these, 197 were analyzed by APISA allowing for an analysis of both temporal and spatial variability in CCC. Less samples were analyzed in Lake Wingra due to lack of PCR amplification of the PC-IGS, which is consistent with lower levels of phycocyanin in this lake (Figure S1). Correspondence analysis was undertaken to explore relationships between CCC and environmental variables between sampling stations (Figure 3). The analysis was either unconstrained (Figure 3A and B) or constrained (Figure 3C and D) by environmental variables, the latter assumes all variation in CCC is accounted for by measured environmental variables. Overall, CCC was segregated between deep and shallow (<3.7 m) stations (ANOSIM R=0.23 ,p < 0.001) in both types of ordinations along the first axis and accounted for 36.4% or 30.8% of the variability in the dataset using constrained or unconstrained analyses, respectively (left to right, Figure 2B and 2D). Average ΔT was higher at deep-water sampling locations (Table 1) and variability in CCC along the first axis correlates best with ΔT (Table 2).
Figure 3. Spatial variability in cyanobacterial community composition based on correspondence analysis of all samples (n= 197), constrained (A and B) or not constrained (C and D) by environmental variables.

Samples are coded by sampling site (A and C) or depth of the water column at each sampling site (B and D). Spatial variability in CCC is most strongly segregated between shallow (<3.7 m) and deep- water sites (> 10 m). Stratification (difference in water temperature from surface to lake bottom, ∆T) correlates best with spatial variability in CCC along the first axis.
Table 2. Spatial trends in cyanobacterial community composition with environmental variables based on correspondence analysis.
| Variable |
Unconstrained (R=)
|
Constrained (R=)
|
||
|---|---|---|---|---|
| Axis 1 | Axis 2 | Axis 1 | Axis 2 | |
| TP | 0.15 | 0.27 | 0.24 | 0.14 |
| SRP | 0.15 | 0.34 | 0.28 | 0.29 |
| Nitrate | 0.25 | 0.33 | 0.35 | 0.39 |
| Nitrite | 0.26 | 0.25 | 0.32 | 0.29 |
| ΔT | 0.38 | 0.02 | 0.40 | -0.07 |
| Temp | 0.07 | -0.11 | 0.06 | -0.35 |
| DO | 0.25 | 0.14 | 0.32 | -0.09 |
| PZD | 0.2 | 0.19 | 0.28 | 0.27 |
Of all taxa, the average relative abundance of the Microcystis OTU designated Mcy215 was significantly higher at shallow water stations and was twice as high in the shallowest lake, Lake Wingra than in all other lakes (Figure S3). The same was not true for the other Microcystis taxa, which followed an opposite trend with lowest abundance in Lake Wingra. This accounted for the segregation in CCC between deep and shallow locations.
Shallow locations were also segregated from lower left to the upper left quadrants in the ordination (Figure 3A and 3C). This split was primarily between Lake Wingra samples and all other shallow locations (ANOSIM R= 0.3-0.7, p < 0.002). Variability in CCC along this axis correlated best with nitrate, SRP, and nitrite (Table 2) due to the fact that nitrate and nitrite were below detection limits in Lake Wingra and average SRP was 2-3 times lower in Lake Wingra than in other locations (Table 1). There were some significant but modest differences (ANOSIM R = 0.1-0.4, p < 0.05) in CCC between several locations in the same lake indicating within lake heterogeneity. Many of these significant differences were between deep and shallow locations (Table S1).
We asked whether relative abundance of individual taxa followed the same trend at different sampling locations in the same lake. The unidentified taxa were not included in this analysis since individual unidentified taxa were only detected on a few sampling days at each location. There was a striking difference between Lake Mendota and all other lakes. In Lake Mendota, the trends in relative abundance for any given cyanobacterium, except for one (i.e. Chr132), were similar at all three stations with Pearson R values of 0.6 or greater (p < 0.05). In other words, the same OTU followed the same trend at all three stations. In contrast, with just a few exceptions, individual cyanobacterial taxa in the other three lakes (Monona, Kegonsa, and Wingra) did not follow the same trend between locations in the same lake at a significance cutoff of p < 0.05. For example, in Lake Kegonsa on Sept. 5th, Aph680 was responsible for a bloom at the KEDH location, but was not detected on the same date, 1.9 km away in the same lake at the KEBE location. Only a few taxa followed similar trends in these three lakes. For example, Cylindrospermopsis in Lake Monona (R = 0.6, p = 0.05), Aph700 in Lake Kegonsa (R = 0.9, p < 0.001), and two Aphanizomenon taxa and Chr352 in Lake Wingra. Overall, these data indicate that with the exception of Lake Mendota, most cyanobacterial taxa follow contrasting trends in relative abundance between locations in the same lake.
In 7 out of 9 locations, temporal variability in CCC correlated best with either PZD (R = 0.60-0.66) or SRP (R = 0.60-0.85) along the first axis accounting for 21.2-34.9% of the variability (Table 3). The second axis was most strongly correlated with either ΔT, temp, or SRP accounting for 18.9-23.8% of the variability. Variability in CCC was not strongly correlated with time (i.e. days since sampling began) and a temporal or seasonal trajectory in CCC was not evident at most locations. The data suggest that variability in CCC through time at each location was most strongly influenced by changes in PZD and SRP.
