

Abstract
BACKGROUND AND PURPOSE
We evaluated the anti-emetic and anti-nausea properties of the acid precursor of Δ9-tetrahydrocannabinol (THC), tetrahydrocannabinolic acid (THCA), and determined its mechanism of action in these animal models.
EXPERIMENTAL APPROACH
We investigated the effect of THCA on lithium chloride- (LiCl) induced conditioned gaping (nausea-induced behaviour) to a flavour, and context (a model of anticipatory nausea) in rats, and on LiCl-induced vomiting in Suncus murinus. Furthermore, we investigated THCA's ability to induce hypothermia and suppress locomotion [rodent tasks to assess cannabinoid1 (CB1) receptor agonist-like activity], and measured plasma and brain THCA and THC levels. We also determined whether THCA's effect could be blocked by pretreatment with SR141716 (SR, a CB1 receptor antagonist).
KEY RESULTS
In rats, THCA (0.05 and/or 0.5 mg·kg−1) suppressed LiCl-induced conditioned gaping to a flavour and context; the latter effect blocked by the CB1 receptor antagonist, SR, but not by the 5-hydroxytryptamine-1A receptor antagonist, WAY100635. In S. murinus, THCA (0.05 and 0.5 mg·kg−1) reduced LiCl-induced vomiting, an effect that was reversed with SR. A comparatively low dose of THC (0.05 mg·kg−1) did not suppress conditioned gaping to a LiCl-paired flavour or context. THCA did not induce hypothermia or reduce locomotion, indicating non-CB1 agonist-like effects. THCA, but not THC was detected in plasma samples.
CONCLUSIONS AND IMPLICATIONS
THCA potently reduced conditioned gaping in rats and vomiting in S. murinus, effects that were blocked by SR. These data suggest that THCA may be a more potent alternative to THC in the treatment of nausea and vomiting.
Keywords: THCA, THC, conditioned gaping, vomiting, SR141716, CB1 receptor
Introduction
The cannabis plant is a source of at least 70 phytocannabinoids, including the psychoactive component Δ9-tetrahydrocannabinol (THC). THC is effective in interfering with nausea and vomiting in human cancer patients (see Cotter, 2013 for review). Comparisons of oral THC with the common anti-emetic agents of the time showed that THC was at least as effective (Frytak et al., 1979; Carey et al., 1983; Ungerleider et al., 1984; Crawford and Buckman, 1986; Cunningham et al., 1988; Tramer et al., 2001) if not more effective (Orr et al., 1980; Orr and McKernan, 1981) at reducing nausea and vomiting in human patients.
In animal models, THC reduces vomiting in ferrets (Van Sickle et al., 2001) and has been shown to suppress acute vomiting induced by cisplatin (Darmani, 2001b), SR141716 (SR), a cannabinoid 1 (CB1) receptor antagonist (Darmani, 2001a), radiation (Darmani et al., 2007) and 5-hydroxytryptophan (an indirect 5-HT receptor agonist, Darmani and Johnson, 2004) in Cryptotis parva (least shrews), a well-established animal model for assessing vomiting (see Darmani, 1998). In another animal model for assessing vomiting, Suncus murinus (house musk shrew), THC reduces cisplatin- (Kwiatkowska et al., 2004), LiCl- (Parker et al., 2004) and motion-induced vomiting (Cluny et al., 2008). Additionally, the administration of THC prior to reintroduction to a context previously associated with illness suppressed the expression of conditioned retching in shrews (Parker and Kemp, 2001; Parker et al., 2006).
Using the taste reactivity (TR) test, THC (0.5 mg·kg−1) has also been shown to interfere with the establishment and expression of conditioned gaping in rats produced by cyclophosphamide, a commonly used chemotherapy drug (Limebeer and Parker, 1999) and LiCl (Parker and Mechoulam, 2003; Parker et al., 2003). Unlike conditioned taste avoidance, which can be produced by both rewarding drugs and emetic drugs, conditioned gaping reactions are produced only by drugs that induce vomiting in emetic species, such as shrews (Parker, 2003; Parker et al., 2008). In a rodent model of anticipatory nausea (AN), THC (0.5 mg·kg−1) also reduces contextually elicited conditioned gaping in rats (Limebeer et al., 2006). The THC-induced suppression of nausea-induced behaviours and vomiting can be reversed by administration of the CB1 receptor antagonists SR or AM251, so it seems that the anti-emetic and anti-nausea effects of THC are mediated by the CB1 receptor (Darmani, 2001b; Parker et al., 2004; Darmani and Johnson, 2004; Darmani et al., 2007; Cluny et al., 2008).
THC is formed in cannabis from an acidic precursor, tetrahydrocannabinolic acid (THCA, Gaoni and Mechoulam, 1964). In the fresh plant, THCA is decarboxylated to THC by heating or burning. Interestingly, no psychotomimetic activity was observed with THCA administration to rhesus monkeys (doses ≤ 5 mg·kg−1, i.v.), mice (doses ≤ 20 mg·kg−1, i.p.) and dogs (doses ≤ 7 mg·kg−1; Grunfeld and Edery, 1969), perhaps making THCA a more desirable treatment than THC because it is devoid of psychoactive activity. No in vivo studies to date have evaluated THCA's mechanism of action, however in vitro, THCA's ability to inhibit the TNF-α levels in culture supernatants from U937 macrophages was not blocked by administration of the CB1 receptor antagonist AM281, or the cannabinoid 2 receptor (CB2) receptor antagonist AM630 (Verhoeckx et al., 2006). In addition, binding assays indicate that THCA is not active at the CB1 receptor (Ahmed et al., 2008).
The present investigation sought to explore whether THCA (0.5 and/or 0.05 mg·kg−1) could (i) inhibit LiCl-induced conditioned gaping to a flavour in rats; (ii) reduce LiCl-induced conditioned gaping to a context in rats, and whether this effect was CB1 or 5-hydroxytryptamine-1A (5-HT1A) receptor dependent; and (iii) reduce LiCl-induced vomiting in S. murinus, and whether this effect was CB1 receptor mediated. Plasma and brain samples from the AN study were also analysed for levels of THCA, as well as THC. Additionally, to determine if an equally low dose of THC could reduce nausea and AN in these models, we also added a low dose THC group (0.05 mg·kg−1) to each of our rat studies. To further assess THCA's CB1 receptor mechanism of action, we also examined whether THCA (0.5 mg·kg−1) exhibited any CB1 agonist-like behavioural activity, as defined in rodents by their characteristic action on a tetrad of tasks developed by Martin et al. (1991), including the hypothermia and locomotor activity assays.
