

Abstract
We describe the use of carbon stable isotope (13C) labeled n-alkanes as a potential internal tracer to assess passage kinetics of ingested nutrients in ruminants. Plant cuticular n-alkanes originating from intrinsically 13C labeled ryegrass plants were pulse dosed intraruminally in four rumen-cannulated lactating dairy cows receiving four contrasting ryegrass silage treatments that differed in nitrogen fertilization level (45 or 90 kg nitrogen ha−1) and maturity (early or late). Passage kinetics through the gastrointestinal tract were derived from the δ13C (i.e. the ratio 13C:12C) in apparently undigested fecal material. Isotopic enrichment was observed in a wide range of long-chain n-alkanes (C27–C36) and passage kinetics were determined for the most abundant C29, C31 and C33 n-alkanes, for which a sufficiently high response signal was detected by combustion isotope ratio mass spectrometry. Basal diet treatment and carbon chain length of n-alkanes did not affect fractional passage rates from the rumen (K 1) among individual n-alkanes (3.71–3.95%/h). Peak concentration time and transit time showed a quantitatively small, significant (p≤0.002) increase with carbon chain length. K 1 estimates were comparable to those of the 13C labeled digestible dry matter fraction (3.38%/h; r = 0.61 to 0.71; p≤0.012). A literature review has shown that n-alkanes are not fermented by microorganisms in the rumen and affirms no preferential depletion of 13C versus 12C. Our results suggest that 13C labeled n-alkanes can be used as nutrient passage tracers and support the reliability of the δ13C signature of digestible feed nutrients as a tool to measure nutrient-specific passage kinetics.
Introduction
Production animals need to be fed according to their nutritional requirements in order to reach their maximum performance, reduce the loss of waste products into the environment from undigested feed nutrients, and prevent nutrient-related disorders due to an unbalanced or insufficient supply of nutrients. Knowledge on the behavior of ingested feed nutrients in the different compartments of the gastrointestinal tract is essential to understand the fate of nutrients and related digestive mechanisms [1]. In ruminants, which are particularly adapted to yield energy from poor-quality forages, the reticulorumen is the main site of fermentative degradation of nutrients through the action of microorganisms, mixing of the ingesta and particle size reduction. Once ingested feed particles reach a specific particle size [2] and specific density [3], they pass in aboral direction into the following digestive compartment at specific fractional rates (i.e., as a fraction per hour; %/h) depending on a number of feed and animal characteristics [4], [5]. If quantified accurately, knowledge on the fractional passage rate from the reticulorumen may be used to predict the extent of degradation and excretion of nutrients [1]. Fractional passage rates are, therefore, an essential part of modern feed evaluation systems as well as of mechanistic models describing the dynamics of microbial population in the rumen and their contribution to methane emissions from ruminants [6]. Yet, quantitative knowledge on feed- and nutrient-specific passage kinetics are limited.
Passage kinetics are commonly estimated by tracer techniques. External tracers were frequently used but are not intrinsic to the diet and may, therefore, not reflect the passage behavior of dietary components. Recently, the carbon isotope signature of feed nutrients (δ13C; i.e. the ratio of the stable isotopes 13C to 12C) has been proposed as an internal passage tracer [5]. In particular, δ13C in fecally excreted undigested fibers allowed researchers to quantify nutrient-specific passage in dairy cows. It was further proposed to use stable isotopes in combination with n-alkanes as an internal passage tracer [7]. n-Alkanes are saturated aliphatic hydrocarbon chains naturally present in plant cuticular wax. They have a high fecal recovery in ruminants depending on the carbon chain length [8], [9], are neither degraded nor synthesized in the rumen [10], [11], and their analytical determination is sensitive and specific [7]; hence, they possess close-to-ideal tracer characteristics.
The objective of this study was to evaluate whether the δ13C signature of n-alkanes can be used to estimate passage kinetics in ruminants. To our knowledge no literature is available on the application of stable isotope labeled n-alkanes as fractional passage rate tracers, although δ13C of individual n-alkanes were successfully employed to assess feed digestibility of ruminants [12]. The present study describes for the first time in vivo δ13C labeled n-alkanes and their relevance to passage kinetics studies in dairy cattle.
Materials and Methods
Animals and Housing
The present study was part of a larger experiment on passage kinetics of nutrients through the gastrointestinal tract of dairy cows, approved by the Institutional Animal Care and Use Committee of Wageningen University, Wageningen, The Netherlands (no. 2010095). Four multiparous Holstein-Friesian dairy cows in their second to fourth lactation, fitted with a permanent rumen cannula (10 cm i.d., Type 1C, Bar Diamond, Parma, ID), were individually housed in tie stalls. Animals averaged (mean ± SEM) 561±13 kg in bodyweight, and, during measurement weeks (n = 4), had a dry matter (DM) intake of 17.0±0.35 kg/d and produced 26.2±0.87 kg milk/d. Animals were milked twice daily during feeding times.
Diet and Treatments
Animals were fed a total mixed ration consisting of 455 g/kg DM ryegrass silage, 195 g/kg DM corn silage and 350 g/kg DM of a specifically designed compound feed ( Table 1 ). The compound feed ingredients originated from cool-season C3 plants to keep the background level of 13C low [13] and similar to that of the natural levels of the grass silage mixed in the experimental diet. Grass silage was prepared from perennial ryegrass (Lolium perenne) from the second regrowth, fertilized at two nitrogen (N; as potassium phosphorus nitrate) levels, and harvested at two maturity stages. Levels of fertilization were either 45 kg N/ha (N45), or 90 kg N/ha (N90). Maturity stages were set to obtain a target DM yield in the range of 1800–2000 kg/ha (early) and 4600–4800 kg/ha (late). The grass plants were harvested in September 2010, wilted and ensiled. Animals were offered daily rations of the diet as two equal meals at 0600 and 1700 h, and had free access to water. The diet was prepared twice weekly; feed ingredient samples were collected each time the diet was prepared and pooled per animal over each experimental period.
