

Abstract
We studied 17 ABC1 genes in Populus trichocarpa, all of which contained an ABC1 domain consisting of about 120 amino acid residues. Most of the ABC1 gene products were located in the mitochondria or chloroplasts. All had a conserved VAVK-like motif and a DFG motif. Phylogenetic analysis grouped the genes into three subgroups. In addition, the chromosomal locations of the genes on the 19 Populus chromosomes were determined. Gene structure was studied through exon/intron organization and the MEME motif finder, while heatmap was used to study the expression diversity using EST libraries. According to the heatmap, PtrABC1P14 was highlighted because of the high expression in tension wood which related to secondary cell wall formation and cellulose synthesis, thus making a contribution to follow-up experiment in wood formation. Promoter cis-element analysis indicated that almost all of the ABC1 genes contained one or two cis-elements related to ABA signal transduction pathway and drought stress. Quantitative real-time PCR was carried out to evaluate the expression of all of the genes under abiotic stress conditions (ABA, CdCl2, high temperature, high salinity, and drought); the results showed that some of the genes were affected by these stresses and confirmed the results of promoter cis-element analysis.
1. Introduction
The protein kinase-like (PKL) superfamily, which can be divided into eukaryotic protein kinases (ePKs) and atypical protein kinases (aPKs), displays enormous variability in structure and sequence [1]. The ABC1 protein family is found widely in eukaryotes and prokaryotes and has been described as a new kind of putative kinase [2]. Before ABC1 genes were found in plants, they were identified in yeast, the first of which was named ABC1/COQ8 (hereafter called ScCOQ8). ScCOQ8 is a nuclear-encoded protein required for ubiquinone (UQ) synthesis in the mitochondria. This gene was found to be necessary for redox activity of the mitochondrial bc1 complex involved in cellular respiration, and was thus given the name abc1 because of its activity [3]. Loss of the ScCOQ8 gene causes a UQ deficiency and accumulation of the biosynthetic precursor 3-hexaprenyl-4-hydroxybenzoic acid, leading to instability of the bc1 complex and a lack of bc1 activity [4]. Missteps in the original analysis of ScCOQ8 gene function have caused confusion. The ScCOQ8 gene was initially believed to suppress a deleterious mutation in a cytochrome b translational activator (cbs2-223), leading to the incorrect conclusion that ScCOQ8p functions as a chaperone of cytochrome b [3, 5]. Not until a decade later was it found that ABC1 was the ScCOQ8 gene and that suppression of the translational activator mutant (cbs2-223) was due to a neighboring tRNATrp gene [4, 6].
Many studies have shown the localization of the ABC1 gene products in Arabidopsis, rice, and maize. Analysis of purified plastoglobules (PGs) from Arabidopsis chloroplasts identified six ABC1 kinase proteins (AtABC1P1, 3, 7) that localize predominantly to the plastid [7–9]. A seventh protein (AtABC1P8/OSA1) was shown to localize to the inner plastid envelope [10]. Furthermore, ABC1P13 is expected to localize to the mitochondria, based on the localization of its functional homolog in Baker's yeast (Saccharomyces cerevisiae) [11, 12]. Proteomic analysis of maize leaf fractions further supported the plastid localization of eight ABC1 kinase proteins, all of which were identified in maize proplastids, chloroplast fractions, or plastid nucleoids [13].
Jasinski et al. showed that the ABC1 proteins have the most conserved kinase motifs, including the VAIK catalytic motif (VAVK and VAMK) and the VAVK motif exists in the ABC1 domain, which also contains another motif, DFG [11, 14]. Many studies have also examined the ABC1 kinase protein domain. According to Lundquist et al. [15], a common ABC1 kinase domain is observed spanning approximately 350 residues and containing 12 conserved motifs based on an alignment of 100 full-length ABC1 kinase protein sequences from seven angiosperm species. Furthermore, eight of the ten key residues of the PKL superfamily identified in their study are present in the ABC1 kinase family and correspond to motifs III, IVa, IVb, VIIb, and VIII, which are involved in ATP binding and orientation (III, IVa, and IVb), catalysis (VIIb), and Mg2+ chelation (VIII) [2, 16–18]. The other seven motifs of the ABC1 kinase domain (I, II, V, VI, VIIa, IX, and X) do not have homologous sequences in ePKs, but can be found in several proteins outside of the PKL superfamily that have diverse enzyme activities.
A recent survey in Arabidopsis suggested that the chloroplast AtOSA1 protein, a homolog of the ABC1 proteins, was a new factor that plays a role in the balance of oxidative stress [11, 12]. There has also been research showing that ABC1 proteins are related to abiotic stress tolerance. Wang et al. showed that TaABC1, a member of the ABC1 protein kinase family in wheat, could enhance tolerance to drought, salt, and cold stress when overexpressed in Arabidopsis [19]. While research on the ABC1 protein family has been reported in Arabidopsis and rice, it has not been studied in Populus.
In this study, we identified 17 ABC1 genes in Populus trichocarpa and carried out phylogenetic and bioinformatic analysis using a number of databases and websites. We then constructed a chromosomal localization diagram using drawing tools based on transcription direction and plotting scale. Finally, we examined the relative expression levels of all 17 genes in response to various abiotic stresses by qRT-PCR using a four-step program to remove primer dimers. Our results showed the phylogenetic relationships and chromosomal locations of the ABC1 genes in P. trichocarpa. Nearly all of the genes contained some cis-elements involved in ABA and drought stress responses, which corresponded with the results of qRT-PCR analysis. Some of the genes had high expression levels in response to stress and might be important for subsequent studies.
2. Materials and Methods
2.1. Identification of ABC1 Genes
The Pfam (http://pfam.sanger.ac.uk/), Phytozome (http://www.phytozome.net/search.php), and NCBI (http://www.ncbi.nlm.nih.gov/) databases were searched for ABC1 protein and CDSs. The SMART database (http://smart.embl-heidelberg.de/) was used to analyze the ABC1 domain. WoLF PSORT (http://wolfpsort.org/) was used to predict the localization of the ABC1P gene products [22].
2.2. Phylogenetic Tree
Multiple sequence alignment was done using the ClustalX 1.83 software [23] and adjusted manually with BioEdit [24]. A phylogenetic tree was constructed using the neighbor-joining method with bootstrapping analysis in MEGA5 [25]. Due to the distant relationship between Populus and rice, maximum parsimony (MP) method was not suitable [26]. Since maximum likelihood (ML) method was tedious, it was also not an appropriate method. The neighbor-joining method was suitable for distant relationship plants and had a simple algorithm. Taken together, NJ method was a preferred tool to construct a phylogenetic tree. Rice protein sequences were obtained from the rice annotation database (http://rice.plantbiology.msu.edu/) based on the results of Yang et al. [14].