Table 3. Temporal Trends in Cyanobacterial Community Composition with Environmental Variables Based on Canonical Correspondence Analysis.
| Site |
FirstAxis(Variable,Ra) |
%Var |
SecondAxis(Variable,R) |
%Var | ||
|---|---|---|---|---|---|---|
| MEDH | PZD, 0.60 | 31.7 | SRP, 0.54 | 20.2 | ||
| MEPP | SRP, 0.85 | DO, 0.59 | 34.9 | PZD, -0.61 | 18.9 | |
| MEGA | SRP, -0.60 | 30.5 | SRP, 0.58 | DO, 0.55 | 21.0 | |
| MODH | PZD, 0.60 | 31.7 | DO, 0.53 | 20.2 | ||
| MOMB | NO3, -0.54 | 21.2 | Temp, 0.61 | TP, -0.51 | 19.3 | |
| MOBE | Temp, -0.56 | 25.7 | None | 23.8 | ||
| KEDH | PZD, 0.60 | 31.7 | SRP, 0.54 | 20.2 | ||
| KEYA | SRP, -0.60 | 29.5 | SRP, 0.58 | DO, 0.55 | 19.6 | |
| KEBE | PZD, -0.66 | 28.6 | Temp, -0.65 | SRP, -0.53 | 20.9 | |
| WIDHb | DO, -0.73 | SRP, 0.61 | 52.8 | PZD, -0.57 | 16.0 | |
| WIARb | None | 35.0 | None | 23.1 | ||
| WIBEb | Temp, -0.66 | PZD, 0.58 | 36.1 | None | 22.2 | |
Variables that correlate with temporal changes in CCC having R>0.5 (or R< -0.5) are shown. SRP= soluble reactive phosphorus, PZD= photosynthetically active radiation, Temp= average water temperature across the depths sampled, NO3= nitrate.
The analysis of Lake Wingra samples did not include nitrate since nitrate was below detection limits in the majority of samples from all stations in this lake.
Timing of bloom events
Significant increases in phycocyanin were observed in all lakes during the sampling period resulting in blue-green colored scums on the water surface (Figure S2), primarily during late summer and early Autumn. Phycocyanin levels varied by up to two orders of magnitude between weekly samplings (Figure 4). At station MEDH, where adaptive sampling was practiced, phycocyanin concentrations were observed to increase 100-fold, from 62 μg/L to nearly 600 μg/L phycocyanin within a 24-hour period (i.e. Sept 12 – Sept. 13) indicating significant temporal variability in cyanobacterial biomass at sub-weekly time-scales in a given location. The coefficient of variation for phycocyanin was 1.19 with an average and standard deviation of 113 +/- 135 μg/L.
Figure 4. Trends in phycocyanin concentration (line graphs) and water temperature by depth (contour maps) compared to relative abundance of cyanobacterial taxa (bottom heat maps).

Closed circles represent dates when a bloom was occurring. At the MEDH location an in- situ fluorometer attached to a monitoring buoy measured phycocyanin fluorescence once per minute (white line graph).
A bloom event was defined as sustained levels of phycocyanin 1.5X above the simple running average of phycocyanin concentrations not including bloom samples. This definition generally agreed with qualitative observations of water quality during sampling. However, there were several events where this definition grouped multiple, often, lesser phycocyanin increases into one bloom event. These occurred in Lakes Kegonsa and Wingra (Figure 4). Therefore, our bloom definition is a conservative estimate of the number of bloom events that occurred in each location, which is consistent with the significant temporal variability observed from adaptive sampling in Lake Mendota.
The average bloom duration was 29 +/- 25 days. Some blooms lasted for as little as 4 days while others slowly accumulated and then declined over the entire sampling period resulting in a single event lasting > 3 months (Table 4 and Figure 4). Since sampling was on a weekly basis it is possible that some events were missed, or that multiple blooms were grouped into one. Trends in phycocyanin concentration between any two sampling stations, (i.e. within or between lakes) were not significantly correlated with each other at a cutoff of p < 0.05 indicating significant within lake bloom heterogeneity. However, the start of the cyanobacterial bloom season at all stations occurred simultaneously, within one to two weeks of each other in mid-June to early-July. This was coincident with increasing water temperature, declining dissolved nutrient concentrations, and an increase in ΔT (Figure S1).
Table 4. Dominant taxa during mild, moderate, or severe cyanobacterial blooms at each sampling location.
| Site | Bloom# | aDate | Days | bLevel | cDominant Taxa |
|---|---|---|---|---|---|
| MEDH | 1 | Jun 26 | 13 | Mild | Aph680, Aph700, Chr352 |
| 2 | Jul 18 | 14 | Moderate | Aph680, Chr352 | |
| 3 | Sep 15 | 19 | Severe | Mcy215, Mcy506 | |
| 4 | Oct 23 | ND | Mild | Mcy215, Mcy506 | |
| MEPP | 5 | Jun 24 | 8 | Mild | Aph700 |
| 6 | Aug 13 | 35 | Mild | AnaAphChr690, Mcy215 | |
| 7 | Sep 24 | 48 | Severe | Mcy215, Mcy506 | |
| MEGA | 8 | Jun 26 | 20 | Severe | None |
| 9 | Jul 30 | 13 | Mild | ND | |
| 10 | Sep 15 | 26 | Moderate | Mcy215 | |
| 11 | Oct 17 | 6 | Moderate | Mcy215, Mcy661 | |
| MODH | 12 | Jun 24 | 8 | Mild | AnaAphChr690, Aph680, Aph700 |
| 13 | Jul 9 | 7 | Mild | ND | |
| 14 | Jul 23 | 21 | Mild | Mcy215 | |
| 15 | Aug 27 | 50 | Moderate | None | |
| MOBE | 16 | Jun 11 | 13 | Mild | Aph700, Mcy215 |
| 17 | Jul 16 | 34 | Moderate | Mcy215 | |
| 18 | Aug 20 | 23 | Moderate | Mcy215 | |
| 19 | Sep 11 | 4 | Mild | ND | |
| 20 | Sep 24 | 15 | Moderate | Mcy215 | |
| MOMB | 21 | Jul 30 | 41 | Mild | None |
| 22 | Sep 24 | 41 | High | Mcy215 | |
| KEDH | 23 | Sep 3 | 98 | Moderate | Aph680 |
| 24 | Oct 16 | ND | Moderate | Mcy215 | |
| KEBE | 25 | Jun 24 | 9 | Mild | None |
| 26 | Sep 17 | 98 | Severe | Mcy215, Mcy661 | |
| KEYA | 27 | Jul 24 | 42 | Mild | ND |
| 28 | Sep 3 | 41 | Moderate | Aph700, Aph680 | |
| WIDH | 29 | Jun 26 | 21 | Moderate | Mcy215 |
| 30 | Aug 7 | 41 | Moderate | AnaAphChr690, Mcy215 | |
| 31 | Sep 11 | 28 | Moderate | Mcy215 | |
| 32 | Oct 16 | >6 | Moderate | ND | |
| WIBE | 33 | Jun 26 | 7 | Mild | Mcy215 |
| 34 | Aug 14 | 91 | Severe | Mcy215 | |
| WIAR | 35 | Jun 26 | 28 | Moderate | ND |
| 36 | Aug 7 | 7 | Mild | AnaAphChr690, Mcy215 | |
| 37 | Sep 11 | 54 | Severe | Mcy215 |
The peak of the bloom is defined as the date when maximum phycocyanin concentrations occurred.
