

Abstract
In in vitro fertilization cycles, both HP-hMG and rFSH gonadotropin treatments are widely used to control human follicle development. The objectives of this study are (i) to characterize and compare gene expression profiles in cumulus cells (CCs) of periovulatory follicles obtained from patients stimulated with HP-hMG or rFSH in a GnRH antagonist cycle and (ii) to examine their relationship with in vitro embryo development, using Human Genome U133 Plus 2.0 microarrays. Genes that were upregulated in HP-hMG-treated CCs are involved in lipid metabolism (GM2A) and cell-to-cell interactions (GJA5). Conversely, genes upregulated in rFSH-treated CCs are implicated in cell assembly and organization (COL1A1 and COL3A1). Interestingly, some genes specific to each gonadotropin treatment (NPY1R and GM2A for HP-hMG; GREM1 and OSBPL6 for rFSH) were associated with day 3 embryo quality and blastocyst grade at day 5, while others (STC2 and PTX3) were related to in vitro embryo quality in both gonadotropin treatments. These genes may prove valuable as biomarkers of in vitro embryo quality.
1. Introduction
The gonadotropin-releasing hormone (GnRH) antagonist and agonist protocols with either highly purified human menopausal gonadotropin (HP-hMG) or recombinant FSH (rFSH) preparations are the most widely used protocols for controlled ovarian stimulation (COS) for both intracytoplasmic sperm injection (ICSI) and in vitro fertilization (IVF) [1–3]. At present, most of the mature oocytes retrieved after COS are capable of fertilization; however, only half of them develop into good embryos and only a few implants. There is increasing evidence that cumulus cells (CCs), which are somatic cells that surround the oocyte, play a crucial role in folliculogenesis and oocyte developmental competence acquisition [4, 5]. Several authors propose the use of CC gene expression as a noninvasive approach to predict oocyte aneuploidy, and oocyte competence, as well as embryo and pregnancy outcomes during assisted reproductive technology (ART) procedures [6–17]. Despite the recent molecular advances in the knowledge of human CCs, our understanding is far from complete. We believe that the characterization of the biology of these cells following COS might explain observed changes in in vitro embryo development. Several studies have compared the effects of HP-hMG and rFSH on oocyte and embryo quality, follicular fluid biochemical profile, and pregnancy rate [18–23]. However, their specific effects on the gene expression profile of individual CC samples have not been investigated. To date, only two such studies have been reported. They compared the gene expression profiles of pooled human granulosa cells (GCs) from periovulatory follicles of six patients in one study and eight patients in the other study. In both studies, the patients were treated with HP-hMG or rFSH in a GnRH agonist long protocol. Significant differences have been observed [24, 25]. The aims of the present study were (i) to compare the gene expression profiles of large cohorts of individual CCs isolated from periovulatory follicles of patients stimulated with HP-hMG or rFSH in a GnRH antagonist protocol and (ii) to determine the relationship between in vitro embryo development and expression profiles of CCs isolated from mature oocytes after COS.
2. Materials and Methods
2.1. Study Oversight
This research was approved by our Institutional Review Board. All patients provided their written informed consent for the use of CC samples for research.
2.2. Sample Collection and Treatment Cycle
This study is a retrospective analysis of data from of a subgroup of eleven randomly selected patients, who participated in an open-label, assessor-blind, parallel groups, multicenter trial (ClinicalTrials.gov Identifier: NCT00884221) that was previously described [26]. CCs (n = 146) were collected from all oocytes retrieved from four patients treated with HP-hMG (Menopur, Ferring Pharmaceuticals) and seven patients treated with rFSH (Follitropin beta, Puregon; MSD) following a GnRH antagonist protocol (Ganirelix Acetate, Orgalutran; MSD), respectively. Stimulation with HP-hMG or rFSH was started at a dose of 150 IU/day (first 5 days of the COS protocol), and the patients' follicular response during stimulation was monitored by transvaginal ultrasound. The GnRH antagonist (daily dose of 0.25 mg) was initiated at day 6 and continued throughout the stimulation period. Transvaginal ultrasound echo guidance, FSH, LH, and estradiol levels were used to monitor the ovarian response. A single injection of 250 μg human chorionic gonadotropin (hCG) (choriogonadotropin alfa, Ovitrelle; Merck Serono) was administered to induce the final follicular maturation when three or more follicles ≥17 mm in diameter were observed. Cumulus-oocyte-complexes were collected 36 h after hCG administration (day 0). Supplemental Table SI (see Supplementary Materials available online at http://dx.doi.org/10.1155/2013/354582) shows a summary of the patients' clinical features, end-of-stimulation data, and the number of retrieved oocytes/patients. All CCs were mechanically removed shortly after oocyte retrieval, washed in culture medium, and frozen immediately prior to total RNA extraction. MII oocytes were used for ICSI. All embryos and blastocysts were assessed daily by the embryologists until 5 days after oocyte retrieval. Embryo quality was assessed at 26 ± 2 and 92 ± 2 hours after insemination. On day 5, the quality evaluations of blastocysts consisted of expansion and hatching status, inner cell mass grading (grade A-C), and trophectoderm grading (grade A-C) [26–28]. Each CC sample included only CCs from a single oocyte. The number of CCs isolated from oocytes at GV, MI, and MII stages and the in vitro embryo outcome for the two patients' groups (HP-hMG or rFSH) are reported in (Figure 1).
Figure 1.
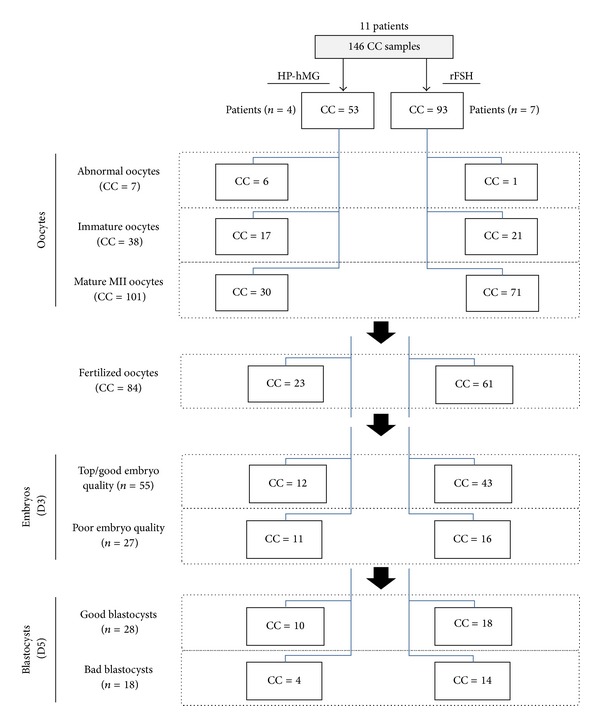
Distribution tree of cumulus cell (CC) samples and embryo outcome relative to the used COS protocol.
2.3. Cumulus Cells RNA Extraction
The RNeasy Micro kit (ref. 74004, Qiagen) was used to extract total RNA from each CCs sample (n = 146) according to the manufacturers' recommended protocols. The quantity and purity of the total RNAs were determined by using a NanoDrop ND-1000 spectrophotometer (NanoDrop ND-Thermo Fisher Scientific, Wilmington, DE, USA) and their integrity by using the Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, http://www.agilent.com/). All RNA samples were stored at −80°C until the microarray experiments.