2.3. Chromosomal Localization
The chromosomal localization of all of the genes was determined with PopGenIE (http://www.popgenie.org/). A diagram of their locations on the 19 chromosomes was created with drawing tools on the basis of transcription direction and plotting scale.
2.4. Exon Intron Structure and Motif Analysis
Exon/intron structure was investigated using the Gene Structure Display Server (CSDS) (http://gsds.cbi.pku.edu.cn/chinese.php). The coding and genomic sequences were obtained from the NCBI database (http://www.ncbi.nlm.nih.gov/). Conserved motifs for each ABC1 gene were investigated using the Multiple Expectation Maximization for Motif Elucidation (MEME) system (Version 3.0 http://meme.sdsc.edu/meme/cgi-bin/meme.cgi).
2.5. Heatmap Analysis
To study the expression differences among different tissues in Populus, heatmap analysis was carried out using EST libraries representing gene models within PopulusDB [27, 28], which included 17 libraries that were derived from several taxa of Populus; aspen (Populus tremula), a hybrid aspen (P. tremula × tremuloides T89), and black cottonwood (http://popgenie.org/book/digital-northern).
2.6. Promoter Cis-Elements Identification and Analysis
Promoter sequences, the 2000 bp sequence upstream of the translational start site, of the genes were found in NCBI and analyzed in the Plant Care database (http://bioinformatics.psb.ugent.be/webtools/plantcare/ html/) which listed all of the cis-elements of these genes.
2.7. Plant Materials Treatment
Leaves of P. trichocarpa were treated with five abiotic stresses including ABA, CdCl2, NaCl, drought and high temperature. For ABA, CdCl2, and NaCl treatment, the leaves were steeped in 100 μM ABA, 10 mM CdCl2, or 200 μM NaCl. For drought treatment, the leaves were steeped in 4% PEG6000. All of these treatments were carried out for 4 or 8 h. For high temperature treatment, the leaves were incubated at 42°C for 0.5 or 1 h. Harvested leaves were frozen in liquid nitrogen immediately and stored at −80°C until RNA isolation.
2.8. RNA Isolation and Reverse Transcription
Total RNA was extracted from leaves that were treated under both control and stress conditions using the CTAB method. RNA integrity was verified by 2% agar gel electrophoresis. First-strand cDNA was synthesized using ReverTra Ace qPCR RT Master Mix with a gDNA Remover Kit (TOYOBO, Osaka, Japan), which eliminated the need for a DNA removal step. RNA samples were incubated at 65°C for 5 min. Then, 2 μL of 4× DN Master Mix (gDNA Remover) was added, and ddH2O was used to make the volume up to 8 μL. After incubation at 37°C for 5 min, 2 μL of 5× RT Master Mix II was added to give a total volume of 10 μL. Finally, the samples were incubated at 37°C for 15 min and 98°C for 5 min to obtain the first-strand cDNA. The reverse transcription products were used to carry out real-time quantitative PCR. Gene-specific primers for all 17 ABC1 genes were designed based on the CDSs using Primer Premier 5 with the following parameters: melting temperature 60–63°C, primer length 20–25 bp, and PCR product length 80–180 bp. All primer sequences are shown in Table 4.
Table 4.
Primers for the 17 ABC1 genes and Actin used in qRT-PCR. F represents forward primer while R represents reverse primer. All primers are listed in the 5′-3′ direction.
| Gene name | Primer (5′-3′) |
|---|---|
| PtrABC1P1 | F TTGAATTGGCTGAGACTGGATG R CACAGGCTTGGGAAAAGAAACA |
| PtrABC1P2 | F TTGAAGAGGTTCCTGTAGCATCTGG R GTCTAACCTTTACAGCAACCACCGT |
| PtrABC1P3 | F CGAGGTACATTTACGATCTGCGA R GCAACAAACTGTCCAGCTTTCAC |
| PtrABC1P4 | F TGACGGACAGCTCACCACAGTT R TGTTCTGGCACGAAGGGACTCT |
| PtrABC1P5 | F CGTAACTTCTTTGATGACGCACTC R AACAGCACCCAGACCATTCACTAG |
| PtrABC1P6 | F GGTGTTTACTGCATTTGCTACCG R TACCATCCCCAAATCGTACTGTG |
| PtrABC1P7 | F AAGGACCTTCCACAGGTTGTTG R GCTTTTCTCCCTCGATCCACTC |
| PtrABC1P8 | F CCGAAACAGTGAGGAGTAAAGTGC R CCAGATTAGACCAATACAGTCCCA |
| PtrABC1P9 | F TTCCAATTCCAAGCAGGAGATC R CCACAAGTTTGGGCAAGTTTTC |
| PtrABC1P10 | F TTCCATGCTGATCCTCATCCAG R TTGCCAAACCCAGAGAGTCACG |
| PtrABC1P11 | F GGACAAATGATACTGAAGAGCGG R CAAGCCTCAACTTATCTGGGAGA |
| PtrABC1P12 | F TCAGACAATGAGGGAGTCAATGCT R GAGCTACATGAACTTGTGCAAGGG |
| PtrABC1P13 | F TTGGGAATACACCACACCGC R AGGGTCGGCATGGAAGAATC |
| PtrABC1P14 | F AGAAAAAGAAGCAACGTCTGGC R CAACAAATGTGAAAGTGGCAGG |
| PtrABC1P15 | F GGATGAGCTTGCCAAGTTGC R GCTGGTCCTCAAACGCCTTA |
| PtrABC1P16 | F AGCAATAGTCTCCGTCACCGAT R ATTCTTTGGGCTTAGGAGGCGT |
| PtrABC1P17 | F CACAAGTCCATCGTGCGACATT R GGGCTGAAGTTATACTGCGGTT |
| Actin | F CATCAAAGCATCGGTGAGGTC R GTTGCCATCCAGGCTGTCC |
2.9. Real-Time Quantitative PCR
qRT-PCR was carried out using SYBR Premix Ex Taq II (TaKaRa, Dalian, China). Reactions were prepared in a total volume of 20 μL containing 10 μL of 2× SYBR Premix, 6 μL of ddH2O, 2 μL cDNA template, and 2 μL of F + R primers whose concentrations were 10 μM. Before the final experiment, a pretest was carried out to determine the optimal reaction condition. Usually, two-step method was applied to qRT-PCR. However, this method might produce more primer dimers. To solve this problem, we tried a four-step program and got the most precise results. The reactions were performed under the following conditions: an initial denaturation step of 95°C for 1 min, followed by a three-step thermal cycling profile of denaturation at 95°C for 5 s, primer annealing at 55°C for 30 s, and extension at 72°C for 30 s. After that, an additional step of 80°C for 1 s was carried out to remove primer dimers, followed by plate reading. These procedures were conducted for 45 cycles. To verify the specificity of the amplicon for each primer pair, a melting curve analysis was performed ranging from 55°C to 99°C with a temperature increase rate of 0.2°C/s, held for 1 s. The relative quantification method (2−ΔΔCT) was used to evaluate quantitative variation between replicates [29]. Three technical replicates were performed for each sample to ensure the accuracy of the results.
