

Abstract
The widely used insecticide beta-cypermethrin has become a public concern because of its environmental contamination and toxic effects on mammals. In this study, a novel beta-cypermethrin degrading enzyme designated as CMO was purified to apparent homogeneity from a Streptomyces sp. isolate capable of utilizing beta-cypermethrin as a growth substrate. The native enzyme showed a monomeric structure with a molecular mass of 41 kDa and pI of 5.4. The enzyme exhibited the maximal activity at pH 7.5 and 30°C. It was fairly stable in the pH range from 6.5–8.5 and at temperatures below 10°C. The enzyme activity was significantly stimulated by Fe2+, but strongly inhibited by Ag+, Al3+, and Cu2+. The enzyme catalyzed the degradation of beta-cypermethrin to form five products via hydroxylation and diaryl cleavage. A novel beta-cypermethrin detoxification pathway was proposed based on analysis of these products. The purified enzyme was identified as a monooxygenase by matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry analysis (MALDI-TOF-MS) and N-terminal protein sequencing. Given that all the characterized pyrethroid-degrading enzymes are the members of hydrolase family, CMO represents the first pyrethroid-degrading monooxygenase identified from environmental microorganisms. Taken together, our findings depict a novel pyrethroid degradation mechanism and indicate that the purified enzyme may be a promising candidate for detoxification of beta-cypermethrin and environmental protection.
Introduction
Pyrethroid insecticides have been used worldwide due to their potent toxic activity against various insect pests, and in particular, they have become the dominate insecticides in retail markets [1], [2]. Since 2000, the usage of these pesticides has been increased by as much as 25%, and their application is anticipated to be further increased due to the reduced use of organophosphate insecticides like diazinon and chlorpyrifos [3]. Beta-cypermethrin (beta-CP) [cyano-(3-phenoxyphenyl) methyl 3-(2,2-dichloroethenyl)-2,2-dimethylcyclopropane-1-carboxylate] is one of the most frequently used pyrethroid insecticides. It has been widely used in agriculture, forestry, horticulture, public health, and homes, as well as for protection of textiles and buildings [4], [5].
Persistent and large scale use of beta-CP has resulted in a serious environmental contamination problem, and which poses a serious threat to the health of human beings and ecosystems [6]–[10]. For instance, the pesticide has been detected in nearly all the urban creeks in California [3], [11]–[13]. Beta-CP is highly toxic to fish and aquatic invertebrates [8], [14]–[16]. Moreover, it shows carcinogenic and endocrine disrupting activities on mammals [6], [17]–[19]. Furthermore, the pesticide is known to adversely affect reproductive function, and the development of sexual and nervous systems [20]–[24]. The findings press the critical need for developing effective and economic approaches to remove this contaminate from environments.
Biodegradation has been attracting much attention in cleanup of the contaminated environments because conventional physical and chemical methods for disposal of persistent pollutants are low in efficiency and need comparatively high operating cost [25]–[30]. Several beta-CP degradation mechanisms have been identified in recent years, such as beta-CP degrading bacterial isolates Serratia sp. JCN13 [31]; Ochrobactrum lupini DG-S-01 [32], and Pseudomonas aeruginosa CH7 [5], and the three genes, i.e., Estp, pytH, and PytZ, encoding pyrethroid-degrading hydrolases from Klebsiella sp. ZD112, Sphingobium sp. JZ-1, and Ochrobactrum anthropi YZ-1, respectively [33]–[35]. In addition, one thermostable enzyme Sys410 involved in pyrethroid degradation has recently been isolated from Tuban Basin soil using metagenomic approach [36]. However, little information is available regarding the ability of actinomycetes in degradation of beta-CP. In this study, we described the purification and characterization of a novel beta-CP degrading enzyme from the actinomycete Streptomyces sp., previously isolated from the pyrethroid-contaminated soils [4]. The purpose of this study was to investigate its specific role in beta-CP degradation. To the best of our knowledge, this is the first pyrethroid-degrading enzyme purified to homogeneity from actinomycetes.
Materials and Methods
Chemicals
Beta-CP (94.8% purity) was obtained from Zhongshan Aestar Fine Chemical Inc., Ltd, China. Sephacryl™ S-100 (16/60) and diethylaminoethyl cellulose (DEAE) were purchased from General Electric Company, USA. Chromatographic grade acetonitrile were purchased from Sigma-Aldrich, USA. Sodium dodecyl sulfate (SDS) and polyacrylamide were purchased from Amresco, USA. All the other chemicals and solvents used in this study were at analytical grade.
Microorganisms and Culture Conditions
The mineral salt medium (MSM) containing (g·L−1) (NH4)2SO4, 2; MgSO4·7 H2O, 0.2; CaCl2·2 H2O, 0.01; FeSO4·7 H2O, 0.001, Na2HPO4·12 H2O, 1.5; and KH2PO4, 1.5 was used for the cultivation of pyrethroid-degrading strains. The actinomycete isolate, which was used in this study, was isolated from the pyrethroid-contaminated soils using an enrichment culture technique [4].
For enzyme production, fresh MSM medium supplemented with 50 mg·L−1 of beta-CP was inoculated with Streptomyces sp. viable spores. The culture was incubated at 28°C for 5 days in 500 mL-Erlenmeyer flasks containing 200 mL of medium on a rotary shaker at 150 rpm, harvested by centrifugation at 8000×g for 30 min at 4°C, washed twice with cold phosphate buffer solution (PBS, 50 mM, pH 7.5), and stored at −20°C until further use.
