

Abstract
Expression of genes in the endoplasmic reticulum (ER) beyond its protein folding capacity activates signaling pathways that are collectively referred to as the Unfolded Protein Response (UPR). A major branch of the UPR pathway is mediated by IRE1, an ER-tethered endonuclease. Upon ER stress-induced activation, IRE1 splices the mRNA of XBP1, thereby generating an active isoform of this transcription factor. During normal Drosophila development, tissues with high protein secretory load show signs of IRE1/XBP1 activity indicative of inherent ER stress associated with those cell types. Here, we report that the XBP1 promoter activity itself is enhanced in secretory tissues of Drosophila, and it can be induced by excessive ER stress. Specifically, we developed a Drosophila XBP1 transcription reporter by placing dsRed under the control of the XBP1 intergenic sequence. DsRed expression in these xbp1p>dsRed transgenic flies showed patterns similar to that of xbp1 transcript distribution. In healthy developing flies, the reporter expression was highest in salivary glands and the intestine. In the adult, the male reproductive organs showed high levels of dsRed. These tissues are known to have high protein secretory load. Consistently, the xbp1p>dsRed reporter was induced by excessive ER stress caused by mutant Rhodopsin-1 overexpression. These results suggest that secretory cells suffer from inherent ER stress, and the xbp1p>dsRed flies provide a useful tool in studying the function and homeostasis of those cells.
Introduction
Eukaryotic cells express most of their membrane and secretory proteins in the endoplasmic reticulum (ER), where those proteins undergo folding before being trafficked to their ultimate destination. Expression of mutant or wild type proteins that exceed the protein folding capacity of this organelle can impose stress that disrupts many aspects of cellular function. In response, most eukaryotic cells are equipped with signaling pathways that can induce the expression of ER quality control genes, which are widely referred to as the Unfolded Protein Response (UPR) [1].
Excessive ER stress and defective UPR underlies a wide variety of diseases in humans [2], [3]. Similarly, a number of human disease models of Drosophila, including Retinitis Pigmentosa, Alzheimer’s disease and Amyotrophic Lateral Sclerosis are also associated with ER stress [4], [5], [6], [7], [8]. A UPR pathway that is conserved between yeast, C. elegans, Drosophila and mammals is mediated by IRE1 and XBP1. IRE1 is an ER-tethered protein with a luminal domain that can bind to misfolded peptides [9]. Upon recognizing misfolded proteins, IRE1 forms oligomers on the ER membrane and activates its cytoplasmic RNAse domain [10], [11]. A major target of IRE1 is the mRNA of the transcription factor xbp1 [12], [13], [14], [15]. IRE1 mediated cleavage, and a subsequent ligation reaction results in an unconventional mRNA splicing event that occurs in the cytoplasm, and results in the shift of the XBP1 reading frame, thereby generating an isoform of XBP1 that acts as an active transcription factor. Once activated, XBP1 induces the expression of a number of ER quality control genes, including chaperones that fold misfolded proteins, and those involved in the degradation of misfolded ER proteins [16], [17].
We had previously shown that xbp1 is an essential gene for Drosophila development [4]. That observation raised the possibility that there are physiological levels of ER stress associated with normal development. Supporting that idea, we recently reported an XBP1 mRNA splicing reporter that indicated signs of IRE1 activity in healthy tissues of Drosophila [18]. Specifically, those tissues with constitutive IRE1 activity included the salivary gland, intestine and the male reproductive system, which are known for their high protein secretory load. Here, we report the development of an xbp1 transcription reporter, which drives dsRed under the control of the xbp1 promoter. We find that this reporter is also expressed in cells with high protein secretory load and is further induced by experimentally imposed ER stress. These results suggest that secretory cells suffer from inherent ER stress, and cells respond by inducing the transcription of xbp1, in addition to the IRE1-mediated splicing of these mRNAs. These tools may further facilitate the study of secretory protein homeostasis and stress response in Drosophila.
Materials and Methods
Constructs
The 0.85 kb intergenic sequence between xbp1 and CG9406 was amplified through genomic PCR. Specifically, the amplified DNA began from the end of CG9406, up to the start codon of the xbp1 coding sequence. This DNA was subcloned into the pPelican plasmid with a dsRed reporter [19], which we refer to as xbp1p>dsRed.