3. Results
According to the Pfam, Phytozome, and NCBI databases, we identified 17 ABC1 genes in Populus, which are shown in Table 1. WoLF PSORT was used to predict the localization of their gene products in the plant cell. Most of the ABC1 gene products were located in the mitochondria or chloroplasts. The SMART database (http://smart.embl-heidelberg.de) was used to search the domains of these genes and showed that all 17 ABC1 genes had an ABC1 domain that contained about 120 amino acid residues with a highly conserved core region. In addition, the ABC1 domain contained a VAVK-like motif (such as VAVK, VVIK, VAMK, or VVVK) and a DFG-like motif (such as DFG, DHG, or DVG) (Figure 1). Interestingly, these results coincided with those of Yang et al. [14].
Table 1.
ABC1 gene family in P. trichocarpa.
| Gene namea | Accession numberb | NCBI Locus IDc | Lengthd | Localizatione |
|---|---|---|---|---|
| PtrABC1P1 | POPTR_0004s03280 | XM_002305610.1 | 630 | chlo |
| PtrABC1P2 | POPTR_0011s04110 | XM_002316632.1 | 628 | chlo |
| PtrABC1P3 | POPTR_0004s07340 | XM_002305222.1 | 518 | chlo |
| PtrABC1P4 | POPTR_0002s06180 | XM_002300851.1 | 734 | nucl |
| PtrABC1P5 | POPTR_0002s00200 | XM_002301843.1 | 704 | chlo |
| PtrABC1P6 | POPTR_0010s22780 | XM_002316324.1 | 826 | chlo |
| PtrABC1P7 | POPTR_0001s14410 | XM_002297781.1 | 807 | mito |
| PtrABC1P8 | POPTR_0006s29050 | XM_002330776.1 | 737 | chlo |
| PtrABC1P9 | POPTR_0019s08950 | XM_002329202.1 | 543 | mito |
| PtrABC1P10 | POPTR_0018s03080 | XM_002324321.1 | 712 | mito |
| PtrABC1P11 | POPTR_0012s09520 | XM_002318076.1 | 471 | cyto |
| PtrABC1P12 | POPTR_0008s06780 | XM_002311182.1 | 538 | mito |
| PtrABC1P13 | POPTR_0007s06450 | XM_002310562.1 | 616 | chlo |
| PtrABC1P14 | POPTR_0005s08490 | XM_002307087.1 | 1235 | chlo |
| PtrABC1P15 | POPTR_0014s16660 | XM_002320491.1 | 719 | cyto |
| PtrABC1P16 | POPTR_0002s18630 | XM_002515113.1 | 616 | chlo |
| PtrABC1P17 | POPTR_0018s04280 | XM_002324089.1 | 242 | nucl |
aSystematic designation given to P. trichocarpa ABC1Ps.
bPhytozome locus ID of full-length cDNA sequence available at Phytozome (http://www.phytozome.net/search.php).
cAccession number of NCBI locus ID(http://www.ncbi.nlm).
dLength of predicted gene product (amino acids).
eLocalization of ABC1 proteins as predicted by WoLF PSORT (http://wolfpsort.org/).
Figure 1.

Conserved amino acid conservation sequences in the ABC1 domain of the ABC1P gene family in P. trichocarpa. The multiple alignment of the ABC1 domain was obtained with Clustal X. Fully and partially conserved (present in more than 50% of aligned sequences) residues are highlighted in black and gray, respectively. VAVK and DFG motifs are marked.
To evaluate the evolutionary relationships of the ABC1 gene family, we constructed a phylogenetic tree from P. trichocarpa and rice protein sequences using MEGA5, which was divided into three groups (groups I, II, and III) (Figure 2) in accordance with the results of Yang et al. [14]. In their study, the rice ABC1 genes were also divided into three groups on the basis of orthologs or paralogs, which indicated that most ABC1P genes in this family were not formed by duplication after the divergence of Arabidopsis and rice and might have expanded in a non-species-specific manner.
Figure 2.
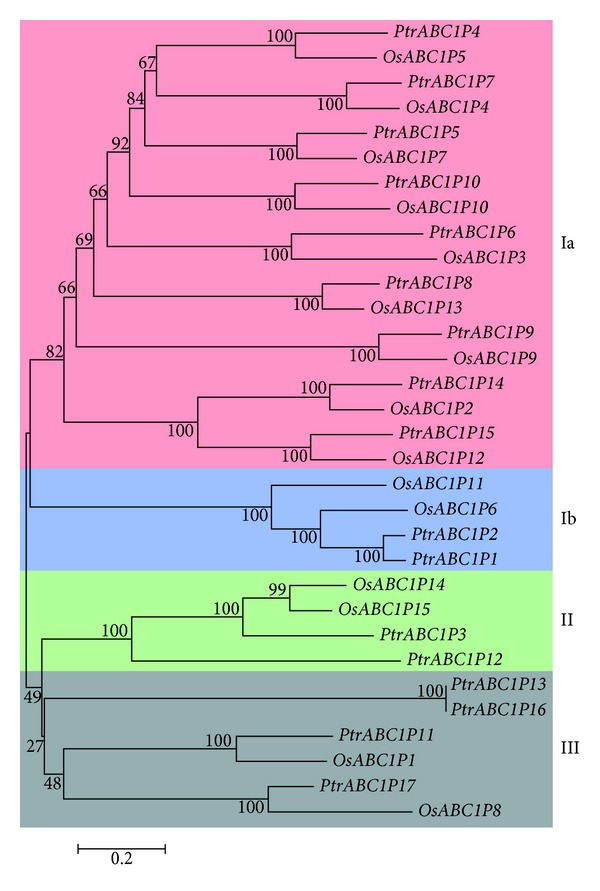
Phylogenetic tree of the ABC1 protein families in P. trichocarpa and rice. The tree was generated using the MEGA5 program with the neighbor-joining method. Pink represents group Ia, blue represents group Ib, green represents group II, and gray represents group III. OsABC1P represents ABC1 proteins in rice, while PtrABC1P represents ABC1 proteins in P. trichocarpa.