Preparation of Crude Cell Extracts
To prepare crude cell extracts, the harvested mycelia were re-suspended in 50 mM PBS buffer (pH 7.5) to a final concentration of 25 g cell dry weight per litre. The mycelia cells were disrupted in an ultrasonic cell disruption system at 4°C. The crude cell extracts were obtained after removing cellular debris by centrifugation at 12,000×g for 30 min at 4°C. The resulting supernatants were used as an enzyme source for further purification. Protein concentration was determined by the method of Bradford [37] with bovine serum albumin as a standard.
Enzyme Assays
Pesticide catalytic activity was assayed by adding 0.05 mL enzyme solution to 5 mL of 50 mM PBS (pH 7.5) containing 5 mg·L−1 of beta-CP and incubating for 10 min at 30°C. After that, samples were extracted as described below, and the residues of pesticide were determined by high performance liquid chromatography (HPLC) according to the method described by Chen et al. [38]. Every treatment was performed in triplicate with inactivated enzyme as control. One unit of enzyme activity (U) was defined as the amount required to catalyze formation of 1 µmol of product per minute.
Enzyme Purification
All of the experiments below were carried out at 4°C unless otherwise specified. Purification was performed according to the methods described in previous reports [39]–[41] with minor modifications.
Ammonium sulfate precipitation. The crude cell extracts was brought to 55% ammonium sulfate saturation and stirred for 30 min, the cloudy suspension was centrifuged at 12,000×g for 20 min, and the supernatant was brought to 60% ammonium sulfate saturation; the pellet obtained by centrifugation was dissolved in the smallest possible volume of 50 mM PBS (pH 7.5) and dialyzed 1000-fold against 50 mM PBS (pH 7.5), and the supernatant was finally concentrated to 2 mL.
DEAE-Sepharose ion exchange chromatography. The dialyzed enzyme solution after ammonium sulfate saturation was loaded onto DEAE-Sepharose Fast Flow anion-exchange column (2.5×30 cm) that had been pre-equilibrated with 50 mM PBS (pH 7.5). The column was washed with 20 mL of the same buffer, and proteins were eluted with a linear gradient of NaCl solution in the range of 0 to 1.0 M in the equilibrating buffer. Fractions (5 mL) were collected every 5 min and screened for enzyme activity. Active fractions were pooled for further purification.
Sephacryl™ S-100 (16/60) gel filtration. The concentrated enzyme solution after DEAE-Sepharose Fast Flow anion-exchange was further purified by a Sephacryl™ S-100 column (2.5×100 cm). The column was washed at a flow rate of 0.5 mL·min−1 with 180 mL of 50 mM PBS buffer (pH 7.5), and 5-mL fractions were collected. The fractions with high specific activity were then pooled and concentrated for subsequent studies.
Determination of Molecular Mass and pI
The molecular mass of the denatured protein was determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). An SDS-12.5% polyacrylamide gel was prepared according to the method of Laemmli [42]. Proteins were stained with Coomassie brilliant blue R-250. The molecular mass of the native protein was estimated by gel filtration on the Sephacryl™ S-100 (16/60). Ovalbumin (43.0 kDa), carbonic anhydrase (31.0 kDa), and lysozyme (14.4 kDa) were used as the standard proteins. Isoelectric point (pI) was determined by isoelectric focusing calibration [39]–[41].
Effects of Temperature and pH on Activity and Stability of the Enzyme
To determine the optimal temperature and pH, the enzyme catalytic activity was investigated by incubating enzyme (0.05 mL) with 5 mg·L−1 beta-CP as a substrate for 10 min in 50 mM PBS. At pH 7.5, catalytic activity was determined in various temperatures ranging from 18°C to 40°C. At temperature 30°C, catalytic activity was assayed at pH values ranging from 4.0 to 11.0 [34], [41]. The relative residual activity was measured immediately as described previously. Here, the relative catalytic activity of the pre-incubated sample at 30°C was regarded as 100%.
Thermostability was measured by pre-incubation of the enzyme for different times in 50 mM PBS (pH 7.5) at different temperatures. The pH stability was tested after incubation of the enzyme for 2 h at 30°C [36]. The retaining catalytic activity of the enzyme was determined as mentioned above.
Effects of Different Metal Ions on Enzyme Catalytic Activity
For determination of the effects of different metal ions on catalytic activity, enzyme assay was performed in 50 mM PBS (pH 7.5) with 5 mg·L−1 beta-CP as a substrate and with various metal ions at a final concentration of 10 mM. The activity assayed in the absence of metal ions was defined as control [35]. The metal ions tested including Fe2+, Mg2+, Ca2+, K+, Na+, Zn2+, Ba2+, Mn2+, Cu2+, Al3+, and Ag+.
NH2-terminal Amino Acid Sequencing
The purified enzyme was electrophoretically transferred from a 12.5% SDS-PAGE to a polyvinylidene fluoride (PDVF) membrane (Millipore, USA). The region containing the beta-CP catalysis band was cut out, and the protein was sent to Shanghai GeneCore BioTechnologies Co., Ltd., China, for NH2-terminal amino acid sequencing [40], [41]. Sequencing was performed by matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometer (MALDI-TOF-MS; Bremen, Germany) according to the method of Peng et al. [43].
