

Abstract
BACKGROUND AND PURPOSE
Epilepsy is the most prevalent neurological disease and is characterized by recurrent seizures. Here, we investigate (i) the anticonvulsant profiles of cannabis-derived botanical drug substances (BDSs) rich in cannabidivarin (CBDV) and containing cannabidiol (CBD) in acute in vivo seizure models and (ii) the binding of CBDV BDSs and their components at cannabinoid CB1 receptors.
EXPERIMENTAL APPROACH
The anticonvulsant profiles of two CBDV BDSs (50–422 mg·kg−1) were evaluated in three animal models of acute seizure. Purified CBDV and CBD were also evaluated in an isobolographic study to evaluate potential pharmacological interactions. CBDV BDS effects on motor function were also investigated using static beam and grip strength assays. Binding of CBDV BDSs to cannabinoid CB1 receptors was evaluated using displacement binding assays.
KEY RESULTS
CBDV BDSs exerted significant anticonvulsant effects in the pentylenetetrazole (≥100 mg·kg−1) and audiogenic seizure models (≥87 mg·kg−1), and suppressed pilocarpine-induced convulsions (≥100 mg·kg−1). The isobolographic study revealed that the anticonvulsant effects of purified CBDV and CBD were linearly additive when co-administered. Some motor effects of CBDV BDSs were observed on static beam performance; no effects on grip strength were found. The Δ9-tetrahydrocannabinol and Δ9-tetrahydrocannabivarin content of CBDV BDS accounted for its greater affinity for CB1 cannabinoid receptors than purified CBDV.
CONCLUSIONS AND IMPLICATIONS
CBDV BDSs exerted significant anticonvulsant effects in three models of seizure that were not mediated by the CB1 cannabinoid receptor and were of comparable efficacy with purified CBDV. These findings strongly support the further clinical development of CBDV BDSs for the treatment of epilepsy.
Keywords: seizure, epilepsy, cannabinoid, cannabidivarin, cannabidiol, anticonvulsant, tolerability, isobologram, radioligand binding assays
Introduction
Epilepsy is a chronic neurological disorder characterized by recurrent seizures, which affects approximately 50 million people worldwide (Leonardi and Ustun, 2002). Epilepsy's co-morbidities include cognitive decline, depressive disorders and schizophrenia (Hermann et al., 2000; Kanner et al., 2012), which are worsened by poorly controlled seizures (Perucca et al., 2000). There are many treatments available (BNF, 2011); however, all have notable side effects (Ortinski and Meador, 2004; Schachter, 2007) and ∼30% of the cases remain pharmacoresistant, resulting in poorly controlled seizures (Hitiris et al., 2007). This represents a major unmet clinical need for new well-tolerated antiepileptic drugs (AEDs) able to control previously pharmacoresistant epilepsies.
We and others have previously reported that cannabidiol (CBD), a non-psychoactive phytocannabinoid (pCB) of the cannabis plant, is anticonvulsant in several in vivo seizure models (Consroe et al., 1982; Wallace et al., 2001; Jones et al., 2010; 2012) and in humans (Cunha et al., 1980). Similarly, cannabidivarin (CBDV; the propyl analogue of CBD) is anticonvulsant in vivo (Hill et al., 2012a). While the anticonvulsant mechanisms of CBD and CBDV are unidentified, their anticonvulsant and tolerability profiles do not suggest interaction with the CB1 cannabinoid receptor (Wallace et al., 2001; Jones et al., 2010; Hill et al., 2012a).
An increasing body of evidence suggests that cannabis extracts enriched with a specific pCB can possess as much, or more, pharmacological efficacy/potency than the purified pCB (Wilkinson et al., 2003; Whalley et al., 2004; Ryan et al., 2006; De Petrocellis et al., 2011; Sagredo et al., 2011; Valdeolivas et al., 2012). As an example, amelioration of spasticity associated with multiple sclerosis by Δ9-tetrahydrocannabinol (THC) and CBD was greater in clinical trials when these two pCBs were combined as two plant extracts rather than as purified pCBs (Russo and Guy, 2006). The apparent benefits of extracts may arise from polypharmacological effects of the pCBs (McPartland and Russo, 2001) or the terpenoids present in cannabis extracts that possess their own pharmacology and can modulate pCB effects to enhance activity or reduce off-target effects (Formukong et al., 1988; McPartland and Russo, 2001), the ‘entourage effect’ (Russo, 2011).