The severity of bloom events was defined by the concentration of phycocyanin (µg/L) at the peak of the bloom event. Mild = <150, Moderate = 150-500, Severe = >500.
Dominant taxa detected at the peak of the bloom event. ND = Not determined, None = no dominant taxa representing >20% relative abundance.
Dominant taxa during blooms
Microcystis and Aphanizomenon dominated (i.e. ≥ 20% relative abundance) most bloom events (Figure 4, Table 4). While there was significant variability in phycocyanin over time the relative abundance of individual taxa did not always vary with blooms. The heat maps in Figure 4 show that Mcy215 was consistently detected at most locations and on most sampling days regardless of whether a bloom was occurring. The relative abundance of the other two Microcystis taxa grew significantly on only a few days in primarily the deepest locations (e.g. MEDH and MODH), but their peak relative abundances did not coincide with most bloom events (Table 4). Of the 31 bloom events analyzed by APISA, twelve were dominated by Mcy215 alone, while 9 of the 31 events were co-dominated by Mcy215 and other taxa. While no single cyanobacterium was responsible for all bloom events, Mcy215 was involved in the majority of them (i.e. 68%) and all of these were severe (i.e. phycocyanin > 500 µg/L). Of the 21 Microcystis - dominated bloom events all but seven occurred in August and September, corresponding to periods when SRP and nitrate were at their lowest and thermal stratification was strong.
The Aphanizomenon taxa dominated two bloom events alone (i.e. Aph680 or Aph700) and five together and/or with other taxa. Nearly all Aphanizomenon -dominated bloom events (5 out of 7) occurred early in the sampling period (i.e. June and July) and/or occurred prior to the first Microcystis bloom (Table 4). These five events occurred at five different stations, all early in summer (1st or 2nd week of June). Of all stations, only two had a Microcystis dominated bloom early in summer, but these occurred at the end of June and both stations were in Wingra where Aphanizomenon was generally absent. Seven stations did not have an Aphnizomenon- dominated bloom early in summer. In a few blooms, no dominant taxa were present at the peak of the bloom (i.e. none with > 20% relative abundance) indicating multiple taxa contributed to the bloom or the dominant organism could not be detected by our methods.
Relative Abundance of Aphanizomenon and Microcystis Are Anti-Correlated
While Microcystis and Aphanizomenon were the dominant taxa during most bloom events, they rarely bloomed together and only co-dominated the same bloom event on one occasion (Table 4, bloom# 16). These two genera were nearly mutually exclusive across all lakes. The sum of relative abundances of Aphanizomenon and Microcystis taxa were significantly (p < 0.001) anti-correlated with each other at nine of twelve stations with Pearson R values of -0.7--0.9 (Figure 5). The community cycled between Aphanizomenon and Microcystis dominance on multiple occasions at each of these nine stations and Microcystis was always the dominant taxa at the end of the sampling period in late October (Figure S1). No other genera (e.g. Chroococcus ) or taxa (e.g. AnaAphChr690) were significantly anti-correlated with either Aphanizomenon or Microcystis .
Figure 5. Significant negative linear relationships between Microcystis and Aphanizomenon relative abundance at nine of twelve sampling sites.

The linear fit of the relationship at each of the nine sites is given by dotted lines.
Linear trends in the dominant taxa during blooms including two Aphanizomenon taxa, Mcy215 and the sum of Microcystis or Aphanizomenon taxa were significantly correlated with nutrients and/or physical variables at 9 of 12 stations (Table S2). The Microcystis and Aphanizomeon taxa showed opposing correlations to the same variable at all three stations in Mendota, two in Monona and one in Kegonsa, suggesting that these genera show contrasting responses to environmental conditions. For example, at these stations Aphanizomenon showed strong positive correlations with SRP and/or ΔT while Microcystis showed negative correlations to these same variables.