Identification of the Beta-CP Transformation Products
The beta-CP transformation products in the reaction solution were collected at different catalytic times. After acidification to pH 2 with 2 M HCl, the reaction solution was extracted according to the method described in a previous report [44]. The products identified by mass spectrometry analysis were matched with authentic standard compounds from the National Institute of Standards and Technology (NIST, USA) library database.
The transformation products of beta-CP by the enzyme were identified on an Agilent 6890N/5975 gas chromatopraphy-mass spectrometry (GC-MS) system equipped with auto-sampler, an on-column, split/splitless capillary injection system, and with HP-5MS capillary column (30.0 m×250 µm×0.25 µm) with array detection from 30–500 nm (total scan). Helium was used as a carrier gas at a flow rate of 1.5 mL·min−1. The column was held initially at a temperature of 90°C for 2 min, then at 6°C·min−1 to 150°C for 1 min, at 10°C·min−1 to 180°C for 4 min, and finally at 20°C·min−1 to 260°C for 10 min. The temperatures corresponding to transfer line and the ion trap were 280°C and 230°C, respectively. The column outlet was inserted directly into the electron ionization source block, operating at 70 eV. The injection volume was 1.0 µL with splitless sampling at 250°C [45], [46].
Results
Production of the Enzyme
Streptomyces sp. was aerobically cultured in the MSM medium supplemented with beta-CP as a growth substrate. Beta-CP degrading enzyme activity was detected at the late log phase and reached the maximum level after 5 days of cultivation (Fig. S1). However, when the pesticide in the MSM medium was replaced by an equal amount of sucrose or glucose, the crude cell extracts showed no or only a trace level of the enzyme activity. These results suggest that the enzyme was induced by beta-CP in the medium. This observation is reminiscent of a previous finding that the pyrethroid hydrolase from Aspergillus niger ZD11 was inducible by trans-permethrin [39].
Purification and the Molecular Mass of the Enzyme
The beta-CP degrading enzyme was purified from the crude cell extracts of Streptomyces sp. by ammonium sulfate precipitation, DEAE-Sepharose Fast Flow anion-exchange chromatography, and Sephacryl™ S-100 gel filtration chromatography. The results of purification were summarized in Table 1. The enzyme was purified by 26.1-fold to a specific activity of 21.4 U·mg−1 protein from the cells with a yield of 21.2%. The purified enzyme gave a single band in SDS-PAGE, suggesting the purified sample was electrophoretically homogeneous under the dissociating conditions. The molecular mass of the purified enzyme determined by SDS-PAGE analysis was approximately 41 kDa (Fig. 1). SDS-PAGE analysis of the fractions showing beta-CP degrading enzyme activity obtained during the purification was given in Fig. S2. The relative molecular mass of native enzyme estimated by gel filtration on a calibrated column of Sephacryl™ S-100 was around 41 kDa. These results suggest that the purified enzyme is a monomer. The pI value was determined to be 5.4. Herewith, we designated the purified enzyme as CMO for the convenience of description.
Table 1. Purification of beta-CP degrading enzyme from Streptomyces sp.
| Purification steps | Total protein (mg) | Total activity (U)a | Specific activity (U·mg−1) | Yield (%) | Purification fold |
| Crude cell extracts | 1702.4 | 1674.8 | 1.0 | 100 | 1.0 |
| Ammonium sulfate precipitation | 108.6 | 599.0 | 5.5 | 35.8 | 5.6 |
| DEAE-Sepharose Fast Flowanion-exchange chromatography | 36.3 | 382.8 | 10.5 | 22.9 | 10.7 |
| Sephacryl™ S-100 gel filtration | 16.6 | 355.7 | 21.4 | 21.2 | 26.1 |
Note: aOne unit of enzyme activity (U) was defined as the amount required to catalyze formation of 1 µmol of product per minute.
Figure 1. SDS-PAGE analysis of the purified enzyme from Streptomyces sp. (Lane 2 and 3) and protein markers (Lane 1) stained with Coomassie blue.
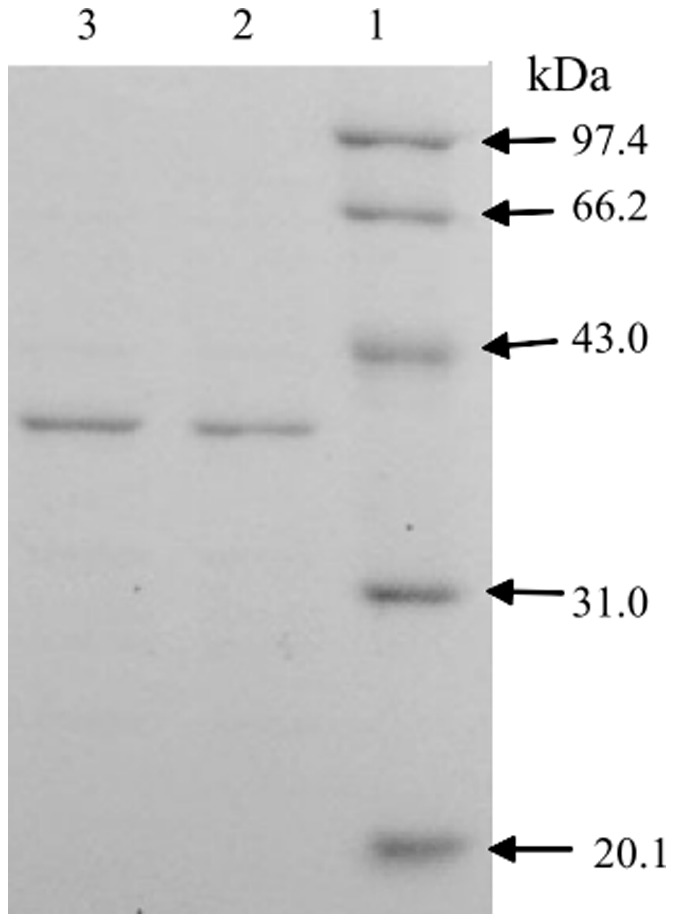
Markers from top to bottom are phosphorylase b (97.4 kDa), bovine serum albumin (66.2 kDa), ovalbumin (43.0 kDa), carbonic anhydrase (31.0 kDa), and trypsin inhibitor (20.1 kDa).