Therefore, we extend our previous investigations of the anticonvulsant effects of CBDV and CBD (Jones et al., 2010; 2012; Hill et al., 2012a) to assess the anticonvulsant potential of extracts rich in CBDV and CBD [‘botanical drug substances’ (BDSs)]. Here, we test two such extracts: one with pharmacologically significant (Järbe et al., 2002; Hill et al., 2010) Δ9-THC and Δ9-tetrahydrocannabivarin (THCV) content (‘unmodified CBDV BDS’) and another with Δ9-THC and Δ9-THCV removed (‘modified CBDV BDS’). This allows assessment of the effects of Δ9-THC and Δ9-THCV on the anticonvulsant and tolerability profiles of the compounds; both Δ9-THC and Δ9-THCV can be anticonvulsant in their own right (Corcoran et al., 1973; Fried and McIntyre, 1973; Wallace et al., 2001; 2003; Hill et al., 2010), although there are also some reports that Δ9-THC can be proconvulsant (Chesher and Jackson, 1974; Karler and Turkanis, 1980).
Here, for the first time, we demonstrate significant anticonvulsant actions of CBDV BDSs in rat and mouse in vivo seizure models. Investigation of the interactions between the principal constituent pCBs indicated that CBD and CBDV act additively to suppress seizures. In motor assays, while unmodified CBDV BDS adversely affected balance and coordination at all doses, limited effects were only seen at the highest dose of modified CBDV BDS; neither CBDV BDS caused deficits in forelimb grip strength. Radioligand binding indicated that the Δ9-THC/Δ9-THCV content of the unmodified CBDV BDS was responsible for the majority of CB1 cannabinoid receptor binding; parallel behavioural experiments demonstrated that they were not required for anticonvulsant effects.
Methods
Animals
All animals were housed on a 12 h light–dark cycle, with food and water available ad libitum. Studies using male Wistar Kyoto rats (70–110 g; Harlan, Bicester, UK) were undertaken at the University of Reading. Mouse whole brain membranes for binding studies (University of Aberdeen) were obtained from adult (25–40 g) male MF1 mice (Harlan, Blackthorn, UK). In both cases, work was conducted in accordance with the Animals (Scientific Procedures) Act of 1986. Audiogenic seizure experiments used DBA/2 mice (10–14 g, 3–4 weeks old; Elevage Janvier, Le Genest-Saint-Isle, France), were designed and analysed by the authors, and performed by Porsolt Research Laboratory (Le Genest-Saint-Isle, France) in accordance with the French legislation under licence from the French Ministry for Agriculture and Fisheries. All animal work was carried out in accordance with the ARRIVE guidelines for reporting experiments involving animals (Kilkenny et al., 2010; McGrath et al., 2010); 595 rats and 290 mice were used for in vivo studies in total.
Drug administration
Animals received either cannabis-derived CBDV BDSs or purified pCBs. pCBs (10–200 mg·kg−1) and cannabis-derived CBDV BDSs (50–422 mg·kg−1; GW Pharmaceuticals Ltd, Salisbury, UK); were suspended in ethanol, Cremophor EL and saline [0.9% (w/v) NaCl, 2:1:17, respectively; all from Sigma-Aldrich, Poole, UK]; and administered via i.p. injection 1 h prior to experimental procedures to achieve brain Tmax (Deiana et al., 2012). Each experiment contained a control group that received vehicle, to which other groups were compared. In seizure experiments, group sizes were n = 10 for mice and n = 15 for rats. Unmodified CBDV BDS contained 47.4% CBDV, 13.9% CBD, 1% Δ9-THC and 2.5% Δ9-THCV; modified CBDV BDS lacked Δ9-THC/Δ9-THCV and contained 57.8% CBDV and 13.7% CBD; remaining content comprised plant matter. A CBDV BDS with most pCB content removed (termed BDS-pCB) contained no Δ9-THCV/Δ9-THC and 4.3% CBDV and 0.1% CBD. For clarity, in vivo experiments are numbered and detailed in Table 1 where the treatments used in each experiment and the doses of CBDV, CBD, Δ9-THC and Δ9-THCV received are outlined. The standardization and reproducibility of all CBDV BDSs employed in this study complied with the US Food and Drug Administration (FDA) guidelines for botanical drug products (FDA, 2004).
Table 1.
