

Abstract
BACKGROUND AND PURPOSE
The cannabinoid 1 (CB1) receptor inverse agonists/antagonists, rimonabant (SR141716, SR) and AM251, produce nausea and potentiate toxin-induced nausea by inverse agonism (rather than antagonism) of the CB1 receptor. Here, we evaluated two phytocannabinoids, cannabidivarin (CBDV) and Δ9-tetrahydrocannabivarin (THCV), for their ability to produce these behavioural effect characteristics of CB1 receptor inverse agonism in rats.
EXPERIMENTAL APPROACH
In experiment 1, we investigated the potential of THCV and CBDV to produce conditioned gaping (measure of nausea-induced behaviour) in the same manner as SR and AM251. In experiment 2, we investigated the potential of THCV and CBDV to enhance conditioned gaping produced by a toxin in the same manner as CB1 receptor inverse agonists.
KEY RESULTS
SR (10 and 20 mg·kg−1) and AM251 (10 mg·kg−1) produced conditioned gaping; however, THCV (10 or 20 mg·kg−1) and CBDV (10 or 200 mg·kg−1) did not. At a subthreshold dose for producing nausea, SR (2.5 mg·kg−1) enhanced lithium chloride (LiCl)-induced conditioned gaping, whereas Δ9-tetrahydrocannabinol (THC, 2.5 and 10 mg·kg−1), THCV (2.5 or 10 mg·kg−1) and CBDV (2.5 or 200 mg·kg−1) did not; in fact, THC (2.5 and 10 mg·kg−1), THCV (10 mg·kg−1) and CBDV (200 mg·kg−1) suppressed LiCl-induced conditioned gaping, suggesting anti-nausea potential.
CONCLUSIONS AND IMPLICATIONS
The pattern of findings indicates that neither THCV nor CBDV produced a behavioural profile characteristic of CB1 receptor inverse agonists. As well, these compounds may have therapeutic potential in reducing nausea.
Keywords: CB1 receptor inverse agonism, cannabidivarin, tetrahydrocannabivarin, CB1 receptor antagonism, nausea, depression, anhedonia, rimonabant, THC
Introduction
Side effects of the cannabinoid 1 (CB1) receptor inverse agonist/antagonist, rimonabant (SR141716; SR), in humans include nausea and depression (Despres, 2009; de Mattos Viana et al., 2009). The nausea produced by SR and AM251 (another CB1 inverse agonist/antagonist) is the result of inverse agonism at the CB1 receptor (Sink et al., 2008) because equivalent doses of AM251, but not the neutral CB1 receptor antagonists AM4113 (Sink et al., 2008), AM6527 or AM6545 (Cluny et al., 2010), produce lithium chloride (LiCl)-induced conditioned gaping reactions, a measure of nausea-induced behaviour in rats. Only drugs that produce emesis in species capable of vomiting produce conditioned gaping reactions and anti-emetic agents prevent the establishment of these nausea-induced gaping reactions in rats (see Parker et al., 2008; 2009 for a review).
The conditioned gaping model has been shown to detect the nauseating side effect of several compounds, including selective serotonin reuptake inhibitors (Limebeer et al., 2009; Tuerke et al., 2009), PDE-4 inhibitors (Rock et al., 2009) and CB1 receptor inverse agonists (McLaughlin et al., 2005; Sink et al., 2008). As well as producing nausea on their own, the CB1 receptor inverse agonists (at subthreshold doses that do not produce nausea on their own), SR (Parker and Mechoulam, 2003; Parker et al., 2003) and AM251 (Limebeer et al., 2010), potentiate the nausea produced by the toxin LiCl. This potentiation of nausea is the result of CB1 receptor inverse agonism because neither the neutral antagonist, AM6545 (peripherally restricted; Tam et al., 2010) nor AM6527 (which crosses the blood–brain barrier; Sink et al., 2009), produced potentiation of LiCl-induced nausea in the gaping model (Limebeer et al., 2010). Interestingly, Chambers et al. (2007) also reported that AM251, but not the neutral CB1 receptor antagonist AM4113, also potentiated morphine-6-glucoronide toxin-induced vomiting (at a dose that did not produce vomiting on its own) in the ferret. These findings suggest that it is the CB1 receptor inverse agonist effects rather than their CB1 receptor antagonist effects that mediate the nausea produced as a side effect of these compounds.
Phytocannabinoids have recently become candidates for therapeutic applications; however, their usefulness is limited if they exhibit CB1 receptor inverse agonist activity. Much research on phytocannabinoids has concentrated mostly on the psychoactive compound, Δ9-tetrahydrocannabinol (THC), and the primary non-psychoactive cannabinoid, cannabidiol (CBD), found in marijuana (e.g. Mechoulam et al., 2002). Low doses (0.5 mg·kg−1 THC and 1–5 mg·kg−1 CBD) of both of these compounds have been shown to suppress LiCl-induced conditioned gaping in the rodent model of conditioned nausea (Limebeer and Parker, 1999; Parker et al., 2002; 2003; Rock et al., 2008; 2012).
The effects of the phytocannabinoids cannabidivarin (CBDV) and tetrahydrocannabivarin (THCV), although identified in the 1970s (Merkus, 1971), have only recently been investigated. Under acidic conditions, CBDV isomerizes into THCV, and in the plant, CBDV is the precursor of THCV (Deiana et al., 2012). Higher levels of CBDV and THCV are more common in Cannabis indica than in Cannabis sativa (Hillig and Mahlberg, 2004).
Recent in vivo work has highlighted CBDV's anti-inflammatory effects in mice (Tubaro et al., 2010). Most recently, in vivo, Hill et al. (2012) demonstrated CBDV's effectiveness as an anti-convulsant (≥50–100 mg·kg−1) in various animal seizure models, suggesting possible anti-epilepsy effects, but little else is understood about the behavioural effects of CBDV. CBDV has also been shown to act as an agonist at human transient receptor potential (TRP) channels of the ankyrin type-1 (TRPA1), TRP channels of vanilloid type-1 and type-2 agonist, but as a TRP channel of melastatin type-8 antagonist (De Petrocellis et al., 2011; 2012). These varied effects warrant further investigations into CBDV's mechanism of action.