Chromosomal localization was determined using PopGenIE (http://www.popgenie.org/), and we created a diagram of the results using drawing tools according to transcription direction and plotting scale. This is shown in Figure 3. We found that no ABC1 genes were located on chromosomes 3, 6, 9, 13, 15, 16, or 17. Chromosome 2 had the largest number of PtrABC1 genes (three) followed by chromosomes 4 and 18 (two each). Only one PtrABC1 gene was found on chromosomes 1, 5, 6, 7, 8, 10, 11, 12, 14, and 19. No substantial clustering of Populus ABC1 genes was observed.
Figure 3.
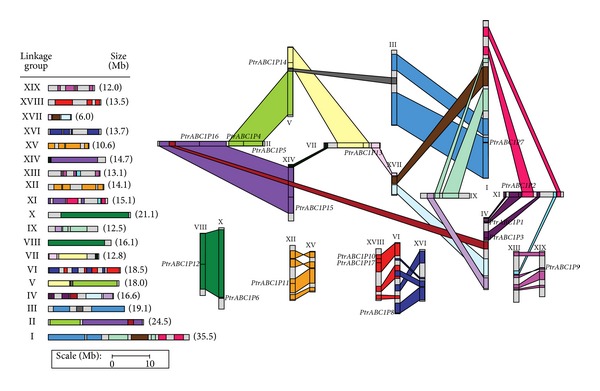
Genomic distribution of ABC1 genes on the P. trichocarpa chromosomes. A schematic view of chromosome reorganization by recent whole-genome duplication in Populus is shown (adapted from [20]). Regions that are assumed to correspond to homologous genome blocks are shaded in the same color and connected with lines.
A diagram of exon/intron structure and motifs is shown in Figure 4. To make the results clearer, we first constructed a phylogenetic tree using only the P. trichocarpa protein sequences, which were grouped into three subgroups in accordance with the phylogenetic tree mentioned above (Figure 4(a)). From the exon/intron structure diagram in Figure 4(b), we found that almost all of the genes had twelve exons but in different distribution ranges. However, PtrABC1P1 and PtrABC1P2, the members of group Ib, had roughly the same structure; both contained four exons that shared the same distribution. MEME motif detection revealed the diversification of the Populus ABC1 genes. The details of the 17 putative motifs identified are shown in Table 2. Figure 4(c) shows that genes in the same group share the same motifs and all of the genes contain motifs 1 and 7. This demonstrated the reliability of the phylogenetic classification.
Figure 4.
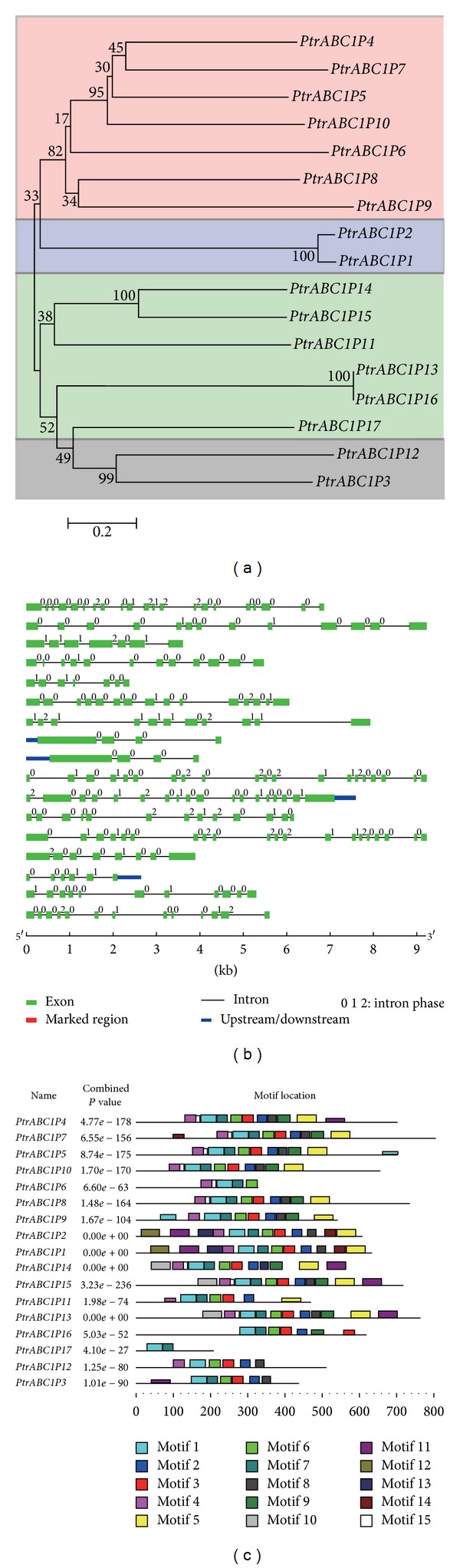
Phylogenetic analysis of the 17 P. trichocarpa ABC1 genes (a) with the integration of exon/intron structures (b) and motifs (c). Exon/intron structures were obtained from the Gene Structure Display Server. Motifs were investigated using the Multiple Expectation Maximization for Motif Elucidation (MEME) system.
Table 2.
Motif sequences of the P. trichocarpa ABC1 genes identified using MEME tools.