Experimental design and pCB content of in vivo Experiments 1.1–4.2
| Experiment/Figures | Compound | Doses (mg·kg−1) | CBDV/CBD content (mg·kg−1) | THC/THCV content (mg·kg−1) |
|---|---|---|---|---|
| Experiment 1.1, Figure 1A and 1B | Modified CBDV BDS | 50 | 29/7 | 0/0 |
| 100 | 58/14 | 0/0 | ||
| 200 | 116/27 | 0/0 | ||
| 275 | 159/38 | 0/0 | ||
| 346 | 200/47 | 0/0 | ||
| Experiment 1.2, Figure 1C and 1D | Purified CBDV | 50 | 50/0 | 0/0 |
| 100 | 100/0 | 0/0 | ||
| 200 | 200/0 | 0/0 | ||
| Modified CBDV BDS | 50 | 29/7 | 0/0 | |
| 100 | 58/14 | 0/0 | ||
| 200 | 116/27 | 0/0 | ||
| Experiment 1.3, Figure 1E and 1F | Purified CBDV + CBD | Low | 50/12 | 0/0 |
| Mid | 100/23 | 0/0 | ||
| High | 200/47 | 0/0 | ||
| Modified CBDV BDS | 50 | 50/12 | 0/0 | |
| 100 | 100/23 | 0/0 | ||
| 200 | 200/47 | 0/0 | ||
| Experiment 1.4, Figure 1G and 1H | pCB-free BDS | 50 | 2/<1 | 0/0 |
| 100 | 4/<1 | 0/0 | ||
| 200 | 9/<1 | 0/0 | ||
| Experiments 2.1 and 2.2, Figure 2A–F | Unmodified CBDV BDS | 150 | 71/21 | 2/4 |
| 275 | 130/38 | 3/7 | ||
| 422 | 200/59 | 4/11 | ||
| Modified CBDV BDS | 150 | 87/21 | 0/0 | |
| 275 | 159/38 | 0/0 | ||
| 346 | 200/47 | 0/0 | ||
| Experiment 3.1, Figure 3A | Unmodified CBDV BDS | 50 | 24/7 | 1/1 |
| 100 | 47/14 | 1/3 | ||
| 200 | 95/28 | 2/5 | ||
| Experiment 3.2, Figure 3B | Modified CBDV BDS | 50 | 29/7 | 0/0 |
| 100 | 58/14 | 0/0 | ||
| 200 | 116/27 | 0/0 | ||
| Experiment 4.1, Figure 4A | Unmodified CBDV BDS | 100 | 47/14 | 1/3 |
| 200 | 95/28 | 2/5 | ||
| 300 | 142/42 | 3/8 | ||
| Experiment 4.2, Figure 4B | Modified CBDV BDS | 200 | 116/27 | 0/0 |
| Unmodified CBDV BDS | 200 | 116/28 | 2/5 | |
| CBDV + CBD | – | 116/27 | 0/0 |
Rat seizure experiments
Pentylenetetrazole (PTZ; 85 mg·kg−1; Experiments 1.1–1.4 and 2.1) or pilocarpine (380 mg·kg−1; Experiments 4.1 and 4.2) was administered i.p. 1 h after pCB/CBDV BDS/vehicle treatment in 0.9% (w/v) NaCl. Methylscopolamine [1 mg·kg−1; in 0.9% (w/v) NaCl] was administered i.p. 45 min before pilocarpine to minimize pilocarpine's peripheral effects. Seizure activity was recorded (Farrimond et al., 2009) for 30 min (PTZ) or 1 h (pilocarpine); video records were blinded and subsequently coded offline using the Observer XT software (Noldus, Wageningen, the Netherlands) and modified Racine scales (PTZ: 0, normal behaviour; 0.5, abnormal behaviour; 1, isolated myoclonic jerk; 2, atypical clonic seizure; 3, bilateral forelimb clonus; 3.5, bilateral forelimb clonus with body twist; 4, tonic–clonic seizure with suppressed tonic phase; 5, fully developed tonic–clonic seizure. Pilocarpine: 0, normal behaviour; 1, mouth clonus; 2, unilateral forelimb clonus; 3, bilateral forelimb clonus; 4, bilateral forelimb clonus with rearing and falling; 4.5, tonic–clonic convulsions; 5, tonic–clonic convulsions with righting reflex lost). In pilocarpine experiments, purified CBDV was added to unmodified CBDV BDS to match the CBDV content of modified CBDV BDS.