2.4. Preparation of cRNA and Microarray Hybridization
Total RNA (50 ng) was used to prepare cRNA (one cycle of amplification) using the Affymetrix 3′ IVT express protocol. An oligo-dT primer with a T7 promoter sequence was used to synthesize the first-strand cDNA. After generating the second strand, the complete cDNA was amplified by in vitro transcription (linear amplification) with a T7 RNA polymerase. The amplified RNA (aRNA) was generated and quantified by using a NanoDrop ND-1000 spectrophotometer (NanoDrop ND-Thermo Fisher Scientific, Wilmington, DE, USA), and biotinylated nucleotide analog was incorporated during in vitro transcription step. RNA from the GeneChip Eukaryotic Poly-A RNA Control Kit (Affymetrix, Santa Clara, CA), which contains mRNAs from Bacillus subtilis genes (lys, phe, thr, and dap), was amplified and labeled under the same conditions as positive controls. After fragmentation, the labeled antisense aRNA (15 μg) was hybridized to HG-U133 Plus 2.0 GeneChip pan-genomic oligonucleotide arrays (Affymetrix) containing 54,675 sets of oligonucleotide probes (probeset) which correspond to ≈25,000 unique human genes or predicted genes. Each cumulus cell sample was put individually on a microarray chip. Microarray experiments were performed in DNA microarray platform of our Institute of Research in Biotherapy at the Montpellier University Hospital.
2.5. Data Processing and Gene Expression Profile Analysis
After image processing with the Affymetrix GeneChip Operating 1.4 software (GCOS), the CEL files were analyzed using the Affymetrix Expression Console Software v1.3.1 and normalized with the MAS5.0 algorithm by scaling each array to a target value (TGT) of 100 using the global scaling method to obtain an intensity value signal for each probe set. This algorithm also determines whether a gene is expressed with a defined confidence level or not (“detection call”). This “call” can either be “present” (when the perfect match probes are significantly more hybridized than the mismatch probes, P < 0.04), “marginal” (for P values of >0.04 and <0.06) or “absent” (P > 0.06). Gene annotation was performed using NetAffx (http://www.affymetrix.com/, March 2009). A first selection of microarray data was based on the detection call (present in at least 50% of the CC samples of each group). Then, the Significant Analysis of Microarrays (SAM) (http://www-stat.stanford.edu/~tibs/SAM/) with the Wilcoxon test and sample label permutation (n = 300) was used to identify genes of which expression varied significantly between the HP-hMG and rFSH CC samples. The lists of significant genes (fold change, FC ≥1.5 and false discovery rate, FDR ≤5%) as well as common genes were analyzed using the Ingenuity Pathway Analysis (IPA) software (http://www.ingenuity.com/) to identify the biological functions that were specific of each CC group and in common between the two treatments, respectively. Only annotations with significant P value (P < 0.05) were considered. Then, the SAM analysis (FC ≥1.5, FDR ≤5%) was used to link gonadotropin-specific genes in CCs or those that are irrespective of gonadotropin treatment to subsequent embryo outcome at day 3 (top, good embryo versus poor) or day 5 (good blastocyst versus bad). Hierarchical clustering analyses based on the expression levels of the differentially expressed genes were performed by using the Cluster and Treeview software packages [29]. Box-and-whisker plots depicted the comparisons of the expression levels of candidate genes carried out using SPSS 12.0 (SPSS, Chicago, IL, USA) software.
2.6. Microarray Data Validation by Quantitative RT-PCR
Quantitative RT-PCR was performed to validate the expression of selected genes identified as differentially expressed between the two CC groups by using mRNAs from HP-hMG (n = 4) and rFSH (n = 4) CC samples as described in [30]. The primer sequences are shown in (Supplementary data, Table SII). Briefly, cDNA was reverse transcribed (RT) following the manufacturer's instructions using 500 ng of amplified RNA in a 20 μL reaction volume that included Superscript II (ref. 18064-014, Invitrogen), oligo-dT primer, dNTP mixture, MgCl2, and RNase inhibitor. Quantitative PCR was performed using a LightCycler 480 apparatus with the LC480 SYBR Green I Master kit (Roche Diagnostics, Mannheim, Germany) and 2 μL of diluted cDNA (1/25) and 0.6 mMol primers in a total volume of 10 μL. After 10 min of activation at 95°C, cycling conditions were 10 s at 95°C, 30 s at 63°C, and 1 s at 72°C for 45 cycles. Gene expression levels were normalized to the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH), because its expression was stable between all CC groups using the following formula 100/2ΔΔCt, where ΔΔCt = ΔCt unknown − ΔCt positive control.
2.7. Statistical Analysis
Statistical analyses were performed with SPSS 12.0 software. A repartition difference between sample groups was considered significant when the Kruskal-Wallis nonparametric test and Wilcoxon test gave a P value ≤0.05. For q-RT-PCR, a statistical analysis was performed with the GraphPad InStat software (Mann-Whitney U test; GraphPad, San Diego, CA). A value of P ≤ 0.05 was considered to be statistically significant.
3. Results
3.1. Identification of Differentially Expressed Genes in Human CCs following Stimulation with HP-hMG or rFSH
A first selection is based on the detection call between all the CC samples from patients stimulated with HP-hMG or rFSH delineated 9,899 genes. Then, using SAM, 94 genes that significantly differentiated between HP-hMG and rFSH CCs were identified. Among them, 45 and 49 genes were upregulated in HP-hMG and rFSH CC samples, respectively (fold-change, FDR, and annotation are in Tables 1 and 2). The HP-hMG CC list included genes implicated in lipid metabolism such as GM2A (x2.3, FDR = 0), AKR1C1 (x1.5, FDR = 0), AKR1C2 (x1.6, FDR = 0.005), and in cell-to-cell interaction like GJA5 (x1.9, FDR = 0), NTS (x1.8, FDR = 0.005), FOS (x1.6, FDR = 0), and NPY1R (x2.1, FDR = 0), NPY2R (x1.6, FDR = 0). Conversely, the rFSH CC list was significantly enriched in genes important for cellular assembly and organization such as COL3A1 (x2, FDR = 0.015), COL1A1 (x1.5; FDR = 0), MT3 (x1.5; FDR = 0), and CAMK1D (x1.5; FDR = 0). Other genes of the rFSH list are members of the tumour necrosis factor (TNF) family such as TNFAIP6 (x1.7; FDR = 0.01) and TNFAIP8 (x1.6, FDR = 0.005). The clustering based on these 94 genes segregates the majority of the HP-hMG (85%) from the rFSH CC samples (Figure 2). RT-qPCR validated the differential expression of some of these genes (Supplementary data, Figure SI).
Table 1.
List of genes that were significantly upregulated in HP-hMG CCs compared with rFSH CCs.