Average dissolved nutrient concentrations, including nitrate, nitrite, and SRP were significantly different (p < 3.6 x 10-5) between periods of Microcystis and Aphanizomenon dominance (Figure 6). Among other variables, PAR > DO > ΔT were also important with decreasing significance (p > 0.01). The nitrate plus nitrite (N+N) to SRP ratio (N+N:SRP) also differed between Microcystis and Aphanizomenon abundance (i.e. lower during Aphanizomenon abundance), but was not significant at all stations. Together, these data suggest that Aphanizomenon was most often dominant during periods when nutrient concentrations were relatively high compared to periods of Microcystis dominance, but the ratio of available inorganic N relative to P was low, possibly favoring this nitrogen fixing cyanobacterium. At two of the three stations where Microcystis and Aphanizomenon were not anti-correlated (i.e. WIAR, and WIBE), Aphanizomenon relative abundance was negligible while in the third station (i.e. MOMB), Aphanizomenon and Microcystis were significantly anti-correlated with the exception of June samples.
Figure 6. Average concentrations of dissolved nutrients during periods of Aphanizomenon and Microcystis abundance.

All except N+N:SRP are significant differences (P<0.01).
Discussion
In this study, we attempted to capture both spatial and temporal variability of CCC in multiple eutrophic lakes and stations using a culture-independent molecular method. We discovered that most bloom events across all lakes and stations were dominated by just three of the more than 90 taxa or genotypes identified (e.g. Aph680, Aph700, and Mcy215). Although other taxa were involved in some bloom events, at least one of these three taxa were involved in all of them. Furthermore, we found that not all Microcystis taxa were equal with respect to bloom formation, and that only one of the three Microcystis taxon identified (Mcy215) was responsible for most cyanobacterial bloom events across all stations.
With respect to accumulations in cyanobacterial biomass, our observations were consistent with previous work [40]. Blooms formed when water temperatures were above 20°C, stratification (∆T) was increasing, and dissolved nutrients were either declining or at a minimum. However, the molecular analyses showed that most of the first blooms involved the two Aphanizomenon taxa and occurred within 1-2 weeks of one another at all stations, while most of the subsequent blooms were dominated specifically by Microcystis , Mcy215. Additionally, the Microcystis blooms were not concurrent across stations. This suggests that Aphanizomenon responds to the change in season while Microcystis benefits from a suite of intersecting station- specific conditions. We found that seasonal trends in water column stratification predict Microcystis abundance well in two similar deep water stations in Lake Mendota, but not in any of the other lakes or stations sampled. In neighboring Lake Monona Microcystis was correlated with SRP, but not stratification. However, this was likely due to the fact that 2 of 3 Lake Monona stations were shallow. In other studies, temporal variability in Microcystis has been correlated with both stratification and SRP, but usually only in a single lake or multiple lakes with poor temporal resolution. Hence, heterogeneity in cyanobacterial ecology, genetics, and physiology within and between lakes is likely even greater than currently recognized. We should note that only a subset of all possible environmental variables were measured in this study.
We showed that there is significant within lake spatial heterogeneity, particularly with respect to cyanobacterial genetic structure and overall abundance. For example, trends in phycocyanin concentration were not correlative between any two locations and except for Lake Mendota, the relative abundance of individual taxa at locations within the same lake followed contrasting trajectories through time (Figure 4). These data suggests that conclusions based on single sampling points in a lake are unlikely to be directly transferrable to other locations within a lake. Furthermore the data support further research focused on improving three- dimensional coupled hydrodynamic-ecosystem modeling approaches (e.g. 41) in order to capture this heterogeneity.
We observed that CCC was significantly segregated between stations according to depth of the water column. Accordingly, of all environmental variables measured, ∆T was most strongly correlated with spatial variability in CCC (Figure 2, Table 2). Water column stability is thought to favor growth of some cyanobacteria and previous studies have noted differences in CCC between deep and shallow lakes [42]. For example, the Oscillatoriales are often observed to dominate shallow systems [43,44]. However, this is clearly not always the case since Chroococcales such as Microcystis have been observed to dominate many shallow lakes including Lake Taihu, a well-studied shallow eutrophic lake in Eastern China [45]. In this study, Microcystis genera were more abundant in shallow locations, especially Lake Wingra, but also emerged later in the Lake Mendota deep locations when stratification was strong. In a study by Brock et al. (1985) in Lake Mendota, Microcystis were more often observed in deep sediment traps than Aphanizomenon . Thus, strong vertical stability may be important in order for Microcystis maintain significant biomass in the photic zone of deep locations, whereas in shallow locations, the water column in thoroughly mixed with light penetrating throughout. This may be a result of poorer buoyancy control compared to Aphanizomenon , or perhaps Microcystis are better adapted to high light conditions in these lakes.
The affect of depth of the water column on cyanobacterial ecology is well studied and may be explained by light and nutrient availability regulated by water column stratification [46,47]. In this study, PZD extended through nearly the entire water column on most sampling days at shallow locations. The PZD was important for temporal variability in CCC (Table 3), but not spatial variability between stations (Table 2). Aside from ΔT, dissolved inorganic nutrients were also significantly correlated with spatial variability in overall CCC. All shallow stations had slightly lower mean concentrations of standing stock SRP, which may have been due to co-occurrence of macrophytes in these areas. Thus, it appears that nutrient availability and eukaryotic competition may play a larger role in structuring CCC between shallow and deep locations within these lakes than does PZD. This agrees with the average higher coefficient of variation in nutrient levels (0.9-1.6) compared to the PZD (0.4-0.6) at all locations.