Audiogenic seizures in mice (Experiments 3.1–3.4)
DBA/2 mice were individually placed in a Plexiglas jar (40 cm wide, 35 cm high) containing an electric bell that sounded for up to 60 s (110–120 dB) to induce seizure activity (wild running, clonic convulsions and tonic convulsions), which was recorded by experienced technicians during experiments (Jensen et al., 1983).
Isobolographic experimental design and analysis determines whether two compounds that exert similar (e.g. anticonvulsant) pharmacological effects behave synergistically, additively or antagonistically when co-administered (Tallarida, 2006). Here, this approach was applied using the audiogenic seizure model to investigate any pharmacological interactions between CBD and CBDV. Initially, purified CBDV and CBD (each 10–200 mg·kg−1, i.p.) were administered separately and their dose–response relationships (DRRs) calculated for protection against development of clonic convulsions (Experiment 3.3). CBDV and CBD were then co-administered in 1:1 effect size ratios (10 equally separated effect sizes, ED10–ED100) calculated from the previously calculated DRRs (Experiment 3.4). Thereafter, the individual DRRs of each pCB were used to create isoboles describing theoretical lines of additivity at multiple effect sizes (e.g. ED30, ED50) for the two compounds in combination. Finally, experimental effect sizes obtained from the co-administration study were statistically compared with the theoretical line of additivity on isoboles in accordance with the isobolographic method for full agonists described by Tallarida (2006).
Motor assays (Experiment 2.2)
Static beam and grip strength tasks were used as described in Jones et al. (2012) to assess motor function after administration of unmodified CBDV BDS (150, 275 or 422 mg·kg−1, i.p.), modified CBDV BDS (150, 275 or 346 mg·kg−1, i.p.) or CBDV BDS vehicle (control). Treatment order was randomized, and each animal (n = 10) received all treatments during the study with a minimum of 48 h between each treatment.
Radioligand binding
Materials
CP55940 was supplied by Tocris (Bristol, UK) and [3H]CP555940 (160 Ci·mmol−1) by PerkinElmer Life Sciences, Inc. (Boston, MA, USA).
CHO cells
CHO cells stably transfected with complementary DNA encoding human CB1 cannabinoid receptors (PerkinElmer Life Sciences, Inc.) were maintained in DMEM nutrient mixture F-12 HAM, supplemented with 1 mM L-glutamine, 10% FBS, 0.6% penicillin-streptomycin and Geneticin G418 (600 μg·mL−1). Cells were maintained at 37°C with 5% CO2 and were passaged twice a week using non-enzymatic cell dissociation solution (EDTA, 1 mM).
Membrane preparation
Binding assays with [3H]CP55940 were performed with membranes obtained from MF1 mouse whole brain or hCB1-CHO cells (Ross et al., 1999; Thomas et al., 2004) and frozen as a pellet at −20°C until required. Before use cells were defrosted, diluted in Tris buffer (50 mM Tris–HCl and 50 mM Tris–base) and homogenized with a 1 mL handheld homogenizer. Protein assays were performed using a Bio-Rad DC Kit (Hercules, CA, USA).
Radioligand displacement assay
Assays were carried out in Tris-binding buffer (50 mM Tris–HCl, 50 mM Tris–base, 0.1% BSA, pH 7.4), total assay volume 500 μL, using the filtration procedure described by Ross et al. (1999). Binding was initiated by addition of mouse brain membranes (33 μg protein per well) or hCB1-CHO cell membranes (50 μg protein per well). All assays were performed at 37°C for 60 min before termination by addition of ice cold Tris-binding buffer and vacuum filtration using a 24 well sampling manifold (Brandel Cell Harvester; Brandel Inc., Gaithersburg, MD, USA) and Brandel GF/B filters that had been soaked in wash buffer at 4°C for at least 24 h. Each reaction well was washed six times with Tris-binding buffer (1.2 mL). The filters were oven-dried for 60 min and placed in 5 mL of scintillation fluid (Ultima Gold XR, PerkinElmer, Seer Green, Buckinghamshire, UK). Radioactivity was quantified by liquid scintillation spectrometry. Specific binding was defined by the presence and absence of 1 μM unlabelled CP55940. The concentration of [3H]CP55940 used in our displacement assays was 0.7 nM. All CBDV BDSs and CBDV were stored as stock solutions (10 mM) in dimethyl sulphoxide (DMSO); the vehicle concentration in all assay wells was 0.1% DMSO. [3H]CP55940 binding parameters were 2336 fmol·mg−1 (Bmax) and 2.31 nM (Kd) in mouse brain membranes (Thomas et al., 2004), and 57.0 pmol·mg−1 (Bmax) and 1.1 nM (Kd) for human CB1 CHO cells.