Work with both plant-derived THCV and synthetic THCV (0–4394) has elucidated its behavioural effects and mechanism of action. In vitro, THCV has been shown to act as a CB1 and CB2 receptor antagonist (Thomas et al., 2005; Pertwee et al., 2007), and also to activate CB2 receptors (Bolognini et al., 2010). As well, in contrast to SR, THCV (0–4394) does not produce inverse agonist activity in the [35S] GTPγS binding assay in mouse whole brain membrane and fails to produce stimulation of [35S] GTPγS binding to such membranes (Pertwee et al., 2007). Indeed, THCV shares the ability of AM251 to reduce the food intake and body weight of non-fasted and fasted mice (Riedel et al., 2009), and at a dose of 2.5 mg·kg−1 (i.p.), THCV has recently been shown not to modify saccharin palatability (O'Brien et al., 2013). If THCV is devoid of the CB1 receptor inverse agonist properties of SR, which are responsible for nausea (McLaughlin et al., 2005; Sink et al., 2008) and potentially depression (Beyer et al., 2010; Sink et al., 2010), then THCV may be a potential candidate for therapeutic use. Indeed, THCV has been shown in animal models to suppress seizure activity (Hill et al., 2010), reduce inflammation and inflammatory pain (Bolognini et al., 2010), reduce weight due to hypophagia (Riedel et al., 2009) and most recently to even reduce Parkinson's disease symptoms, as well as disease progression (Garcia et al., 2011). In addition, unlike SR (2.5 mg·kg−1), THCV (2.5 mg·kg−1) did not produce an anxiogenic-like response of avoidance of an open lit area in the light/dark immersion test (O'Brien et al., 2013), an effect consistently observed with CB1 receptor inverse agonists, but not with neutral CB1 receptor antagonists such as AM4113 (Sink et al., 2010). Therefore, further examination of potential nauseating side effects of these phytocannabinoids is important to exclude potential in vivo CB1 receptor inverse agonist activity.
If a compound acts as a CB1 receptor inverse agonist, like SR and AM251, it is expected to (i) produce conditioned gaping reactions to a novel flavour with which it is paired (experiment 1), and (ii) at subthreshold doses for producing nausea on their own, enhance the nauseating effects of another toxin, that is, produce potentiation of LiCl-induced conditioned gaping (experiment 2). If CBDV and THCV do not produce conditioned gaping on their own and do not enhance LiCl-induced conditioned gaping, it is likely that they do not exhibit CB1 receptor inverse agonist activity. These experiments can also be used to evaluate the potential of the compounds to reduce nausea-induced conditioned gaping in rats, like THC (Limebeer and Parker, 1999), HU-210 (Parker and Mechoulam, 2003; Parker et al., 2003), URB597 (Cross-Mellor et al., 2007), CBD (Parker et al., 2002; Rock et al., 2012) and cannabidiolic acid (Bolognini et al., 2013; Rock and Parker, 2013). Therefore, if a compound reduces nausea, these effects will be detected by the following experiments as well.
Methods
Experimental procedures
Experiment 1: does the compound produce conditioned gaping on its own?
Rats were implanted with intraoral cannulae under isoflurane anaesthesia (as described by Limebeer et al., 2010). Following recovery from surgery (at least 3 days), rats received an adaptation trial. They were placed in the taste reactivity (TR) chamber with their cannula attached to an infusion pump (model KDS100; KD Scientific, Holliston, MA, USA) for fluid delivery, and water was infused into their intraoral cannula for 2 min at a rate of 1 mL·min−1. The TR chambers were made of clear Plexiglas (22.5 × 26 × 20 cm) that sat on a table with a clear glass top. A mirror beneath the chamber on a 45° angle facilitated viewing of the ventral surface of the rat to observe the orofacial responses. On the following day, the rats received a conditioning trial, during which they were intraorally infused with a 0.1% saccharin solution for 2 min at a rate of 1 mL·min−1. Immediately after the saccharin infusion, they (n = 10 per group, except AM251 with n = 6) were injected with either VEH (1/1/18: ethanol/Cremophor/saline), 10 mg·kg−1 SR, 20 mg·kg−1 SR, 10 mg·kg−1 AM251, 10 mg·kg−1 THCV, 20 mg·kg−1 THCV, 10 mg·kg−1 CBDV or 200 mg·kg−1 CBDV.
Seventy-two hours after the conditioning trial, rats were returned to the TR chamber for a test and intraorally infused with 0.1% saccharin solution. Their orofacial reactions were video recorded with the feed from the video camera (Sony DCR-HC48; Henry's Cameras, Waterloo, ON, Canada) fire-wired into a computer. The video tapes were later scored (at ½ speed) by a trained observer blind to the experimental conditions using ‘the Observer’ (Noldus Information Technology Inc., Leesburg, VA, USA) for the behaviour of gaping (large openings of the mouth and jaw, with lower incisors exposed). The videotapes were scored by two trained raters, resulting in an extremely high inter-rater reliability score (r = 0.97).
Experiment 2: does the compound modify saccharin palatability and/or potentiate (or reduce) the nausea produced by LiCl?
Following recovery from intraoral cannulation surgery, the rats received the adaptation trial and the TR test as described in experiment 1, except as noted. On the day of conditioning, the rats were injected with vehicle (VEH; n = 10), 2.5 mg·kg−1 SR (n = 10), 2.5 mg·kg−1 THC (n = 6), 10 mg·kg−1 THC (n = 7), 2.5 mg·kg−1 THCV (n = 10) or 10 mg·kg−1 THCV (n = 6), 2.5 mg·kg−1 CBDV (n = 10) or 200 mg·kg−1 CBDV (n = 13). Thirty minutes later, each rat was intraorally infused with 0.1% saccharin solution, while their orofacial responses were video recorded from the mirror beneath the chamber. Immediately following the 2 min intraoral infusion of saccharin, the rats were injected with 20 mg·kg−1 of 0.15 M LiCl.