Fly genetics
This study did not involve vertebrate animal models. The ire1f02170 obtained from the Bloomington Stock Center had background lethal mutations, which were cleaned up by outcrossing to the Ryoo lab w1118. To generate mosaic clones, ire1f02170 was recombined to the FRT82 chromosome. Maternal zygotic ire1f02170 −/− flies were generated by utilizing the ovoD1 based protocol [20]. Specifically, we crossed FRT82, ire1f02170 flies to hs-flp; FRT82, ovoD1 flies to generate ire1 −/− clones in the germ line of the progeny. The female progeny of this cross were crossed to ire1f02170, xbp1p>dsRed/TM6B/TM6B-GFP males. The maternal zygotic mutants were distinguished by the presence of xbp1p>dsRed and the absence of GFP. IRE1 activity was assessed by crossing the UAS-xbp1-EGFP HG line [18] to the tubulin>Gal4 driver.
Immunohistochemistry
anti-Rhodopsin-1 antibody (4C5) was from DSHB of Univ. of Iowa and was used at 1∶500. xbp1p>dsRed was visualized through natural red fluorescence. For validation purposes, we used anti-dsRed antibody from Clonetech. Rabbit anti-GFP was from Invitrogen.
Results
The XBP1 expression reporter is active in highly secretory tissues
In a previous study, we had shown that xbp1 is primarily expressed in tissues that are notable for their secretory function, including the embryonic salivary gland and the midgut [4] (also Figure 1). To better understand how this gene expression is controlled, we developed an xbp1 gene expression reporter. Sequence comparison of the xbp1 locus in a number of Drosophila species showed that the intergenic sequence between xbp1 and its adjacent gene, CG9406, is conserved, raising the likelihood that this region may encode essential regulatory DNA sequences (Figure 1A). Based on this, we placed a dsRed reporter under the control of the xbp1 upstream intergenic DNA. We found that the transgenic flies expressed dsRed in a pattern that was indistinguishable from that of xbp1 in situ hybridization (Figure 1B). In embryos, the salivary gland and the midgut showed prominent expression of the xbp1p>dsRed reporter (Figure 1C). Also in the larva, dsRed was prominently expressed in the salivary gland and the larval intestine (Figure 1D, E).
Figure 1. XBP1 regulatory sequence drives dsRed expression in developing secretory tissues.

(A) Comparison of the xbp1 locus sequence between D. melanogaster, D. simulans, D. yakuba and D. erecta. Sequence conservation is not only found in the coding sequence of genes (purple) and UTRs (light blue), but also in the intergenic region (red) that may encode enhancers and promoters. The bracket (xbp1p) indicates the intergenic sequence that was placed upstream of the dsRed reporter. (B) xbp1 in situ hybridization in embryos. (C-E) xbp1p>dsRed reporter expression in the embryo (C), the larval salivary gland (D) and the larval intestine (E). Images in (D, E) are composites of several frames taken with the 10x confocal microscope lens. There was no detectable xbp1p>dsRed expression in the larval brain and imaginal discs (D). Abbreviations: Salivary Gland (SG), Mid Gut (MG).
IRE1 activity is detected in and beyond the cells expressing the XBP1 transcription reporter
In a previous study, we had described another reporter suitable for detecting IRE1’s enzymatic activity. That reporter took advantage of the fact that IRE1 mediated XBP1 mRNA splicing results in a frame shift in translation. By fusing GFP C-terminal to the XBP1 coding sequence, we were able to have GFP express in frame only in cells that have active IRE1 [18]. When this splicing sensor was expressed ubiquitously through the tubulin-Gal4 driver, we were able to detect IRE1 activity in the secretory cells that express xbp1p>dsRed (Figure 2A) [18]. These observations suggest that xbp1p>dsRed marks many cell types with inherent IRE1/XBP1 pathway activity.
Figure 2. Co-labeling of xbp1p>dsRed with an IRE1-mediated XBP1 splicing reporter.

Shown is a composite image of multiple 10x confocal microscopy images of the 3rd instar larval brain, eye discs and salivary glands. Labeled in red is the xbp1p>dsRed reporter, and in green is the IRE1-mediated XBP1 mRNA splicing reporter. The latter (IRE1 reporter) was driven ubiquitously through the tubulin-Gal4 driver. (A) Double labeling of dsRed and GFP shows overlap in the salivary glands. On the other hand, the brain and the eye discs primarily show IRE1 activity without XBP1 expression. (B) A GFP only channel of the image shown in (A).