Effect of Temperature on CMO Activity and Stability
The effect of temperature on CMO activity was investigated by using beta-CP as a substrate at pH 7.5, with temperature ranging from 18°C to 40°C. Catalytic activity was increased as temperature increased up to 30°C, and decreased beyond that level. CMO displayed a high catalytic activity at temperatures between 25°C and 35°C with an optimal temperature at 30°C (Fig. 2a).
Figure 2. Effect of temperature on enzyme activity (a) and stability (b) of CMO.

The purified enzyme was pre-incubated at 0°C (♦), 10°C (▪), 20°C (▴), and 30°C (•) for 6 h. Error bar represents the standard deviation of the mean of three replicates.
Thermostability of the purified enzyme was determined by analysis of residual activity at regular intervals after pre-incubation for 6 h, at temperatures ranging from 0 to 30°C. CMO was highly stable below 10°C, with a residual activity exceeding 85% after incubation for 6 h. At temperatures above 10°C, catalytic activity was decreased slowly along with increased incubation time. The enzyme retained approximately 60% and 50% of its activity after incubation for 6 h at 20°C and 30°C, respectively (Fig. 2b).
Effect of pH on CMO Activity and Stability
The effect of pH on CMO activity was determined using beta-CP as a substrate at 30°C, with pH values ranging from 4.0 to 11.0. Maximal activity was observed at pH 7.5. The enzyme showed high activity in a broad pH range with 80% of residual activity between pH values 6.5 and 8.5 (Fig. 3a).
Figure 3. Effect of pH on enzyme activity (a) and stability (b) of CMO.

Error bar represents the standard deviation of the mean of three replicates.
pH stability was measured via 2 h pre-incubation of the purified enzyme in the above buffer at different pH values ranging from 4.0 to 11.0 at 30°C. The results in Fig. 3b show that the enzyme was fairly stable at pH values between 6.5 to 8.5, retaining more than 80% of the original activity after pre-incubation for 2 h. However, at pH values of 4.0 and 11.0, CMO only kept about 20% and 40% of its activity after incubation for 2 h, respectively. These results suggest that the purified enzyme is more sensitive to acidic conditions than alkaline conditions.
Effect of Metal Ions on CMO Activity
The influence of various metal ions on the enzyme activity of CMO was investigated by addition of metal ions separately into the reaction mixture at a final concentration of 1.0 mM. The CMO activity was then measured with beta-CP as a substrate and expressed as a percentage of the activity obtained in the absence of the added compounds (Table 2). Ag+, Al3+, and Cu2+ had a strong inhibitory effect (40–50% inhibition); while Zn2+, Mn2+, and Ba2+ showed only slight inhibitory effect (5–10% inhibition), respectively. Other metal ions such as Na+ and K+ had no obvious effect on the enzyme activity. In contrast, the presence of Ca2+ and Mg2+ resulted in slight stimulation (10–20% stimulation) of the enzymatic activity. In particular, the CMO activity was drastically boosted by Fe2+ up to 317.2%, suggesting that Fe2+ may be associated with the catalytic center of the enzyme.
Table 2. Effects of various metal ions on beta-CP degrading enzyme activity.
| Metal ions | Relative activity (%) | Metal ions | Relative activity (%) |
| Fe2+ | 317.2±16.3a | Ba2+ | 98.9±14.7b |
| Mg2+ | 109.7±13.9b | Mn2+ | 94.8±8.4b |
| Ca2+ | 117.4±11.0b | Cu2+ | 62.2±9.2c |
| K+ | 106.3±14.9b | Al3+ | 58.6±7.4c |
| Na+ | 101.1±8.6b | Ag+ | 51.5±10.0c |
| Zn2+ | 89.4±10.6b |
Note: The data presented are means of three replicates with standard deviation, which is within 5% of the mean.
Protein Identification
The purified protein was identified by matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry analysis (MALDI-TOF-MS) and N-terminal protein sequencing (Fig. 4). Database searches showed that the protein matched well with the monooxygenase from Streptomyces genus.
Figure 4. Matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry analysis (MALDI-TOF-MS) of the purified enzyme.