Methods
Experimental procedures
Effect of THCA on LiCl-induced conditioned gaping to a flavour in rats
All rats were surgically implanted with an intraoral cannula under isofluorane anaesthesia according to the procedures described by Limebeer et al. (2010). Following recovery from surgery (at least 3 days), the rats received an adaptation trial in which they were placed in the TR chamber with their cannula attached to an infusion pump (Model KDS100, KD Scientific, Holliston, MA, USA) for fluid delivery. The TR chambers were made of clear Plexiglas (22.5 × 26 × 20 cm) that sat on a table with a clear glass top. A mirror beneath the chamber on a 45° angle facilitated viewing of the ventral surface of the rat to observe orofacial responses. Water was infused into their intraoral cannulae for 2 min at the rate of 1 mL·min−1. On the day following the adaptation trial, the rats received a conditioning trial in which they were administered a pretreatment injection of THCA (0.05, 0.5 mg·kg−1), THC (0.5 mg·kg−1) or VEH [ethanol/Cremophor/saline (SAL), 1:1:18]. Thirty minutes after the pretreatment injection, the rats were individually placed in the chamber and intraorally infused with 0.1% saccharin solution for 2 min at the rate of 1 mL·min−1 while the orofacial responses were video recorded from a mirror at a 45° angle beneath the chambers, with the feed from the video camera (Sony DCR-HC48, Henry's Cameras, Waterloo, ON, Canada) into a computer via FireWire connection. Immediately after the saccharin infusion, all rats were injected with 20 mL·kg−1 of 0.15 M LiCl and returned to their home cage. The groups were as follows: VEH (n = 8), 0.05 mg·kg−1 THCA (n = 8), 0.5 mg·kg−1 THCA (n = 7), 0.05 mg·kg−1 THC (n = 8). Seventy-two hours later, rats were tested drug free. Rats were again intraorally infused with 0.1% saccharin solution for 2 min at the rate of 1 mL·min−1 while the orofacial responses were video recorded. Rats were then returned to their home cages. The videotapes were later scored (at 1/2 speed) by an observer blind to the experimental conditions using ‘The Observer’ (Noldus Information Technology Inc., Leesburg, VA, USA) for the behaviour of gaping (large openings of the mouth and jaw, with lower incisors exposed). Conditioned taste avoidance was assessed in a single bottle test. Rats were water-restricted at 4:00 pm. The following morning, a single bottle containing 0.1% saccharin was placed on the cage at 9:00 am. Measures of saccharin consumption were taken at 30, 120 and 360 min. The number of gapes during the TR test trial was entered into a one-way anova. The amount of saccharin consumed during the conditioned taste avoidance test for each group was entered into a 4 × 3 mixed factors one-way anova. For all analyses, P-values of <0.05 were taken as significant.
Assessment of CB1 agonist-like behavioural activity
To assess whether THCA induced hypothermia, in a separate group of rats, rectal temperatures were monitored using a rectal digital thermometer (Model # 2483, Life Brand; Shoppers Drug Mart Inc., Guelph, ON, Canada). Rats were placed on a towel and mildly restrained with the tail lifted. The probe tip was covered with petroleum jelly and inserted 5 cm into the rat's rectum and held in place for 30 s. A baseline temperature was established, consisting of the average of two baseline measures, taken 15 min apart. Rats were then pretreated with either VEH or THCA (0.5 mg·kg−1, n = 5/group) and temperatures were taken at 30, 45, 60, 75 and 90 min after pretreatment. The temperatures recorded for each group were entered into a 2 × 6 mixed factors one-way anova. P-values of <0.05 were taken as significant.
To assess whether THCA reduced spontaneous motor activity, a separate group of rats were pretreated with either VEH or THCA (0.5 mg·kg−1, n = 8/group) 30 min prior to being placed in a novel activity chamber. The activity chamber was constructed of white Plexiglass with the dimensions of 60 cm × 25 cm × 25 cm illuminated with a red light. A video camera mounted on an extension pole captured the activity of the rat, which was sent to a computer for analysis of distance (cm) travelled using the Ethovision software program (Noldus Information Technology Inc., Leesburg, VA, USA). The distance (cm) travelled in the locomotor activity test was entered into an independent-samples t-test. P-values of <0.05 were taken as significant.
Wiley and Martin (2003) have, however, indicated that because other drug classes (such as dopamine antagonists) are also capable of producing cannabinoid-like effects on some of the tetrad tasks, CB1 agonist activity should be verified by blocking the behavioral effects using a selective CB1 antagonist/inverse agonist such as SR (Rinaldi-Carmona et al., 1994). Therefore, we went on to further investigate THCA's effect on LiCl-induced conditioned gaping to a context, as well as the mechanism of action, by examining whether SR or WAY100635 (WAY) could reverse THCA's effect.
Effect of SR or WAY pretreatment with THCA on LiCl-induced conditioned gaping to a context in rats
The distinctive context utilized for conditioning used location, visual, and tactile cues different from those in the home cage environment. The room was dark with two 25 Watt lights beside the conditioning chamber. The conditioning apparatus was identical to that used in the LiCl-induced conditioned gaping experiments, except that the chamber was made of black opaque Plexiglas. The rats received four conditioning trials, during which the contextual chamber was paired with 127 mg·kg−1 LiCl. In each conditioning trial, each rat was injected with LiCl and immediately placed in the distinctive context for a 30 min period. This procedure was followed for a total of four conditioning trials, with 72 h between each trial. On the test trial, rats were given a pretreatment of SR (2.5 mg·kg−1), WAY (0.1 mg·kg−1), or VEH to investigate THCA's mechanism of action. Fifteen minutes later, rats received a second pretreatment of VEH, THCA (0.05 mg·kg−1) or THC (0.05 mg·kg−1). Thirty minutes later, rats were taken individually to the conditioning context, given a SAL injection (20 mL·kg−1) and placed in the chamber for 5 min while their orofacial responses were video-recorded from a mirror beneath the chamber. The groups were as follows: VEH-VEH (n = 7), VEH-THCA (n = 8), SR-THCA (n = 8), SR-VEH (n = 8), WAY-THCA (n = 7), WAY-VEH (n = 7), VEH-THC (n = 8). The number of gapes during the test trial was analysed using a one-way anova. P-values of <0.05 were taken as significant.