Table 1. Chemical composition of the diet consisting of grass silage of early or late maturity at two nitrogen (N) fertilization levels (N45∶45 kg N/ha; N90∶90 kg N/ha), corn silage and compound feed.
| N45 | N90 | |||||
| Chemical composition1 | early | late | early | late | Corn silage | Compound feed2 |
| Dry matter (g/kg fresh) | 366 | 723 | 567 | 520 | – | – |
| Dry matter (kg/ha) | 1840 | 4734 | 2020 | 4860 | – | – |
| Organic matter | 879 | 912 | 907 | 903 | 961 | 933 |
| Crude protein | 197 | 137 | 249 | 168 | 77 | 262 |
| Starch | – | – | – | – | 403 | 218 |
| Neutral detergent fiber | 432 | 545 | 429 | 556 | 356 | 293 |
| Acid detergent fiber | 294 | 296 | 257 | 326 | 201 | 151 |
In g/kg dry matter unless specified otherwise (starch not determined for grass silage).
Ingredients (g/kg dry matter): wheat (80.0), sunflower seeds (140.0), soybean hulls (26.5), palm kernel expeller (90.0), soybeans (185.0), sugar beet pulp (75.0), potato starch (200.0), rumen-protected soybean meal (185.0; MervoBest, Pre-Mervo, Utrecht, The Netherlands), phosphoric acid limestone (7.5), salt (3.0), mineral premix (8.0).
Tracer Preparation
The external tracer chromium mordanted fiber (Cr-NDF) was prepared as described by Udén [14] from wheat straw, dried and ground to pass a 0.5-mm screen. The 13C tracer was prepared from δ13C labeled and ensiled ryegrass originating from the field that also provided the basal experimental diet. In brief, representative ryegrass shoots from the second regrowth were randomly collected from the field and grown on hydroponics (8.6 shoots per m2) under climate-controlled greenhouse conditions in hermetically sealed isotope assimilation chambers, specifically designed for homogeneous atmospheric isotope labeling [15]. Grass was continuously enriched under high levels of 13carbon dioxide (released from 99.98 atom% 13C bicarbonate) from plant emergence onwards in a commercial facility (IsoLife BV, Wageningen, The Netherlands). The labeled grass plants were exposed to similar conditions to the field plants to account for potential known sources of variation for an altered cell wall structure and n-alkanes content in temperate forages [16]; e.g., by adjusting the light schedule in the greenhouse to the field conditions, and inducing wind stress to the labeled plants. Grass plants received the identical fertilization regimen and harvested at a similar physiological stage (172 g DM/m2 and 498 g DM/m2 for early and late maturity stage, respectively) as the field plants. Plants were subsequently wilted, cut to size (2 cm), placed into several bags of larger mashed grit gauze (pore size 212 µm; PA-74, Sefar Nytal, Heiden, Switzerland) and distributed over silage bales to be ensiled together with the field plants over an 8-week period. The stable isotope labeling was successfully terminated by collecting ensiled ryegrass plants enriched up to 7.75–9.21 atom% 13C for the different treatments relative to a background level of 1.01 atom% 13C (SD 0.001) for the unlabeled field plants.
Sampling and Measurements
Four experimental periods of three weeks each were used, which included an adaptation after diet changeover from day 1 through 14, followed by fecal sampling from day 15 through 19. From day 12 onwards, animals were fed 95% of the individual DM intake measured during the preceding adaptation days to minimize feed residuals. Feed and water uptake were monitored daily and animals were milked twice daily.
Animals received a ruminal pulse dose of 100 g DM Cr-NDF (45.9 g Cr/kg DM Cr mordant) and 15 g DM δ13C labeled grass silage. Prior to pulse-dosing, the labeled tracer material was cut to pieces of about 0.5 cm to resemble ingested bulk grass silage particles.
From day 15 onwards, directly after administration of tracers into the rumen (0900 h), 20 spot samples of feces were collected after defecation in sampling blocks of three hours each. Ten fecal samples collected at times t = 0, 12, 18, 24, 30, 36, 48, 72, 96 and 120 h after pulse dose administration were analyzed for their tracer concentrations. Feces were weighed, thoroughly homogenized by hand and a representative sample of about 400 g fresh matter was stored at −20°C pending analyses.
Chemical Analyses
All samples were freeze-dried and ground over a hammer mill to pass a 1-mm screen (Peppink 100 AN, Olst, The Netherlands). Dry matter, ash, crude protein, starch, NDF and ADF were analyzed as described by Abrahamse [17], [18]. Fecal Cr concentrations were determined using an atomic absorption spectrophotometer (AA240FS, Varian, Palo Alto, CA) after oxidation by wet-destruction [5]. n-Alkane extraction was carried out after Mayes [19] with modifications after Olívan [20] and Bezabih [21] and using tetratriacontane (C34) as an internal standard. In brief, test samples were pulverized (MM2000, Retsch, Haan, Germany; 3 min at 85 Hz) prior to n-alkane extraction. Full base line separation of n-alkanes (C27 to C36) was achieved using a gas chromatograph (GC; Finnigan Trace GC Ultra, Milan, Italy) fitted with a capillary column (40 m x 0.32 mm i.d. fused silica capillary SPB-1 and 0.10 µm film thickness) and using helium as a carrier gas at a constant flow of 2.5 ml/min. n-Alkane extracts, previously diluted with 125 µL of heptane, were injected using a split/splitless-type injector operating on split mode (split ratio of 1∶5). The temperature for the injector was 270°C. The oven temperature program started at 210°C (maintained for 1 min), increased at a rate of 7.2°C/min to a temperature of 300°C (maintained for 6 min). To determine the δ13C of individual n-alkanes, the column outlet was fitted to a combustion interface (Thermo Finnigan GC Combustion III, Bremen, Germany) that was connected to an isotope ratio mass spectrometer (IRMS; Delta V Advantage, Thermo Scientific, Bremen Germany). For δ13C analyses of the apparent undigested fecal DM fraction (13C-DM), a test sample was pulverized, and δ13C was determined by elemental analysis using an IRMS as describe above. The relative atom% 13C in the substrate is expressed as the 13C:12C ratio in the samples relative to the 13C:12C ratio of the international Vienna Pee Dee Belemnite standard, and presented as δ13C. After correction for natural 13C abundance, fecal excretion patterns of atom% 13C excess were established.