Identification of the Beta-CP Transformation Products
The enzyme catalyzed the degradation of beta-CP to five products in the PBS buffer, including 2-(4-hydroxyphenoxy) benzoic acid methyl ester, 3,5-dihydroxy benzoic acid, 3,5-dimethoxy phenol, 4-hydroxy-3-methoxy benzoic acid, and phenol. The transformation products were identified and confirmed by GC-MS, based on the similarity of their fragment and molecular ions with those of corresponding authentic compounds (Fig. S3). The retention time (RT) of these compounds was presented in Table 3. On the basis of the transformation products formed, a novel detoxification pathway for beta-CP was proposed (Fig. 5). The monooxygenase first converted the parent beta-CP [A] to 2-(4-hydroxyphenoxy) benzoic acid methyl ester [B] via hydroxylation. The intermediate was then subjected to diaryl cleavage, resulting in formation of 3,5-dihydroxy benzoic acid [C], 3,5-dimethoxy phenol [D], 4-hydroxy-3-methoxy benzoic acid [E], and phenol [F]. This is the first evidence of a novel beta-CP detoxification mechanism by hydroxylation and diaryl cleavage, which we propose is of vital importance in the beta-CP biogeocycle.
Table 3. Chromatographic properties of the beta-CP degradation products by the enzyme from Streptomyces sp.
| Code | RT (min) | m/z | Compounds |
| A1 | 31.145 | 415 | trans-Beta-CP |
| A2 | 30.961 | 459 | cis-Beta-CP |
| B | 15.903 | 497 | 2-(4-Hydroxyphenoxy) benzoic acid methyl ester |
| C | 9.650 | 429 | 3,5-Dihydroxy benzoic acid |
| D | 9.576 | 460 | 3,5-Dimethoxy phenol |
| E | 8.046 | 232 | 4-Hydroxy-3-methoxy benzoic acid |
| F | 3.653 | 281 | Phenol |
Figure 5. The proposed pathway for detoxification of beta-CP by the enzyme from Streptomyces sp. I, hydroxylation; II, diaryl cleavage.

Discussion
In this study, a novel pyrethroid-degrading enzyme designated as CMO was purified to apparent homogeneity from a Streptomyces sp. isolate capable of utilizing beta-CP as a growth substrate. The pyrethroid-degrading enzyme activities have been reported from several bacterial and fungal isolates [33], [34], [35], [39], [40], [47], but not from actinomycetes. This is the first report to our knowledge on the production, purification, and characterization of a pyrethroid-degrading enzyme from actinomycetes.
CMO has an apparent molecular mass of 41 kDa, which is smaller than the reported pyrethroid-hydrolyzing enzymes, such as permethrinase (61 kDa) from Bacillus cereus SM3 [47], pyrethroid hydrolase (56 kDa) from A. niger ZD11 [39], and EstP (73 kDa) from Klebsiella sp. ZD112 [33]. However, it was bigger than esterase (31.2 kDa) from the metagenome [48], pytH (31 kDa) from Sphingobium sp. JZ-1 [34], PytZ (24.2 kDa) from Ochrobactrum anthropi YZ-1 [35], and Sys410 (30.8 kDa) from the metagenome [36]. The pI value of CMO was estimated to be 5.4, which was similar to the value of the pyrethroid hydrolase from A. niger ZD11 (5.4), but lower than that recorded for Klebsiella sp. ZD112 (8.6) [33], [39]. The low pI suggests that the enzyme is rich in acidic amino acid residues [40], [41]. Taken together, the above findings suggest that CMO could be a novel pyrethroid-degradation enzyme.
CMO displayed a high catalytic activity at temperatures between 25°C and 35°C with an optimal temperature at 30°C. This result was in consistent with the observation that the optimal temperature for pyrethroid-degrading enzymes is usually between 30°C and 45°C [33], [35], [39], [40], [47], [48]. However, it was far less than that recorded for the thermostable pyrethroid-hydrolyzing enzyme Sys410 (55°C) [36].
CMO showed high activity in a broad pH range between pH values 6.5 and 8.5. Maximal activity was observed at pH 7.5. A range of previous characterized pyrethroid-degrading enzymes show maximal activity at pH 7.5 [35], [40], [47], but the pyrethroid esterase from various bacterial isolates or metagenome are known to exhibit maximal activity at pH range from 6.5–7.0 [33], [36], [39], [48]. Similar results were observed in most other pyrethroid-degrading enzymes [33], [34], [35], [39], [40]. Notably, CMO retained a high activity at a broad range of temperatures and pH, suggesting the promising potential of using this enzyme to clean up beta-CP contamination under various environmental conditions.
The influence of various metal ions on the enzyme activity of CMO was investigated. The enzyme activity was significantly stimulated by Fe2+. This result contrasts with previous findings of Zhai et al. [35] who reported that Fe2+ had no significant effect on the pyrethroid-hydrolyzing carboxylesterase (PytZ) isolated from O. anthropi YZ-1. Our results suggested that Fe2+ may be involved in the catalytic center of the enzyme. However, the CMO activity was strongly inhibited by Ag+, Al3+, and Cu2+. Similar results were observed in other pyrethroid-degrading enzymes isolated from A. niger ZD11, Klebsiella sp. ZD112, Sphingobium sp. JZ-2, and O. anthropi YZ-1 [33], [35], [39], [40].
The purified enzyme was identified as a monooxygenase by matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry analysis (MALDI-TOF-MS) and N-terminal protein sequencing. Until now, all the reported pyrethroid-degrading enzymes from environmental isolates belong to hydrolase family [33], [34], [35], [39], [40], [47], whereas our results suggest that the purified enzyme is most likely to be a monooxygenase.