| Gene name | Gene title | Probesets | Fold change | FDR (%) |
|---|---|---|---|---|
| PHACTR2 | Phosphatase and actin regulator 2 | 244774_at | 2.9 | 0 |
| GM2A | GM2 ganglioside activator | 235678_at | 2.3 | 0 |
| LOC654433 | Homo sapiens, clone IMAGE:4826696, mRNA | 228425_at | 2.2 | 0 |
| LOC201651 | Similar to esterase/N-deacetylase (EC 3.5.1.-), 50 K hepatic-rabbit | 1569582_at | 2.1 | 0 |
| PAX8 | Transcribed locus, moderately similar to XP_375099.1 hypothetical protein LOC283585 (Homo sapiens) | 227474_at | 2.1 | 0 |
| NPY1R | Neuropeptide Y receptor Y1 | 205440_s_at | 2.1 | 0 |
| GJA5 | Gap junction protein, alpha 5, 40 kDa (connexin 40) | 226701_at | 1.9 | 0 |
| FOXG1B | Forkhead box G1B | 206018_at | 1.9 | 0 |
| SPP1 | Secreted phosphoprotein 1 | 209875_s_at | 1.9 | 0.58 |
| NTS | Neurotensin | 206291_at | 1.8 | 0.58 |
| THAP4 | THAP domain containing 4 | 220417_s_at | 1.8 | 0 |
| SPESP1 | Sperm equatorial segment protein 1 | 229352_at | 1.8 | 0.58 |
| SEMA6D | Sema domain, transmembrane domain (TM), and cytoplasmic domain, (semaphorin) 6D | 233882_s_at | 1.8 | 0.58 |
| DOCK8 | Dedicator of cytokinesis 8 | 225502_at | 1.8 | 0.58 |
| SERPINB2 | Serine (or cysteine) proteinase inhibitor, clade B (ovalbumin), member 2 | 204614_at | 1.7 | 0.58 |
| PPP1R14C | Protein phosphatase 1, regulatory (inhibitor) subunit 14C | 226907_at | 1.7 | 0 |
| CTIF | CBP80/20-dependent translation initiation factor | 243090_at | 1.7 | 0 |
| SSFA2 | Sperm-specific antigen 2 | 236207_at | 1.7 | 0 |
| HS3ST1 | Heparan sulfate (glucosamine) 3-O-sulfotransferase 1 | 205466_s_at | 1.7 | 0 |
| CYP1B1 | Cytochrome P450, family 1, subfamily B, polypeptide 1 | 202437_s_at | 1.7 | 0 |
| TMEM37 | Transmembrane protein 37 | 1554485_s_at | 1.6 | 0 |
| BBS12 | Hypothetical protein FLJ35630 | 229603_at | 1.6 | 0 |
| AKR1C2 | Aldo-keto reductase family 1, member C2 | 211653_x_at | 1.6 | 0.58 |
| MALL | BENE protein | 209373_at | 1.6 | 0 |
| NPY2R | Neuropeptide Y receptor Y2 | 210729_at | 1.6 | 0 |
| METTL7B | Hypothetical protein MGC17301 | 227055_at | 1.6 | 0 |
| RNF128 | Ring finger protein 128 | 219263_at | 1.6 | 0 |
| ARL4C | ADP-ribosylation factor-like 7 | 202207_at | 1.6 | 0 |
| PAPPA | Pregnancy-associated plasma protein A, pappalysin 1 | 240450_at | 1.6 | 0 |
| USP45 | Ubiquitin-specific protease 45 | 224441_s_at | 1.6 | 0 |
| FOS | v-fos FBJ murine osteosarcoma viral oncogene homolog | 209189_at | 1.6 | 0 |
| PDK4 | Pyruvate dehydrogenase kinase, isozyme 4 | 225207_at | 1.6 | 0 |
| ZNF718 | Hypothetical protein FLJ90036 | 1553269_at | 1.6 | 0 |
| ARHGAP20 | Rho GTPase activating protein 20 | 228368_at | 1.5 | 0 |
| FLJ43663 | CDNA FLJ26188 fis, clone ADG04821 | 238619_at | 1.5 | 0 |
| HOP | Homeodomain-only protein | 211597_s_at | 1.5 | 0 |
| ENPP2 | Ectonucleotide pyrophosphatase/phosphodiesterase 2 (autotaxin) | 209392_at | 1.5 | 2.95 |
| LYZ | Lysozyme (renal amyloidosis) | 213975_s_at | 1.5 | 1.05 |
| SKAP2 | src family associated phosphoprotein 2 | 204361_s_at | 1.5 | 0 |
| ABHD12 | Chromosome 20 open reading frame 22 | 228124_at | 1.5 | 0 |
| RUNX1 | Runt-related transcription factor 1 | 236114_at | 1.5 | 0 |
| AKR1C1 | Aldo-keto reductase family 1, member C2 | 216594_x_at | 1.5 | 0 |
| BRE | Brain and reproductiveorgan-expressed(TNFRSF1A modulator) | 211566_x_at | 1.5 | 0 |
| SERPINI1 | Serine (or cysteine) proteinase inhibitor, clade I (neuroserpin), member 1 | 205352_at | 1.5 | 0 |
| RASL11B | RAS-like, family 11, member B | 219142_at | 1.5 | 0 |
Table 2.
List of genes that were significantly upregulated in rFSH CCs compared with HP-hMG CCs.
| Gene name | Gene title | Probesets | Fold change | FDR (%) |
|---|---|---|---|---|
| ITM2A | Integral membrane protein 2A | 202746_at | 4.2 | 0 |
| H19 | H19, imprinted maternally expressed transcript (nonprotein coding) | 224646_x_at | 3.8 | 0 |
| PSPH | Phosphoserine phosphatase | 205048_s_at | 2.4 | 0 |
| GAL | Galanin | 214240_at | 2.4 | 0 |
| ZNF528 | Zinc finger-like | 232315_at | 2.3 | 0 |
| NFKBIZ | Nuclear factor of kappa light polypeptide gene enhancer in B-cell inhibitor, zeta | 223217_s_at | 2.2 | 4.73 |
| FAM84B | Breast cancer membrane protein 101 | 225864_at | 2 | 0 |
| COL3A1 | Collagen, type III, alpha 1 (Ehlers-Danlos syndrome type IV, autosomal dominant) | 211161_s_at | 2 | 1.53 |
| DKFZp451A211 | DKFZp451A211 protein | 1556114_a_at | 1.8 | 0 |
| SPARCL1 | SPARC-like 1 (mast9, hevin) | 200795_at | 1.8 | 0 |
| PTER | Phosphotriesterase related | 222798_at | 1.8 | 0 |
| NFIB | Nuclear factor I/B | 213032_at | 1.8 | 0 |
| MXRA5 | Adlican | 209596_at | 1.8 | 0 |
| GALNTL2 | UDP-N-acetyl-alpha-D-galactosamine:polypeptide N-acetylgalactosaminyltransferase-like 2 | 228501_at | 1.8 | 0 |
| SUPT3H | Suppressor of Ty 3 homolog (S. cerevisiae) | 211106_at | 1.7 | 0 |
| DDX17 | DEAD (Asp-Glu-Ala-Asp) box polypeptide 17 /// DEAD (Asp-Glu-Ala-Asp) box polypeptide 17 | 208151_x_at | 1.7 | 4.15 |
| TNFAIP6 | Tumor necrosis factor, alpha-induced protein 6 | 206026_s_at | 1.7 | 1.05 |
| MTUS1 | Mitochondrial tumor suppressor 1 | 212096_s_at | 1.7 | 4.73 |
| RP1-93H18.5 | Similar to RIKEN cDNA A630077B13 gene, RIKEN cDNA 2810048G17 | 229390_at | 1.7 | 0 |
| LOC92196 | Hypothetical LOC92196 (uncharacterized) | 229290_at | 1.6 | 0 |
| LOC401317 | Hypothetical LOC402472 (uncharacterized) | 242329_at | 1.6 | 0 |
| CHAC1 | Hypothetical protein MGC4504 | 219270_at | 1.6 | 0 |
| STRN3 | Striatin, calmodulin binding protein 3 | 215505_s_at | 1.6 | 0 |
| OSBPL10 | Oxysterol binding protein-like 10 | 219073_s_at | 1.6 | 0 |
| GLIPR1 | HIV-1 rev binding protein 2 | 214085_x_at | 1.6 | 0 |
| BTRC | Beta-transducin repeat containing E3 ubiquitin protein ligase | 237862_at | 1.6 | 0 |
| TNFAIP8 | Tumor necrosis factor, alpha-induced protein 8 | 208296_x_at | 1.6 | 0.54 |
| PMAIP1 | Phorbol-12-myristate-13-acetate-induced protein 1 | 204286_s_at | 1.6 | 0 |
| RBM24 | RNA binding motif protein 24 | 235004_at | 1.6 | 1.53 |
| LOC388796 | Hypothetical LOC388796 (uncharacterized) | 65588_at | 1.6 | 0 |
| LOC157278 | Homo sapiens, clone IMAGE:5285282, mRNA (uncharacterized) | 238716_at | 1.6 | 0 |
| GREM1 | Gremlin 1 | 218468_s_at | 1.6 | 0 |
| OSBPL6 | Oxysterol binding protein-like 6 | 223805_at | 1.6 | 0 |
| CREB5 | cAMP responsive element binding protein 5 | 205931_s_at | 1.5 | 0 |
| CAMK1D | Calcium/calmodulin-dependent protein kinase ID | 235626_at | 1.5 | 0 |