Curve Fitting and Statistical Analyses
Fractional passage rates were derived from tracer excretion patterns, fitted iteratively with a nonlinear multicompartmental model [22]:
where C t denotes the fecal tracer concentration at time = t; t is the average time span of collection after tracer administration; K 1 and K 2 refer to the fractional rate constants for the compartment with the longest (reticulorumen) and the second longest retention time (proximal colon-cecum) in the gastrointestinal tract, respectively; N refers to the model-derived number of mixing compartments; and A forms a scalable parameter dependent on K 1, K 2 and N.
Before curve fitting, fecal tracer concentrations were scaled to the tracer peak concentration [23]. Curve fitting was performed using nonlinear least squares regression procedures of SAS (version 9.2, Cary, NC) based on the least square Levenberg-Marquardt algorithm. Initial values for the iterative procedure were obtained through a grid search and curve fits were solved after, on average, 6 to 9 iterations. Transit time (TT; i.e. moment of first appearance of the tracer in the feces) and moment of peak concentration (PCT) were derived for fecal tracer excretion patterns from the estimated parameters as described by Dhanoa [22]. Total mean retention time (TMRT) in the reticulorumen was calculated as the sum of the reciprocals of K 1 and K 2 plus TT; total tracer clearance time was calculated as described by France [24]. Accuracy of curve fits were evaluated by comparing predicted tracer concentrations with observed values using the root mean squared prediction error relative to the observed mean, thus obtaining the mean prediction error (MPE). The MPE was decomposed into errors due to random variation, errors of central tendency and errors due to regression [25].
Model parameters were log transformed due to asymmetrical distribution patterns of residuals and tested by analysis of variance in a split plot, with factorial main plots in a Latin square and subplots representing the type of n-alkane, using the mixed model procedure of SAS (version 9.2, Cary, NC), according to the model:
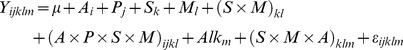 |
where Y ijklm is the dependent variable; μ is the overall mean; Ai (animal; i = 4), Pj (period; j = 4), Sk (silage; k = 2), Ml (maturity; l = 2) and its interaction term (S × M)kl represent effects assigned to the main plots in a Latin square; Alkm (type of n-alkane; m = 3) and (S × M × Alk)klm represent effects related to the subplots. Main plot variables were tested against the interaction term (A × P × S × M)ijkl and subplot variables were tested against the pooled residual error (εijklm). Covariance parameters were estimated using the residual maximum likelihood (REML) method and denominator degrees of freedom were estimated using the Satterthwaite approximation. Tables report back-transformed values. Passage kinetics of 13C-n-alkanes were compared to the commonly used external tracer Cr-NDF and the internal tracer 13C-DM originating from the same substrate by means of the Pearson correlation coefficient r.
Results
Concentrations of 13C isotopes were detected in a wide range of n-alkanes ( Table 2 ). A high signal amplitude voltage by combustion isotopic ratio mass spectrometry was obtained for the odd-chain n-alkanes C29, C31 and C33, which were also used to assess fractional passage (in total, 48 curve fits). The signal amplitude voltage obtained for the even-chain n-alkanes (C28–C36; amplitude always below 0.3 V) and for the odd-chain n-alkanes C27 and C35 (amplitude always below 0.5 V, with up to 72% of values below 0.3 V) were below or close to the limit of quantitation (defined here as 0.3 V) and considered too low to obtain a reliable δ13C based on the analytical procedure and equipment used ( Figure 1 ). Background levels of δ13C (i.e. natural enrichment) decreased with carbon chain length of odd-chain n-alkanes ( Table 2 ). Differences between δ13C background and peak levels were generally higher for the odd-chain n-alkanes and highest for C29, C31, C33 and C35.
Table 2. Mean background and peak concentrations of δ13C for individual n-alkanes in feces as a mean of four grass silage treatments.
| n-Alkane | Background (δ) | Peak (δ) | Difference (δ)1 |
| Even-chain | |||
| C28 | −33.07 (0.592) | −29.84 (0.279) | 3.30 (0.531) |
| C30 | −35.10 (0.526) | −30.29 (0.751) | 5.19 (0.512) |
| C32 | −33.81 (0.753) | −30.03 (0.579) | 3.79 (0.611) |
| C36 | −29.51 (0.302) | −27.86 (0.271) | 1.66 (0.370) |
| Odd-chain | |||
| C27 | −32.90 (0.577) | −29.23 (0.611) | 3.67 (0.546) |
| C29 | −35.58 (0.241) | −29.25 (0.714) | 6.33 (0.557) |
| C31 | −36.08 (0.195) | −27.57 (0.888) | 8.51 (0.753) |
| C33 | −36.69 (0.223) | −29.95 (0.800) | 6.73 (0.615) |
| C35 | −37.27 (0.518) | −30.37 (0.840) | 6.90 (0.692) |
Standard error of the mean given in parenthesis. C34: internal standard. δ13C refers to the relative atom% 13C in the sample relative to the atom% 13C of the international Vienna Pee Dee Belemnite standard.
Difference between δ13C background and peak concentration based on least square means.
Figure 1. Mass spectra for δ13C labeled n-alkanes (C27 to C36) collected in feces upon an intraruminal pulse dose.

Plots A–B show mass chromatograms with signal amplitude voltage (V; lower plot segments) of individual n-alkanes for the carbon isotopes 12C and 13C (from lowest to highest concentration, respectively), and their respective 13C:12C ratio (upper plot segments). C34 is the internal standard. Mass spectra illustrate the distinctive peaks of the most abundant n-alkanes C29, C31 and C33 originating from a highly enriched fecal sample collected at peak concentration time (mean δ13C −28.84; SEM 1.736). Although signals were detected and distinctive peaks were identified for lesser abundant n-alkanes (plot A), the 13C:12C ratio indicates that the respective δ13C levels were below or close to quantitation limit (0.3 V; plot B). Lower enriched fecal samples often provided no response signal for lesser abundant n-alkanes. Test samples were pre-concentrated by reducing the amount of solvent to 125 µL, and were injected at a split ratio 1∶5 using a split/splitless-type injector operating on split mode to obtain a high peak resolution.