Intriguingly, we noticed that many cells in the brain show signs of IRE1 activity, but do not show xbp1p>dsRed expression (Figure 2). That observation suggests that there are cells with active IRE1, but not its downstream target XBP1.
The XBP1 reporter is highly expressed in the adult male reproductory organs
In adults, the xbp1p>dsRed reporter was expressed at high levels in the male abdomen, but not in females (Figure 3A, B). Upon dissection, we found prominent expression of the xbp1p>dsRed in the male reproductory organs within the abdomen. Specifically, dsRed fluorescence was found in the accessory glands and the ejaculatory duct, but not in the testis itself. Notably, the accessory gland and the ejaculatory duct are known for their high secretory activity (see Discussion).
Figure 3. xbp1p>dsRed expression in the male reproductive system.
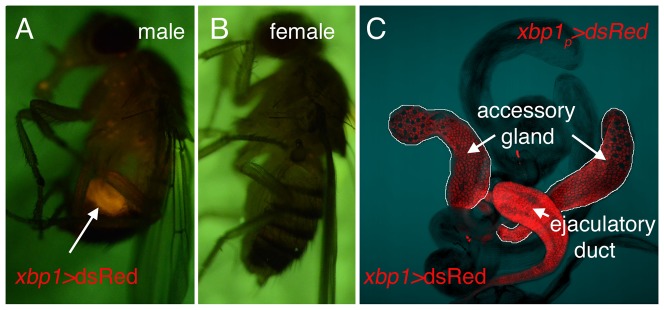
Prominent dsRed signal was found in the adult abdomen of males (A), but not in females (B). Dissected adult tissues show the strongest dsRed signal in the accessory gland and the ejaculatory duct (C).
The XBP1 promoter is activated by excessive ER stress
In a previous study, we had shown that the IRE1/XBP1 pathway can induce the ER chaperone, BiP, in imaginal discs [4]. However, we found no detectable expression of xbp1p>dsRed in that tissue (Figure 1D). Such inconsistencies led us to explore the possibility that xbp1p>dsRed is induced in these tissues primarily in response to ER stress. To impose ER stress, we expressed in larval eye discs a mutant membrane protein, Rhodopsin-1G69D, which fails to fold properly in the ER [4], [21]. Under these conditions, we found xbp1p>dsRed induction in that tissue (Figure 4A, B), supporting the idea that xbp1 promoter itself is induced in response to ER stress.
Figure 4. xbp1p>dsRed expression is induced by misfolded membrane proteins.

Shown are larval eye imaginal discs with xbp1p>dsRed (red). Control eye discs show no basal level of reporter activity (A). xbp1p>dsRed is induced by the overexpression of a mutant membrane protein, Rhodopsin-1G69D (green) (B), or the spliced xbp1 isoform xbp1-RB (D) in the eye using the GMR promoter. On the other hand, the unspliced xbp1 isoform, xbp1-RA, did not induce xbp1p>dsRed expression (C). (E) ire1 is not required for xbp1p>dsRed induction by mutant Rhodopsin-1G69D. ire1 mosaic clones were generated in the eye discs and marked by the absence of GFP. (E’, E’’) show higher magnification images of the inset in (E). GMR promoter driven Rhodopsin-1G69D (blue) induces xbp1p>dsRed in both ire1+ and ire1 −/− cells, although the intensity of dsRed is moderately reduced in ire1 −/− clones. (E’’) is a dsRed only channel of (E’).
Interestingly, we found that the IRE1/XBP1 pathway can induce xbp1p>dsRed. Specifically, we activated the IRE1/XBP1 pathway by overexpressing the spliced isoform of XBP1 (xbp1-RB)[22] in Drosophila eye discs, which led to the induction of xbp1p>dsRed (Figure 4C, D). Conversely, we inactivated the IRE1/XBP1 pathway by generating mosaic clones of ire1f02170 mutants. There is a single ire1 gene in Drosophila, unlike the presence of two IRE1 genes in humans. Drosophila ire1f02170 is a null allele with a PiggyBac transposon inserted within the coding sequence [23]. When mutant Rhodopsin-1G69D was expressed in the eye discs with ire1f02170 −/− mosaic clones, we still observed xbp1p>dsRed induction, albeit at a reduced level (Figure 4E-E’’). These results suggest that xbp1p>dsRed expression is controlled by the UPR, but there are pathways other than IRE1/XBP1 that play redundant roles in this process.