Seventy-two hours later, the rats received a drug-free test trial, during which they received a 2 min intraoral infusion of 0.1% saccharin solution and the frequency of gaping was measured. The videotapes were later scored (at ½ speed) by an observer blinded to the experimental conditions using ‘the Observer’ for conditioned gaping as well as tongue protrusions (extensions of the tongue, both forward and lateral, from the mouth as a measure of hedonic reactions) during conditioning. As well, to determine other non-specific effects of the drug during conditioning, the conditioning tapes were scored for the duration (s) of bouts of active locomotion (forward propulsion of the rats' body by movement of the forepaws on the floor of the chamber).
Animals
Animal procedures were according to the Canadian Council on Animal Care (CCAC). The protocol was approved by the Institutional Animal Care Committee, which is accredited by the CCAC. The authors consulted with the ARRIVE guidelines of the British Journal of Pharmacology for reporting experiments involving animals (Kilkenny et al., 2009; McGrath et al., 2010). Naïve male Sprague-Dawley rats (experiment 1, n = 76; experiment 2, n = 73), obtained from Charles River Laboratories (St Constant, QC, Canada), were single-housed in shoebox cages in the colony room at an ambient temperature of 21°C with a 12/12 light/dark schedule (lights off at 0800 hours) and maintained on ad libitum food and water. Rats weighed between 236 and 455 g on the day of conditioning.
Drugs and materials
All cannabinoid compounds were prepared in a VEH of ethanol/Cremophor (Sigma, St Louis, MO, USA)/saline (SAL; 1:1:18) and administered i.p. LiCl (Sigma) was prepared in a 0.15 M solution with sterile water and administered i.p. at a volume of 20 mL·kg−1 (127.2 mg·kg−1) in experiment 2.
SR was prepared at 2.5, 10 and 20 mg·kg−1 and administered at a volume of 2 mL·kg−1. Two sources of SR were used at the 10 mg·kg−1 dose (n = 8 per group): one purchased from Sequoia laboratories, GB, and the other provided by B. Whalley's laboratory from Otsuka Pharmaceuticals (Osaka, Japan). A comparison of the gaping produced by these two sources of SR revealed no significant difference in potential to produce conditioned gaping [Sequoia laboratories SR (mean gaping = 3.6); Otsuka SR (mean gaping = 3.8)]; therefore, the data collected with 10 mg·kg−1 SR were combined. The 20 mg·kg−1 dose of SR was from the Otsuka supply.
CBDV (GW Pharmaceuticals, Porton Down, Wiltshire, UK) was prepared at 2.5 and 10 mg·kg−1 and administered at a volume of 2 mL·kg−1. CBDV prepared at 200 mg·kg−1 and was administered at a volume of 10 mL·kg−1. This high dose of CBDV was evaluated, given recent findings that it has anti-epileptic potential at this high dose (Hill et al., 2012). THCV (GW Pharmaceuticals) was prepared at 2.5, 10 and 20 mg·kg−1 and administered at a volume of 2 mL·kg−1. AM251 (Cayman Chemicals, Ann Arbor, MI, USA) was prepared at 10 mg·kg−1 and administered at a volume of 2 mL·kg−1. THC (GW Pharmaceuticals) was prepared at 2.5 and 10 mg·kg−1 and administered at a volume of 2 mL·kg−1.
Results
Experiment 1: does the compound produce conditioned gaping on its own?
The CB1 inverse agonists/antagonists, SR and AM251, produced more conditioned gaping reactions (nausea-like behaviours) than any other group. Figure 1 presents the mean (±SEM) number of gapes displayed by the various groups in experiment 1. The one-way anova revealed a main effect of group [F(7, 74) = 4.9; P < 0.001]. Using Fisher's least significant difference (LSD) comparison tests, all doses of the CB1 receptor inverse agonists/antagonists (SR and AM251) produced significantly more conditioned gaping than any of the other compounds when paired with saccharin solution (P's <0.05). As well, none of the other compounds (at any dose tested) significantly differed from VEH controls. Finally, SR produced marginally more gaping at 20 mg·kg−1 than at 10 mg·kg−1 (P = 0.057). Therefore, at all doses tested, CBDV and THCV did not produce nausea-induced gaping (inverse agonist effect) when compared with VEH, but SR and AM251 both produced the inverse agonist effect of nausea that is not produced by CB1 receptor neutral antagonists (Sink et al., 2008).
Figure 1.
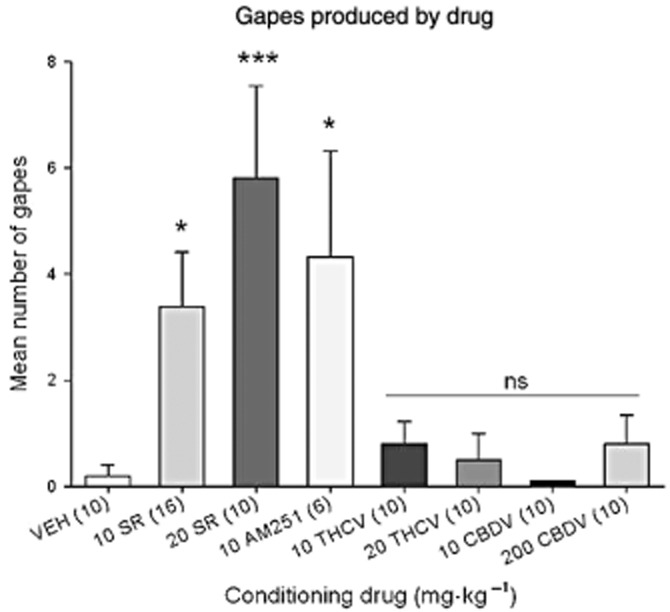
Mean number (±SEM) of gapes at test elicited by 0.1% saccharin solution previously paired with each compound during the drug-free test trial. Numbers in parentheses indicate n per group. *P < 0.05; ***P < 0.001 from other groups indicated by line [10 mg·kg−1 THCV, 20 mg·kg−1 THCV, 10 mg·kg−1 CBDV, 200 mg·kg−1 CBDV and VEH, which did not differ from one another (ns)]. Those conditioned with 10 mg·kg−1 SR, 20 mg·kg−1 SR and AM251 did not significantly differ from one another, although there was a trend for 20 mg·kg−1 SR to display more gaping than 10 mg·kg−1 SR (P = 0.07).
Experiment 2: does the compound modify saccharin palatability and/or potentiate (or reduce) the nausea produced by LiCl?