Fecal excretion patterns of 13C-n-alkanes were characterized by an initial quickly ascending phase until moment of tracer peak concentration (PCT) followed by a slowly descending phase ( Figure 2 ). The 13C concentrations in n-alkanes were close to their natural abundance when fecal sampling was terminated as shown by the mean total tracer clearance time of 134±4.9 h (mean ± SEM). The 48 curve fits established for 13C-n-alkanes showed a mean prediction error (MPE) of 9.9±4.68% ( Table 3 ), of which 92.7±1.14% were related to errors due to random variation, 2.7±0.45% to errors of central tendency and 4.6±0.70% to errors due to regression. n-Alkane carbon chain length did not affect MPE of curve fits.
Figure 2. Fecal dilution curves of δ13C labeled C31 n-alkane (•) and chromium mordanted fiber (×).

Dilution curves show mean fecal tracer concentrations with standard error bars upon an intraruminal pulse dose in dairy cows fed grass silages of early and late maturity at two nitrogen (N) fertilization levels (N45∶45 kg N/ha; N90∶90 kg N/ha).
Table 3. Passage kinetics of δ13C labeled odd-chain n-alkanes (C29, C31, C33) in dairy cows fed grass silages of early and late maturity at two nitrogen (N) fertilization levels (N45∶45 kg N/ha; N90∶90 kg N/ha).
| Item | K 1 | K 2 | PCT | TT | TMRT | MPE |
| n-Alkanes (Alk) | ||||||
| C29 | 3.95 (0.158) | 21.3 (2.81) | 22.6 (2.93) | 13.4 (0.37) | 43.9 (0.74) | 9.7 (2.08) |
| C31 | 3.71 (0.148) | 20.8 (2.74) | 22.7 (2.94) | 13.0 (0.36) | 44.9 (0.75) | 7.8 (1.66) |
| C33 | 3.93 (0.157) | 18.8 (2.47) | 23.9 (3.10) | 14.0 (0.39) | 45.3 (0.76) | 9.8 (2.10) |
| Fertilization level (F) | ||||||
| N45 | 4.05 (0.204) | 17.0 (3.13) | 26.7 (4.89) | 13.8 (0.40) | 45.0 (1.24) | 8.4 (2.40) |
| N90 | 3.67 (1.849) | 24.2 (4.46) | 19.9 (3.64) | 13.1 (0.44) | 44.3 (1.22) | 9.8 (2.81) |
| Maturity stage (M) | ||||||
| Early | 3.76 (0.232) | 20.6 (4.65) | 22.6 (5.07) | 13.1 (0.48) | 45.1 (1.15) | 12.9 (4.53) |
| Late | 3.96 (0.244) | 20.0 (4.51) | 23.5 (5.27 ) | 13.7 (0.50) | 44.3 (1.13) | 6.4 (2.23) |
| P-values1 | ||||||
| Main plots | ||||||
| Animal | 0.370 | 0.673 | 0.614 | 0.265 | 0.472 | 0.746 |
| Period | 0.010 | 0.908 | 0.771 | 0.147 | 0.010 | 0.424 |
| F | 0.216 | 0.225 | 0.299 | 0.332 | 0.612 | 0.709 |
| M | 0.618 | 0.936 | 0.920 | 0.535 | 0.680 | 0.264 |
| F × M | 0.940 | 0.523 | 0.379 | 0.651 | 0.178 | 0.667 |
| Subplots | ||||||
| Alk | 0.120 | 0.002 | <0.001 | 0.047 | 0.088 | 0.100 |
| Alk × F × M | 0.050 | 0.173 | 0.022 | 0.167 | 0.018 | 0.167 |
K 1: fractional passage rate constant (%/h) from the reticulorumen; K 2: fractional passage rate constant (%/h) from the proximal colon-cecum; PCT: tracer peak concentration time (h); TT: tracer transit time (h); TMRT: total mean retention time (h); MPE: mean prediction error (% of observed mean); values represent means (n = 16 per n-alkane type) and respective standard error of the mean in parenthesis.
Analyses of variance on log-transformed means.
Basal diet had no effect on passage kinetics ( Table 3 ). n-Alkane carbon chain length did not affect the respective K 1 estimates (3.71–3.95%/h). Quantitatively small, significant changes in some passage kinetic parameters occurred with increasing carbon chain length, such as a decrease of K 2 (p = 0.002) and an increase in PCT (p<0.001) and TT (p = 0.047). Total mean retention time (TMRT) was not different among n-alkanes (43.9–45.3 h; p = 0.088). The model parameters N (model-derived hypothetical number of mixing compartments) and A (scalable model parameter) as obtained after fitting excretion data with a multicompartmental model were 13±2.5 and 3.8±0.42, respectively (data not shown). Fecal 13C-DM gave a mean K 1 value of 3.38±0.315%/h, a K 2 value of 24.1±1.84%/h, and a TMRT of 47.1±2.52 h. Fecal Cr-NDF gave a mean K 1 value of 5.25±0.490%/h, a K 2 value of 31.2±2.39%/h, and a TMRT of 35.7±1.91 h (data not shown).
Discussion
The Labeling and Analytical Determination of Carbon Isotope n-Alkanes
The present study is the first describing stable isotope (13C) labeled n-alkanes from in vivo isotopic labeled plant material and its application in digesta passage studies in ruminants. A first attempt to estimate fractional rumen passage rates in small ruminants from radioactive isotope (14C/3H) labeled n-alkanes (originating from fresh perennial ryegrass) has been made by Mayes [26] and was published as a conference proceedings abstract. Details on the labeling procedure were not provided but appear to involve immersion of fresh grass in a solution of 14C labeled acetate followed by a short exposure to a high-intensity light source [8] for temporary 14C assimilation.