Immediately following the AN test, the rats in groups VEH-VEH and VEH-THCA were sacrificed and the trunk blood was collected and prepared and separated (under refrigeration) into clean tubes. Plasma samples was stored at −20°C until they were sent in dry ice to Quotient Bioresearch (Fordham, Cambridgeshire, UK) for analysis of both THCA and THC levels present.
Effect of THCA on LiCl-induced vomiting in S. murinus.
This experiment evaluated the effect of pretreatment with THCA on vomiting induced by LiCl in S. murinus. The shrews were moved into the experimental room from the colony room and given four meal worms in an empty cage 15 min prior to receiving their pretreatment injection. Shrews were injected with THCA (0.05, n = 10 or 0.5 mg·kg−1, n = 8) or VEH (ethanol/Cremophor/SAL, 1:1:18, n = 10), followed 30 min later by an injection of LiCl (390 mg·kg−1). An additional group to assess the CB1 receptor mediation of THCA's effect was given SR (2.5 mg·kg−1, n = 6) 15 min prior to THCA (0.05 mg·kg−1). They were then immediately placed, individually in the chamber and observed for 45 min. The Plexiglas observation chambers (22.5 × 26 × 20 cm) sat on a table with a clear glass top. A mirror beneath the chamber on a 45° angle facilitated viewing of the ventral surface of the shrew to observe vomiting episodes. An observer blind to experimental conditions counted the frequency of vomiting episodes. Data were expressed as the mean ± SEM of vomiting responses (n = 6–10) and analysed using anova. P-values of <0.05 were taken as significant.
Animals
Animal procedures complied with the Canadian Council on Animal Care. The protocols were approved by the Institutional Animal Care Committee, which is accredited by the Canadian Council on Animal Care. The authors also followed the ARRIVE guidelines of the British Journal of Pharmacology, for reporting experiments involving animals (Kilkenny et al., 2010; McGrath et al., 2010). Naïve male Sprague-Dawley rats, weighing between 230 and 311 g on the day of conditioning, obtained from Charles River Laboratories (St Constant, Quebec, Canada) were used for assessment of anti-nausea-like behaviour. They were single-housed (except in the assessment of conditioned gaping to a context, for which they were pair-housed) in shoebox cages in a colony room at an ambient temperature of 21°C with a 12/12 h light-dark schedule (lights off at 8:00 am) and maintained on food and water ad libitum. LiCl-induced emesis experiments were performed with male (36.3–47.4 g) and female (21.6–30.2 g) S. murinus, ranging from 98 to 814 days of age at time of test, bred and raised in the University of Guelph colony. They were single-housed in cages in a colony room at an ambient temperature of 22°C on a 10/14 hlight-dark schedule (lights off at 7:00 pm). Shrews were tested during their light cycle.
Drugs and materials
Samples of THCA and THC, extracted from cannabis, were provided by GW Pharmaceuticals (Porton Down, Wiltshire, UK). Ethanol, Cremophor and LiCl were provided by Sigma (St Louis, MO, USA). THCA and THC were prepared in a vehicle (VEH) consisting of a 1:1:18 mixture of ethanol, Cremophor and SAL and was administered i.p. in a volume of 2 mL·kg−1. The initial dose for THCA (0.5 mg·kg−1) was selected based on the effective doses of THC in our models (Parker and Kemp, 2001; Parker and Mechoulam, 2003; Parker et al., 2003; 2006) and the selection of a lower dose of THCA was based on the finding that CBD's acidic precursor was far more potent than CBD in our models (Bolognini et al., 2013; Rock and Parker, 2013). LiCl was prepared as a 0.15 M solution with sterile water and administered i.p. in a volume of 60 mL·kg−1 (390 mg·kg−1) to shrews and in a volume of 20 mL·kg−1 (127.2 mg·kg−1) to rats. SR, 2.5 mg·kg−1 – a dose that on its own does not potentiate conditioned gaping or vomiting over that of VEH (Rock et al., 2008; Parker et al., 2009) – was prepared in a VEH consisting of a 1:1:18 mixture of ethanol, Tween and SAL and administered i.p. in a volume of 1 mL·kg−1 in rats and shrews. WAY (0.1 mg·kg−1, Rock et al., 2012) was mixed in SAL and administered i.p. in a volume of 1 mL·kg−1.
Results
THCA suppresses LiCl-induced conditioned gaping to a flavour in rats
We investigated whether THCA could attenuate LiCl-induced gaping (Figure 1A). A one-way anova revealed a significant effect of pretreatment, F(3, 27) = 4.4; P < 0.02. Least significant difference (LSD) post hoc comparisons revealed that both 0.05 and 0.5 mg·kg−1 THCA significantly (P-values < 0.05) reduced LiCl-induced gaping relative to the VEH-pretreated controls. Group THC (0.05 mg·kg−1) did not differ from the VEH-pretreated controls (P > 0.05). It is unlikely that THCA attenuated conditioned gaping in rats through a direct effect on learning because none of the gape-attenuating doses of THCA (0.05 and 0.5 mg·kg−1) or the low dose of THC (0.05 mg·kg−1) interfered with LiCl-induced conditioned taste avoidance. The mean amounts of saccharin consumed during the conditioned taste avoidance test at 30, 120 and 360 min by groups of rats treated with VEH, THCA (0.05 or 0.5 mg·kg−1) or THC (0.05 mg·kg−1) are presented in Figure 1B. A 4 × 3 mixed factors anova revealed only a significant effect of time of test, F(2,54) = 309.1, P < 0.001.
Figure 1.