| CCDC58 | Hypothetical LOC131076 | 235244_at | 1.5 | 0 |
| LRRN3 | Leucine-rich repeat neuronal 3 | 209840_s_at | 1.5 | 0 |
| HS3ST3A1 | Heparan sulfate (glucosamine) 3-O-sulfotransferase 3A1 | 219985_at | 1.5 | 0 |
| ARSD | Arylsulfatase D | 232423_at | 1.5 | 0 |
| ENDOD1 | KIAA0830 protein | 212570_at | 1.5 | 0 |
| ZNF521 | Zinc finger protein 521 | 226676_at | 1.5 | 0 |
| DFNA5 | Deafness, autosomal dominant 5 | 203695_s_at | 1.5 | 0 |
| PSD3 | Pleckstrin and Sec7 domain containing 3 | 203354_s_at | 1.5 | 0 |
| LOC283070 | Hypothetical protein LOC283070 (uncharacterized) | 226382_at | 1.5 | 0 |
| COL1A1 | Collagen, type I, alpha 1 | 1556499_s_at | 1.5 | 0 |
| SPOCK2 | Sparc/osteonectin, cwcv and kazal-like domains proteoglycan (testican) 2 | 202523_s_at | 1.5 | 0 |
| ATP7A | ATPase, Cu++ transporting, alpha polypeptide (Menkes syndrome) | 205197_s_at | 1.5 | 0 |
| MT3 | Metallothionein 3 (growth inhibitory factor (neurotrophic)) | 205970_at | 1.5 | 0 |
| DDIT3 | DNA-damage-inducible transcript 3 | 209383_at | 1.5 | 0 |
Figure 2.

Gene expression patterns of the HP-hMG and rFSH CC samples. Supervised hierarchical clustering of CC samples based on the 94 genes that are differentially expressed between the two treatment groups (HP-hMG and rFSH). We can see a distinct signature in each CCs category. The color intensity indicates the level of gene expression (red for upregulated genes and green for downregulated genes).
3.2. Common Transcriptional Gene Profile in HP-hMG/rFSH CCs
In view of few differences between the two gonadotropin treatments, we examined the list of genes in common to HP-hMG and rFSH groups (list of 9,805 genes; see Supplementary data, Table SIII). We used IPA software to explore the specific functional properties of this common molecular signature. Estrogen receptor signaling (83 genes) (P value = 8.17E − 08) was one of the top canonical pathways related to this molecular signature. On the other hand, the top network involving 35 genes was articulated around the “cell death and survival, DNA replication, recombination, and repair” functions. The detailed list of genes involved in this network can be found in (Supplementary data, Table SIV). Interestingly, the most common HP-hMG/rFSH genes were associated with multiple signaling pathways including FGF signaling (FGFR and GRB2), IGF signaling (IGF1R and IGFBP3), EGF signaling (EGFR and MAPK1), and PDGF signaling (PDGFRA and PDGFD). It is important to note that no difference was observed in the mRNA CC level between treatments for receptors (LHCGR and BMPR2), aromatase (CYP19A1), cytochrome P450 (CYP11A1), or steroidogenic genes (StAR, HSD3B2, ACVR1, ACVR1B, INHBC, and INHBB).
3.3. Relationship between the HP-hMG or rFSH CC Expression Profiles and In Vitro Embryo Development
Of the 146 CC samples, 101 were isolated from MII mature oocytes which underwent ICSI. In the HP-hMG group, 77% of injected oocytes were fertilized and 61% achieved blastocyst stage at day 5. In the rFSH group, these values were, respectively, 86% and 52%. Fertilized MII oocytes (n = 23 in the HP-hMG and n = 61 in the rFSH group) were divided into oocytes that developed into (i) top/good quality (52% in the HP-hMG and 70% in the rFSH group, no significant difference (ε = 1.65)) or poor quality embryos at day 3; and then into (ii) good (AA and AB) (43% for the HP-hMG and 29% for the rFSH group, no significant difference (ε = 1.28)) or bad grade (AC, BC, CC, and CB) blastocysts at day 5 (Figure 1). Then, the transcription profile of the cumulus cell samples isolated from these 101 MII oocytes was evaluated relative to day 3 embryo quality and blastocyst grading at day 5. In the HP-hMG group, NPY1R (x1.58, FDR = 0.0004) and NPY2R (x1.67, FDR = 0.0004) upregulation was observed in CCs isolated from MII oocytes that developed into top/good day 3 embryos, whereas GM2A (x2.10, FDR = 0.0005) and USP45 (x2.32, FDR = 0.0005) were upregulated in cumulus cells from MII oocytes with good blastocyst grading (Figure 3(a)). After rFSH treatment, upregulation of GREM1 (x1.59, FDR = 0) and PSPH (x1.6, FDR = 0) was significantly associated with top/good quality day 3 embryos; OSBPL6 (x1.59, FDR = 0) upregulation was found in CCs from oocytes that developed into good blastocyst at day 5 (Figure 3(b)). In the two gonadotropin groups, PTX3 (x-1.81, FDR = 0) downregulation and STC2 (x1.76, FDR = 0) upregulation were observed in CCs isolated from MII oocytes that developed into top/good day 3 embryos, whereas TRIM65 (x-1.62, FDR = 0) and GSTM2 (x-1.67, FDR = 0) expressions were downregulated in CCs associated with good blastocyst grading (Figure 3(c)).
Figure 3.

Gonadotropin gene expression associated with in vitro embryo development. (a) and (b) Box-and-whisker plots comparing the expression level of gonadotropin-specific gene in CCs from oocytes that developed into top/good quality embryos (n = 43 in the rFSH and n = 12 in the HP-hMG group) or poor quality embryos (n = 16 in the rFSH and n = 11 in the HP-hMG group) and into good blastocysts (n = 18 in the rFSH and n = 10 in the HP-hMG group) or bad blastocysts (n = 14 in the rFSH and n = 4 in the HP-hMG group). (c) Box-and-whisker plots comparing the expression level of gonadotropin common genes in CCs from oocytes that developed into top/good quality embryos (n = 55 CCs) or poor quality embryos (n = 27 CCs) and into good blastocysts (n = 28 CCs) or bad blastocysts (n = 18 CCs). The signal intensity for each gene is shown on the y-axis as arbitrary units determined by the Affymetrix GCOS software. *A significant difference with FDR ≤0.05.
3.4. CC mRNA Content and In Vitro Blastocyst Outcome at Day 5
Independently of the type of gonadotropin treatment used, the relation between amplified mRNA content of CC samples and in vitro blastocyst development at day 5 was also investigated. Seventeen CC samples, isolated from MII oocytes that developed into top quality 8-cell embryos at day 3, were selected and divided in three groups: (i) CCs from MII oocytes that developed into good quality (grade AA-AB, n = 7), (ii) intermediary (grade BB, n = 6), and (iii) bad (grade CC and others, n = 4) blastocysts. The amount (mean ± SEM) of amplified mRNA from CCs from MII oocytes leading to good quality blastocysts was 1044.28 ± 159.18 ng/μL. This value decreased to 796.66 ± 150 ng/μL in the intermediary group and to 627.50 ± 76.25 ng/μL in the bad blastocyst grade group (Figure 4).