Statistical analyses
Statistical procedures were performed using GraphPad Prism 5 (GraphPad Software, Inc., San Diego, CA, USA). Effects on seizure severity were analysed by Kruskal–Wallis with post hoc Mann–Whitney U-tests. Parameters reported as percentages were analysed by chi-squared tests with post hoc Fisher exact tests. Body temperatures were analysed using a Student's t-test. Grip strength, distance travelled and foot slips/metre parameters in motor assays were analysed using one-way anova with post hoc Tukey tests, performed on the daily averages of each animal. Isoboles were analysed by comparing the Cartesian coordinates for the experimental result with those of the two points on the theoretical line of additivity. The x coordinate was calculated by setting the y coordinate of the experimental value and solving for x on the line of additivity, and vice versa for the y coordinate. The three coordinates were compared using a chi-squared test. Binding assay values were expressed as means and variability as SEM or as 95% confidence limits. The concentrations of test compounds that produced a 50% displacement of radioligand from specific binding sites (IC50 values) were determined, and the corresponding Ki values were calculated using the equation of Cheng and Prusoff (1973). All receptor and ion channel nomenclature conforms to Alexander et al. (2011).
Results
Effects of modified CBDV BDS and constituents on PTZ-induced seizures in rats
Initially, we investigated the effects of modified CBDV BDS (containing no Δ9-THCV or Δ9-THC; Table 1) on PTZ-induced seizures in Experiments 1.1–1.4. The highest modified CBDV BDS dose was selected to contain the most effective anticonvulsant dose of purified CBDV tested to date (200 mg·kg−1; Hill et al., 2012a). In Experiment 1.1, modified CBDV BDS had a significant anticonvulsant effect on seizure severity (U = 13.84, P ≤ 0.05; Figure 1A); 200 and 275 mg·kg−1 significantly suppressed the observed severity (P ≤ 0.05). Seizure-associated mortality was also affected by modified CBDV BDS administration [χ2(5) = 29.97, P ≤ 0.01; Figure 1B]; ≥200 mg·kg−1 significantly reduced mortality (P ≤ 0.05). We then compared the anticonvulsant effects of modified CBDV BDS with those of purified CBDV in Experiment 1.2. Administration of purified CBDV or modified CBDV BDS, matched by absolute weight, significantly affected the observed seizure severity (U = 12.72, P ≤ 0.05; Figure 1C); both ≥50 mg·kg−1 of purified CBDV (P ≤ 0.05) and 100 mg·kg−1 (P ≤ 0.01) of modified CBDV BDS significantly suppressed seizure severity; additionally, 200 mg·kg−1 of modified CBDV BDS produced a strong trend (P ≤ 0.1) towards seizure severity suppression. Mortality was also significantly reduced by purified CBDV and modified CBDV BDS [χ2(6) = 21.05, P ≤ 0.01; Figure 1D], with significant reductions caused by administration of ≥100 mg·kg−1 (P ≤ 0.01) of either substance.
Figure 1.

Effects of CBDV BDS components in the PTZ model of acute convulsion. (A, B) Experiment 1.1: dose response of modified BDS. (C, D) Experiment 1.2: purified CBDV compared with modified BDS. (E, F) Experiment 1.3: comparison of modified CBDV BDS against matching levels of CBDV and CBD. (G, H) Experiment 1.4: BDS-pCB. (A, C, E, G) Maximum observed convulsion severity (median severity in grey, box represents interquartile range, whiskers represent maxima and minima (Kruskal–Wallis test, with a post hoc Mann–Whitney U-tests). (B, D, F, H) Mortality (chi-squared test, with post hoc Fisher exact test); n = 15 for each dose; #P ≤ 0.1, *P ≤ 0.05, **P ≤ 0.01. In all panels ‘V’ represents vehicle treatment.