Temporal variations in CCC were most strongly correlated with PZD and/or SRP, and to a lesser extent with nitrate, water temperature, and DO. In 2008 when this study was conducted, spring rainfall was abnormally high resulting in significant flooding across the region in June. Accordingly, dissolved inorganic nutrients were highest in June. This came during a period of increasing stratification and well after the spring mixing event that begins during ice-off (March or April). The strong pulse of nutrients delivered during these rain events could have disrupted any seasonal trend in CCC, which was not observed in our dataset. Rather the influx of inorganic nutrients, particularly SRP, may have structured CCC during this year. Cyanobacterial blooms of Aphanizomenon and, to a lesser extent, Microcystis quickly increased after the June rain events as nutrients and PZD declined while water temperature increased (Table 4, Figure S1). Thus, it is likely that June rainfall events and associated nutrient influx played a large role in shaping the temporal variation observed in bloom formation and CCC during 2008. As climate change is expected to induce extreme weather events, the 2008 flooding may be representative of future years.
We observed a strong anti-correlation between Aphanizomenon and Microcystis . This anti-correlation is primarily due to the occurrence of Aphanizomenon blooms early in summer while Microcystis had peak abundances later in summer into early autumn at most locations (Figure S1). In addition, at some stations (e.g. MEDH, KEBE) the population cycled between Microcystis and Aphanizomenon dominance two or three times throughout the sampling period. Our data do not allow for a definitive explanation of this phenomenon. However, there are several possibilities.
The N2- fixing cyanobacteria such as Aphanizomenon are expected to out compete other non-N2- fixing cyanobacteria when N:P ratios decline below some threshold, which could be higher than the Redfield ratio [48]. At most stations, nutrients were just beginning to decline from peak concentrations in early June when Aphanizomenon blooms were observed and the N+N:SRP ratio was low or declining. It is possible this decline was significant enough to trigger N2- fixation giving some advantage to early blooms of Aphanizomenon and not Microcystis . A recent study by Beversdorf et al. in Lake Mendota suggests that N2- fixation by Aphanizomenon during nutrient draw down early in the summer provides new N input supporting toxic Microcystis blooms later in summer [20]. As this new N is consumed, N2- fixing Aphanizomenon are favored thus explaining cycling between Aphanizomenon and Microcystis .
On the other hand, early blooms of Aphanizomenon may be explained by a slightly lower optimal growth temperature for Aphanizomenon compared to that of Microcystis . At the start of our study in June, water temperatures were at 15 °C, increased to 27 °C in early July and then reached a maximum of 30 °C at all stations between mid- July and early August. Most, but not all Aphanizomenon blooms occurred before or at the time max temperatures were reached, while many of the Microcystis blooms occurred after max temperatures were reached. A literature review by Robarts and Zohary indicate that growth of Microcystis stops below 15 °C with optimal growth temperatures of 27.5-32 °C. In contrast, the optimal growth temperature for four strains of Aphanizomenon was 15-28 °C [49]. This suggests that Aphanizomenon may have the ability to outgrow Microcystis at lower temperatures, such as earlier in the summer of temperature lakes. This agrees with our observations of earlier blooms of Aphanizomenon . However, we did not examine optimal growth temperatures of the Aphanizomenon taxa in our lakes and other drivers may be more important.
Another possibility is that anti-predation characteristics specifically possessed by Aphanizomenon in these lakes may be important for establishing conditions that allow cyanobacteria to dominate, transitioning out of the “clear water” phase when zooplankton are abundant. Both Microcystis and Aphanizomenon display anti-predation phenotypes including colony formation, toxin production, and overall prey avoidance [50,51]. It is possible that a combination of lower optimal growth temperature, anti- predation characteristics, and N2- fixation provide a competitive advantage for Aphanizomenon earlier in the summer during the transition out of the zooplankton dominated clear- water phase. This hypothesis requires more investigation, but should be fruitful since much of the cyanobacterial bloom literature has focused on Microcystis blooms.
As we look to forecast the effects of management decisions, climate and land use change on the occurrence and frequency of algal bloom events, it is necessary to characterize taxa responsible for producing this biomass. We can draw some very salient parallels to the infectious disease literature, which is right now focused on the necessity of differentiating bacterial pathogens based on genetic markers rather than phenotypic morphological markers [51]. Should we treat cyanobacteria any differently? This study shows that not all taxa of the bloom- forming genera in lakes (e.g. Microcystis or Aphanizomenon ) are responsible for producing the bulk of biomass in lakes. In fact, only one of three Microcystis OTUs was responsible for most bloom events indicating that these “species” differ greatly in their ability to dominate the water column. Therefore, the identification of taxa that specifically contribute to blooms at a genetic level is likely to provide a better understanding of CCC and overall cyanobacterial bloom ecology.
Conclusions
This study provided the most comprehensive analysis to date of spatial and temporal variability in CCC across multiple lakes and within-lake stations using a culture- independent molecular approach. The CCC across all stations was most strongly segregated by depth of the sampling location and was correlated most strongly with thermal stratification and nutrients. Spatial variability in CCC and temporal trends in taxa abundance were rarely correlative between sampling stations in the same lake indicating significant heterogeneity in the x-y dimensions. More than 90 taxa were identified, yet a single Microcystis , and two different Aphanizomenon taxa were the dominant cyanobacteria detected during all bloom events at all stations including multiple stations in the same lake. The Microcystis and Aphanizomenon taxa were significantly anti-correlated with each other at most stations and Aphanizomenon was more abundant when nutrients were higher suggesting interesting interactions between these genera potentially driven by N2-fixation or other phenotypes (e.g. optimal growth temperature, anti- predation characteristics). Overall, this study shows that temporal trends in biomass and individual taxa are heterogeneous between lakes and locations in the same lake. Furthermore, the molecular approach shows that just a few individual genotypes were responsible for most bloom events.
Supporting Information
Results of pair-wise comparisons of cyanobacterial community composition between locations in the same lake using ANOSIM.