Figure 4.
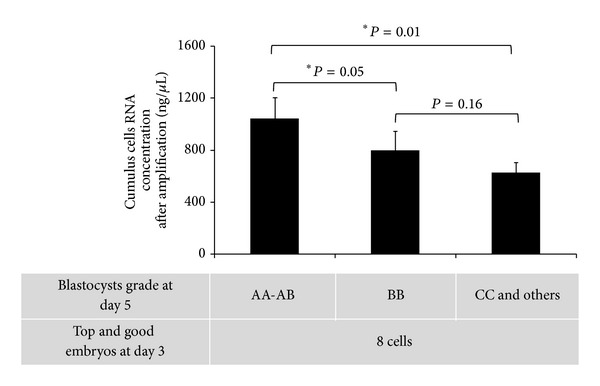
Relationship between amount of amplified CCs mRNA and blastocyst quality. Three groups of blastocysts (good, intermediary, or bad quality) were obtained from top and good 8-cell embryos at day 3. The Kruskal-Wallis test was used to indicate that at least one of the groups is different from the others (P = 0.011, Kruskal-Wallis test), and the Wilcoxon test was used to establish whether group AA-AB is significantly different from group BB and/or group CC. *A significant difference in the concentration of amplified CC mRNA between two groups of blastocysts. CC samples (n = 17) were from oocytes that developed in top and good 8-cell embryos at day 3. AA-AB: good blastocyst grades (n = 7); BB: intermediary blastocyst grades (n = 6); CC and others: bad blastocyst grades (n = 4). Bars represent the mean ± SEM.
4. Discussion
Following global genomic assessment of 146 human CCs transcriptome under HP-hMG and rFSH treatments, the present study revealed a small but significant distinct molecular signature of 94 genes between the two treatments, suggesting that these treatments impact differentially the CC gene expression profile. This may be accounted for by the differences in the origin of the two pharmaceutical preparations. More precisely, overexpression of genes involved in the metabolism of lipids such as GM2A, AKR1C1 and AKR1C2, as well as genes related to the intercellular signaling (GJA5 and FOS) was observed in the CCs treated with HP-hMG, while genes involved in “cellular assembly and organization” (COL1A1, COL3A1, MT3, TNFAIP6, and TNFAIP8) were overexpressed in the rFSH CCs. Each of these functions plays a central role in oocyte maturation and/or oocyte competence [31–33]. Indeed, the metabolism of lipids represents the main energy source for protein synthesis during oocyte nuclear maturation and early embryo development [34, 35]. Simultaneously, adequate communication between oocyte and CCs and appropriate assembly and organization of the CC matrix are required for both oocyte maturation and competence [36–38]. Most of the genes, identified in the present investigation as differentially expressed in CCs treated with HP-hMG and rFSH, were reported for the first time, except for TNFAIP6 and GJA5 (connexin 40) which have been previously identified as potential markers of oocyte competence in CCs from bovine preovulatory follicles [39] and biomarker of oocyte maturation in canine cumulus-oocyte complexes matured in vitro, respectively [38].
Furthermore, the comparison of our data with the two other transcriptomic studies comparing the same gonadotropin treatment in granulosa cells (GCs) using the GnRH agonist long protocols [24, 25] indicates that GM2 ganglioside activator is upregulated in HP-hMG CCs (this study) and rFSH GCs [24]. GM2A is known to play an important role in the hydrolysis of phospholipids or small glycolipids [40]. In addition, among the 9 common genes of our study and the one by Brannian et al. [25], six genes (ATP7A, BTRC, LRRN3, STRN3, PTER, and SUPT3) are upregulated in both CCs and GCs after rFSH treatment; one (H19) was upregulated in both rFSH CCs and HP-hMG GCs and the two others (SERPINI1 and SSFA2) in HP-hMG CCs and rFSH. The use of different GnRH analogs might explain these discrepancies, but we cannot exclude the possibility that gonadotropin stimulation might have different effects on CCs and GCs. More investigations are required to address this issue.
On the other hand, we reported an important common CC molecular signature revealing the preservation of numerous growth factor signaling between the two types of treatments including the IGF, PDGF, FGF, and EGF pathways (See Figure SIII). These signaling pathways have been previously reported to play a central role in the control of the intrafollicular androgen/estrogen ratio for the IGF members [41], in angiogenesis and embryo development for the FGF and PDGF members [42] and in oocyte maturation for the members of the EGF family [43–45]. The interactions between these signaling pathways in CCs under COS will be a precious itinerary to explore in future works in order to complete the oocyte competence puzzle.
Another important finding of this study is that the mRNA level for key genes involved in ovulation process including hormonal receptors (LHCGR and BMPR2) and regulators of steroidogenesis (StAR, HSD3B2, Activins, and Inhibins) was comparable in the HP-hMG and rFSH CC groups. This suggests a similar potency of the two protocols to induce hormonal receptors and similar estrogenic capacity of the CC samples stimulated by HP-hMG and rFSH. This is in line with several studies reporting that CCs in vitro were able to secrete estradiol during COCs culture from patients undergoing stimulated cycles, probably as a consequence of the action of gonadotropins [46].
We also identified a significant relationship between some CC genes that were specifically upregulated following stimulation with HP-hMG or rFSH and in vitro embryo development. In the HP-hMG group, upregulation of NPY1R and NPY2R in CCs was associated with top/good embryo quality at day 3. NPY modulates steroid production through NPY receptors [47] and plays a role in human ovarian steroidogenesis directly at the level of the granulosa cells of the follicles in the early stage of luteinization [48, 49]. Additionally, the association of ubiquitin specific protease 45 (USP45) with good blastocyst quality suggests the requirement of proteasomal activity in HP-hMG-treated CCs. Proteasomal activity has been reported to have multiple functions in CCs expansion, in oocyte meiosis, and in the modification of cumulus-oocyte communication [50].
In the rFSH group, upregulation of gremlin 1 (GREM1) in CCs was associated with top/good embryo quality at day 3 and OSBPL6 upregulation with good blastocyst grading at day 5. Only CC expression of GREM1, a member of the bone morphogenic protein (BMP) antagonist family, has been reported as positively correlated with embryo quality [7, 12, 51]. The regulation of BMP through GREM1 is thought to contribute to CCs expansion and therefore to the final maturation of oocytes [52]. The gene OSBPL6 codes for the oxysterol binding protein-like-6 receptor. Oxysterols, which bind to this receptor, are potent modulators of expression of cholesterol synthesis in human granulosa cells [53]. Recently, Watanabe et al. [54] reported that variation in cholesterol contents in cumulus-oocyte complexes during in vitro maturation of porcine oocytes affected their ability to be fertilized, suggesting that, under rFSH regime, cholesterogenesis at a nearby site of oocyte growth and maturation might also be involved in in vitro blastocyst outcome.
On the other hand, we also identified CC genes associated with day 3 embryo quality and blastocyst grading at day 5, independently of the type of gonadotropins. Among these genes, we report for the first time the expression of STC2, GSTM2, and TRIM65, as well as PTX3 which has been shown in previous studies to either be associated with fertilization rate [55] or to have no relationship with high-quality embryo on day 3 [51]. A possible reason for higher stanniocalcin 2 (STC2) expression in the CCs isolated from MII oocytes that developed into top/good day 3 is the modulation of the angiogenic [56] or steroidogentic pathways [57] or principal processes in ovarian function [58–60]. Conversely, we observed an increased expression of GSTM2 and TRIM65 in CCs from oocytes that developed into bad blastocyst grading. GSTM2 and TRIM65 play a role in the protection against lipid peroxidation [61] and in DNA repair [62] respectively, suggesting an increase in cellular resistance against oxidative stress and damaged DNA. The implications of these genes, at the CC level, deserve to be addressed in future studies in order to understand their function in follicular growth.