We quantified passage kinetics for the most abundant 13C-n-alkanes C29, C31 and C33 in apparent undigested feces. The use of a combustion isotope ratio mass spectrometer allowed detection of δ13C for the long-chain C27–36 n-alkanes. The even-chain and the odd-chain C27 and C35 n-alkanes offered particular weak δ13C signals, most probably because of their low natural concentrations generally observed in plant biomass and feces [19]. In ryegrass species, concentrations of the most abundant n-alkanes C29, C31 and C33 typically ranged from 77 to 338 mg/kg DM [27], [28]. The GC-IRMS was set up to present a high amount of sample for analyses (see Figure 1 ) and thus allow for high analytical sensitivity. A higher sensitivity of the low enriched n-alkanes was not feasible due to the relatively high enrichment levels and corresponding strong signal response of some of the adjacent n-alkanes approaching the upper analytical detection limit for δ13C.
Continuous intrinsic isotope labeling applied in our study was shown to provide uniformly labeled plant material [15], [29]. Differences in δ13C background levels between n-alkanes ( Table 2 ) appeared to depend on the respective chain length and suggest that carbon isotope discrimination occurs during the enzymatic n-alkane biosynthesis by decarbonylase in the plant cuticula. Similar observations were reported for the biosynthesis of lignin [30] and starch [31] in plant tissue. Differential carbon allocation as observed in the plant tissue does not occur in the animal organism due to the absence of ruminal synthesis and degradation of n-alkanes [10], [11]. Microbial fermentation in the hindgut of ruminants is minor [32] and a hypothetical degradation of n-alkanes and preferential carbon isotope disappearance from the hindgut is therefore unlikely to affect passage kinetics estimations of n-alkanes from fecal samples. Disappearance of some ingested n-alkanes in the gastrointestinal tract has been reported and generally decreased with carbon chain length [8], [9]. However, a literature review suggests that absorption from the small intestine rather than ruminal degradation accounts for the main loss of n-alkanes [8].
Passage Kinetics Assessment of Isotope Labeled n-Alkanes
Basal diet treatments had no effect on passage kinetics in our study despite a considerable change in the dietary nutritional composition. The carbon chain length of n-alkanes affected various passage kinetic parameters but did not affect K 1 estimates. Overall, TMRT and total tracer clearance time from the gastrointestinal tract were similar among n-alkanes. When compared to a commonly used external digesta passage tracer, passage kinetics of 13C-n-alkanes differed considerably from those of Cr-NDF (on average 3.86 and 5.25%/h, respectively). No significant correlations between the two tracers were observed for K 1 and TMRT (p>0.10; Figure 3 ). The discrepancy in TMRT between the two tracers is in line with observations on 14C/3H labeled n-alkanes and Cr-NDF in small ruminants [26]. The discrepancy in K 1 between the two tracers might be explained by potential differences in particle size [33] and the increased feed particle density [34] with a resulting reduced buoyancy [35] commonly observed for Cr-NDF particles. The degree of tracer association with the particulate matter might be a confounding factor throughout passage studies [36], although Mayes [26] found a nearly full affinity (0.98) of those tracers.
Figure 3. Relationship between δ13C labeled n-alkanes and chromium mordanted fiber or δ13C labeled dry matter.

n-Alkanes: C29 (□), C31 (○), C33 (Δ). Pearson correlation coefficients shown in plots. K 1 = fractional rumen passage rate (%/h); TMRT = total mean retention time (h). Mean K 1 is (mean ± SEM) 3.38±0.315%/h for δ13C labeled dry matter and 5.25±0.490%/h for chromium mordanted fiber; mean TMRT is 47.1±2.52 h for δ13C labeled dry matter and 35.7±1.91 h for chromium mordanted fiber.
When compared to an alternative internal digesta passage tracer, passage kinetics of 13C-n-alkanes was comparable to that of 13C-DM originating from the same labeled plant material ( Figure 3 ). n-Alkanes were shown to associate well to the particulate DM pool [26], which, given the few data points in our study, sustains the overall satisfactory resemblance in passage kinetics observed for 13C-n-alkanes and 13C-DM. Pearson correlation coefficient r ranged from 0.61 to 0.71 (p≤0.012; n = 16) for K 1 (on average 3.38%/h), and from 0.75 to 0.80 (p≤0.001; n = 16) for TMRT (47.1 h). Our K 1 estimates are in line with recent data on δ13C labeled ryegrass silage of high and low digestibility [5]. They reported mean K 1 estimates of 3.52–3.85%/h and 4.76–5.03%/h for 13C-DM and Cr-NDF, respectively. The Pellikaan [5] study employed two dairy cows fed grass silage and compound feed, and their 13C labeling approach consisted of pulse labeling plants up to eight times on field. For even-chain n-alkanes sprayed onto ryegrass leaves or stems, K 1 values were considerably higher (7.5–9.5%/h [37]), whereas K 1 values for Cr-NDF (4.0%/h) were in line with studies using Cr-NDF in dairy cows [5], [38]. These exceptionally high rates for forages were partly explained by the binding association of the sprayed n-alkanes onto the plant cell wall matrix, which can be low for a synthetic matrix [8], resulting in migration of sprayed n-alkanes to the liquid phase [39]. Observations by Mayes [26] further suggest a considerably lower binding association of dosed even-chain n-alkanes (0.92) compared to natural or 14C/3H labeled n-alkanes from ryegrass plants (0.98). They reported considerably lower K 1 values of 2.85–2.98%/h (based on even-chain n-alkanes sprayed onto ryegrass leaves and stems), and 2.54–2.56%/h (based on 14C/3H-n-alkanes from ryegrass) for small ruminants, in contrast to the findings of Giráldez [37]. Assuming that animal species have a minor effect on K 1 [40], [41], differences in feed intake level between the various studies might explain the somewhat higher K 1 values for Cr-NDF in our study. Reservations on the spraying technique, which implies an external application of the even-chain n-alkanes, have been expressed with regard to rare earth elements as it was observed that plant tissue did not uniformly absorb the sprayed-on tracer [42]. Passage kinetics are therefore highly dependent on the absorption capacity of the plant cell wall matrix. In contrast, intrinsically isotope labeled plants will circumvent this problem, as the 13C isotopes are homogeneously distributed in the plant tissue when continuously labeled in a greenhouse [15], [29].