Saccharin palatability during conditioning
Figure 2 presents the mean (±SEM) number of tongue protrusions displayed during the conditioning trial following exposure to the pretreatment injections, prior to receiving LiCl. A one-way anova revealed a significant pretreatment group effect [F(7, 62) = 4.3; P = 0.001]. VEH pretreated rats displayed significantly more hedonic reactions than those pretreated with 2.5 mg·kg−1 SR (P < 0.02) or 10 mg·kg−1 THCV (P < 0.02). Those rats pretreated with CBDV (2.5 and 200 mg·kg−1), THCV (2.5 mg·kg−1) and THC (2.5 and 10 mg·kg−1) did not significantly differ from VEH-pretreated controls (P's > 0.05). The anova for total activity [F(7, 62) = 1.4; P = 0.2] was non-significant (data not shown). Therefore, the drugs did not affect overall activity. These results suggest that both 2.5 mg·kg−1 SR and 10 mg·kg−1 THCV reduced saccharin palatability, which may reflect an anti-appetite effect of these compounds, both shown to reduce feeding (e.g. Riedel et al., 2009).
Figure 2.
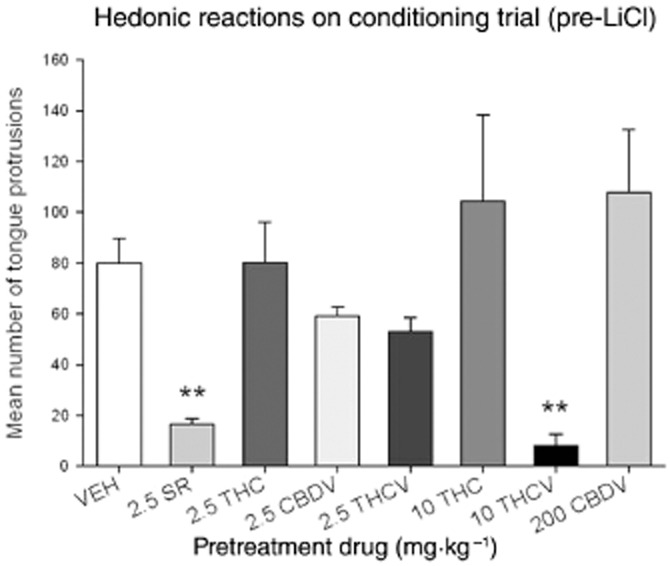
Mean number (±SEM) of tongue protrusions during conditioning elicited by 0.1% saccharin solution 30 min following pretreatment with VEH, 2.5 mg·kg−1 SR, 2.5 mg·kg−1 THC, 10 mg·kg−1 THC, 2.5 mg·kg−1 CBDV, 200 mg·kg−1 CBDV, 2.5 mg·kg−1 THCV or 10 mg·kg−1 THCV. Both groups pretreated with 2.5 mg·kg−1 SR and 10 mg·kg−1 THCV displayed suppressed (**P < 0.02) hedonic reactions relative to VEH-pretreated controls.
Potentiation of LiCl-induced conditioned gaping during the test trial
Figure 3 presents the mean (±SEM) number of conditioned gaping reactions during the drug-free test, 72 h following the conditioning trial. The one-way anova revealed a significant pretreatment group effect [F(7, 62) = 8.0; P < 0.001]. Subsequent LSD tests revealed that only those rats pretreated with 2.5 mg·kg−1 SR displayed potentiated LiCl-induced conditioned gaping reactions relative to VEH-pretreated controls (P < 0.025).
Figure 3.
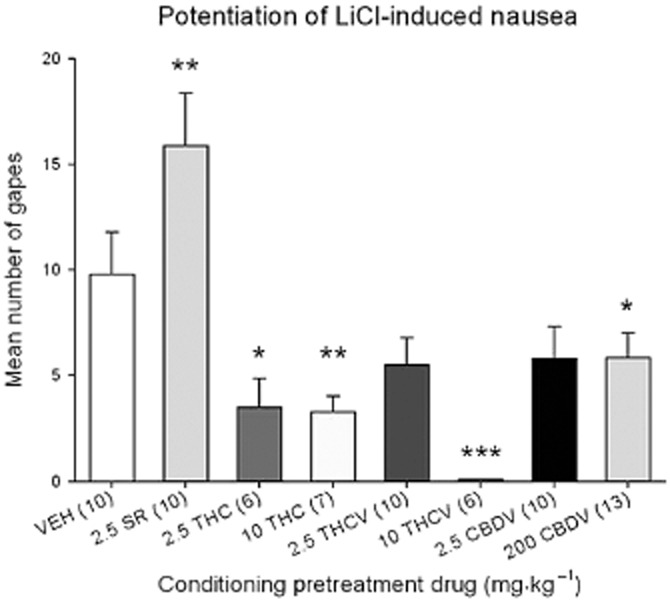
Mean number (±SEM) of gapes elicited by 0.1% saccharin solution previously paired with LiCl during the drug-free TR test trial. Only those pretreated with SR displayed potentiated gaping (**P < 0.025 relative to VEH-pretreated controls) during the drug-free test, suggesting that pretreatment during conditioning enhanced the nausea produced by LiCl. In contrast, 10 mg·kg−1 THCV completely eliminated LiCl-induced gaping (***P < 0.001 relative to VEH) and 200 mg·kg−1 CBDV, 2.5 mg·kg−1 THC (*P < 0.05 relative to VEH) and 10 mg·kg−1 THC (**P < 0.01), reduced LiCl-induced gaping during the drug-free test trial, suggesting that pretreatment during conditioning eliminated the nausea produced by LiCl.
In fact, 10 mg·kg−1 THCV (P < 0.001), 200 mg·kg−1 CBDV (P < 0.05) and both doses of THC (P's < 0.02) produced suppression of LiCl-induced conditioned gaping reactions relative to VEH-pretreated controls, a pattern suggesting an anti-nausea effect of these compounds. In addition, rats pretreated with 2.5 mg·kg−1 CBDV (P = 0.08) and 2.5 mg·kg−1 THCV (P = 0.07) showed marginally attenuated LiCl-induced conditioned gaping reactions relative to VEH-pretreated controls.