Furthermore, independently of the type of gonadotropin treatment, we found an association between blastocyst grading at day 5 and the amount of amplified mRNA in CC samples from MII mature oocytes with comparable top/good embryo quality at day 3. Lower mRNA values were detected in CCs from MII oocytes that developed into bad blastocysts as compared to CC samples from oocytes that developed into intermediary or good quality blastocysts at day 5. This suggests that CCs surrounding an incompetent oocyte are less transcriptionally active.
These results are in line with our previously published data showing a general reduction in transcriptomic activity of CCs associated with poor oocyte competence and negative clinical outcome [6].
5. Conclusion
Analysis of the microarray data of CCs from patients, who underwent GnRH-antagonist COS, highlights a significant difference in the gene expression profile of CCs following treatment with HP-hMG or rFSH. Components of signaling pathways (the EGF, IGF, FGF, and PDGF cascades) were conserved in CCs under the two gonadotropin stimulation regimens. Some genes specific to each gonadotropin treatment or commonly expressed in both groups were associated with in vitro embryo development. Moreover, independently of the gonadotropin preparation used, the amount of amplified mRNA in each CC was associated with blastocyst grading at day 5. These genes may prove valuable as biomarkers of in vitro embryo quality and can be useful for understanding the biology of stimulation.
Supplementary Material
Supplementary Material available online includes: (i) Baseline clinical characteristics, end-of-stimulation data and number of oocytes retrieved from each patient following controlled ovarian stimulation with HP-hMG or rFSH. (ii) Primer pairs used for validation of the array data by qRT-PCR. (iii) List of the 9,805 common genes expressed in both HP-hMG and rFSH CC samples. (iv) Validation by qRT-PCR of some of the genes that were differentially expressed in HP-hMG and rFSH CC samples. (v) Top-ranked networks evidenced in common HP-hMG/rFSH signature by Ingenuity Pathway software. (vi) Detailed list of the genes presented in the network. (vii) The major signaling pathways that occur in CCs following the gonadotropin (HP-hMG and rFSH) stimulation.
Acknowledgments
This work was supported by Ferring Pharmaceuticals A/S. The authors thank the direction of the Montpellier 1 University, University Hospital of Montpellier for their support, and Dr. Aït-ahmed Ounissa for the insightful discussions and the critical review of the paper.
References
- 1.Trew GH, Brown AP, Gillard S, et al. In vitro fertilisation with recombinant follicle stimulating hormone requires less IU usage compared with highly purified human menopausal gonadotrophin: results from a European retrospective observational chart review. Reproductive Biology and Endocrinology. 2010;8, article 137 doi: 10.1186/1477-7827-8-137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.van Wely M, Kwan I, Burt AL, et al. Recombinant versus urinary gonadotrophin for ovarian stimulation in assisted reproductive technology cycles. A cochrane review. Human Reproduction Update. 2012;18(2):p. 111. doi: 10.1093/humupd/dmr048. [DOI] [PubMed] [Google Scholar]
- 3.van Wely M, Kwan I, Burt AL, et al. Recombinant versus urinary gonadotrophin for ovarian stimulation in assisted reproductive technology cycles. Cochrane Database of Systematic Reviews. 2011;(2) doi: 10.1002/14651858.CD005354.pub2.CD005354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hutt KJ, Albertini DF. An oocentric view of folliculogenesis and embryogenesis. Reproductive BioMedicine Online. 2007;14(6):758–764. doi: 10.1016/s1472-6483(10)60679-7. [DOI] [PubMed] [Google Scholar]
- 5.Albertini DF, Combelles CMH, Benecchi E, Carabatsos MJ. Cellular basis for paracrine regulation of ovarian follicle development. Reproduction. 2001;121(5):647–653. doi: 10.1530/rep.0.1210647. [DOI] [PubMed] [Google Scholar]
- 6.Assou S, Haouzi D, Mahmoud K, et al. A non-invasive test for assessing embryo potential by gene expression profiles of human cumulus cells: a proof of concept study. Molecular Human Reproduction. 2008;14(12):711–719. doi: 10.1093/molehr/gan067. [DOI] [PubMed] [Google Scholar]
- 7.McKenzie LJ, Pangas SA, Carson SA, et al. Human cumulus granulosa cell gene expression: a predictor of fertilization and embryo selection in women undergoing IVF. Human Reproduction. 2004;19(12):2869–2874. doi: 10.1093/humrep/deh535. [DOI] [PubMed] [Google Scholar]
- 8.van Montfoort APA, Geraedts JPM, Dumoulin JCM, Stassen APM, Evers JLH, Ayoubi TAY. Differential gene expression in cumulus cells as a prognostic indicator of embryo viability: a microarray analysis. Molecular Human Reproduction. 2008;14(3):157–168. doi: 10.1093/molehr/gam088. [DOI] [PubMed] [Google Scholar]
- 9.Gebhardt KM, Feil DK, Dunning KR, Lane M, Russell DL. Human cumulus cell gene expression as a biomarker of pregnancy outcome after single embryo transfer. Fertility and Sterility. 2011;96(1):47.e2–52.e2. doi: 10.1016/j.fertnstert.2011.04.033. [DOI] [PubMed] [Google Scholar]
- 10.Hamel M, Dufort I, Robert C, et al. Identification of differentially expressed markers in human follicular cells associated with competent oocytes. Human Reproduction. 2008;23(5):1118–1127. doi: 10.1093/humrep/den048. [DOI] [PubMed] [Google Scholar]
- 11.Assou S, Boumela I, Haouzi D, et al. Dynamic changes in gene expression during human early embryo development: from fundamental aspects to clinical applications. Human Reproduction Update. 2011;17(2):272–290. doi: 10.1093/humupd/dmq036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Anderson RA, Sciorio R, Kinnell H, et al. Cumulus gene expression as a predictor of human oocyte fertilisation, embryo development and competence to establish a pregnancy. Reproduction. 2009;138(4):629–637. doi: 10.1530/REP-09-0144. [DOI] [PubMed] [Google Scholar]
- 13.Adriaenssens T, Wathlet S, Segers I, et al. Cumulus cell gene expression is associated with oocyte developmental quality and influenced by patient and treatment characteristics. Human Reproduction. 2010;25(5):1259–1270. doi: 10.1093/humrep/deq049. [DOI] [PubMed] [Google Scholar]
- 14.Ouandaogo ZG, Haouzi D, Assou S, et al. Human cumulus cells molecular signature in relation to oocyte nuclear maturity stage. PLoS One. 2011;6(11) doi: 10.1371/journal.pone.0027179.e27179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Assou S, Anahory T, Pantesco V, et al. The human cumulus-oocyte complex gene-expression profile. Human Reproduction. 2006;21(7):1705–1719. doi: 10.1093/humrep/del065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fragouli E, Wells D, Iager AE, Kayisli UA, Patrizio P. Alteration of gene expression in human cumulus cells as a potential indicator of oocyte aneuploidy. Human Reproduction. 2012;27:2559–2568. doi: 10.1093/humrep/des170. [DOI] [PubMed] [Google Scholar]