| Motif | Width | Best possible match |
|---|---|---|
| 1 | 41 | VEEELGAPVDDIFDQFDYEPIAAASLGQVHRARLKGQEVVI |
| 2 | 25 | AVESYLEQILSHGFFHADPHPGNIA |
| 3 | 29 | EYVKVPAIYWEYTTPQVLTMZYVPGIKIN |
| 4 | 29 | PTFVKLGQGLSTRPDICPPEYLEELAELQ |
| 5 | 50 | IGEDLLSIAADQPFRFPATFTFVVRAFSVLDGIGKGLDPRFDITEIAKPY |
| 6 | 29 | IYDECASVLYQEIDYTKEAANAELFASNF |
| 7 | 27 | KVQRPGLKDLFDIDLKNLRVIAEYLQK |
| 8 | 22 | GRLIFYDFGMMGSISPNIREGL |
| 9 | 33 | YGIYEKDPDKVLEAMIQMGVLVPTGDMTAVRRT |
| 10 | 50 | YSTIQRTLEIWGSVLTFIFKAWLNNQKFSYRGGMTEEKKMVRRKALAKWL |
| 11 | 50 | RQSRAFHNLFRQADRVQKLAETIQRLEQGDLKLRVRTLEAERAFQRVAAV |
| 12 | 48 | HGTVVSVCLHLPQFRNYSQYSFPSREYSSFALCNIKEQFGRRCFTRNY |
| 13 | 41 | AIYLAILFSPSMMMAPFADSCGPEFRKIWLHVVHRTLEKAG |
| 14 | 29 | QNCPNPKAFIEEVEESFTFWGTPEGDLVH |
| 15 | 11 | DQVPPFPSETA |
Publicly available Expressed Sequence Tags (ESTs) provided a useful tool to survey gene expression profiles in the form of a heatmap. We first carried out a preliminary analysis of ABC1 gene expression under various growth conditions and across different tissues by manual counting (Figure 5). Complete searching of the digital expression profiles from PopGenIE (http://www.popgenie.org/) yielded a total of 11 Populus ABC1 genes in the cDNA libraries. In the heatmap, most of the genes were represented by one or two ESTs in the cDNA libraries. PtrABC1P14 and PtrABC1P8 had notably high expression in senescing leaves. These expression profiles demonstrated that most of the ABC1 genes had a broad expression pattern across different tissues.
Figure 5.
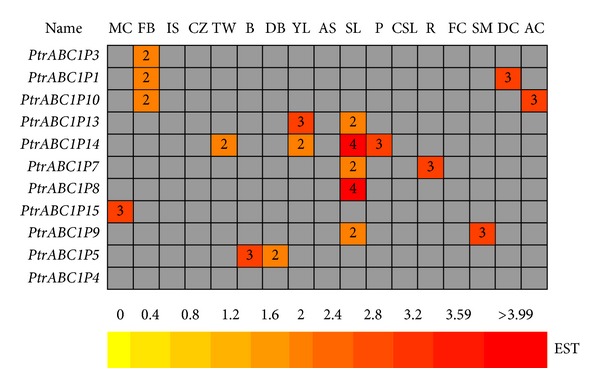
In silico EST analysis of P. trichocarpa ABC1 genes. The EST frequency for each gene was calculated by evaluating its EST representation among 17 cDNA libraries available at PopGenIE (http://www.popgenie.org/) [21]. The heatmap was visualized using the Heatmapper Plus tool by counting the corresponding ESTs for each particular gene in the database. The color bar at the bottom represents the frequencies of the EST counts. MC: male catkins, FB: flower buds, IS: imbibed seeds, CZ: cambial zone, TW: tension wood, B: bark, DB: dormant buds, YL: young leaves, AS: apical shoot, SL: senescing leaves, P: petioles, CSL: cold stressed leaves, R: roots, FC: female catkins, SM: shoot meristem, DC: dormant cambium, and AC: active cambium.
Promoter sequences (−2000 bp) were obtained from the NCBI database, and the cis-elements of these 17 genes were examined using the Plant Care database (Table 3). Many cis-elements were involved in abiotic stress responses such as the ABRE, MYB, MYC, and DRE core motifs and the LTRE motif. Most of the 17 genes contained ABRE and G-box motifs, both of which are related to the ABA signal transduction pathway. In addition, PtrABC1P1, 5, 14, and 15 also contained the W-box, which is found in the promoter regions of disease resistance-related genes and has been correlated with drought and ABA stress.
Table 3.
Cis-elements of the 17 ABC1 genes related to abiotic stresses such as drought and ABA. These cis-elements were obtained from the Plant Care database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/).
| Gene name | Cis-elements related to abiotic stress |
|---|---|
| PtrABC1P1 | G-box; W box |
| PtrABC1P2 | ABRE; G-box |
| PtrABC1P3 | ABRE; G-box |
| PtrABC1P4 | ABRE |
| PtrABC1P5 | G-box; W box |
| PtrABC1P6 | ABRE; G-box |
| PtrABC1P7 | ABRE; G-box |
| PtrABC1P8 | ABRE; G-box |
| PtrABC1P9 | ABRE; G-box |
| PtrABC1P10 | ABRE; G-box |
| PtrABC1P11 | G-box |
| PtrABC1P12 | G-box |
| PtrABC1P13 | ABRE; G-box |
| PtrABC1P14 | ABRE; G-box; W box |
| PtrABC1P15 | ABRE; G-box; W box |
| PtrABC1P16 | ABRE; G-box |
| PtrABC1P17 | None |
To study the relative expression levels of the 17 Populus ABC1 genes and select genes that had high relative expression levels, qRT-PCR was carried out using the SYBR Premix Ex Taq II kit. Figure 6 shows the results. According to the phylogenetic tree, we grouped these genes into three groups, which are shown in the figure. Most of the genes had higher expression under ABA stress after 8 h treatment compared with 4 h, except PtrABC1P17. Among these genes, the relative expression levels of PtrABC1P1 and PtrABC1P16 were notably high, though PtrABC1P1 was the highest. In the PEG treatment, we found that PtrABC1P10 and PtrABC1P16 had the highest expression. Interestingly, five genes (PtrABC1P8, 9, 10, 13, and 14) had higher expression after 4 h than after 8 h, while PtrABC1P16 presented the opposite tendency. The expression levels of PtrABC1P12 and PtrABC1P16 were high, with maximum relative expression values of 8, under Cd stress. The expression values were generally low under NaCl and high temperature (HT) stress. Only PtrABC1P7 and PtrABC1P13 showed high relative expression values (3 and 4, resp.) under NaCl treatment, while the expression values of PtrABC1P4, 6, and 16 were about 3 under HT treatment.
Figure 6.
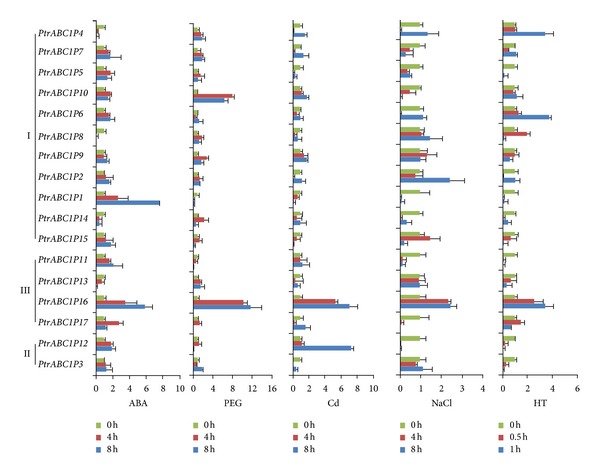
Relative expression of the 17 P. trichocarpa ABC1 genes. The five treatments investigated were ABA, PEG, Cd, NaCl, and HT (high temperature). I, II, and III represent the three groups shown in the phylogenetic tree in Figure 2. 0 h represents no treatment. 0.5, 1, 4, and 8 h represent the corresponding times under different treatments.