Discussion
The results of experiment 1 revealed that when explicitly paired with a novel saccharin solution, neither CBDV (10 or 200 mg·kg−1) nor THCV (10 or 20 mg·kg−1) produced the nausea-like profile of conditioned gaping produced by the CB1 receptor inverse agonists SR and AM251. Previous studies revealed that the nausea produced by both SR and AM251 is mediated by the inverse agonism of the CB1 receptor because CB1 receptor neutral antagonists do not produce this side effect (Sink et al., 2008; Cluny et al., 2010). As conditioned gaping in rats is produced only by compounds that produce nausea and emesis in other species (see Parker and Limebeer, 2008), these in vivo results support previous in vitro findings, suggesting that THCV acts as a CB1 receptor neutral antagonist (Pertwee et al., 2007). Neither CBDV nor THCV should produce the symptoms of nausea, an effect that appears to be caused by the CB1 receptor inverse agonist (and not the CB1 receptor antagonist) effects of SR and AM251. Therefore, these results suggest that the inverse agonism effect of nausea will not hinder the proposed clinical applications for CBDV and THCV, (i.e. anti-inflammatory and anti-convulsant properties).
Experiment 2 evaluated the potential of CBDV and THCV to potentiate (or reduce) nausea produced by LiCl. At doses subthreshold for producing nausea on their own (2.5 mg·kg−1), both SR (Parker and Mechoulam, 2003) and AM251 (Limebeer et al., 2010) pretreatments prior to a saccharin-LiCl pairing potentiate the nausea produced by LiCl, as revealed by potentiated conditioned gaping displayed in the subsequent drug-free test trial 72 h after conditioning. In contrast, CB1 neutral antagonists, AM6527 or AM6545 do not potentiate LiCl-induced nausea (Limebeer et al., 2010). Experiment 2 revealed that only the group pretreated with 2.5 mg·kg−1 SR showed potentiated LiCl-induced conditioned gaping relative to all other groups. In fact, all other phytocannabinoids tested appeared to produce an anti-nausea effect, at least at the higher doses. Rats pretreated with THCV, CBDV or THC during conditioning showed suppressed gaping during the subsequent drug-free test, relative to those pretreated with VEH. This suppression of conditioned gaping by pretreatment with THCV, CBDV and THC indicates that these compounds exhibit anti-nausea potential. The low dose effects of THCV shown here are consistent with those previously reported (Pertwee et al., 2007). At a low dose (2.5 mg·kg−1), THCV acts as a CB1 receptor antagonist (Pertwee et al., 2007) and does not enhance LiCl-induced nausea evidenced as potentiation of conditioned gaping (like AM4113), suggesting that it is a neutral antagonist.
The potential of SR, CBDV, THCV and THC to modify saccharin palatability (the hedonic reaction of tongue protrusions) was assessed during the conditioning trial of experiment 2. We have previously shown that doses of SR and AM251 that are subthreshold for producing nausea (2.5 mg·kg−1) reduce sucrose or saccharin palatability (Jarrett et al., 2007; Limebeer et al., 2010) and at a low dose of 0.5 mg·kg−1 THC enhances sucrose palatability in the TR test (Jarrett et al., 2005). Here, we show that 30 min following injection of both 2.5 mg·kg−1 SR and 10 mg·kg−1 THCV (but not 2.5 mg·kg−1 THCV, or 2.5 or 200 mg·kg−1 CBDV) saccharin palatability was suppressed. These results may reflect the potential of THCV, like SR, to modify appetite for sweet tastes (Riedel et al., 2009).
Interestingly, rats pretreated with 10 mg·kg−1 THCV showed no conditioned gaping reactions during intraoral infusion of saccharin that had been previously paired with LiCl, suggesting that it completely blocked LiCl-induced nausea, an effect evident with drugs that are anti-nausea agents such ondansetron (Limebeer and Parker, 2000), palonosetron (Limebeer et al., 2009) and 8-hydroxy-2-(dipropylamino)tetralin (Limebeer and Parker, 2003). Given that the same dose of THCV also suppressed saccharin palatability during conditioning, it is unlikely that the suppression of nausea was mediated by CB1 receptor agonism (e.g. Pertwee et al., 2007). Further experiments are warranted to determine the mechanism of action of the anti-nausea effects of this phytocannabinoid.
In these studies, CBDV was found to be devoid of inverse agonist properties (unlike SR and AM251), not to modify hedonics, and to reduce conditioned gaping (at a high dose). These actions of CBDV are not consistent with CB1 receptor antagonism because CBDV does not potentiate conditioned gaping, nor does CBDV act like a CB1 receptor agonist (like 0.5 mg·kg−1 THC) to enhance hedonics (Jarrett et al., 2005). CBDV reduced LiCl-induced conditioned gaping as is observed with THC and HU-210 (Parker et al., 2003), but this suppression of gaping was not accompanied by enhanced hedonic reactions during conditioning, suggesting that it may not be due to a CB1 mechanism of action. Alternatively, CBDV could also be acting at the TRP channel receptors in this model, as De Petrocellis et al. (2011; 2012) have shown that CBDV may act at these receptors. Sharkey et al. (2007) have indeed shown that the dual TRVP1/CB1 agonist, arvanil, has anti-emetic effects in ferrets that are mediated by activation of both the TRVP1 and the CB1 receptor. Further experiments are warranted to elucidate the mechanism of action of CBDV's anti-nausea effect.
The finding that higher doses of THC (2.5 and 10 mg·kg−1) reduced LiCl-induced conditioned gaping extend previous findings that a low dose (0.5 mg·kg−1) of THC interferes with cyclophosphamide- (Limebeer and Parker, 1999) and LiCl-induced conditioned gaping reactions (Parker and Mechoulam, 2003; Parker et al., 2003). These previous experiments also demonstrated that the THC-induced suppression of conditioned gaping can be reversed by SR, indicating a CB1 receptor mechanism of action for THC's anti-nausea effects (Parker and Mechoulam, 2003; Parker et al., 2003). These findings are also consistent with the human literature. In cancer patients undergoing chemotherapy treatment, administration of oral THC significantly reduces nausea and vomiting, in comparison to placebo controls (Sallan et al., 1975; Chang et al., 1979; Frytak et al., 1979; Orr et al., 1980; Sweet et al., 1981).