As 100 mg·kg−1 modified CBDV BDS and purified CBDV each suppressed seizure activity to a similar extent despite modified CBDV BDS containing less CBDV, we proceeded to compare the anticonvulsant properties of the modified CBDV BDS with combinations of purified CBD and CBDV to assess potential benefits of BDSs in Experiment 1.3. Administration of both modified CBDV BDS and matched levels of purified CBDV plus CBD caused a trend in the observed seizure severity (U = 12.28, P ≤ 0.1; Figure 1E), where the highest dose of the purified pCBs (P ≤ 0.05) and middle dose of the modified CBDV BDS (P ≤ 0.01) significantly suppressed seizure severity. Mortality was significantly affected by administration of modified CBDV BDS or matched doses of pCBs [χ2(6) = 26.81, P ≤ 0.001; Figure 1F], where middle or greater purified pCBs (P ≤ 0.05) and all modified CBDV BDS (P ≤ 0.05) doses significantly reduced mortality. In a further experiment (Experiment 1.4), we examined the effects of the BDS-pCB in the PTZ model of seizure; only a near-significant trend in seizure severity (U = 6.555, P ≤ 0.1; Figure 1G) was seen due to 100 mg·kg−1, suggesting an increase in seizure severity (P ≤ 0.1) but with no effect on mortality (Figure 1H).
Thus, when these data are considered as a whole, modified CBDV BDS had a strong anticonvulsant effect in the PTZ model of seizures, producing comparable anticonvulsant effects as purified constituent pCBs (Figure 1E–G), but with lower pCB levels.
Effects of modified and unmodified CBDV BDS on PTZ-induced convulsions and motor function
Next, we compared modified and unmodified CBDV BDSs to determine whether the presence of Δ9-THC and Δ9-THCV affects the anticonvulsant profile of CBDV BDS (Experiment 2.1; Table 1). Administration of both modified and unmodified CBDV BDS significantly affected the observed seizure severity (U = 21.57, P ≤ 0.01; Figure 2A); seizure severity was significantly suppressed by ≥150 mg·kg−1 of both unmodified (P ≥ 0.05) and modified (P ≥ 0.01) CBDV BDS. Mortality was also significantly affected by CBDV BDS administration [χ2(6) = 26.81, P ≤ 0.001; Figure 2B]; 150–275 mg·kg−1 (P ≤ 0.05) of unmodified and ≥150 mg·kg−1 (P ≤ 0.05) of modified CBDV BDS significantly reduced mortality.
Figure 2.

Anticonvulsant and motor effects of modified and unmodified BDSs. (A, B) Experiment 2.1: effect of BDSs on the severity (A) and associated mortality (B) of PTZ-induced convulsion. In (A) median severity is shown in grey, box represents interquartile range and whiskers represent maxima and minima (Kruskal–Wallis test, with a post hoc Mann–Whitney U-tests); in (B), mortality is given as a percentage (chi-squared test, with post hoc Fisher exact test). (C–F) Experiment 2.2: side effect profile of BDSs in motor assays. (C–E) Performance on the static beam assay showing the failure rate (C; chi squared, Fisher exact post hoc test), the mean number of foot slips per metre (D) and the mean distance covered (E) after treatment with BDS or vehicle. (F) Mean forelimb grip strength (kgf). Data in (D–F) presented as mean ± SEM and analysed by anova with Tukey post hoc test. (A, B) n = 15, (C–F) n = 10; *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001.
We next evaluated the effects of modified and unmodified CBDV BDSs on motor function, using static beam and grip strength assays (Experiment 2.2; Table 1). CBDV BDS significantly affected the number of animals that failed the static beam [χ2(6) = 15.77, P ≤ 0.05; Figure 2C]; ≥150 mg·kg−1 of unmodified (P ≤ 0.05) and 346 mg·kg−1 of modified (P ≤ 0.01) CBDV BDS significantly increased the failure rate. Notably, the greater failure rate of unmodified CBDV BDS-treated animals arose from exceeding the time limit. The foot slips/metre and distance travelled on the static beam were unaffected by either modified or unmodified CBDV BDS, as was forelimb grip strength. The data presented in Figure 2 taken as a whole demonstrate that CBDV BDS anticonvulsant activity is unaffected by the presence or absence of Δ9-THC and Δ9-THCV. However, poor static beam performance following a lower dose of unmodified CBDV BDS than modified CBDV BDS indicates that, despite their opposing pharmacological actions at CB1 cannabinoid receptors (see the Discussion section), Δ9-THC and/or Δ9-THCV content negatively affects motor.
Modified and unmodified CBDV BDSs and isobolographic analysis of CBDV and CBD in the audiogenic seizure model
Having demonstrated significant anticonvulsant effects of all CBDV BDSs in the PTZ model, we next evaluated their anticonvulsant activity in the audiogenic model of generalized seizure in mouse (Experiments 3.1 and 3.2).