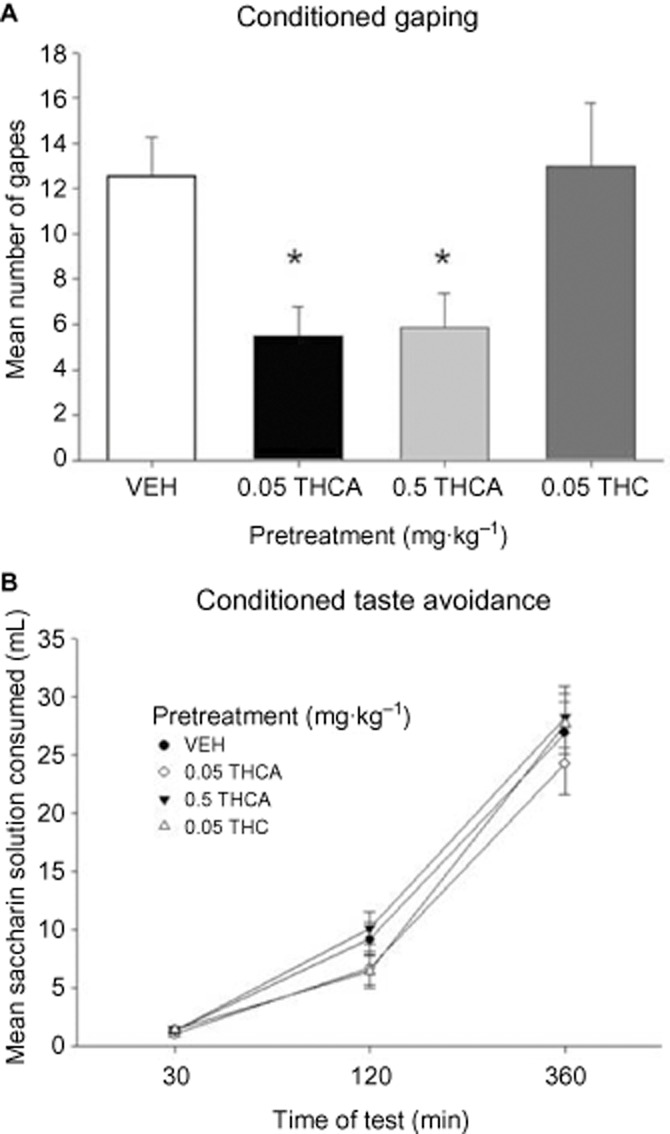
Effect of THCA (0.05, 0.5 mg·kg−1), THC (0.05 mg·kg−1) or VEH (n = 7–8) administered i.p. 30 min prior to LiCl. The mean number of conditioned gaping responses elicited by the LiCl-paired saccharin solution was measured during the test trial (A). Each bar represents the mean ± SEM (n = 7–8). The asterisks indicate a significant difference from the VEH-treated control animals (*P < 0.05). The mean cumulative amount of saccharin solution consumed (mL ± SEM) during a one-bottle consumption test was measured at 30, 120 and 360 min after introduction of the bottle to fluid-restricted rats (B).
THCA does not induce the CB1-agonist-like effects of hypothermia or reduced locomotor activity
We investigated whether THCA (0.5 mg·kg−1) could induce hypothermia, as a measure of CB1-agonist-like activity. The core temperatures measured at baseline, 30, 45, 60, 75 and 90 min after pretreatment are presented in Figure 2. A 2 × 6 mixed factors anova revealed no significant main effects or interaction (Ps > 0.05).
Figure 2.
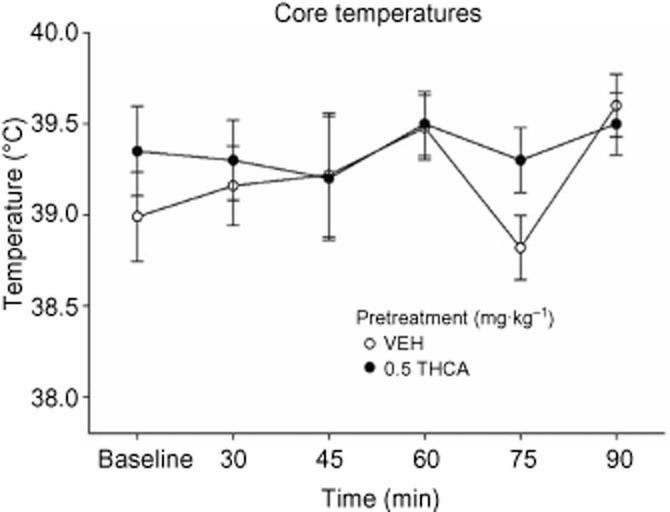
Effect THCA (0.5 mg·kg−1) or VEH (n = 5 per group) administered i.p. on core temperatures, relative to baseline, measured at 30, 45, 60, 75 and 90 min after pretreatment.
THCA (0.5 mg·kg−1) also did not alter locomotor activity, relative to VEH-pretreated controls during the 15 min activity test (Figure 3). A one-way anova revealed no significant differences in distance moved, t(14) = −0.8, P > 0.05.
Figure 3.
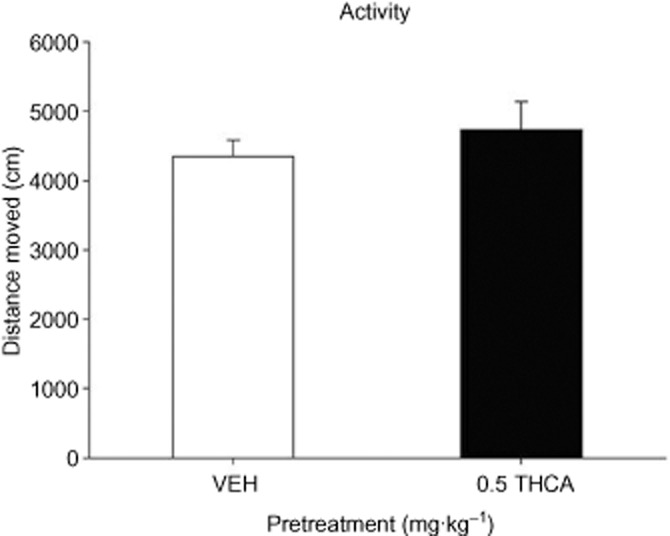
Effect of THCA (0.5 mg·kg−1) or VEH (n = 8 per group) administered i.p. 30 min prior to placement in the activity chamber. The distance moved (cm) was measured for 15 min. Each bar represents the mean ± SEM.
THCA suppresses LiCl-induced conditioned gaping to a context in rats: a CB1 receptor mechanism of action
THCA (0.05 mg·kg−1) interfered with the nausea-inducing effects of the context previously paired with LiCl, and SR, but not WAY, blocked this effect. The mean number of gapes during the test trial is presented in Figure 4. A one-way anova revealed a main effect of pretreatment group, F(6, 46) = 4.7, P = 0.001. LSD post hoc comparisons revealed that pretreatment group VEH-THCA gaped significantly less than VEH controls (P = 0.001). The suppressed gaping by THCA was reversed in Group SR-THCA (P < 0.001), but not in group WAY-THCA. Groups VEH-THC, SR-THCA, WAY-VEH, SR-VEH did not differ from VEH-VEH-pretreated controls (P > 0.05).