(DOCX)
Significant (P<0.05) correlations (Pearson R) between the major bloom forming Aphanizomenon , or Microcystis taxa and environmental variables.
(DOCX)
Trends in biological, physical, and chemical variables across all lakes and locations. For each location the top graph represents the sum of the relative abundance of all Microcystis taxa compared to that of Aphanizomenon taxa. Arrows indicate peak bloom dates based on phycocyanin. The second graph shows changes in phycocyanin and water temperature in the photic zone. The photic zone depth (PZD) was estimated from secchi disk depth, which is shown in the third graph along with the difference in water temperature from surface to lake bottom as a measure of water column stratification (ΔT). The fourth graph shows trends in nutrients (TP = total phosphorus, SRP = soluble reactive phosphorus). and the nitrate+nitrite to SRP ratio (N+N:SRP). Nitrate and nitrite were below detection limits in Lake Wingra.
(TIF)
Cyanobacterial surface scums on Lake Kegonsa, WI, August 2008.
(TIF)
Differences in mean relative abundance of identified taxa between deep and shallow locations. Chr352, Mcy215, and Unmatched taxa are significant differences.
(TIF)
Acknowledgments
We would like to thank Holly Powell, and Adrienne Rust for their invaluable help obtaining water samples; Katherine Waugh, Jay Hawley, Ian Conlin, and Beomkang Huh for their help with laboratory analyses; and Jordan Read, Paul Hanson, Luke Winslow, Ashley Shade, and Stuart Jones for their help with statistical analyses. In addition, we wish to thank Dave Harring for maintenance of boats, vehicles, trailers, and other sampling equipment necessary for this project.
Funding Statement
Funding provided by a Wisconsin SeaGrant (R/BT-24; http://www.seagrant.wisc.edu/home/), and the National Science Foundation (CAREER CBET-0644949; NTL-Microbial Observatory MCB-0702395; NTL-LTER DEB-0822700; http://www.nsf.gov/). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kanoshina I, Lips U, Leppanen J-M (2003) The influence of weather conditions (temperature and wind) on cyanobacterial bloom development in the Gulf of Finland (Baltic Sea). Harmful Algae 2: 29-41. doi:10.1016/S1568-9883(02)00085-9. [Google Scholar]
- 2. Paerl HW (1996) A comparison of cyanobacterial bloom dynamics in freshwater, estuarine and marine environments. Phycologia 35: 25-35. doi:10.2216/i0031-8884-35-6S-25.1. [Google Scholar]
- 3. Dodds WK, Bouska WW, Eitzmann JL, Pilger TJ, Pitts KL et al. (2009) Eutrophication of US freshwaters: analysis of potential economic damages. Environ Sci Technol 43: 12-19. doi:10.1021/es801217q. PubMed: 19209578. [DOI] [PubMed] [Google Scholar]
- 4. Paerl HW, Huisman J (2008) Climate. Blooms like it hot. Science 320: 57-58. doi:10.1126/science.1155398. PubMed: 18388279. [DOI] [PubMed] [Google Scholar]
- 5. O’Neil JM, Davis TW, Burford MA, Gobler CJ (2012) The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 14: 313-334. doi:10.1016/j.hal.2011.10.027. [Google Scholar]
- 6. Miller TR, McMahon KD (2011) Genetic diversity of cyanobacteria in four eutrophic lakes. FEMS Microbiol Ecol 78: 336-348. doi:10.1111/j.1574-6941.2011.01162.x. PubMed: 21707672. [DOI] [PubMed] [Google Scholar]
- 7. Willen T, Mattsson R (1997) Water-blooming and toxin-producing cyanobacteria in Swedish fresh and brackish waters, 1981-1995. Hydrobiologia 353: 181-192. doi:10.1023/A:1003047019422. [Google Scholar]
- 8. Padisák J (1997) Cylindrospermopsis raciborskii (Wołoszy´nska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: worldwide distribution and review of its ecology. Arch Hydrobiol Supplement 107: 563–593. [Google Scholar]
- 9. Walsby AE (1972) Gas-filled structures providing buoyancy in photosynthetic organisms. Symp Soc Exp Biol 26: 233-250. PubMed: 4196325. [PubMed] [Google Scholar]
- 10. Yang Z, Kong FX, Cao HS, Shi XL (2005) Observation on colony formation of Microcystis aeruginosa induced by filtered lake water under laboratory conditions. Ann Limnol - International Journal of Limnology 41: 169-173 [Google Scholar]
- 11. Price GD, Badger MR, Woodger FJ, Long BM (2008) Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J Exp Bot 59: 1441-1461. PubMed: 17578868. [DOI] [PubMed] [Google Scholar]
- 12. Jacobson L, Halmann M (1982) Polyphosphate metabolism in the blue-green alga Microcystis aeruginosa. J Plankton Res 4: 481-488. doi:10.1093/plankt/4.3.481. [Google Scholar]
- 13. Simon RD (1971) Cyanophycin granules from the blue-green alga Anabaena cylindrica: a reserve material consisting of copolymers of aspartic acid and argenine. Proc Natl Acad Sci USA 68: 265-267. doi:10.1073/pnas.68.2.265. PubMed: 16591901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Steinberg CEW, Hartmann HM (1988) Planktonic bloom-forming Cyanobacteria and the eutrophication of lakes and rivers. Freshw Biol 20: 279-287. doi:10.1111/j.1365-2427.1988.tb00452.x. [Google Scholar]
- 15. Deblois CP, Juneau P (2012) Comparison of resistance to light stress in toxic and non-toxic strains of Microcystis aeruginosa (Cyanophyta). J Phycol 48: 1002-1011. doi:10.1111/j.1529-8817.2012.01191.x. [DOI] [PubMed] [Google Scholar]