The XBP1 expression reporter is expressed in the ire1 −/− larva
We further examined the role of ire1 in xbp1p>dsRed expression of developing larvae. At 48 – 72 hours after egg laying, the ire1f02170 −/− larvae were significantly smaller than that of the ire1 −/+ siblings, as the homozygous mutant flies failed to develop beyond the 1st instar larval stage (Figure 5). Live ire1f02170 −/− larvae were no longer found after 72 hours of egg laying.
Figure 5. xbp1p>dsRed is expressed in the salivary gland of ire1 −/− larva.
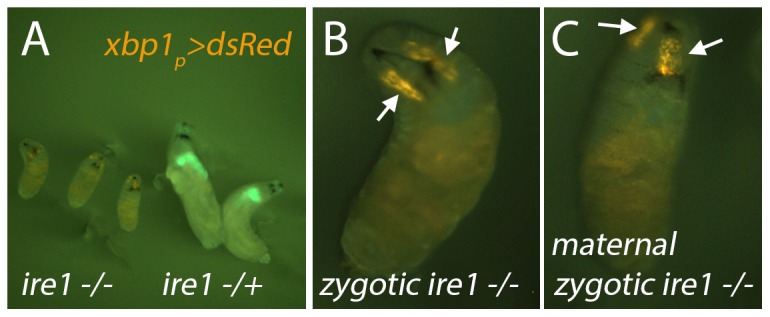
(A) Shown are 48 – 72 hour larvae of the ire1f02170 −/− (left three) and ire1 −/+ (right two) genotypes. xbp1p>dsRed was recombined to the ire1f02170 chromosome to facilitate genotyping. The ire1+ chromosome is marked with GFP. No ire1 −/− larvae were found to grow beyond this 1st instar larval stage. (B, C) A higher magnification view of an ire1 −/− larva. xbp1p>dsRed expression in the salivary gland is visible (arrows) in the ire1 −/− zygotic (B), as well as maternal, zygotic mutants (C).
As XBP1 is highly expressed in salivary glands, we examined whether the salivary glands form at all in the ire1f02170 −/− flies. To test this, we introduced the xbp1p>dsRed reporter into the background of ire1 mutants. The red fluorescence of dsRed was readily detected in these mutants (Figure 5B). To test whether there is a maternal ire1 mRNA contribution that allows the formation of the salivary gland, we generated maternal zygotic ire1 mutant flies, which also showed 1st instar larval lethality. These larvae still retained the xbp1p>dsRed in the salivary gland (Figure 5C), indicating that ire1 is neither required for the salivary gland formation nor the expression of xbp1p>dsRed in this tissue.
Discussion
Here, we report the development of an XBP1 expression reporter, which marked cells and tissues with high protein secretory load. Among the tissues with xbp1p>dsRed expression are salivary glands that secrete large amounts of digestive enzymes. Intestines of embryos and larvae were also prominent organs with xbp1p>dsRed expression. The intestine secretes a variety of proteins, not limited to those involved in digestion, but also in anti-microbial activity. In fact, studies in C. elegans, mice and humans indicate that xbp1 plays an essential role in maintaining the integrity of anti-microbial peptide secreting cells, and mutation of xbp1 in humans enhances risks for the inflammatory bowel syndrome [24], [25]. We also observed high xbp1p>dsRed expression in the male reproductive organs, such as the accessory gland and the ejaculatory duct. The Drosophila accessory gland is a tissue that produces a number of secretory proteins that are referred to as ACPs (Accessory Gland Proteins). There are now more than 50 ACPs that have been identified in Drosophila, indicating that the accessory gland is a major secretory tissue [26], [27]. Similarly, the ejaculatory duct also produces secretory proteins, including those that have anti-microbial activity [28]. These observations are consistent with the idea that XBP1 is expressed in cells with high secretory load. In addition, this promoter was induced by excessive ER stress. The latter property may be useful to researchers seeking facile methods to detect ER stress in live Drosophila tissues.