Generally, carboxylic ester hydrolysis by carboxylesterases is considered as the first step of degradation and detoxification of pyrethroid insecticides in many species, from mammals and insects to bacteria [5], [26], [49], [50]. In this study, the transient accumulation of intermediate 2-(4-hydroxyphenoxy) benzoic acid methyl ester suggests that transformation of beta-CP by CMO was initiated by hydroxylation (Fig. 5, step I), which is differing from other pyrethroid hydrolases involved in pyrethroid degradation through hydrolysis [33], [34], [35], [39], [40]. Moreover, the enzyme further transformed the intermediate 2-(4-hydroxyphenoxy) benzoic acid methyl ester by cleavage of the diaryl bond (Fig. 5, step II), leading to detoxification of beta-CP. This is a sharp contrast to the reported pyrethroid hydrolases, which are known to undertake one-step reaction only (hydrolysis) [39], [40]. Given that the CMO enzyme is capable of complete degradation of beta-CP to generate simple aromatic compounds, the enzyme may be an ideal candidate for bioremediation of the beta-CP contaminated environments.
In conclusion, a novel beta-CP degrading enzyme was successfully purified from Streptomyces sp. in the present study. The strong catalytic activity and broad temperature and pH range of the enzyme were necessary to meet the practical requirements of bioremediation to enable its use in situ for detoxification of beta-CP. Moreover, we presented evidence that the enzyme harbors the catalytic mechanisms for complete degradation of beta-CP to yield simple aromatic products. To our knowledge, this is the first report that a pyrethroid-degrading enzyme transforms beta-CP through hydroxylation and diaryl cleavage, which we propose is of vital importance in beta-CP biogeocycle. Finally, this is the first pyrethroid-degrading monooxygenase purified to homogeneity from environmental isolates.
Supporting Information
The crude enzyme activity on degrading beta-CP over the incubation time.
(TIF)
SDS-PAGE analysis of the fractions showing beta-CP degrading enzyme activity obtained during the purification. Lane M: protein markers, 14.4–94.0 kDa; Lane 1: total proteins; Lane 2: fractions from ammonium sulfate precipitation; Lane 3: fractions from DEAE Sepharose Fast Flow anion-exchange.
(TIF)
GC-MS spectra of products produced from beta-CP degradation by CMO. a, trans-Beta-CP; b, cis-Beta-CP; c, 2-(4-Hydroxyphenoxy) benzoic acid methyl ester; d, 3,5-Dihydroxy benzoic acid; e, 3,5-Dimethoxy phenol; f, 4-Hydroxy-3-methoxy benzoic acid; g, Phenol.
(TIF)
Funding Statement
The authors are grateful to the National Natural Science Foundation (No.30871660) of China for the financial support. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Zhuang R, Chen H, Yao J, Li Z, Burnet JE, et al. (2011) Impact of beta-cypermethrin on soil microbial community associated with its bioavailability: a combined study by isothermal microcalorimetry and enzyme assay techniques. J Hazard Mater 189: 323–328. [DOI] [PubMed] [Google Scholar]
- 2. Weston DP, Holmes RW, Lydy MJ (2009) Residential runoff as a source of pyrethroid pesticides to urban creeks. Environ Pollut 157: 287–294. [DOI] [PubMed] [Google Scholar]
- 3. Cao Z, Shafer TJ, Crofton KM, Gennings C, Murray TF (2011) Additivity of pyrethroid actions on sodium influx in cerebrocortical neurons in primary culture. Environ Health Perspect 119: 1239–1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lin QS, Chen SH, Hu MY, Rizwan-ul-Haq M, Yang L, et al. (2011) Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int J Environ Sci Tech 8: 45–56. [Google Scholar]
- 5. Zhang C, Wang SH, Yan YC (2011) Isomerization and biodegradation of beta-cypermethrin by Pseudomonas aeruginosa CH7 with biosurfactant production. Bioresour Technol 102: 7139–7146. [DOI] [PubMed] [Google Scholar]
- 6. Corcellas C, Feo ML, Torres JP, Malm O, Ocampo-Duque W, et al. (2012) Pyrethroids in human breast milk: occurrence and nursing daily intake estimation. Environ Int 15: 17–22. [DOI] [PubMed] [Google Scholar]
- 7. Kuivila KM, Hladik ML, Ingersoll CG, Kemble NE, Moran PW, et al. (2012) Occurrence and potential sources of pyrethroid insecticides in stream sediments from seven U.S. metropolitan areas. Environ Sci Technol 46: 4297–4303. [DOI] [PubMed] [Google Scholar]
- 8. Kumar A, Sharma B, Pandey RS (2012) Assessment of stress in effect to pyrethroid insecticides, λ-cyhalothrin and cypermethrin, in a freshwater fish, Channa punctatus (Bloch). Cell Mol Biol (Noisy-le-grand) 22: 153–159. [PubMed] [Google Scholar]