The Use of Carbon Isotope Labeled n-Alkanes for Digesta Passage Kinetic Studies
Various studies have shown that isotopes from intrinsically labeled plants can be detected in various undigested fractions of feces and digesta of ruminants, such as in various fiber fractions [5], [43] and starch [44]. In addition to the plant cuticular n-alkanes, isotopes can be potentially detected in further plant wax components, such as in long-chain alcohols and very long-chain fatty acids, which are typically present in considerably higher concentrations than n-alkanes in perennial ryegrass [27], [28]. This could be of particular interest for quantifying passage kinetics of a diet composed of plant species containing low concentrations of n-alkanes such as some of the common temperate grass species (e.g. Phleum pratense and Dactylis glomerata [8]) and some tropical forages [12], [21].
The use of isotopic labeled n-alkanes could be of particular interest for comparative passage rate studies to study the rumen physiology and its evolutionary mechanisms [45], [46] of domestic and wild ruminants [47], [48]. The 13C-n-alkane dose can be adjusted to the original type of diet preferably consumed on pasture by wild herbivores [12], [21] or by using confined animals or captive zoo animals under controlled housing and dietary conditions.
n-Alkanes share the linear aliphatic hydrocarbon chain with fatty acids. As the latter pass through the reticulorumen with the particle phase [49], it has been suggested that also the flow characteristics of n-alkanes through the gastrointestinal tract might be alike [50]. Passage kinetics of n-alkanes through the reticulorumen might be therefore similar to that of the common long-chain C10–C18 fatty acids present in dietary lipids. A direct measurement of the fractional rumen passage rate of the latter is difficult because of the substantial transfer of carbon isotopes from the labeled dietary fatty acid into newly formed fatty acid compounds or microbial lipids. Knowledge on passage kinetics of n-alkanes might be therefore a useful indicator of rumen passage kinetics of dietary long-chain fatty acids bypassing the reticulorumen.
Incorporation of carbon isotopes into n-alkanes in the rumen was considered negligible [51], providing further evidence as to the potential use of isotope labeled n-alkanes to measure rumen digesta passage. An early in vivo study with dairy cows [10] suggested that rumen bacteria may incorporate some of the intra-ruminally dosed 14C-n-alkane C18, although no further metabolic process was observed for that specific labeled n-alkane [10], [52]. Yet, no evidence for microbial incorporation is available with regard to the more common natural long-chain forage n-alkanes C27 to C35 used in our study. A recent in vitro study, in which 14C labeled perennial ryegrass was incubated in buffered rumen fluid [11], suggested the complete absence of ruminal degradation and synthesis of long-chain n-alkanes by ruminal bacteria.
In contrast, dietary nutrients are often subjected to extensive fermentative degradation in the rumen. Isotopes originating from an isotopic labeled diet may be incorporated into rumen microbial protein and volatile fatty acids [53]. A bias in the prediction of K 1 can occur if tracer migration occurs; for instance, if stable isotopes from dietary nutrients are incorporated into microbial biomass [5], [54]. Furthermore, imperfect experimental in vivo conditions, such as an inhomogeneous isotope distribution in the diet, non-steady state conditions in the rumen or irregular nutrient uptake, might result in a different δ13C of the indigested nutrients relative to that of the original nutrients ingested by the animal, thereby affecting respective fractional passage rates. The overall satisfactory resemblance in passage kinetics of 13C-n-alkanes and dietary 13C-DM observed in this study supports earlier studies on the use of stable isotopes to measure rumen passage of dietary feed nutrients.
Conclusions
Passage kinetics of 13C labeled n-alkanes are rather comparable to that of the dietary dry matter originating from stable isotope labeled ryegrass plants but differs considerably from that of the external tracer Cr-NDF. In combination with evidence from literature as to the absence of microbial involvement in the passage of n-alkanes, our results suggest that stable isotopes are an appropriate tool to assess passage kinetics of n-alkanes and dietary nutrients through the gastrointestinal tract of ruminants.
Acknowledgments
The authors kindly acknowledge contributions of J.-B. Daniel and J.J.B.W. Leenaars as part of their undergraduate course at Wageningen University, Wageningen, The Netherlands.
Funding Statement
This work was co-financed by the Product Board Animal Feed (Den Haag, The Netherlands). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. No additional external funding received for this study.
References
- 1. Dijkstra J, Kebreab E, Mills JAN, Pellikaan WF, López S, et al. (2007) Predicting the profile of nutrients available for absorption: from nutrient requirement to animal response and environmental impact. Animal 1: 99–111. [DOI] [PubMed] [Google Scholar]
- 2. Poppi DP, Norton BW, Minson DJ, Hendricksen RE (1980) The validity of the critical size theory for particles leaving the rumen. J Agric Sci 94: 275–280. [Google Scholar]
- 3. Siciliano-Jones J, Murphy MR (1986) Passage of inert particles varying in length and specific gravity through the postruminal digestive tract of steers. J Dairy Sci 69: 2304–2311. [DOI] [PubMed] [Google Scholar]
- 4.Owens FN, Goetsch AL (1986) Digesta passage and microbial protein synthesis. In: Milligan LP, Grovum WL, Dobson A, editors. Control of Digestion and Metabolism in Ruminants. Englewood, NJ: Prentice-Hall. 196–223.