The pattern of the xbp1p>dsRed reporter largely overlapped with the XBP1 mRNA splicing reporter that we described in a recent publication (Figure 2) [18]. The tissues that showed signs of XBP1 mRNA splicing included the larval salivary gland, intestine and the adult accessory glands [18]. Intriguingly, we did not see xbp1p>dsRed expression in a few cell types that showed the XBP1 mRNA splicing reporter activity. The cell types of the latter class include a number of glial cells in the embryonic and larval brain (compare Figure 1D and [18]). It is noteworthy that the previously described XBP1 mRNA splicing reporter was expressed in all somatic cells using the beta tubulin promoter, even in those tissues with no detectable xbp1 transcripts. Thus, the splicing reporter did not reflect the expression pattern of xbp1, but rather, reported possible IRE1 activity in tissues. Taken together, the difference in the two reporter activities indicate that there are IRE1 active cells (such as certain glial cells) that do not have xbp1 mRNA. Recent studies indicate that the cytoplasmic RNase domain of IRE1 can cleave mRNAs other than that of xbp1, perhaps to reduce the load of new protein synthesis into the stressed ER [29], [30], [31], [32]. Strikingly, certain species like S. pombe do not encode an XBP1 ortholog, and IRE1’s main role in that species is to engage in the unconventional activity of degrading mRNAs [33]. Based on this, we speculate that the IRE1 activity in certain glial cells may be engaged in an XBP1-independent role for cellular homeostasis.
Acknowledgments
We thank the members of the Ryoo laboratory for comments on the manuscript. J. L. is a recipient of the Intel Science Competition Finalist Prize.
Funding Statement
This work was supported by the National Institutes of Health (NIH) grant R01 EY020866. The web address for NIH is http://www.nih.gov. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Walter P, Ron D (2011) The unfolded protein response: from stress pathway to hemeostatic regulation. Science 334: 1081–1086. [DOI] [PubMed] [Google Scholar]
- 2. Marciniak SJ, Ron D (2006) Endoplasmic reticulum stress signaling in disease. Physiol Rev 86: 1133–1149. [DOI] [PubMed] [Google Scholar]
- 3. Lin JH, Walter P, Yen TS (2008) Endoplasmic reticulum stress in disease pathogenesis. Annu Rev Pathol 3: 399–425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Ryoo HD, Domingos PM, Kang MJ, Steller H (2007) Unfolded protein response in a Drosophila model for retinal degeneration. Embo J 26: 242–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Tsuda H, Han SM, Yang Y, Tong C, Lin YQ, et al. (2008) The amyotriphic lateral sclerosis 8 protein VAPB is cleaved, secreted, and acts as a ligand for Eph receptors. Cell 133: 963–977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Mendes CS, Levet C, Chatelain G, Dourlen P, Fouillet A, et al. (2009) ER stress protects from retinal degeneration. EMBO J 28: 1296–1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Casas-Tinto S, Zhang Y, Sanchez-Garcia J, Gomez-Velazquez M, Rincon-Limas DE, et al. (2011) The ER stress factor XBP1s prevents amyloid-beta neurotoxicity. Hum Mol Genet 20: 2144–2160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Kumita JR, Helmfors L, Williams J, Luheshi LM, Menzer L, et al. (2012) Disease-related amyloidogenic variants of human lysozyme trigger the unfolded protein response and disturb eye development in Drosophila melanogaster. FASEB J 26: 192–202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Gardner BM, Walter P (2011) Unfolded proteins are Ire1-activating ligands that directly induce the unfolded protein response. Science 333: 1891–1894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Korennykh AV, Egea PF, Korostelev AA, Finer-Moore J, Zhang C, et al. (2009) The unfolded protein response signals trhough high-order assembly of Ire1. Nature 457: 687–693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Li H, Korenykh AV, Behrman SL, Walter P (2010) Mammalian endoplasmic reticulum stress sensor IRE1 signals by dynamic clustering. Proc Natl Acad Sci USA 107: 16113–16118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Sidrauski C, Walter P (1997) The transmembrane kinase Ire1p is a site-specific endonuclease that initiates mRNA splicing in the unfolded protein response. Cell 90: 1031–1039. [DOI] [PubMed] [Google Scholar]