- 17.Assou S, Haouzi D, de Vos J, Hamamah S. Human cumulus cells as biomarkers for embryo and pregnancy outcomes. Molecular Human Reproduction. 2010;16(8):531–538. doi: 10.1093/molehr/gaq032. [DOI] [PubMed] [Google Scholar]
- 18.Balasch J, Peñarrubia J, Fábregues F, et al. Ovarian responses to recombinant FSH or HMG in normogonadotrophic women following pituitary desensitization by a depot GnRH agonist for assisted reproduction. Reproductive BioMedicine Online. 2003;7(1):35–42. doi: 10.1016/s1472-6483(10)61726-9. [DOI] [PubMed] [Google Scholar]
- 19.Kilani Z, Dakkak A, Ghunaim S, et al. A prospective, randomized, controlled trial comparing highly purified hMG with recombinant FSH in women undergoing ICSI: ovarian response and clinical outcomes. Human Reproduction. 2003;18(6):1194–1199. doi: 10.1093/humrep/deg252. [DOI] [PubMed] [Google Scholar]
- 20.Platteau P, Andersen AN, Balen A, et al. Similar ovulation rates, but different follicular development with highly purified menotrophin compared with recombinant FSH in WHO Group II anovulatory infertility: a randomized controlled study. Human Reproduction. 2006;21(7):1798–1804. doi: 10.1093/humrep/del085. [DOI] [PubMed] [Google Scholar]
- 21.Daya S. Updated meta-analysis of recombinant follicle-stimulating hormone (FSH) versus urinary FSH for ovarian stimulation in assisted reproduction. Fertility and Sterility. 2002;77(4):711–714. doi: 10.1016/s0015-0282(01)03246-0. [DOI] [PubMed] [Google Scholar]
- 22.Coomarasamy A, Afnan M, Cheema D, van der Veen F, Bossuyt PMM, van Wely M. Urinary hMG versus recombinant FSH for controlled ovarian hyperstimulation following an agonist long down-regulation protocol in IVF or ICSI treatment: a systematic review and meta-analysis. Human Reproduction. 2008;23(2):310–315. doi: 10.1093/humrep/dem305. [DOI] [PubMed] [Google Scholar]
- 23.Ziebe S, Lundin K, Janssens R, Helmgaard L, Arce J-C. Influence of ovarian stimulation with HP-hMG or recombinant FSH on embryo quality parameters in patients undergoing IVF. Human Reproduction. 2007;22(9):2404–2413. doi: 10.1093/humrep/dem221. [DOI] [PubMed] [Google Scholar]
- 24.Grøndahl ML, Borup R, Lee YB, Myrhøj V, Meinertz H, Sørensen S. Differences in gene expression of granulosa cells from women undergoing controlled ovarian hyperstimulation with either recombinant follicle-stimulating hormone or highly purified human menopausal gonadotropin. Fertility and Sterility. 2009;91(5):1820–1830. doi: 10.1016/j.fertnstert.2008.02.137. [DOI] [PubMed] [Google Scholar]
- 25.Brannian J, Eyster K, Mueller BA, Bietz MG, Hansen K. Differential gene expression in human granulosa cells from recombinant FSH versus human menopausal gonadotropin ovarian stimulation protocols. Reproductive Biology and Endocrinology. 2010;8, article 25 doi: 10.1186/1477-7827-8-25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Devroey P, Pellicer A, Nyboe Andersen A, Arce J-C. A randomized assessor-blind trial comparing highly purified hMG and recombinant FSH in a GnRH antagonist cycle with compulsory single-blastocyst transfer. Fertility and Sterility. 2012;97(3):561–571. doi: 10.1016/j.fertnstert.2011.12.016. [DOI] [PubMed] [Google Scholar]
- 27.Gardner DK, Schoolcraft WB. Culture and transfer of human blastocysts. Current Opinion in Obstetrics and Gynecology. 1999;11(3):307–311. doi: 10.1097/00001703-199906000-00013. [DOI] [PubMed] [Google Scholar]
- 28.Gardner DK, Schoolcraft WB, Wagley L, Schlenker T, Stevens J, Hesla J. A prospective randomized trial of blastocyst culture and transfer in in-vitro fertilization. Human Reproduction. 1998;13(12):3434–3440. doi: 10.1093/humrep/13.12.3434. [DOI] [PubMed] [Google Scholar]
- 29.Eisen MB, Spellman PT, Brown PO, Botstein D. Cluster analysis and display of genome-wide expression patterns. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(25):14863–14868. doi: 10.1073/pnas.95.25.14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Assou S, Boumela I, Haouzi D, et al. Transcriptome analysis during human trophectoderm specification suggests new roles of metabolic and epigenetic genes. PLoS One. 2012;7(6) doi: 10.1371/journal.pone.0039306.e39306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dozier BL, Watanabe K, Duffy DM. Two pathways for prostaglandin F2α synthesis by the primate periovulatory follicle. Reproduction. 2008;136(1):53–63. doi: 10.1530/REP-07-0514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Regassa A, Rings F, Hoelker M, et al. Transcriptome dynamics and molecular cross-talk between bovine oocyte and its companion cumulus cells. BMC Genomics. 2011;12, article no. 57 doi: 10.1186/1471-2164-12-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Szöllösi D. On the role of gap junctions between follicle cells and oocyte in the mammalian ovarian follicle. Research Communications in Chemical Pathology and Pharmacology. 1978;10(2):3–4. [PubMed] [Google Scholar]
- 34.Dunning KR, Cashman K, Russell DL, Thompson JG, Norman RJ, Robker RL. Beta-oxidation is essential for mouse oocyte developmental competence and early embryo development. Biology of Reproduction. 2010;83(6):909–918. doi: 10.1095/biolreprod.110.084145. [DOI] [PubMed] [Google Scholar]
- 35.McEvoy TG, Coull GD, Broadbent PJ, Hutchinson JSM, Speake BK. Fatty acid composition of lipids in immature cattle, pig and sheep oocytes with intact zona pellucida. Journal of Reproduction and Fertility. 2000;118(1):163–170. [PubMed] [Google Scholar]
- 36.Russell DL, Robker RL. Molecular mechanisms of ovulation: co-ordination through the cumulus complex. Human Reproduction Update. 2007;13(3):289–312. doi: 10.1093/humupd/dml062. [DOI] [PubMed] [Google Scholar]
- 37.Fülöp C, Szántó S, Mukhopadhyay D, et al. Impaired cumulus mucification and female sterility in tumor necrosis factor-induced protein-6 deficient mice. Development. 2003;130(10):2253–2261. doi: 10.1242/dev.00422. [DOI] [PubMed] [Google Scholar]
- 38.Song HJ, Kang EJ, Maeng GH, et al. Influence of epidermal growth factor supplementation during in vitro maturation on nuclear status and gene expression of canine oocytes. Research in Veterinary Science. 2011;91(3):439–445. doi: 10.1016/j.rvsc.2010.09.003. [DOI] [PubMed] [Google Scholar]
- 39.Assidi M, Dieleman SJ, Sirard M-A. Cumulus cell gene expression following the LH surge in bovine preovulatory follicles: potential early markers of oocyte competence. Reproduction. 2010;140(6):835–852. doi: 10.1530/REP-10-0248. [DOI] [PubMed] [Google Scholar]
- 40.Shimada Y, Li Y-T, Li S-C. Effect of GM2 activator protein on the enzymatic hydrolysis of phospholipids and sphingomyelin. Journal of Lipid Research. 2003;44(2):342–348. doi: 10.1194/jlr.M200234-JLR200. [DOI] [PubMed] [Google Scholar]