In conclusion, at all doses tested, unlike SR and AM251, both CBDV and THCV neither (i) produced conditioned gaping on their own when explicitly paired with saccharin solution nor (ii) potentiated LiCl-induced conditioned gaping. In fact, both compounds produced an anti-nausea-like effect in experiment 2; that is, CBDV partially reduced LiCl-induced nausea, and at the highest dose tested, THCV blocked LiCl-induced nausea. Taken together, these results suggest that neither THCV nor CBDV acts as a CB1 receptor inverse agonist. Therefore, it seems that these two phytocannabinoids may be promising therapeutics, with anti-nausea properties, devoid of symptoms associated with CB1 receptor inverse agonism.
Acknowledgments
The authors would like to thank Cheryl Limebeer for her assistance with all aspects of these experiments. These experiments were funded by G.W. Pharmaceuticals and an NSERC operating grant (92057) to L. A. P.
Glossary
- CB1
cannabinoid 1
- CBD
cannabidiol
- CBDV
cannabidivarin
- LiCl
lithium chloride
- LSD
least significant difference
- SAL
saline
- SR
SR141716
- THC
Δ9-tetrahydrocannabinol
- THCV
Δ9-tetrahydrocannabivarin
- TR
taste reactivity
- TRP
transient receptor potential
- TRPA1
transient receptor potential channels of the ankyrin type-1
- VEH
vehicle
Conflict of interest
The research performed in Guelph was funded by GW Research Ltd, UK. C. S. and M. D. are employees of GW Research Ltd, UK.
References
- Beyer CE, Dwyer JM, Piesla MJ, Platt BJ, Shen R, Rahman Z, et al. Depression-like phenotype following chronic CB1 receptor antagonism. Neurobiol Dis. 2010;39:148–155. doi: 10.1016/j.nbd.2010.03.020. [DOI] [PubMed] [Google Scholar]
- Bolognini D, Costa B, Maione S, Comelli F, Marini P, Di Marzo V, et al. The plant cannabinoid Delta9-tetrahydrocannabivarin can decrease signs of inflammation and inflammatory pain in mice. Br J Pharmacol. 2010;160:677–687. doi: 10.1111/j.1476-5381.2010.00756.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolognini D, Rock EM, Cluny NL, Cascio MG, Limebeer CL, Duncan M, et al. Cannabidiolic acid prevents vomiting in Suncus murinus and nausea-induced behaviour in rats by enhancing 5-HT1A receptor activation. Br J Pharmacol. 2013;168:1456–1470. doi: 10.1111/bph.12043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambers AP, Vemuri VK, Peng Y, Wood JT, Olszewska T, Pittman QJ, et al. A neutral CB1 receptor antagonist reduces weight gain in rat. Am J Physiol Regul Integr Comp Physiol. 2007;293:R2185–R2193. doi: 10.1152/ajpregu.00663.2007. [DOI] [PubMed] [Google Scholar]
- Chang AE, Shiling DJ, Stillman RC, Goldberg NH, Seipp CA, Barofsky I, et al. Delata-9-tetrahydrocannabinol as an antiemetic in cancer patients receiving high-dose methotrexate. A prospective, randomized evaluation. Ann Intern Med. 1979;91:819–824. doi: 10.7326/0003-4819-91-6-819. [DOI] [PubMed] [Google Scholar]
- Cluny NL, Vemuri VK, Chambers AP, Limebeer CL, Bedard H, Wood JT, et al. A novel peripherally restricted cannabinoid receptor antagonist, AM6545, reduces food intake and body weight, but does not cause malaise, in rodents. Br J Pharmacol. 2010;161:629–642. doi: 10.1111/j.1476-5381.2010.00908.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross-Mellor SK, Ossenkopp KP, Piomelli D, Parker LA. Effects of the FAAH inhibitor, URB597, and anandamide on lithium-induced taste reactivity responses: a measure of nausea in the rat. Psychopharmacology (Berl) 2007;190:135–143. doi: 10.1007/s00213-006-0589-7. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Ligresti A, Moriello AS, Allara M, Bisogno T, Petrosino S, et al. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol. 2011;163:1479–1494. doi: 10.1111/j.1476-5381.2010.01166.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Petrocellis L, Orlando P, Moriello AS, Aviello G, Stott C, Izzo AA, et al. Cannabinoid actions at TRPV channels: effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol (Oxf) 2012;204:255–266. doi: 10.1111/j.1748-1716.2011.02338.x. [DOI] [PubMed] [Google Scholar]
- Deiana S, Watanabe A, Yamasaki Y, Amada N, Arthur M, Fleming S, et al. Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Delta-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive-compulsive behaviour. Psychopharmacology (Berl) 2012;219:859–873. doi: 10.1007/s00213-011-2415-0. [DOI] [PubMed] [Google Scholar]
- Despres JP. Pleiotropic effects of rimonabant: clinical implications. Curr Pharm Des. 2009;15:553–570. doi: 10.2174/138161209787315666. [DOI] [PubMed] [Google Scholar]
- Frytak S, Moertel CG, O'Fallon JR, Rubin J, Creagan ET, O'Connell MJ, et al. Delta-9-tetrahydrocannabinol as an antiemetic for patients receiving cancer chemotherapy. A comparison with prochlorperazine and a placebo. Ann Intern Med. 1979;91:825–830. doi: 10.7326/0003-4819-91-6-825. [DOI] [PubMed] [Google Scholar]
- Garcia C, Palomo-Garo C, Garcia-Arencibia M, Ramos J, Pertwee R, Fernandez-Ruiz J. Symptom-relieving and neuroprotective effects of the phytocannabinoid Δ9-THCV in animal models of Parkinson's disease. Br J Pharmacol. 2011;163:1495–1506. doi: 10.1111/j.1476-5381.2011.01278.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill AJ, Weston SE, Jones NA, Smith I, Bevan SA, Williamson EM, et al. Δ9-Tetrahydrocannabivarin suppresses in vitro epileptiform and in vivo seizure activity in adult rats. Epilepsia. 2010;51:1522–1532. doi: 10.1111/j.1528-1167.2010.02523.x. [DOI] [PubMed] [Google Scholar]