- 9. Lu C, Adamkiewicz G, Attfield K, Kapp M, Spengler JD, et al. (2013) Household pesticide contamination from indoor pest control applications in urban low-income public housing dwellings: a community-based participatory research. Environ Sci Technol 47: 2018–2025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Morgan MK (2012) Children’s exposures to pyrethroid insecticides at home: a review of data collected in published exposure measurement studies conducted in the United States. Int J Environ Res Public Health 9: 2964–2985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Lubick N (2008) Pyrethroids are ubiquitous in California’s urban streams. Environ Sci Technol 12: 8622–8622. [DOI] [PubMed] [Google Scholar]
- 12. Weston DP, Lydy MJ (2010) Urban and agricultural sources of pyrethroid insecticides to the Sacramento-San Joaquin Delta of California. Environ Sci Technol 44: 1833–1840. [DOI] [PubMed] [Google Scholar]
- 13. Delgado-Moreno L, Lin K, Veiga-Nascimento R, Gan J (2011) Occurrence and toxicity of three classes of insecticides in water and sediment in two Southern California coastal watersheds. J Agric Food Chem 59: 9448–9456. [DOI] [PubMed] [Google Scholar]
- 14. Jin Y, Zheng S, Fu Z (2011) Embryonic exposure to cypermethrin induces apoptosis and immunotoxicity in zebrafish (Danio rerio). Fish Shellfish Immunol 30: 1049–1054. [DOI] [PubMed] [Google Scholar]
- 15. Ansari AR, Rahman S, Kaur M, Anjum S, Raisuddin S (2011) In vivo cytogenetic and oxidative stress-inducing effects of cypermethrin in freshwater fish, Channa punctata Bloch. Ecotoxicol Environ Saf 74: 150–156. [DOI] [PubMed] [Google Scholar]
- 16. Jin Y, Zheng S, Pu Y, Shu L, Sun L, et al. (2011) Cypermethrin has the potential to induce hepatic oxidative stress, DNA damage and apoptosis in adult zebrafish (Danio rerio). Chemosphere 82: 398–404. [DOI] [PubMed] [Google Scholar]
- 17. Farag AT, Goda NF, Shaaban NA, Mansee AH (2007) Effects of oral exposure of synthetic pyrethroid, cypermethrin on the behavior of F1-progeny in mice. Reprod Toxicol 23: 560–567. [DOI] [PubMed] [Google Scholar]
- 18. George J, Srivastava AK, Singh R, Shukla Y (2011) Cypermethrin exposure leads to regulation of proteins expression involved in neoplastic transformation in mouse skin. Proteomics 11: 4411–4421. [DOI] [PubMed] [Google Scholar]
- 19. Jin Y, Wang L, Ruan M, Liu J, Yang Y, et al. (2011) Cypermethrin exposure during puberty induces oxidative stress and endocrine disruption in male mice. Chemosphere 84: 124–130. [DOI] [PubMed] [Google Scholar]
- 20. Wang X, Liu S, Sun Y, Wu J, Zhou Y, et al. (2009) Beta-cypermethrin impairs reproductive function in male mice by inducing oxidative stress. Theriogenology 72: 599–611. [DOI] [PubMed] [Google Scholar]
- 21. Meeker JD, Barr DB, Hauser R (2009) Pyrethroid insecticide metabolites are associated with sperm hormone levels in adult men. Reprod Toxicol 27: 155–160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Wang H, Wang SF, Ning H, Ji YL, Zhang C, et al. (2011) Maternal cypermethrin exposure during lactation impairs testicular development and spermatogenesis in male mouse offspring. Inc Environ Toxicol 26: 382–394. [DOI] [PubMed] [Google Scholar]
- 23. Shafer TJ, Meyer DA, Crofton KM (2005) Developmental neurotoxicity of pyrethroid insecticides: critical review and future research needs. Environ Health Persp 113: 123–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Wolansky MJ, Harrill JA (2008) Neurobehavioral toxicology of pyrethroid insecticides in adult animals: a critical review. Neurotoxicol Teratol 30: 55–78. [DOI] [PubMed] [Google Scholar]
- 25. Yang L, Chen S, Hu M, Hao W, Geng P, et al. (2011) Biodegradation of carbofuran by Pichia anomala strain HQ-C-01 and its application for bioremediation of contaminated soils. Biol Fert Soils 47: 917–923. [Google Scholar]
- 26. Arora PK, Sasikala C, Ramana CV (2012) Degradation of chlorinated nitroaromatic compounds. Appl Microbiol Biotechnol 93: 2265–2277. [DOI] [PubMed] [Google Scholar]
- 27. Cycoń M, Wojcik M, Piotrowska-Seget Z (2011) Biodegradation kinetics of the benzimidazole fungicide thiophanate-methyl by bacteria isolated from loamy sand soil. Biodegradation 22: 573–583. [DOI] [PubMed] [Google Scholar]
- 28. Cycoń M, Zmijowska A, Wójcik M, Piotrowska-Seget Z (2013) Biodegradation and bioremediation potential of diazinon-degrading Serratia marcescens to remove other organophosphorus pesticides from soils. J Environ Manage 117: 7–16. [DOI] [PubMed] [Google Scholar]
- 29. Chen S, Hu W, Xiao Y, Deng Y, Jia J, et al. (2012) Degradation of 3-phenoxybenzoic acid by a Bacillus sp. PLoS ONE 7: e50456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Chen S, Liu C, Peng C, Liu H, Hu M, et al. (2012) Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-trichloro-2-pyridinol by a new fungal strain Cladosporium cladosporioides Hu-01. PLoS ONE 6: e47205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Zhang C, Jia L, Wang SH, Qu J, Xu LL, et al. (2010) Biodegradation of beta-cypermethrin by two Serratia spp. with different cell surface hydrophobicity. Bioresour Technol 101: 3423–3429. [DOI] [PubMed] [Google Scholar]