In Experiment 3.1 (Table 1), the unmodified CBDV BDS significantly affected the proportion of animals that developed: wild running [χ2(3) = 19.55, P ≤ 0.001; Figure 3A], where ≥100 mg·kg−1 significantly reduced incidence (P ≤ 0.01); clonic convulsions [χ2(3) = 17.94, P ≤ 0.001; Figure 3A], where ≥100 mg·kg−1 significantly reduced incidence (P ≤ 0.01); and tonic convulsions [χ2(3) = 17.14, P ≤ 0.001; Figure 3A], where ≥50 mg·kg−1 (P ≤ 0.05) significantly reduced their incidence. Body temperature was significantly lower in animals administered 100 mg·kg−1 [36.5°C ± 0.2; t(18) = 3.28, P ≤ 0.01] and 200 mg·kg−1 [34.5°C ± 0.2; t(18) = 4.78, P ≤ 0.001] of unmodified CBDV BDS than vehicle (37.3°C ± 0.1). Thereafter, we tested the modified CBDV BDS in Experiment 3.2 (Table 1), where administration significantly affected: wild running [χ2(3) = 26.81, P ≤ 0.001; Figure 3B], where ≥50 mg·kg−1 significantly reduced incidence (P ≤ 0.01); and clonic convulsions [χ2(3) = 21.18, P ≤ 0.001; Figure 3B], where ≥50 mg·kg−1 (P ≤ 0.05) significantly reduced incidence. Body temperature was significantly lower in animals administered 200 mg·kg−1 unmodified CBDV BDS [33.7°C ± 0.6; t(18) = 5.73, P ≤ 0.001] when compared with vehicle-treated animals (37.2°C ± 0.2).
Figure 3.

Effects of CBDV BDS modified and unmodified, and isobolographic study of CBDV and CBD. (A, B) Experiments 3.1–3.2: percentage of animals exhibiting each convulsion parameter (WR, wild running; clonic, clonic convulsions; tonic, tonic convulsions; chi-squared test, with post hoc Fisher exact test) for unmodified BDS (A) and modified BDS (B). (C–E) Experiments 3.3–3.4: isobolographic determination of CBDV and CBD interactions. (C) Dose–response relationships of CBD and CBDV. (D) ED50 isobole of CBDV and CBD with theoretical and actual ED50s marked (chi-squared test). (E) Results from co-administration study and predicted results based on individual pCB studies (two-way anova); n = 10; *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001.
As both modified and unmodified CBDV BDS reduced seizure activity at lower doses than purified cannabinoids in the PTZ model of seizure, we also used the tractable nature of the audiogenic seizure model to investigate any therapeutic interaction between CBDV and CBD using an isobolographic approach (Experiments 3.3 and 3.4). Firstly, in Experiment 3.3 we demonstrated that purified CBDV and CBD both reduced clonic convulsion incidence [χ2(8) = 34.21, P ≤ 0.001; Figure 3C], where ≥100 mg·kg−1 of both CBDV (P ≤ 0.05) and CBD (P ≤ 0.05) significantly reduced incidence. Furthermore, calculation of respective ED50 values revealed that CBDV (64 mg·kg−1) was more potent than CBD (80 mg·kg−1). The clonic convulsion parameter was selected to evaluate seizure suppression due to the linear DRRs, and each dose producing a unique effect (Tallarida, 2006). As DRRs were significantly non-parallel [F(1,4) = 121.63, P ≤ 0.01; Figure 3C], in Experiment 3.4 we co-administered CBDV and CBD in a 1:1 dose effect ratio using 10 theoretical effect sizes (ED10–ED100) derived from the DRRs of the purified CBDV and CBD in Figure 3C. The ED50 of co-administered CBDV and CBD did not differ significantly from the theoretical line of additivity [χ2(2) = 3.44, P ≥ 0.1; Figure 3D; see the Methods section] plotted between the ED50s of each drug when administered in isolation, thereby indicating an additive anticonvulsant action when the two compounds are combined at this effect size. Moreover, this additive interaction was present across the entire dose range, with no significant difference between the theoretical additive DRR [F(1,9) = 0.49, P ≥ 0.1; Figure 3E], derived from the pCBs in isolation, and the experimental DRR follows co-administration.
As a whole, these results demonstrate that modified and unmodified CBDV BDSs each have strong anticonvulsant activity in the mouse audiogenic seizure and complement the data produced in the PTZ seizure model. In addition, we demonstrate that anticonvulsant properties of CBDV and CBD are additive in the mouse audiogenic seizure model.