- 16. Van Gremberghe I, Vanormelingen P, Vanelslander B, Van der Gucht K, D’Hondt S et al. (2009) Genotype-dependent interactions among sympatric Microcystis strains mediated by Daphnia grazing. Oikos 118: 1647-1658. doi:10.1111/j.1600-0706.2009.17538.x. [Google Scholar]
- 17. Briand E, Escoffier N, Straub C, Sabart M, Quiblier C et al. (2009) Spatiotemporal changes in the genetic diversity of a bloom-forming Microcystis aeruginosa (cyanobacteria) population. ISME J 3: 419-429. doi:10.1038/ismej.2008.121. PubMed: 19092863. [DOI] [PubMed] [Google Scholar]
- 18. Briand E, Yéprémian C, Humbert JF, Quiblier C (2008) Competition between microcystin- and non-microcystin-producing Planktothrix agardhii (cyanobacteria) strains under different environmental conditions. Environ Microbiol 10: 3337-3348. doi:10.1111/j.1462-2920.2008.01730.x. PubMed: 18759740. [DOI] [PubMed] [Google Scholar]
- 19. Wang X, Sun M, Wang J, Yang L, Luo L et al. (2012) Microcystis genotype succession and related environmental factors in Lake Taihu during cyanobacterial blooms. Microb Ecol, 64: 986–99. PubMed: 22760733. [DOI] [PubMed] [Google Scholar]
- 20. Beversdorf LJ, Miller TR, McMahon KD (2013) The role of nitrogen fixation in cyanobacterial bloom toxicity in a temperate, eutrophic lake. PLOS ONE 8: e56103. doi:10.1371/journal.pone.0056103. PubMed: 23405255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wilson AE, Sarnelle O, Neilan BA, Salmon TP, Gehringer MM et al. (2005) Genetic variation of the bloom-forming cyanobacterium Microcystis aeruginosa within and among lakes: Implications for harmful algal blooms. Appl Environ Microbiol 71: 6126-6133. doi:10.1128/AEM.71.10.6126-6133.2005. PubMed: 16204530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Vézie C, Rapala J, Vaitomaa J, Seitsonen J, Sivonen K (2002) Effect of nitrogen and phosphorus on growth of toxic and nontoxic Microcystis strains and on intracellular microcystin concentrations. Microb Ecol 43: 443-454. doi:10.1007/s00248-001-0041-9. PubMed: 11953809. [DOI] [PubMed] [Google Scholar]
- 23. Kim SG, Rhee SK, Ahn CY, Ko SR, Choi GG et al. (2006) Determination of cyanobacterial diversity during algal blooms in Daechung Reservoir, Korea, on the basis of cpcBA intergenic spacer region analysis. Appl Environ Microbiol 72: 3252-3258. doi:10.1128/AEM.72.5.3252-3258.2006. PubMed: 16672464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Fewer DP, Köykkä M, Halinen K, Jokela J, Lyra C, et al. (2009) Culture-independent evidence for the persistent presence and genetic diversity of microcystin-producing Anabaena (Cyanobacteria) in the Gulf of Finland. Environmental Microbiology 11: 855-866 [DOI] [PubMed] [Google Scholar]
- 25. Budikova D, Coleman JSM, Strope SA, Austin A (2010) Hydroclimatology of the 2008 Midwest floods. Water Resour Res 46: W12524. [Google Scholar]
- 26. Watson VJ, Loucks OL, Mitchell J, Clesceri NL (1979) Impact of development on watershed hydrologic and nutrient budgets. J Water Pollut Contr Fed 51: 2876-2885. [Google Scholar]
- 27. Birge EA, Juday C (1926) The organic content of lake water. Proc Natl Acad Sci U S A 12: 515-519. doi:10.1073/pnas.12.8.515. PubMed: 16587120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Poole HH, Atkins WRG (1929) Photoelectric measures of submarine illumination throughout the year. J Mar Biol Assoc UK 16: 297-324. doi:10.1017/S0025315400029829. [Google Scholar]
- 29. Yannarell AC, Kent AD, Lauster GH, Kratz TK, Triplett EW (2003) Temporal patterns in bacterial communities in three temperate lakes of different trophic status. Microb Ecol 46: 391-405. doi:10.1007/s00248-003-1008-9. PubMed: 12904915. [DOI] [PubMed] [Google Scholar]
- 30. Association APH, American Water Works Association, publication WEF (1995). Stand Methods Examination Water Wastewater Washington D.C.. [Google Scholar]
- 31. Flowers JJ, He S, Ivanova N, Yilmaz S, Noguera DR et al. (2009) Denitrification capabilities of two enhanced biological phosphorus removal sludges dominated by different 'Candidatus Accumulibacter' clades. Environ Microbiol Rep 1: 583-588. doi:10.1111/j.1758-2229.2009.00090.x. PubMed: 20808723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Marsac NTD, Houmard J (1988) Complementary chromatic adaptation: physiological conditions and action spectra. Methods Enzymol 167. [Google Scholar]
- 33. Neilan BA, Jacobs D, Goodman AE (1995) Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl Environ Microbiol 61: 3875-3883. PubMed: 8526499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Fisher MM, Triplett EW (1999) Automated approach for ribosomal intergenic spacer analysis of microbial diversity and its application to freshwater bacterial communities. Appl Environ Microbiol 65: 4630-4636. PubMed: 10508099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Shade A, Kent AD, Jones SE, Newton RJ, Triplett EW et al. (2007) Interannual dynamics and phenology of bacterial communities in a eutrophic lake. Limnol Oceanogr 52: 487-494. doi:10.4319/lo.2007.52.2.0487. [Google Scholar]