Figure 4.
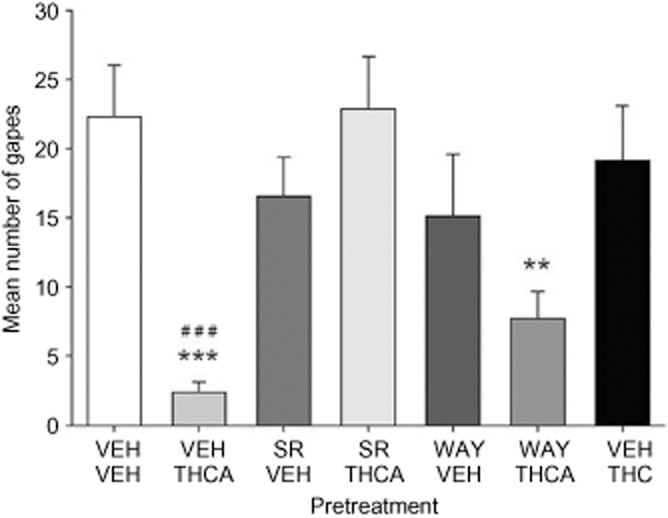
Effect of VEH, SR (2.5 mg·kg−1, i.p.) or WAY (0.1 mg·kg−1, i.p.) on the THCA- (0.05 mg·kg−1, i.p.) induced suppression on the expression of gaping to a context previously paired with LiCl (a model of AN). An additional group was administered VEH-THC (0.05 mg·kg−1, i.p.). Each bar represents the mean number of conditioned gaping responses ± SEM (n = 7–8). The asterisks indicate a significant difference from the VEH-VEH-treated control animals (**P < 0.025, ***P = 0.001). The pound symbols represent a significant difference from the VEH-0.05 THCA group (###P < 0.001).
Analysis of plasma THCA levels
In the VEH-THCA-pretreated rats, THCA, but not THC, was detected in the plasma (mean = 16.1 ng·mL−1, SD = 10.3), ranging in concentration from 3.92–32.5 ng·mL−1. In the VEH-VEH-pretreated controls, neither THCA nor THC was detected in plasma samples. These results indicate that THCA's effects on AN are unlikely to be due to in vivo conversion of THCA to THC.
THCA reduces LiCl-induced vomiting in S. murinus: a CB1 receptor-mediated effect
We finally determined whether THCA could also reduce LiCl-induced emesis in shrews, and whether this effect was CB1 receptor mediated. We found that THCA does indeed inhibit LiCl-induced emesis. In Figure 5, a one-way anova revealed a significant difference among groups, F(3,29) = 18.2, P < 0.001; Bonferroni post hoc tests revealed that those shrews given LiCl after pretreatment with both doses of THCA vomited significantly less than VEH-pretreated controls (Ps < 0.001) The suppressive effect on emesis appeared to be CB1 receptor mediated because shrews in the pretreatment group SR-0.05 THCA vomited significantly more than those in group 0.05 THCA (P = 0.007). When administered by themselves, neither THCA, VEH nor SR produced emesis during the 45 min pretreatment period before toxin administration (data not shown).
Figure 5.
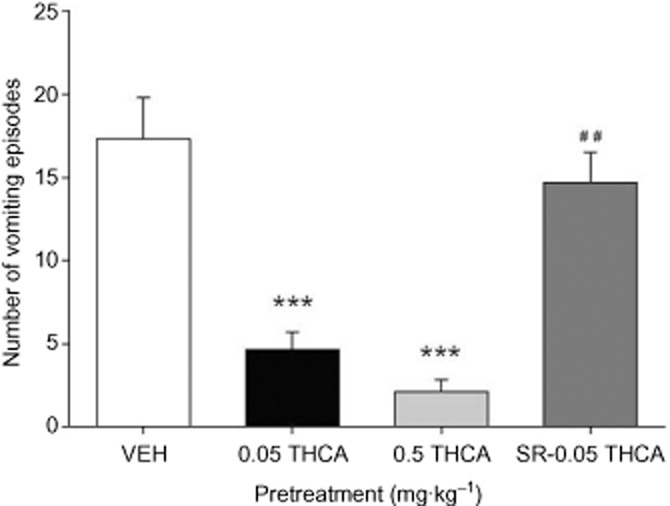
Effect of THCA (0.05, 0.5 mg·kg−1) or VEH administered i.p. 30 min prior to LiCl administration. An additional group was also given SR (2.5 mg·kg−1, i.p.) 15 min prior to THCA (0.05 mg·kg−1). The number of emetic episodes in shrews treated with LiCl was measured. Each bar represents the mean ± SEM (n = 6–10). The asterisk indicates a significant difference from the VEH-treated control animals (***P < 0.001). The pound symbols represent a significant difference from the 0.05 THCA group (##P < 0.01).
Discussion
THCA (0.5 and 0.05 mg·kg−1) reduced LiCl-induced conditioned gaping to a flavour and THCA (0.05 mg·kg−1) also reduced conditioned gaping to a context previously paired with LiCl. SR reversed this latter effect (while having no effect on its own), but WAY did not, indicating that THCA's suppressive effects on contextually elicited conditioned gaping may be CB1 receptor mediated. Furthermore, THCA (0.5 and 0.05 mg·kg−1) was also effective in reducing LiCl-induced vomiting in shrews. The suppressive effect of 0.05 mg·kg−1 THCA was also blocked by SR administration, lending further support to a CB1 receptor-mediated mechanism of action.
Although our in vivo results suggest a CB1 receptor-mediated mechanism for THCA, the few in vitro studies that have been published to date do not seem to support this finding. THCA has been shown to inhibit TNF-α (a pro-inflammatory cytokine), demonstrating its ability to reduce the inflammatory response, albeit through a mechanism independent of CB1 or CB2 receptors (Verhoeckx et al., 2006). Binding assays also suggest that THCA is not active at CB1 receptors (Ahmed et al., 2008). Likewise, N-arachidonoyl-L-serine, a novel endocannabinoid, demonstrated neuroprotective effects, which were blocked in vivo by the CB2 receptor antagonist SR144528, demonstrating a CB2 mechanism of action (Cohen-Yeshurun et al., 2011; 2013); however, it has been shown to bind very weakly to this receptor in vitro (Milman et al., 2006). Therefore, further studies need to be conducted to further elucidate THCA's mechanism of action.