- Hill AJ, Mercier MS, Hill TD, Glyn SE, Jones NA, Yamasaki Y, et al. Cannabidivarin is anticonvulsant in mouse and rat in vitro and in seizure models. Br J Pharmacol. 2012;167:1629–1642. doi: 10.1111/j.1476-5381.2012.02207.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hillig KW, Mahlberg PG. A chemotaxonomic analysis of cannabinoid variation in Cannabis (Cannabaceae) Am J Bot. 2004;91:966–975. doi: 10.3732/ajb.91.6.966. [DOI] [PubMed] [Google Scholar]
- Jarrett MM, Limebeer CL, Parker LA. Effect of Delta9-tetrahydrocannabinol on sucrose palatability as measured by the taste reactivity test. Physiol Behav. 2005;86:475–479. doi: 10.1016/j.physbeh.2005.08.033. [DOI] [PubMed] [Google Scholar]
- Jarrett MM, Scantlebury J, Parker LA. Effect of delta9-tetrahydrocannabinol on quinine palatability and AM251 on sucrose and quinine palatability using the taste reactivity test. Physiol Behav. 2007;90:425–430. doi: 10.1016/j.physbeh.2006.10.003. [DOI] [PubMed] [Google Scholar]
- Kilkenny C, Browne W, Cuthill IC, Emerson M, Altman DG. Animal research: reporting in vivo experiments: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1577–1579. doi: 10.1111/j.1476-5381.2010.00872.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA. Delta-9-tetrahydrocannabinol interferes with the establishment and the expression of conditioned rejection reactions produced by cyclophosphamide: a rat model of nausea. Neuroreport. 1999;10:3769–3772. doi: 10.1097/00001756-199912160-00009. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA. The antiemetic drug ondansetron interferes with lithium-induced conditioned rejection reactions, but not lithium-induced taste avoidance in rats. J Exp Psychol Anim Behav Process. 2000;26:371–384. doi: 10.1037//0097-7403.26.4.371. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Parker LA. The 5-HT1A agonist 8-OH-DPAT dose-dependently interferes with the establishment and the expression of lithium-induced conditioned rejection reactions in rats. Psychopharmacology (Berl) 2003;166:120–126. doi: 10.1007/s00213-002-1309-6. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Litt DE, Parker LA. Effect of 5-HT3 antagonists and a 5-HT(1A) agonist on fluoxetine-induced conditioned gaping reactions in rats. Psychopharmacology (Berl) 2009;203:763–770. doi: 10.1007/s00213-008-1421-3. [DOI] [PubMed] [Google Scholar]
- Limebeer CL, Vemuri VK, Bedard H, Lang ST, Ossenkopp KP, Makriyannis A, et al. Inverse agonism of cannabinoid CB1 receptors potentiates LiCl-induced nausea in the conditioned gaping model in rats. Br J Pharmacol. 2010;161:336–349. doi: 10.1111/j.1476-5381.2010.00885.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGrath J, Drummond G, McLachlan E, Kilkenny C, Wainwright C. Guidelines for reporting experiments involving animals: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1573–1576. doi: 10.1111/j.1476-5381.2010.00873.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaughlin PJ, Winston KM, Limebeer CL, Parker LA, Makriyannis A, Salamone JD. The cannabinoid CB1 antagonist AM 251 produces food avoidance and behaviors associated with nausea but does not impair feeding efficiency in rats. Psychopharmacology (Berl) 2005;180:286–293. doi: 10.1007/s00213-005-2171-0. [DOI] [PubMed] [Google Scholar]
- de Mattos Viana B, Prais HA, Daker MV. Melancholic features related to rimonabant. Gen Hosp Psychiatry. 2009;31:583–585. doi: 10.1016/j.genhosppsych.2008.12.009. [DOI] [PubMed] [Google Scholar]
- Mechoulam R, Parker LA, Gallily R. Cannabidiol: an overview of some pharmacological aspects. J Clin Pharmacol. 2002;42:11S–19S. doi: 10.1002/j.1552-4604.2002.tb05998.x. [DOI] [PubMed] [Google Scholar]
- Merkus FW. Cannabivarin and tetrahydrocannabivarin, two new constituents of hashish. Nature. 1971;232:579–580. doi: 10.1038/232579a0. [DOI] [PubMed] [Google Scholar]
- O'Brien LD, Wills KL, Segsworth B, Dashney B, Rock EM, Limebeer CL, et al. Effect of chronic exposure to rimonabant and phytocannabinoids on anxiety-like behavior and saccharin palatability. Pharmacol Biochem Behav. 2013;103:597–602. doi: 10.1016/j.pbb.2012.10.008. [DOI] [PubMed] [Google Scholar]
- Orr LE, McKernan JF, Bloome B. Antiemetic effect of tetrahydrocannabinol. Compared with placebo and prochlorperazine in chemotherapy-associated nausea and emesis. Arch Intern Med. 1980;140:1431–1433. doi: 10.1001/archinte.140.11.1431. [DOI] [PubMed] [Google Scholar]
- Parker LA, Limebeer CL. Cannabinoids in the management of nausea and vomiting. In: Köfalvi A, editor. Cannabinoids and the Brain. New York: Springer; 2008. pp. 259–273. [Google Scholar]
- Parker LA, Mechoulam R. Cannabinoid agonists and antagonists modulate lithium-induced conditioned gaping in rats. Integr Physiol Behav Sci. 2003;38:133–145. doi: 10.1007/BF02688831. [DOI] [PubMed] [Google Scholar]
- Parker LA, Mechoulam R, Schlievert C. Cannabidiol, a non-psychoactive component of cannabis and its synthetic dimethylheptyl homolog suppress nausea in an experimental model with rats. Neuroreport. 2002;13:567–570. doi: 10.1097/00001756-200204160-00006. [DOI] [PubMed] [Google Scholar]
- Parker LA, Mechoulam R, Schlievert C, Abbott L, Fudge ML, Burton P. Effects of cannabinoids on lithium-induced conditioned rejection reactions in a rat model of nausea. Psychopharmacology (Berl) 2003;166:156–162. doi: 10.1007/s00213-002-1329-2. [DOI] [PubMed] [Google Scholar]