- 36. Ludwig W, Strunk O, Westram R, Richter L, Meier H et al. (2004) ARB: a software environment for sequence data. Nucleic Acids Res 32: 1363-1371. doi:10.1093/nar/gkh293. PubMed: 14985472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Ricke P, Kolb S, Braker G (2005) Application of a newly developed ARB software-integrated tool for in silico terminal restriction fragment length polymorphism analysis reveals the dominance of a novel pmoA cluster in a forest soil. Appl Environ Microbiol 71: 1671-1673. doi:10.1128/AEM.71.3.1671-1673.2005. PubMed: 15746378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Kent AD, Smith DJ, Benson BJ, Triplett EW (2003) Web-based phylogenetic assignment tool for analysis of terminal restriction fragment length polymorphism profiles of microbial communities. Appl Environ Microbiol 69: 6768-6776. doi:10.1128/AEM.69.11.6768-6776.2003. PubMed: 14602639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Braak CT, Smilauer P (2002) CANOCO reference manual and CanoDraw for Windows user’s guide: software for canonical community ordination (version 45). Ithaca, NY: Microcomputer Power. [Google Scholar]
- 40. Paerl H (2008) Chapter 10: Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater-marine continuum10. In: Hudnell HK. International Symposium on Cyanobacterial Harmful Algal Blooms: Berlin: Springer-Verlag; . pp. 217-237 [DOI] [PubMed] [Google Scholar]
- 41. Carraro E, Guyennon N, Hamilton D, Valsecchi L, Manfredi EC et al. (2012) Coupling high-resolution measurements to a three- dimensional lake model to assess the spatial and temporal dynamics of the cyanobacterium Planktothrix rubescens in a medium-sized lake. Hydrobiologia 698: 77-95. doi:10.1007/s10750-012-1096-y. [Google Scholar]
- 42. Nixdorf B, Mischke U, Rücker J (2003) Phytoplankton assemblages and steady state in deep and shallow eutrophic lakes – an approach to differentiate the habitat properties of Oscillatoriales. Hydrobiologia 502: 111-121. doi:10.1023/B:HYDR.0000004274.65831.e5. [Google Scholar]
- 43. Rucker J, Widener C, Zippel P (1997) Factors controlling the dominance of Planktothrix agardhii and Limnothrix redekei in eutrophic shallow lakes. Hydrobiologia 342: 107-115. [Google Scholar]
- 44. Scheffer M, Rinaldi S, Gragnani A, Mur LR, vanNes EH (1997) On the dominance of filamentous cyanobacteria in shallow, turbid lakes. Ecology 78: 272-282. doi:10.1890/0012-9658(1997)078[0272:OTDOFC]2.0.CO;2. [Google Scholar]
- 45. Chen Y, Qin B, Teubner K, Dokulil MT (2003) Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J Plankton Res 25: 445-453. doi:10.1093/plankt/25.4.445. [Google Scholar]
- 46. Dokulil MT, Teubner K (2000) Cyanobacterial dominance in lakes. Hydrobiologia 438: 1-12. doi:10.1023/A:1004155810302. [Google Scholar]
- 47. Reynolds CS, Oliver RL, Walsby AE (1987) Cyanobacterial dominance: The role of buoyancy regulation in dynamic lake environments. N Z J Mar Freshw Res 21: 379-390. doi:10.1080/00288330.1987.9516234. [Google Scholar]
- 48. Tõnno I, Nõges T (2003) Nitrogen fixation in a large shallow lake: rates and initiation conditions. Hydrobiologia 490: 23-30. doi:10.1023/A:1023452828667. [Google Scholar]
- 49. Robarts RD, Zohary T (1987) Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. N Z J Mar Freshw Res 21: 391-399. doi:10.1080/00288330.1987.9516235. [Google Scholar]
- 50. Chakraborty S, Bhattacharya S, Feudel U, Chattopadhyay J (2012) The role of avoidance by zooplankton for survival and dominance of toxic phytoplankton. Ecol Complexity 11: 144-153. doi:10.1016/j.ecocom.2012.05.006. [Google Scholar]
- 51. Lampert W (1987) Laboratory studies on zooplankton-cyanobacteria interactions. N Z J Mar Freshw Res 21: 483-490. doi:10.1080/00288330.1987.9516244. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Results of pair-wise comparisons of cyanobacterial community composition between locations in the same lake using ANOSIM.
(DOCX)
Significant (P<0.05) correlations (Pearson R) between the major bloom forming Aphanizomenon , or Microcystis taxa and environmental variables.
(DOCX)
Trends in biological, physical, and chemical variables across all lakes and locations. For each location the top graph represents the sum of the relative abundance of all Microcystis taxa compared to that of Aphanizomenon taxa. Arrows indicate peak bloom dates based on phycocyanin. The second graph shows changes in phycocyanin and water temperature in the photic zone. The photic zone depth (PZD) was estimated from secchi disk depth, which is shown in the third graph along with the difference in water temperature from surface to lake bottom as a measure of water column stratification (ΔT). The fourth graph shows trends in nutrients (TP = total phosphorus, SRP = soluble reactive phosphorus). and the nitrate+nitrite to SRP ratio (N+N:SRP). Nitrate and nitrite were below detection limits in Lake Wingra.
(TIF)
Cyanobacterial surface scums on Lake Kegonsa, WI, August 2008.
(TIF)
Differences in mean relative abundance of identified taxa between deep and shallow locations. Chr352, Mcy215, and Unmatched taxa are significant differences.
(TIF)