- 13. Shen X, Ellis RE, Lee K, Liu CY, Yang K, et al. (2001) Complementary signaling pathways regulate the unfolded protein response and are required for C. elegans development. Cell 107: 893–903. [DOI] [PubMed] [Google Scholar]
- 14. Yoshida H, Okada T, Haze K, Yanagi H, Yura T, et al. (2001) Endoplasmic reticulum stress-induced formation of transcription factor complex ERSF including NF-Y (CBF) and activating transcription factors 6alpha and 6beta that activates the mammalian unfolded protein response. Mol Cell Biol 21: 1239–1248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Calfon M, Zeng H, Urano F, Till JH, Hubbard SR, et al. (2002) IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature 415: 92–96. [DOI] [PubMed] [Google Scholar]
- 16. Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, et al. (2000) Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 101: 249–258. [DOI] [PubMed] [Google Scholar]
- 17. Yamamoto K, Sato T, Matsui T, Sato M, Okada T, et al. (2007) Transcriptional induction of mammalian ER quality control proteins is mediated by single, or combined action of ATF6alpha and xbp1. Dev Cell 13: 365–376. [DOI] [PubMed] [Google Scholar]
- 18. Sone M, Zeng X, Larese J, Ryoo HD (2013) A modified UPR stress sensing system reveals a novel tissue distribution of IRE1/XBP1 activity during normal Drosophila development. Cell Stress Chaperones 18: 307–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Barolo S, Castro B, Posakony JW (2004) New Drosophila transgenic reporters: insulated P-element vectors expressing fast-maturing RFP. Biotechniques 36: 436–440. [DOI] [PubMed] [Google Scholar]
- 20. Chou TB, Perrimon N (1992) Use of a yeast site-specific recombinase to produce female germline chimeras in Drosophila. Genetics 131: 643–653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Kang MJ, Ryoo HD (2009) Suppression of retinal degeneration in Drosophila by stimulation of ER-Associated Degradation. Proc Natl Acad Sci USA 106: 17043–17048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Ryoo HD, Steller H (2007) Unfolded protein response in Drosophila: why another model can make it fly. Cell Cycle 6: 830–835. [DOI] [PubMed] [Google Scholar]
- 23. Kang MJ, Chung J, Ryoo HD (2012) CDK5 and MEKK1 mediate pro-apoptotic signalling following endoplasmic reticulum stress in an autosomal dominant retinitis pigmentosa model. Nat Cell Biol 14: 409–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kaser A, Lee AH, Franke A, Glickman JN, Zeissig S, et al. (2008) XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 134: 743–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Richardson CE, Kooistra T, Kim DH (2010) An essential role for XBP-1 in host protection against immune activation in C. elegans. Nature 463: 1092–1095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Wolfner MF (1997) Tokens of love: functions and regulation of Drosophila male accessory gland products. Insect Biochem Mol Biol 27: 179–192. [DOI] [PubMed] [Google Scholar]
- 27. Mueller JL, Ravi Ram K, McGraw LA, Bloch Qazi MC, Siggia ED, et al. (2005) Cross-species comparison of Drosophila male accessory gland protein genes. Genetics 171: 131–143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Lung O, Kuo L, Wolfner MF (2001) Drosophila males transfer antibacterial proteins from their accessory gland and ejaculatory duct to their mates. J Insect Physiol 47: 617–622. [DOI] [PubMed] [Google Scholar]
- 29. Hollien J, Weissman JS (2006) Decay of endoplasmic reticulum-localized mRNAs during the unfolded protein response. Science 313: 104–107. [DOI] [PubMed] [Google Scholar]
- 30. Hollien J, Lin JH, Li H, Stevens N, Walter P, Weissman JS (2009) Regulated Ire1-dependent decay of messenger RNAs in mammalian cells. J Cell Biol 186: 323–331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Han D, Lerner AG, Vande Walle L, Upton JP, Xu W, et al. (2009) IRE1alpha kinase activation modes control alternate endoribonuclease outputs to determine divergent cell fates. Cell 138: 562–572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Cross BC, Bond PJ, Sadowski PG, Jha BK, Zak J, et al. (2012) The molecular basis for selective inhibition of unconventional mRNA splicing by an IRE1-binding small molecule. Proc Natl Acad Sci USA 109: E869–878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Kimmig P, Diaz M, Zheng J, Williams CC, Lang A, et al. (2012) The unfolded protein response in fission yeast modulates stability of select mRNAs to maintain protein homeostasis. Elife 1: e00048. [DOI] [PMC free article] [PubMed] [Google Scholar]