- Parker LA, Rana SA, Limebeer CL. Conditioned nausea in rats: assessment by conditioned disgust reactions, rather than conditioned taste avoidance. Can J Exp Psychol. 2008;62:198–209. doi: 10.1037/a0012531. [DOI] [PubMed] [Google Scholar]
- Parker LA, Limebeer CL, Rana SA. Conditioned disgust, but not conditioned taste avoidance, may reflect conditioned nausea in rats. In: Reilly S, Schachtman TR, editors. Conditioned Taste Aversions: Behavioral and Neural Processes. New York: Oxford University Press; 2009. [Google Scholar]
- Pertwee RG, Thomas A, Stevenson LA, Ross RA, Varvel SA, Lichtman AH, et al. The psychoactive plant cannabinoid, Delta9-tetrahydrocannabinol, is antagonized by Delta8- and Delta9-tetrahydrocannabivarin in mice in vivo. Br J Pharmacol. 2007;150:586–594. doi: 10.1038/sj.bjp.0707124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riedel G, Fadda P, McKillop-Smith S, Pertwee RG, Platt B, Robinson L. Synthetic and plant-derived cannabinoid receptor antagonists show hypophagic properties in fasted and non-fasted mice. Br J Pharmacol. 2009;156:1154–1166. doi: 10.1111/j.1476-5381.2008.00107.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock E, Bolognini D, Limebeer C, Cascio M, Anavi-Goffer S, Fletcher P, et al. Cannabidiol, a non-psychotropic component of cannabis, attenuates vomiting and nausea-like behaviour via indirect agonism of 5-HT(1A) somatodendritic autoreceptors in the dorsal raphe nucleus. Br J Pharmacol. 2012;165:2620–2634. doi: 10.1111/j.1476-5381.2011.01621.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock EM, Parker LA. Effect of low doses of cannabidiolic acid and ondansetron on LiCl-induced conditioned gaping (a model of nausea-induced behaviour) in rats. Br J Pharmacol. 2013;169:685–692. doi: 10.1111/bph.12162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rock EM, Limebeer CL, Mechoulam R, Piomelli D, Parker LA. The effect of cannabidiol and URB597 on conditioned gaping (a model of nausea) elicited by a lithium-paired context in the rat. Psychopharmacology (Berl) 2008;196:389–395. doi: 10.1007/s00213-007-0970-1. [DOI] [PubMed] [Google Scholar]
- Rock EM, Benzaquen J, Limebeer CL, Parker LA. Potential of the rat model of conditioned gaping to detect nausea produced by rolipram, a phosphodiesterase-4 (PDE4) inhibitor. Pharmacol Biochem Behav. 2009;91:537–541. doi: 10.1016/j.pbb.2008.09.005. [DOI] [PubMed] [Google Scholar]
- Sallan SE, Zinberg NE, Frei E., 3rd Antiemetic effect of delta-9-tetrahydrocannabinol in patients receiving cancer chemotherapy. N Engl J Med. 1975;293:795–797. doi: 10.1056/NEJM197510162931603. [DOI] [PubMed] [Google Scholar]
- Sharkey KA, Cristino L, Oland LD, Van Sickle MD, Starowicz K, Pittman QJ, et al. Arvanil, anandamide and N-arachidonoyl-dopamine (NADA) inhibit emesis through cannabinoid CB1 and vanilloid TRPV1 receptors in the ferret. Eur J Neurosci. 2007;25:2773–2785. doi: 10.1111/j.1460-9568.2007.05521.x. [DOI] [PubMed] [Google Scholar]
- Sink KS, McLaughlin PJ, Wood JA, Brown C, Fan P, Vemuri VK, et al. The novel cannabinoid CB1 receptor neutral antagonist AM4113 suppresses food intake and food-reinforced behavior but does not induce signs of nausea in rats. Neuropsychopharmacology. 2008;33:946–955. doi: 10.1038/sj.npp.1301476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sink KS, Vemuri VK, Wood J, Makriyannis A, Salamone JD. Oral bioavailability of the novel cannabinoid CB1 antagonist AM6527: effects on food-reinforced behavior and comparisons with AM4113. Pharmacol Biochem Behav. 2009;91:303–306. doi: 10.1016/j.pbb.2008.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sink KS, Segovia KN, Sink J, Randall PA, Collins LE, Correa M, et al. Potential anxiogenic effects of cannabinoid CB1 receptor antagonists/inverse agonists in rats: comparisons between AM4113, AM251, and the benzodiazepine inverse agonist FG-7142. Eur Neuropsychopharmacol. 2010;20:112–122. doi: 10.1016/j.euroneuro.2009.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweet DL, Miller NJ, Weddington W, Senay E, Sushelsky L. delta 9-Tetrahydrocannabinol as an antiemetic for patients receiving cancer chemotherapy. A pilot study. J Clin Pharmacol. 1981;21:70S–75S. doi: 10.1002/j.1552-4604.1981.tb02577.x. [DOI] [PubMed] [Google Scholar]
- Tam J, Vemuri VK, Liu J, Batkai S, Mukhopadhyay B, Godlewski G, et al. Peripheral CB1 cannabinoid receptor blockade improves cardiometabolic risk in mouse models of obesity. J Clin Invest. 2010;120:2953–2966. doi: 10.1172/JCI42551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas A, Stevenson LA, Wease KN, Price MR, Baillie G, Ross RA, et al. Evidence that the plant cannabinoid Delta9-tetrahydrocannabivarin is a cannabinoid CB1 and CB2 receptor antagonist. Br J Pharmacol. 2005;146:917–926. doi: 10.1038/sj.bjp.0706414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tubaro A, Giangaspero A, Sosa S, Negri R, Grassi G, Casano S, et al. Comparative topical anti-inflammatory activity of cannabinoids and cannabivarins. Fitoterapia. 2010;81:816–819. doi: 10.1016/j.fitote.2010.04.009. [DOI] [PubMed] [Google Scholar]
- Tuerke KJ, Leri F, Parker LA. Antidepressant-like effects of paroxetine are produced by lower doses than those which produce nausea. Pharmacol Biochem Behav. 2009;93:190–195. doi: 10.1016/j.pbb.2009.05.011. [DOI] [PubMed] [Google Scholar]