4. Discussion
According to Lundquist et al. [15], there are 23 predicted ABC1 genes in P. trichocarpa. However, the results of our modeling were a little different, and only 17 genes were found using the Pfam database. One of the reason for this might be that the other six genes are redundant or do not have ABC1 domains. In addition, in NCBI database, sequences of several genes were similar, and some of them were also incomplete. Taken together, only 17 genes were nonredundant and precise.
Previous data showed that the ABC1 proteins are involved in electron transfer in the respiratory chain and are located in mitochondria [6]. The first representative ABC1 gene in plants, AtABC1, is a structural and functional homolog of yeast ABC1, and it allows partial restoration of the complex III activity of a yeast abc1 mutant, suggesting the subcellular localization of AtABC1 in mitochondria [4, 12]. However, AtOSA1, another ABC1 gene in Arabidopsis, is located in chloroplasts and plays a role in the balance of oxidative stress [11]. In this study, most of the genes were located in mitochondria or chloroplasts. This further confirms that ABC1 family proteins contain the conserved ABC1 domain but have different kinase domains that lead to different localizations and presumably the diverse functions they perform.
Construction of a phylogenetic tree by Lundquist et al. from the ABC1 kinase proteins of 42 diverse species of Archaea, bacteria, and eukaryotes revealed a division into 15 subfamilies, which were named ABC1 K1 to ABC1 K15 [15]. On the other hand, the phylogenetic tree was divided into three clear primary clades characterized by evolutionary origins and subcellular localization. The first clade comprised eight subfamilies (1–8) and was specific for photosynthetic organisms. The second clade consisted of subfamilies 11–15, which are all likely targeted to the mitochondria based on localization predictions. Among these families, subfamily 13 was singled out as it corresponds with ScCOQ8P. Experimental evidence supported the mitochondrial location assignment of subfamily 13, because ScCOQ8P is localized to the inner mitochondrial envelope [3], and the UQ pathway is localized within the mitochondria. The third, central clade with subfamilies 9 and 11 contained the majority of the nonphotosynthetic bacterial ABC1Ps. This taxonomy placed PtrABC1 in the first clade, which was specific for photosynthetic organisms. In another study, Yang and Li constructed a phylogenetic tree for rice and Arabidopsis, which was divided into three groups on the basis of orthologs or paralogs. In accordance with their study, we also constructed a three-group phylogenetic tree using MEGA5. We used the 17 PtrABC1 genes in P. trichocarpa and 15 ABC1 genes in rice together to create the tree. The genes in groups Ia and Ib were orthologs (PtrABC1P1, 2, 4, 5, 6, 7, 8, 9, 10, 14, and 15), PtrABC1P3 and 12 were in group II, and group III contained PtrABC1P11, 13, 16, and 17.
Previous studies have revealed that the Populus genome has undergone at least three rounds of genome-wide duplication followed by multiple segmental duplication, tandem duplication, and transposition events such as retroposition and replicative transposition [30]. In particular, the segmental duplication associated with the salicoid duplication event that occurred 65 million years ago contributed remarkably to the expansion of many multigene families [31–36]. However, tandem duplication can easily occur in hotspots of chromosome recombination and has been closely correlated with the response of biotic and abiotic stress-related genes. In this study, about 82% (14 of 17) of the Populus ABC1 genes were located in duplicated regions, while only three genes (PtrABC1P5, 6, and 9) were located outside of any duplicated blocks. Among these 14 genes, six were preferentially retained duplicates that were located in both duplicated regions, while the others were only on one of the blocks and lacked duplicates on the corresponding block, suggesting that dynamic rearrangement might have occurred that led to the loss of some genes and might explain why we only found 17 PtrABC1 genes, while Lundquist et al. predicted 23. We draw attention to PtrABC1P1 and PtrABC1P2, as they were in the same branch in the phylogenetic tree and were also found in homologous regions on chromosomes 4 and 11, respectively. This discovery might explain the homology of the two genes.
The heatmap showed that PtrABC1P1, 3, and 10 had high expression in flower buds, with expression values of 2, probably indicating that these genes have a close relationship with flower differentiation and floral organ formation. PtrABC1P14 had high expression in young leaves, senescing leaves, and petioles, suggesting another relationship between this gene and leaves. In contrast, PtrABC1P4 had no expression in the 17 EST libraries. It might have expression in other libraries for which data was not available. For PtrABC1P14, it also highly expressed in TW (tension wood), which had a probability that this gene related to cellulose synthesis. Compared with herbaceous plant, P. trichocarpa had abundant wood as a woody plant. Wood is composed chiefly of cell walls, and the wall of a wood cell consists of compound middle lamella and secondary wall layers (S1, S2, and S3) composed of a cellulose/hemicellulose network impregnated with lignin. In many angiosperm trees, including Populus, the normal development of secondary wall biosynthesis is greatly modified when tension wood is formed as a gravitational response to stem movements caused, for example, by wind or load [37–39]. One of the main characteristics of tension wood, which distinguishes it from normal wood, is the formation of fibers with a thick inner gelatinous cell wall layer mainly composed of crystalline cellulose. Hence, tension wood is enriched in cellulose and deficient in lignin and hemicelluloses [40, 41]. In view of these factors and high expression in TW, PtrABC1P14 was speculated that it might relate to the secondary cell wall formation and cellulose synthesis in ABC1 gene family which might make a contribution to wood formation study.