We have previously reported that at a dose as low as 0.5 mg·kg−1 ip, THC reduces both the establishment of LiCl-induced conditioned gaping to a flavour (Parker and Mechoulam, 2003; Parker et al., 2003) and the expression of LiCl-induced conditioned gaping to a context (Limebeer et al., 2006). Here, we found that at a lower dose of 0.05 mg·kg−1, THC did not reduce these behaviours reflective of nausea, but at that dose, THCA was effective. Therefore, THCA appears a more potent anti-nausea treatment than THC, even though it is not psychoactive. Additionally, it is unlikely that the anti-nausea effects of THCA in our models could be due to the metabolism of THCA into THC, given that the equivalent dose of THC is ineffective. Furthermore, when plasma samples were analysed, no detectable levels of THC were present, only THCA. This evidence discounts the possibility that the effects were due to conversion of THCA to THC.
Measuring these plasma levels does not, however, rule out the possibility that THCA is modified in vivo to produce another metabolite that has CB1 agonist-like properties. We attempted to assess this possibility by measuring core temperatures and locomotor activity in response to VEH or THCA pretreatments. If the THCA effects seen here were due to its in vivo conversion to a metabolite with CB1 agonist-like properties, then we would expect to see a hypothermic response and reduced activity (characteristic of CB1 agonist-like activity as measured in the tetrad of tasks). Relative to VEH-pretreated controls, there was no significant change in rectal temperatures or activity as a result of THCA pretreatment. Because we saw no such changes, it seems unlikely that these effects can be explained by THCA's conversion to a metabolite with CB1 agonist-like properties.
In contrast to LiCl-induced conditioned gaping reactions, THCA and THC pretreatment did not interfere with the establishment of conditioned taste avoidance at any dose tested. This pattern is similar to that evident in previous studies with ondansetron (e.g. Limebeer and Parker, 2000), cannabidiol (CBD; Parker et al., 2002; Rock et al., 2012), cannabidiolic acid (CBDA; Bolognini et al., 2013) and THC (Limebeer and Parker, 1999). Since conditioned taste avoidance is produced by emetic drugs, but also by rewarding drugs, and because anti-emetic drugs do not interfere with taste avoidance, unlike conditioned gaping, we have argued that conditioned taste avoidance is not a selective measure of nausea (see Parker et al., 2008).
THCA may be a more desirable therapeutic treatment for nausea and vomiting than THC because it is both more potent and devoid of psychoactive properties. There is an intriguing parallel between the effects of THCA and the acid precursor of CBD, CBDA, on nausea and vomiting in our models. Our group (Bolognini et al., 2013; Rock and Parker, 2013) has recently also evaluated the potential of CBDA to prevent the establishment of nausea-induced conditioned gaping reactions elicited by a flavour (as a model of acute nausea), to prevent the expression of contextually elicited conditioned gaping reactions (a model of AN) in rats and to prevent vomiting in shrews. Like THCA, CBDA was much more potent in reducing these nausea-induced responses than has been shown with CBD (e.g. Parker et al., 2002; Rock et al., 2008) and these effects like those of CBD (Rock et al., 2012) are 5-HT1A receptor mediated. As both CBDA and THCA are present in the unheated cannabis plant, future work will evaluate potential synergistic effects (potentially by an interaction between CB1 and 5-HT1A mechanisms) of the two compounds on these behaviours reflective of nausea in rats as well as on vomiting in shrews.
Acknowledgments
This research was supported by grants from GW Pharmaceuticals to L. A. P. and from the Natural Sciences and Engineering Council of Canada (NSERC-92057) to L. A. P. The authors would like to thank Quotient Bioresearch (Fordham, Cambridgeshire, UK) for the analysis of plasma samples. The authors would also like to thank Linda Groocock for management of the shrew colony at the University of Guelph, and Martin Sticht for assistance with these studies.
Glossary
- 5-HT1A
5-hydroxytryptamine-1A
- AN
anticipatory nausea
- CB1
cannabinoid 1
- CB2
cannabinoid 2
- CBD
cannabidiol
- CBDA
cannabidiolic acid
- LiCl
lithium chloride
- LSD
least significant difference
- SAL
saline
- SR
SR141716
- THC
Δ9-tetrahydrocannabinol
- THCA
tetrahydrocannabinolic acid
- TR
taste reactivity
- VEH
vehicle
- WAY
WAY100635
Conflict of interest
The research performed in Guelph was funded by GW Research Ltd, UK.