- 5. Pellikaan WF, Verstegen MWA, Tamminga S, Dijkstra J, Hendriks WH (2013) δ 13C as a marker to study digesta passage kinetics in ruminants; a combined in vivo and in vitro study. Animal 7: 754–767. [DOI] [PubMed] [Google Scholar]
- 6. Kebreab E, Mills JAN, Crompton LA, Bannink A, Dijkstra J, et al. (2004) An integrated mathematical model to evaluate nutrient partition in dairy cattle between the animal and its environment. Anim Feed Sci Technol 112: 131–154. [Google Scholar]
- 7. Dove H, Mayes RW (2006) Protocol for the analysis of n-alkanes and other plant-wax compounds and for their use as markers for quantifying the nutrient supply of large mammalian herbivores. Nature Prot 1: 1680–1697. [DOI] [PubMed] [Google Scholar]
- 8. Dove H, Mayes RW (1991) The use of plant wax alkanes as marker substances in studies of the nutrition of herbivores: a review. Austr J Agric Res 42: 913–952. [Google Scholar]
- 9. Ferreira LMM, Celaya R, García U, Rodrigues MAM, Osoro K (2009) Differences between domestic herbivores species in alkane faecal recoveries and the accuracy of subsequent estimates of diet composition. Anim Feed Sci Technol 151: 128–142. [Google Scholar]
- 10. Bartley EE, Helmer LG, Meyer RM (1971) Metabolism of 14C-labeled octadecane by cattle. J Anim Sci 33: 1351–1355. [DOI] [PubMed] [Google Scholar]
- 11. Keli A, Mayes RW, De Vega A (2008) In vitro studies of the metabolism of [14C]-n-alkanes using ruminal fluid of sheep as substrate. Animal 2: 1748–1752. [DOI] [PubMed] [Google Scholar]
- 12. Bezabih M, Pellikaan WF, Hendriks WH (2011) Using n-alkanes and their carbon isotope enrichments (δ13C) to estimate the botanical composition of pasture mixes from the Mid Rift Valley grasslands of Ethiopia. Livestock Sci 142: 298–304. [Google Scholar]
- 13. Südekum KH, Ziggers W, Roos N, Sick H, Tamminga S, et al. (1995) Estimating the passage of digesta in steers and wethers using the ratio of 13C to 12C and titanium(IV)-oxide. Isot Environ Health Stud 31: 219–227. [Google Scholar]
- 14. Udén P, Colucci PE, Van Soest PJ (1980) Investigation of chromium, cerium and cobalt as markers in digesta. Rate of passage studies. J Sci Food Agric 31: 625–632. [DOI] [PubMed] [Google Scholar]
- 15. Gorissen A, Kuikman PJ, Ginkel JH, Beek H, Jansen AG (1996) ESPAS – An advanced phytotron for measuring carbon dynamics in a whole plant-soil system. Plant Soil 179: 81–87. [Google Scholar]
- 16. Dove H, Mayes RW, Freer M (1996) Effects of species, plant part, and plant age on the n-alkane concentrations in the cuticular wax of pasture plants. Austr J Agric Res 47: 1333–1347. [Google Scholar]
- 17. Abrahamse PA, Dijkstra J, Vlaeminck B, Tamminga S (2008) Frequent allocation of rotationally grazed dairy cows changes grazing behavior and improves productivity. J Dairy Sci 91: 2033–2045. [DOI] [PubMed] [Google Scholar]
- 18. Abrahamse PA, Vlaeminck B, Tamminga S, Dijkstra J (2008) The effect of silage and concentrate type on intake behavior, rumen function, and milk production in dairy cows in early and late lactation. J Dairy Sci 91: 4778–4792. [DOI] [PubMed] [Google Scholar]
- 19. Mayes RW, Lamb CS, Colgrove PM (1986) The use of dosed and herbage n-alkanes as markers for the determination of herbage intake. J Agric Sci 107: 70. [Google Scholar]
- 20. Oliván M, Osoro K (1999) Effect of temperature on alkane extraction from faeces and herbage. J Agric Sci 132: 305–312. [Google Scholar]
- 21. Bezabih M, Pellikaan WF, Tolera A, Hendriks WH (2011) Evaluation of n-alkanes and their carbon isotope enrichments (δ13C) as diet composition markers. Animal 5: 57–66. [DOI] [PubMed] [Google Scholar]
- 22. Dhanoa MS, Siddons RC, France J, Gale DL (1985) A multicompartmental model to describe marker excretion patterns in ruminant faeces. Br J Nutr 53: 663–671. [DOI] [PubMed] [Google Scholar]
- 23. Sponheimer M, Robinson T, Roeder B, Hammer J, Ayliffe L, et al. (2003) Digestion and passage rates of grass hays by llamas, alpacas, goats, rabbits, and horses. Small Rumin Res 48: 149–154. [Google Scholar]
- 24. France J, Thornley JHM, Siddons RC, Dhanoa MS (1993) On incorporating diffusion and viscosity concepts into compartmental models for analysing faecal marker excretion patterns in ruminants. Br J Nutr 70: 369–369. [DOI] [PubMed] [Google Scholar]
- 25.Bibby J, Toutenburg H (1977) Prediction and improved estimation in linear models. Wiley New York.
- 26.Mayes RW, Giráldez FJ, Lamb CS (1997) Estimation of gastrointestinal passage rates of different plant components in ruminants using isotopically-labelled plant wax hydrocarbons or sprayed even-chain alkanes. Proc Nutr Soc 56: 187 A.