- 41.Erickson GF, Garzo VG, Magoffin DA. Insulin-like growth factor-I regulates aromatase activity in human granulosa and granulosa luteal cells. Journal of Clinical Endocrinology and Metabolism. 1989;69(4):716–724. doi: 10.1210/jcem-69-4-716. [DOI] [PubMed] [Google Scholar]
- 42.Friedman CI, Seifer DB, Kennard EA, Arbogast L, Alak B, Danforth DR. Elevated level of follicular fluid vascular endothelial growth factor is a marker of diminished pregnancy potential. Fertility and Sterility. 1998;70(5):836–839. doi: 10.1016/s0015-0282(98)00301-x. [DOI] [PubMed] [Google Scholar]
- 43.Park J-Y, Su Y-Q, Ariga M, Law E, Jin S-LC, Conti M. EGF-like growth factors as mediators of LH action in the ovulatory follicle. Science. 2004;303(5658):682–684. doi: 10.1126/science.1092463. [DOI] [PubMed] [Google Scholar]
- 44.Ashkenazi H, Cao X, Motola S, Popliker M, Conti M, Tsafriri A. Epidermal growth factor family members: endogenous mediators of the ovulatory response. Endocrinology. 2005;146(1):77–84. doi: 10.1210/en.2004-0588. [DOI] [PubMed] [Google Scholar]
- 45.Ouandaogo ZG, Frydman N, Hesters L, et al. Differences in transcriptomic profiles of human cumulus cells isolated from oocytes at GV, MI and MII stages after in vivo and in vitro oocyte maturation. Human Reproduction. 2012;27:2438–2447. doi: 10.1093/humrep/des172. [DOI] [PubMed] [Google Scholar]
- 46.Chian R-C, Ao A, Clarke HJ, Tulandi T, Tan S-L. Production of steroids from human cumulus cells treated with different concentrations of gonadotropins during culture in vitro. Fertility and Sterility. 1999;71(1):61–66. doi: 10.1016/s0015-0282(98)00416-6. [DOI] [PubMed] [Google Scholar]
- 47.Kempná P, Körner M, Waser B, et al. Neuropeptide Y modulates steroid production of human adrenal H295R cells through Y1 receptors. Molecular and Cellular Endocrinology. 2010;314(1):101–109. doi: 10.1016/j.mce.2009.08.010. [DOI] [PubMed] [Google Scholar]
- 48.Barreca A, Valli B, Cesarone A, et al. Effects of the neuropeptide Y on estradiol and progesterone secretion by human granulosa cells in culture. Fertility and Sterility. 1998;70(2):320–325. doi: 10.1016/s0015-0282(98)00133-2. [DOI] [PubMed] [Google Scholar]
- 49.Körner M, Waser B, Reubi JC. Neuropeptide Y receptor expression in human primary ovarian neoplasms. Laboratory Investigation. 2004;84(1):71–80. doi: 10.1038/labinvest.3700009. [DOI] [PubMed] [Google Scholar]
- 50.Yi Y-J, Nagyova E, Manandhar G, et al. Proteolytic activity of the 26S proteasome is required for the meiotic resumption, germinal vesicle breakdown, and cumulus expansion of porcine cumulus-oocyte complexes matured in vitro. Biology of Reproduction. 2008;78(1):115–126. doi: 10.1095/biolreprod.107.061366. [DOI] [PubMed] [Google Scholar]
- 51.Cillo F, Brevini TAL, Antonini S, Paffoni A, Ragni G, Gandolfi F. Association between human oocyte developmental competence and expression levels of some cumulus genes. Reproduction. 2007;134(5):645–650. doi: 10.1530/REP-07-0182. [DOI] [PubMed] [Google Scholar]
- 52.Pangas SA, Jorgez CJ, Matzuk MM. Growth differentiation factor 9 regulates expression of the bone morphogenetic protein antagonist gremlin. Journal of Biological Chemistry. 2004;279(31):32281–32286. doi: 10.1074/jbc.M403212200. [DOI] [PubMed] [Google Scholar]
- 53.Golos TG, Strauss JF., III 8-Bromoadenosine cyclic 3′5′-phosphate rapidly increases 3-hydoxy-3-methylglutaryl coenzyme a reductase mRNA in human granulosa cells: role of cellular sterol balance in controlling the response to tropic stimulation. Biochemistry. 1988;27(9):3503–3506. doi: 10.1021/bi00409a056. [DOI] [PubMed] [Google Scholar]
- 54.Watanabe H, Hirai S, Tateno H, Fukui Y. Variation of cholesterol contents in porcine cumulus-oocyte complexes is a key factor in regulation of fertilizing capacity. Theriogenology. 2013;79:680–686. doi: 10.1016/j.theriogenology.2012.11.024. [DOI] [PubMed] [Google Scholar]
- 55.Zhang X, Jafari N, Barnes RB, Confino E, Milad M, Kazer RR. Studies of gene expression in human cumulus cells indicate pentraxin 3 as a possible marker for oocyte quality. Fertility and Sterility. 2005;83(4, supplement 1):1169–1179. doi: 10.1016/j.fertnstert.2004.11.030. [DOI] [PubMed] [Google Scholar]
- 56.Chakraborty A, Brooks H, Zhang P, et al. Stanniocalcin-1 regulates endothelial gene expression and modulates transendothelial migration of leukocytes. American Journal of Physiology—Renal Physiology. 2007;292(2):F895–F904. doi: 10.1152/ajprenal.00219.2006. [DOI] [PubMed] [Google Scholar]
- 57.Paciga M, McCudden CR, Londos C, DiMattia GE, Wagner GF. Targeting of big stanniocalcin and its receptor to lipid storage droplets of ovarian steroidogenic cells. The Journal of Biological Chemistry. 2003;278(49):49549–49554. doi: 10.1074/jbc.M307302200. [DOI] [PubMed] [Google Scholar]
- 58.Basini G, Bussolati S, Santini SE, et al. Antiangiogenesis in swine ovarian follicle: a potential role for 2-methoxyestradiol. Steroids. 2007;72(8):660–665. doi: 10.1016/j.steroids.2007.05.002. [DOI] [PubMed] [Google Scholar]
- 59.Basini G, Baioni L, Bussolati S, et al. Expression and localization of stanniocalcin 1 in swine ovary. General and Comparative Endocrinology. 2010;166(2):404–408. doi: 10.1016/j.ygcen.2009.12.013. [DOI] [PubMed] [Google Scholar]
- 60.Basini G, Bussolati S, Santini SE, et al. Hydroxyestrogens inhibit angiogenesis in swine ovarian follicles. Journal of Endocrinology. 2008;199(1):127–135. doi: 10.1677/JOE-08-0258. [DOI] [PubMed] [Google Scholar]
- 61.Lim J, Luderer U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biology of Reproduction. 2011;84(4):775–782. doi: 10.1095/biolreprod.110.088583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ziv Y, Bielopolski D, Galanty Y, et al. Chromatin relaxation in response to DNA double-strand breaks is modulated by a novel ATM-and KAP-1 dependent pathway. Nature Cell Biology. 2006;8(8):870–876. doi: 10.1038/ncb1446. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Material available online includes: (i) Baseline clinical characteristics, end-of-stimulation data and number of oocytes retrieved from each patient following controlled ovarian stimulation with HP-hMG or rFSH. (ii) Primer pairs used for validation of the array data by qRT-PCR. (iii) List of the 9,805 common genes expressed in both HP-hMG and rFSH CC samples. (iv) Validation by qRT-PCR of some of the genes that were differentially expressed in HP-hMG and rFSH CC samples. (v) Top-ranked networks evidenced in common HP-hMG/rFSH signature by Ingenuity Pathway software. (vi) Detailed list of the genes presented in the network. (vii) The major signaling pathways that occur in CCs following the gonadotropin (HP-hMG and rFSH) stimulation.