References
- Ahmed SA, Ross SA, Slade D, Radwan MM, Zulfiqar F, Matsumoto RR, et al. Cannabinoid ester constituents from high-potency Cannabis sativa. J Nat Prod. 2008;71:536–542. doi: 10.1021/np070454a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolognini D, Rock EM, Cluny NL, Cascio MG, Limebeer CL, Duncan M, et al. Cannabidiolic acid prevents vomiting in Suncus murinus and nausea-induced behaviour in rats by enhancing 5-HT1A receptor activation. Br J Pharmacol. 2013;168:1456–1470. doi: 10.1111/bph.12043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carey MP, Burish TG, Brenner DE. Delta-9-tetrahydrocannabinol in cancer chemotherapy: research problems and issues. Ann Intern Med. 1983;99:106–114. doi: 10.7326/0003-4819-99-1-106. [DOI] [PubMed] [Google Scholar]
- Cluny NL, Naylor RJ, Whittle BA, Javid FA. The effects of cannabidiol and tetrahydrocannabinol on motion-induced emesis in Suncus murinus. Basic Clin Pharmacol Toxicol. 2008;103:150–156. doi: 10.1111/j.1742-7843.2008.00253.x. [DOI] [PubMed] [Google Scholar]
- Cohen-Yeshurun A, Trembovler V, Alexandrovich A, Ryberg E, Greasley PJ, Mechoulam R, et al. N-arachidonoyl-L-serine is neuroprotective after traumatic brain injury by reducing apoptosis. J Cereb Blood Flow Metab. 2011;31:1768–1777. doi: 10.1038/jcbfm.2011.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen-Yeshurun A, Willner D, Trembovler V, Alexandrovich A, Mechoulam R, Shohami E, et al. N-arachidonoyl-L-serine (AraS) possesses proneurogenic properties in vitro and in vivo after traumatic brain injury. J Cereb Blood Flow Metab. 2013;33:1242–1250. doi: 10.1038/jcbfm.2013.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotter J. Efficacy of crude marijuana and synthetic delta-9-tetrahydrocannabinol as treatment for chemotherapy-induced nausea and vomiting: a systematic literature review. Oncol Nurs Forum. 2009;36:345–352. doi: 10.1188/09.ONF.345-352. [DOI] [PubMed] [Google Scholar]
- Crawford SM, Buckman R. Nabilone and metoclopramide in the treatment of nausea and vomiting due to cisplatinum: a double blind study. Med Oncol Tumor Pharmacother. 1986;3:39–42. doi: 10.1007/BF02934575. [DOI] [PubMed] [Google Scholar]
- Cunningham D, Bradley CJ, Forrest GJ, Hutcheon AW, Adams L, Sneddon M, et al. A randomized trial of oral nabilone and prochlorperazine compared to intravenous metoclopramide and dexamethasone in the treatment of nausea and vomiting induced by chemotherapy regimens containing cisplatin or cisplatin analogues. Eur J Cancer Clin Oncol. 1988;24:685–689. doi: 10.1016/0277-5379(88)90300-8. [DOI] [PubMed] [Google Scholar]
- Darmani NA. Serotonin 5-HT3 receptor antagonists prevent cisplatin-induced emesis in Cryptotis parva: a new experimental model of emesis. J Neural Transm. 1998;105:1143–1154. doi: 10.1007/s007020050118. [DOI] [PubMed] [Google Scholar]
- Darmani NA. Delta(9)-tetrahydrocannabinol and synthetic cannabinoids prevent emesis produced by the cannabinoid CB(1) receptor antagonist/inverse agonist SR 141716A. Neuropsychopharmacology. 2001a;24:198–203. doi: 10.1016/S0893-133X(00)00197-4. [DOI] [PubMed] [Google Scholar]
- Darmani NA. Delta-9-tetrahydrocannabinol differentially suppresses cisplatin-induced emesis and indices of motor function via cannabinoid CB(1) receptors in the least shrew. Pharmacol Biochem Behav. 2001b;69:239–249. doi: 10.1016/s0091-3057(01)00531-7. [DOI] [PubMed] [Google Scholar]
- Darmani NA, Johnson JC. Central and peripheral mechanisms contribute to the antiemetic actions of delta-9-tetrahydrocannabinol against 5-hydroxytryptophan-induced emesis. Eur J Pharmacol. 2004;488:201–212. doi: 10.1016/j.ejphar.2004.02.018. [DOI] [PubMed] [Google Scholar]
- Darmani NA, Janoyan JJ, Crim J, Ramirez J. Receptor mechanism and antiemetic activity of structurally-diverse cannabinoids against radiation-induced emesis in the least shrew. Eur J Pharmacol. 2007;563:187–196. doi: 10.1016/j.ejphar.2007.01.093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frytak S, Moertel CG, O'Fallon JR, Rubin J, Creagan ET, O'Connell MJ, et al. Delta-9-tetrahydrocannabinol as an antiemetic for patients receiving cancer chemotherapy. A comparison with prochlorperazine and a placebo. Ann Intern Med. 1979;91:825–830. doi: 10.7326/0003-4819-91-6-825. [DOI] [PubMed] [Google Scholar]
- Gaoni Y, Mechoulam R. Isolation, structure, and partial synthesis of an active constituent of hashish. J Am Chem Soc. 1964;86:1646–1647. [Google Scholar]
- Grunfeld Y, Edery H. Psychopharmacological activity of the active constituents of hashish and some related cannabinoids. Psychopharmacologia. 1969;14:200–210. doi: 10.1007/BF00404218. [DOI] [PubMed] [Google Scholar]
- Kilkenny C, Browne W, Cuthill IC, Emerson M, Altman DG. Animal research: reporting in vivo experiments: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1577–1579. doi: 10.1111/j.1476-5381.2010.00872.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwiatkowska M, Parker LA, Burton P, Mechoulam R. A comparative analysis of the potential of cannabinoids and ondansetron to suppress cisplatin-induced emesis in the Suncus murinus (house musk shrew) Psychopharmacology (Berl) 2004;174:254–259. doi: 10.1007/s00213-003-1739-9. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA. Delta-9-tetrahydrocannabinol interferes with the establishment and the expression of conditioned rejection reactions produced by cyclophosphamide: a rat model of nausea. Neuroreport. 1999;10:3769–3772. doi: 10.1097/00001756-199912160-00009. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA. The antiemetic drug ondansetron interferes with lithium-induced conditioned rejection reactions, but not lithium-induced taste avoidance in rats. Anim Behav Process. 2000;26:371–384. doi: 10.1037//0097-7403.26.4.371. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Hall G, Parker LA. Exposure to a lithium-paired context elicits gaping in rats: a model of anticipatory nausea. Physiol Behav. 2006;88:398–403. doi: 10.1016/j.physbeh.2006.04.014. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Vemuri VK, Bedard H, Lang ST, Ossenkopp KP, Makriyannis A, et al. Inverse agonism of cannabinoid CB1 receptors potentiates LiCl-induced nausea in the conditioned gaping model in rats. Br J Pharmacol. 2010;161:336–349. doi: 10.1111/j.1476-5381.2010.00885.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGrath J, Drummond G, McLachlan E, Kilkenny C, Wainwright C. Guidelines for reporting experiments involving animals: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1573–1576. doi: 10.1111/j.1476-5381.2010.00873.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin BR, Compton DR, Thomas BF, Prescott WR, Little PJ, Razdan RK, et al. Behavioral, biochemical, and molecular modeling evaluations of cannabinoid analogs. Pharmacol Biochem Behav. 1991;40:471–478. doi: 10.1016/0091-3057(91)90349-7. [DOI] [PubMed] [Google Scholar]