- 32. Chen S, Hu M, Liu J, Zhong G, Yang L, et al. (2011) Biodegradation of beta-cypermethrin and 3-phenoxybenzoic acid by a novel Ochrobactrum lupini DG-S-01. J Hazard Mater 187: 433–440. [DOI] [PubMed] [Google Scholar]
- 33. Wu PC, Liu YH, Wang ZY, Zhang XY, Li H, et al. (2006) Molecular cloning, purification, and biochemical characterization of a novel pyrethroid-hydrolyzing esterase from Klebsiella sp. strain ZD112. J Agric Food Chem 54: 836–842. [DOI] [PubMed] [Google Scholar]
- 34. Wang BZ, Guo P, Hang BJ, Li L, He J, et al. (2009) Cloning of a novel pyrethroid-hadrolyzing carboxylesterase gene from Sphingobium sp. strain JZ-1 and characteization of the gene product. Appl Environ Microbiol 75: 5496–5500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zhai Y, Li K, Song J, Shi Y, Yan Y (2012) Molecular cloning, purification and biochemical characterization of a novel pyrethroid-hydrolyzing carboxylesterase gene from Ochrobactrum anthropi YZ-1. J Hazard Mater 221–222: 206–212. [DOI] [PubMed] [Google Scholar]
- 36. Fan X, Liu X, Huang R, Liu Y (2012) Identification and characterization of a novel thermostable pyrethroid-hydrolyzing enzyme isolated through metagenomic approach. Microb Cell Fact 11: 33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Bradford MM (1976) A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248–253. [DOI] [PubMed] [Google Scholar]
- 38. Chen S, Hu Q, Hu M, Luo J, Weng Q, et al. (2011) Isolation and characterization of a fungus able to degrade pyrethroids and 3-phenoxybenzaldehyde. Bioresour Technol 102: 8110–8116. [DOI] [PubMed] [Google Scholar]
- 39. Liang WQ, Wang ZY, Li H, Wu PC, Hu JM, et al. (2005) Purification and characterization of a novel pyrethroid hydrolase from Aspergillus niger ZD11. J Agric Food Chem 53: 7415–7420. [DOI] [PubMed] [Google Scholar]
- 40. Guo P, Wang BZ, Hang BJ, Li L, Ali SW, et al. (2009) Pyrethroid-degrading Sphingobium sp. JZ-2 and the purification and characterization of a novel pyrethroid hydrolase. Int Biodeterior Biodegrad 63: 1107–1112. [Google Scholar]
- 41. Gao Y, Chen SH, Hu MY, Hu QB, Luo JJ, et al. (2012) Purification and characterization of a novel chlorpyrifos hydrolase from Cladosporium cladosporioides Hu-01. PLoS ONE 7: e38137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Laemmli UK (1970) Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature 227: 680–685. [DOI] [PubMed] [Google Scholar]
- 43. Peng Y, Chi ZM, Wang XH, Li J (2009) Purification and molecular characterization of exo-β-1,3-glucanases from the marine yeast Williopsis saturnus WC91–2. Appl Microbiol Biotechnol 85: 85–94. [DOI] [PubMed] [Google Scholar]
- 44. Tallur PN, Megadi VB, Ninnekar HZ (2008) Biodegradation of cypermethrin by Micrococcus sp. strain CPN 1. Biodegradation 28: 77–82. [DOI] [PubMed] [Google Scholar]
- 45. Chen S, Luo J, Hu M, Lai K, Geng P, et al. (2012) Enhancement of cypermethrin degradation by a coculture of Bacillus cereus ZH-3 and Streptomyces aureus HP-S-01. Bioresour Technol 110: 97–104. [DOI] [PubMed] [Google Scholar]
- 46. Chen SH, Dong YH, Chang CC, Deng YY, Zhang XF, et al. (2013) Characterization of a novel cyfluthrin-degrading bacterial strain Brevibacterium aureum and its biochemical degradation pathway. Bioresour Technol 132: 16–23. [DOI] [PubMed] [Google Scholar]
- 47. Maloeny SE, Maule A, Smith ARW (1993) Purification and preliminary characterization of permethrinase from a pyrethroid-transforming strain of Bacillus cereus . Appl Environ Microbiol 59: 2007–2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Li G, Wang K, Liu YH (2008) Molecular cloning and characterization of a novel pyrethroid-hydrolyzing esterase originating from the metagenome. Microb Cell Fact 7: 38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Chen SH, Luo JJ, Hu MY, Geng P, Zhang YB (2012) Microbial detoxification of bifenthrin by a novel yeast and its potential for contaminated soils treatment. PLoS ONE 7: e30862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Heidari R, Devonshire AL, Campbell BE, Dorrian SJ, Oakeshott JG, et al. (2005) Hydrolysis of pyrethroids by carboxylesterases from Lucilia cuprina and Drosophila melanogaster with active sites modified by in vitro mutagenesis. Insect Biochem Mol Biol 35: 597–609. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The crude enzyme activity on degrading beta-CP over the incubation time.
(TIF)
SDS-PAGE analysis of the fractions showing beta-CP degrading enzyme activity obtained during the purification. Lane M: protein markers, 14.4–94.0 kDa; Lane 1: total proteins; Lane 2: fractions from ammonium sulfate precipitation; Lane 3: fractions from DEAE Sepharose Fast Flow anion-exchange.
(TIF)
GC-MS spectra of products produced from beta-CP degradation by CMO. a, trans-Beta-CP; b, cis-Beta-CP; c, 2-(4-Hydroxyphenoxy) benzoic acid methyl ester; d, 3,5-Dihydroxy benzoic acid; e, 3,5-Dimethoxy phenol; f, 4-Hydroxy-3-methoxy benzoic acid; g, Phenol.
(TIF)