Previous studies showed that ABC1 genes function differently in different plants. In S. cerevisiae, an ABC1 protein was shown to have chaperone-like activity and be essential for the proper conformation of cytochrome b complex III [6]. However, the ABC1 proteins in prokaryotes described as a new family of putative kinases [2] might be involved in the regulation of UQ biosynthesis [7]. Moreover, the homolog of the yeast ABC1 protein in Arabidopsis can partially restore respiratory complex deficiency when expressed in S. cerevisiae [12]. The chloroplast AtOSA1 (AtABC1P17) protein was identified as a new factor playing a role in the balance of oxidative stress. There was also a study that showed that ZmABC1-10 was significantly upregulated and stable under elevated Cd2+ concentrations [42]. Using transgenic plants, Wang et al. demonstrated that TaABC1 overexpression enhanced drought, salt, and cold stress tolerance in Arabidopsis and suggested that TaABC1 might act as a regulatory factor involved in multiple stress responses [19]. In this study, similar results were obtained. All 17 PtrABC1 genes, except for PtrABC1P17, had ABRE or G-box cis-elements, which play an important role in drought and ABA tolerance. This result was supported by qRT-PCR analysis. The qRT-PCR results showed that many genes were highly expressed under ABA and PEG treatment, such as PtrABC1P7, 10, 2, 15, 11, and 16. The expression of some genes under Cd stress also presented a similar tendency. Interestingly, PtrABC1P16 was highly expressed in all five treatments. However, members of the same group were only expressed under a subset of the conditions; PtrABC1P17 was only highly expressed under Cd stress, while PtrABC1P11 and 13 showed high expression under ABA and Cd stress and PEG, Cd, NaCl, and HT stress, respectively. To summarize, the ABC1Ps in Populus were differentially expressed in response to heat, ABA, PEG, NaCl, and CdCl2 stress, which implies that the PtrABC1Ps play different but important roles in response to abiotic stresses. These results might help us to select highly expressed genes that can lay a foundation for subsequent studies on the PtrABC1 genes in Populus.
5. Conclusions
The results of this study provide a phylogenetic and bioinformatic overview of 17 PtrABC1 genes in P. trichocarpa, most of which are localized to the mitochondria or chloroplasts. A phylogenetic tree showed the phylogeny and evolutionary relationships of the PtrABC1 genes, while heatmap analysis showed their expression levels in different tissues. Exon-intron structure and motif analyses were carried out to further investigate the phylogeny. Cis-elements contained in the PtrABC1 genes provide clues to their functions and expression under different abiotic stresses. From this, we should be able to select genes related to stress responses for further studies. However, the specific functions of the PtrABC1 genes remain unknown. Hence, further studies should be carried out to improve on the results of this study.
Conflict of Interests
The authors declare no conflict of interests.
Acknowledgments
This research was supported by the National High Technology Research and Development Program of China (2011AA100203), Fundamental Research Funds for the Central Universities (DL11EA02), and Program for Changjiang Scholars and Innovative Research Team in University (IRT1054).
References
- 1.Kannan N, Taylor SS, Zhai Y, Venter JC, Manning G. Structural and functional diversity of the microbial kinome. PLoS Biology. 2007;5(3):467–478. doi: 10.1371/journal.pbio.0050017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Leonard CJ, Aravind L, Koonin EV. Novel families of putative protein kinases in bacteria and archaea: evolution of the ‘eukaryotic’ protein kinase superfamily. Genome Research. 1998;8(10):1038–1047. doi: 10.1101/gr.8.10.1038. [DOI] [PubMed] [Google Scholar]
- 3.Bousquet I, Dujardin G, Slonimski PP. ABC1, a novel yeast nuclear gene has a dual function in mitochondria: it suppresses a cytochrome b mRNA translation defect and is essential for the electron transfer in the bc1 complex. EMBO Journal. 1991;10(8):2023–2031. doi: 10.1002/j.1460-2075.1991.tb07732.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Do TQ, Hsu AY, Jonassen T, Lee PT, Clarke CF. A defect in coenzyme Q biosynthesis is responsible for the respiratory deficiency in Saccharomyces cerevisiae abc1 mutants. Journal of Biological Chemistry. 2001;276(21):18161–18168. doi: 10.1074/jbc.M100952200. [DOI] [PubMed] [Google Scholar]
- 5.Brasseur G, Tron P, Dujardin G, Slonimski PP, Brivet-Chevillotte P. The nuclear ABC1 gene is essential for the correct conformation and functioning of the cytochrome bc1 complex and the neighbouring complexes II and IV in the mitochondrial respiratory chain. European Journal of Biochemistry. 1997;246(1):103–111. doi: 10.1111/j.1432-1033.1997.t01-1-00103.x. [DOI] [PubMed] [Google Scholar]
- 6.Hsieh EJ, Dinoso JB, Clarke CF. A tRNATRP gene mediates the suppression of cbs2-223 previously attributed to ABC1/COQ8 . Biochemical and Biophysical Research Communications. 2004;317(2):648–653. doi: 10.1016/j.bbrc.2004.03.096. [DOI] [PubMed] [Google Scholar]
- 7.Vidi P-A, Kanwischer M, Baginsky S, et al. Tocopherol cyclase (VTE1) localization and vitamin E accumulation in chloroplast plastoglobule lipoprotein particles. Journal of Biological Chemistry. 2006;281(16):11225–11234. doi: 10.1074/jbc.M511939200. [DOI] [PubMed] [Google Scholar]
- 8.Ytterberg AJ, Peltier J-B, Van Wijk KJ. Protein profiling of plastoglobules in chloroplasts and chromoplasts. A surprising site for differential accumulation of metabolic enzymes. Plant Physiology. 2006;140(3):984–997. doi: 10.1104/pp.105.076083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lundquist PK, Poliakov A, Bhuiyan NH, Zybailov B, Sun Q, van Wijk KJ. The functional network of the Arabidopsis plastoglobule proteome based on quantitative proteomics and genome-wide coexpression analysis. Plant Physiology. 2012;158(3):1172–1192. doi: 10.1104/pp.111.193144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jasinski M, Sudre D, Schansker G, et al. AtOSA1, a member of the ABC1-like family, as a new factor in cadmium and oxidative stress response. Plant Physiology. 2008;147(2):719–731. doi: 10.1104/pp.107.110247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Xie LX, Hsieh EJ, Watanabe S, et al. Expression of the human atypical kinase ADCK3 rescues coenzyme Q biosynthesis and phosphorylation of Coq polypeptides in yeast coq8 mutants. Biochimica et Biophysica Acta. 2011;1811(5):348–360. doi: 10.1016/j.bbalip.2011.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cardazzo B, Hamel P, Sakamoto W, Wintz H, Dujardin G. Isolation of an Arabidopsis thaliana cDNA by complementation of a yeast ABC1 deletion mutant deficient in complex III respiratory activity. Gene. 1998;221(1):117–125. doi: 10.1016/s0378-1119(98)00417-x. [DOI] [PubMed] [Google Scholar]