- Milman G, Maor Y, Abu-Lafi S, Horowitz M, Gallily R, Batkai S, et al. N-arachidonoyl L-serine, an endocannabinoid-like brain constituent with vasodilatory properties. Proc Natl Acad Sci U S A. 2006;103:2428–2433. doi: 10.1073/pnas.0510676103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orr LE, McKernan JF. Antiemetic effect of delta 9-tetrahydrocannabinol in chemotherapy-associated nausea and emesis as compared to placebo and compazine. J Clin Pharmacol. 1981;21:76S–80S. doi: 10.1002/j.1552-4604.1981.tb02578.x. [DOI] [PubMed] [Google Scholar]
- Orr LE, McKernan JF, Bloome B. Antiemetic effect of tetrahydrocannabinol. Compared with placebo and prochlorperazine in chemotherapy-associated nausea and emesis. Arch Intern Med. 1980;140:1431–1433. doi: 10.1001/archinte.140.11.1431. [DOI] [PubMed] [Google Scholar]
- Parker LA. Taste avoidance and taste aversion: evidence for two different processes. Learn Behav. 2003;31:165–172. doi: 10.3758/bf03195979. [DOI] [PubMed] [Google Scholar]
- Parker LA, Kemp SW. Tetrahydrocannabinol (THC) interferes with conditioned retching in Suncus murinus: an animal model of anticipatory nausea and vomiting (ANV) Neuroreport. 2001;12:749–751. doi: 10.1097/00001756-200103260-00027. [DOI] [PubMed] [Google Scholar]
- Parker LA, Mechoulam R. Cannabinoid agonists and antagonists modulate lithium-induced conditioned gaping in rats. Integr Physiol Behav Sci. 2003;38:133–145. doi: 10.1007/BF02688831. [DOI] [PubMed] [Google Scholar]
- Parker LA, Mechoulam R, Schlievert C. Cannabidiol, a non-psychoactive component of cannabis and its synthetic dimethylheptyl homolog suppress nausea in an experimental model with rats. Neuroreport. 2002;13:567–570. doi: 10.1097/00001756-200204160-00006. [DOI] [PubMed] [Google Scholar]
- Parker LA, Mechoulam R, Schlievert C, Abbott L, Fudge ML, Burton P. Effects of cannabinoids on lithium-induced conditioned rejection reactions in a rat model of nausea. Psychopharmacology (Berl) 2003;166:156–162. doi: 10.1007/s00213-002-1329-2. [DOI] [PubMed] [Google Scholar]
- Parker LA, Kwiatkowska M, Burton P, Mechoulam R. Effect of cannabinoids on lithium-induced vomiting in the Suncus murinus (house musk shrew) Psychopharmacology (Berl) 2004;171:156–161. doi: 10.1007/s00213-003-1571-2. [DOI] [PubMed] [Google Scholar]
- Parker LA, Kwiatkowska M, Mechoulam R. Delta-9-tetrahydrocannabinol and cannabidiol, but not ondansetron, interfere with conditioned retching reactions elicited by a lithium-paired context in Suncus murinus: an animal model of anticipatory nausea and vomiting. Physiol Behav. 2006;87:66–71. doi: 10.1016/j.physbeh.2005.08.045. [DOI] [PubMed] [Google Scholar]
- Parker LA, Rana SA, Limebeer CL. Conditioned nausea in rats: assessment by conditioned disgust reactions, rather than conditioned taste avoidance. Can J Exp Psychol. 2008;62:198–209. doi: 10.1037/a0012531. [DOI] [PubMed] [Google Scholar]
- Parker LA, Limebeer CL, Rock EM, Litt DL, Kwiatkowska M, Piomelli D. The FAAH inhibitor URB-597 interferes with cisplatin- and nicotine-induced vomiting in the Suncus murinus (house musk shrew) Physiol Behav. 2009;97:121–124. doi: 10.1016/j.physbeh.2009.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rinaldi-Carmona M, Barth F, Heaulme M, Shire D, Calandra B, Congy C, et al. SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Lett. 1994;350:240–244. doi: 10.1016/0014-5793(94)00773-x. [DOI] [PubMed] [Google Scholar]
- Rock E, Parker L. Effect of low doses of cannabidiolic acid and ondansetron on LiCl-induced conditioned gaping (a model of nausea-induced behaviour) in rats. Br J Pharmacol. 2013;169:685–692. doi: 10.1111/bph.12162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock EM, Limebeer CL, Mechoulam R, Piomelli D, Parker LA. The effect of cannabidiol and URB597 on conditioned gaping (a model of nausea) elicited by a lithium-paired context in the rat. Psychopharmacology (Berl) 2008;196:389–395. doi: 10.1007/s00213-007-0970-1. [DOI] [PubMed] [Google Scholar]
- Rock E, Bolognini D, Limebeer C, Cascio M, Anavi-Goffer S, Fletcher P, et al. Cannabidiol, a non-psychotropic component of cannabis, attenuates vomiting and nausea-like behaviour via indirect agonism of 5-HT(1A) somatodendritic autoreceptors in the dorsal raphe nucleus. Br J Pharmacol. 2012;165:2620–2634. doi: 10.1111/j.1476-5381.2011.01621.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tramer MR, Carroll D, Campbell FA, Reynolds DJ, Moore RA, McQuay HJ. Cannabinoids for control of chemotherapy induced nausea and vomiting: quantitative systematic review. BMJ. 2001;323:16–21. doi: 10.1136/bmj.323.7303.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungerleider JT, Andrysiak TA, Fairbanks LA, Tesler AS, Parker RG, Ungerleider JT, et al. Tetrahydrocannabinol vs. prochlorperazine. The effects of two antiemetics on patients undergoing radiotherapy. Radiology. 1984;150:598–599. doi: 10.1148/radiology.150.2.6318262. [DOI] [PubMed] [Google Scholar]
- Van Sickle MD, Oland LD, Ho W, Hillard CJ, Mackie K, Davison JS, et al. Cannabinoids inhibit emesis through CB1 receptors in the brainstem of the ferret. Gastroenterology. 2001;121:767–774. doi: 10.1053/gast.2001.28466. [DOI] [PubMed] [Google Scholar]
- Verhoeckx KC, Korthout HA, van Meeteren-Kreikamp AP, Ehlert KA, Wang M, van der Greef J, et al. Unheated Cannabis sativa extracts and its major compound THC-acid have potential immuno-modulating properties not mediated by CB1 and CB2 receptor coupled pathways. Int Immunopharmacol. 2006;6:656–665. doi: 10.1016/j.intimp.2005.10.002. [DOI] [PubMed] [Google Scholar]
- Wiley JL, Martin BR. Cannabinoid pharmacological properties common to other centrally acting drugs. Eur J Pharmacol. 2003;471:185–193. doi: 10.1016/s0014-2999(03)01856-9. [DOI] [PubMed] [Google Scholar]