- 27. Ali HAM, Mayes RW, Hector BL, Verma AK, Ørskov ER (2005) The possible use of n-alkanes, long-chain fatty alcohols and long-chain fatty acids as markers in studies of the botanical composition of the diet of free-ranging herbivores. J Agric Sci 143: 85–95. [Google Scholar]
- 28. Ferreira LMM, Carvalho S, Falco V, Celaya R, Garcia U, et al. (2012) Assessment of very long-chain fatty acids as complementary or alternative natural fecal markers to n-alkanes for estimating diet composition of goats feeding on mixed diets J Anim Sci. 87: 2732–2745. [DOI] [PubMed] [Google Scholar]
- 29. Ippel JH, Pouvreau L, Kroef T, Gruppen H, Versteeg G, et al. (2004) In vivo uniform 15N-isotope labelling of plants: Using the greenhouse for structural proteomics. Proteomics 4: 226–234. [DOI] [PubMed] [Google Scholar]
- 30. Benner R, Fogel ML, Sprague EK, Hodson RE (1987) Depletion of 13C in lignin and its implications for stable carbon isotope studies. Nature 329: 708–710. [Google Scholar]
- 31. Scott MP, Jane J-L, Soundararajan M (1999) Carbon isotope ratios of amylose, amylopectin and mutant starches. Phytochemistry 52: 555–559. [Google Scholar]
- 32. Illius AW, Gordon IJ (1992) Modelling the nutritional ecology of ungulate herbivores: evolution of body size and competitive interactions. Oecologia 89: 428–434. [DOI] [PubMed] [Google Scholar]
- 33. Bruining M, Bosch MW (1992) Ruminal passage rate as affected by CrNDF particle size. Anim Feed Sci Technol 37: 193–200. [Google Scholar]
- 34. Ehle FR, Bas F, Barno B, Martin R, Leone F (1984) Particulate rumen turnover rate measurement as influenced by density of passage marker. J Dairy Sci 67: 2910–2913. [Google Scholar]
- 35.Sutherland TM (1988) Particle separation in the forestomachs of sheep. In: Dobson A, editor. Aspects of Digestive Physiology in Ruminants. Ithaca, NY: Cornell University Press. 43–73.
- 36. Owens FN, Hanson CF (1992) External and internal markers for appraising site and extent of digestion in ruminants. J Dairy Sci 75: 2605–2617. [DOI] [PubMed] [Google Scholar]
- 37. Giráldez FJ, López S, Lamb CS, Mayes RW (2006) The use of even-chain alkanes sprayed onto herbage as rate of passage markers in goats. Livestock Sci 100: 195–202. [Google Scholar]
- 38. Bosch MW, Bruining M (1995) Passage rate and total clearance rate from the rumen of cows fed on grass silages differing in cell-wall content. Br J Nutr 73: 41–49. [PubMed] [Google Scholar]
- 39. Bulang M, Elwert C, Spilke J, Rodehutscord M (2008) Suitability of synthetic alkanes as markers for the estimation of passage rate in sheep. Livestock Sci 115: 42–52. [Google Scholar]
- 40. Colucci PE, Macleod GK, Grovum WL, McMillan I, Barney DJ (1990) Digesta kinetics in sheep and cattle fed diets with different forage to concentrate ratios at high and low intakes. J Dairy Sci 73: 2143–2156. [DOI] [PubMed] [Google Scholar]
- 41.Cannas A, Van Soest P (2000) Simple allometric models to predict rumen feed passage. In: McNamara JP, France J, Beever DE, editors. Modelling Nutrient Utilization in Farm Animals. Wallingford, UK: CAB International. 49–62.
- 42.Ellis WC, Lascano C, Teeter R, Owens FN (1982) Solute and particulate flow markers. In: Owens FN, editor. Protein Requirements for Cattle. Stillwater, OK: Oklahoma State Univ. 37–56.
- 43. Huhtanen P, Hristov AN (2001) Estimating passage kinetics using fibre-bound 15N as an internal marker. Anim Feed Sci Technol 94: 29–41. [Google Scholar]
- 44. Warner D, Dijkstra J, Hendriks WH, Pellikaan WF (2013) Passage kinetics of 13C labeled corn silage components through the gastrointestinal tract of dairy cows. J Dairy Sci 96: 5844–5858. [DOI] [PubMed] [Google Scholar]
- 45. Clauss M, Lechner-Doll M (2001) Differences in selective reticulo-ruminal particle retention as a key factor in ruminant diversification. Oecologia 129: 321–327. [DOI] [PubMed] [Google Scholar]
- 46. Clauss M, Lechner-Doll M, Streich WJ (2003) Ruminant diversification as an adaptation to the physicomechanical characteristics of forage. Oikos 102: 253–262. [Google Scholar]
- 47. Hatt J-M, Lechner-Doll M, Mayes B (1998) The use of dosed and herbage n-alkanes as markers for the determination of digestive strategies of captive giraffes (Giraffa camelopardalis). Zoo Biol 17: 295–309. [Google Scholar]
- 48. Lechner-Doll M, Engelhardt Wv (1989) Particle size and passage from the forestomach in camels compared to cattle and sheep fed a similar diet. J Anim Physiol Anim Nutr 61: 120–128. [Google Scholar]
- 49.Dijkstra J, Gerrits WJJ, Bannink A, France J (2000) Modelling lipid metabolism in the rumen. In: McNamara JP, France J, Beever DE, editors. Modelling Nutrient Utilization in Farm Animals. Wallingford, UK: CAB International. 25–36.
- 50. Ohajuruka OA, Palmquist DL (1991) Evaluation of n-alkanes as digesta markers in dairy cows. J Anim Sci 69: 1726–1732. [DOI] [PubMed] [Google Scholar]
- 51.Mayes RW, Lamb CS, Colgrove PM (1988) Digestion and metabolism of dosed even-chain and herbage odd-chain n-alkanes in sheep. In: Flanagan S, editor. Proc 12th Gen Meeting European Grassland Federation. Dublin, Ireland. 159–163.
- 52. McCarthy RD (1964) Mammalian metabolism of straight-chain saturated hydrocarbons. Biochim Biophys Acta 84: 74–79. [DOI] [PubMed] [Google Scholar]
- 53. Alexander CL, Bartley EE, Meyer RM (1969) Incorporation of 14C from labeled alfalfa into rumen bacterial and volatile fatty acid carbon and rumen removal rate of fiber and soluble fractions. J Anim Sci 29: 948–958. [Google Scholar]
- 54. Firkins JL, Allen MS, Oldick BS, St-Pierre NR (1998) Modeling ruminal digestibility of carbohydrates and microbial protein flow to the duodenum. J Dairy Sci 81: 3350–3369. [DOI] [PubMed] [Google Scholar]