- 13.Majeran W, Friso G, Asakura Y, et al. Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: a new conceptual framework for nucleoid functions. Plant Physiology. 2012;158(1):156–189. doi: 10.1104/pp.111.188474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yang SG, Li T, Liu M, et al. Phylogenetic, structure and expression analysis of ABC1Ps gene family in rice. Biologia Plantarum. 2012;56(4):667–674. [Google Scholar]
- 15.Lundquist PK, Davis JI, van Wijk KJ. ABC1K atypical kinases in plants: filling the organellar kinase void. Trends in Plant Science. 2012;17(9):546–555. doi: 10.1016/j.tplants.2012.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hanks SK, Hunter T. The eukaryotic protein kinase superfamily: kinase (catalytic) domain structure and classification. FASEB Journal. 1995;9(8):576–596. [PubMed] [Google Scholar]
- 17.Taylor SS, Radzio-Andzelm E. Three protein kinase structures define a common motif. Structure. 1994;2(5):345–355. doi: 10.1016/s0969-2126(00)00036-8. [DOI] [PubMed] [Google Scholar]
- 18.Dissmeyer N, Schnittger A. The age of protein kinases. Methods in Molecular Biology. 2011;779:7–52. doi: 10.1007/978-1-61779-264-9_2. [DOI] [PubMed] [Google Scholar]
- 19.Wang C, Jing R, Mao X, Chang X, Li A. TaABC1, a member of the activity of bc1complex protein kinase family from common wheat, confers enhanced tolerance to abiotic stresses in Arabidopsis . Journal of Experimental Botany. 2011;62(3):1299–1311. doi: 10.1093/jxb/erq377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Douglas CJ, DiFazio SP. The Populus genome and comparative genomics. In: Jansson S, Bhalerao R, Groover A, editors. Genetics and Genomics of Populus. New York, NY, USA: Springer; 2010. pp. 67–90. [Google Scholar]
- 21.Sjödin A, Street NR, Sandberg G, Gustafsson P, Jansson S. The Populus Genome Integrative Explorer (PopGenIE): a new resource for exploring the Populus genome. New Phytologist. 2009;182(4):1013–1025. doi: 10.1111/j.1469-8137.2009.02807.x. [DOI] [PubMed] [Google Scholar]
- 22.Horton P, Park K-J, Obayashi T, et al. WoLF PSORT: protein localization predictor. Nucleic Acids Research. 2007;35:W585–W587. doi: 10.1093/nar/gkm259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The CLUSTAL X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research. 1997;25(24):4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series. 1999;41:95–98. [Google Scholar]
- 25.Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution. 1987;4(4):406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 26.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution. 2011;28(10):2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Segerman B, Jansson S, Karlsson J. Characterization of genes with tissue-specific differential expression patterns in Populus . Tree Genetics and Genomes. 2007;3(4):351–362. [Google Scholar]
- 28.Sterky F, Bhalerao RR, Unneberg P, et al. A Populus EST resource for plant functional genomics. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(38):13951–13956. doi: 10.1073/pnas.0401641101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang X, Wollenweber B, Jiang D, Liu F, Zhao J. Water deficits and heat shock effects on photosynthesis of a transgenic Arabidopsis thaliana constitutively expressing ABP9, a bZIP transcription factor. Journal of Experimental Botany. 2008;59(4):839–848. doi: 10.1093/jxb/erm364. [DOI] [PubMed] [Google Scholar]
- 30.Tuskan GA, DiFazio S, Jansson S, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray) Science. 2006;313(5793):1596–1604. doi: 10.1126/science.1128691. [DOI] [PubMed] [Google Scholar]
- 31.Wilkins O, Nahal H, Foong J, Provart NJ, Campbell MM. Expansion and diversification of the Populus R2R3-MYB family of transcription factors. Plant Physiology. 2009;149(2):981–993. doi: 10.1104/pp.108.132795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barakat A, Bagniewska-Zadworna A, Choi A, et al. The cinnamyl alcohol dehydrogenase gene family in Populus: phylogeny, organization, and expression. BMC Plant Biology. 2009;9, article 26 doi: 10.1186/1471-2229-9-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kalluri UC, Difazio SP, Brunner AM, Tuskan GA. Genome-wide analysis of Aux/IAA and ARF gene families in Populus trichocarpa . BMC Plant Biology. 2007;7, article 59 doi: 10.1186/1471-2229-7-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hu R, Qi G, Kong Y, Kong D, Gao Q, Zhou G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa . BMC Plant Biology. 2010;10(article 145) doi: 10.1186/1471-2229-10-145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Barakat A, Choi A, Yassin NBM, Park JS, Sun Z, Carlson JE. Comparative genomics and evolutionary analyses of the O-methyltransferase gene family in Populus . Gene. 2011;479(1-2):37–46. doi: 10.1016/j.gene.2011.02.008. [DOI] [PubMed] [Google Scholar]
- 36.Zhuang J, Cai B, Peng R-H, et al. Genome-wide analysis of the AP2/ERF gene family in Populus trichocarpa . Biochemical and Biophysical Research Communications. 2008;371(3):468–474. doi: 10.1016/j.bbrc.2008.04.087. [DOI] [PubMed] [Google Scholar]
- 37.Hellgren JM, Olofsson K, Sundberg B. Patterns of auxin distribution during gravitational induction of reaction wood in poplar and pine. Plant Physiology. 2004;135(1):212–220. doi: 10.1104/pp.104.038927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mellerowicz EJ, Baucher M, Sundberg B, Boerjan W. Unravelling cell wall formation in the woody dicot stem. Plant Molecular Biology. 2001;47(1-2):239–274. [PubMed] [Google Scholar]
- 39.Pilate G, Déjardin A, Laurans F, Leplé J-C. Tension wood as a model for functional genomics of wood formation. New Phytologist. 2004;164(1):63–72. doi: 10.1111/j.1469-8137.2004.01176.x. [DOI] [PubMed] [Google Scholar]
- 40.Fujita M, Saiki H, Harada H. Electron microscopy of microtubules and cellulose microfibrils in secondary wall formation of popular tension wood fibers. Mokuzai Gakkaishi. 1974;20:147–156. [Google Scholar]
- 41.Norberg PH, Meier H. Physical and chemical properties of the gelatinous layer in tension wood fibres of aspen (Populus tremula L.) Holzforschung. 1966;20(6):174–178. [Google Scholar]
- 42.Gao Q-S, Yang Z-F, Zhou Y, et al. Cloning of an ABC1-like gene ZmABC1-10 and its responses to cadmium and other abiotic stresses in maize (Zea mays L.) Acta Agronomica Sinica. 2010;36(12):2073–2083. [Google Scholar]