Effects of CBDV BDSs on pilocarpine-induced convulsions in rats
Both modified and unmodified CBDV BDSs were investigated in the pilocarpine model of acute, temporal lobe convulsion (Experiments 4.1 and 4.2; Table 1). In Experiment 4.1, unmodified CBDV BDS significantly affected convulsion severity (U = 13.15, P ≤ 0.01; Figure 4A); ≥100 mg·kg−1 significantly reduced severity (P ≥ 0.05). No effect on mortality was observed (data not shown). In Experiment 4.2, using the dose at which unmodified CBDV BDS exerted its optimal anticonvulsant effect (200 mg·kg−1; see Figure 4A), the effects of modified and unmodified CBDV BDSs and co-administered purified pCBs with matching doses of CBDV and CBD were compared. Drug treatment significantly affected convulsion severity (U = 10.64, P ≤ 0.05; Figure 4A); unmodified CBDV BDS (P ≤ 0.05) and purified pCBs (P ≤ 0.001) reduced severity, and modified CBDV BDS produced a trend towards severity reduction (P ≤ 0.1). Drug administration produced a trend towards a reduction in mortality [χ2(3) = 6.67, P ≤ 0.1], where unmodified CBDV BDS reduced mortality (P ≤ 0.1; data not shown).
Figure 4.
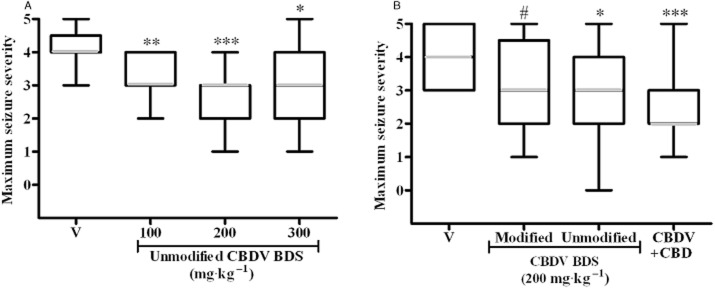
Modified and unmodified CBDV BDS, and purified cannabinoids in the acute pilocarpine model in rat. (A) Experiment 4.1: unmodified CBDV BDS. (B) Experiment 4.2: modified and unmodified CBDV BDS, and matched doses of pure CBDV and CBD. Both panels show maximum observed seizure severity (median severity in grey, box represents interquartile range, whiskers represent maxima and minima; Kruskal–Wallis test, with a post hoc Mann–Whitney U-tests). n = 15; #P ≤ 0.1, *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001.
Radioligand binding assays
First, displacement binding assays to compare the pharmacological profiles of unmodified CBDV BDS with purified CBDV were performed. Unmodified CBDV BDS showed greater affinity for CB1 cannabinoid receptors than purified CBDV in both MF1 mouse brain and hCB1-CHO cell membranes (Figure 5A,B and Table 2); purified CBDV only displaced [3H]CP55940 at the highest concentration tested (10 μM). Next, to investigate the greater affinity of unmodified CBDV BDS over purified CBDV for CB1 cannabinoid receptors, displacement binding assays using MF1 whole brain membranes were performed with unmodified CBDV BDS in parallel with the modified CBDV BDS. Interestingly, removal of Δ9-THC and Δ9-THCV did not significantly affect the potency with which modified CBDV BDS displaced [3H]CP55940 (Figure 5C and Table 2). However, the mean displacement, at the highest concentrations tested (1 and 10 μM), was significantly reduced by this removal (P < 0.05) (Figure 5C and Table 2). To further assess the contribution made by Δ9-THC plus Δ9-THCV to the potency with which unmodified CBDV BDS displaced [3H]CP55940, we tested unmodified CBDV BDS in parallel with BDS-pCB to which a pCB (CBDV, Δ9-THCV or Δ9-THC) had been added. The resulting percentage concentration of the added pCB was the same as the percentage concentration of this pCB in unmodified CBDV BDS. Interestingly, we found that BDS-pCB + purified CBDV displaced [3H]CP55940 with significantly less potency than unmodified CBDV BDS (Figure 5D and Table 3). The addition of Δ9-THCV to BDS-pCB resulted in this BDS displacing [3H]CP55940 with slightly, but not significantly, less potency than unmodified CBDV BDS (Figure 5E and Table 3). Addition of Δ9-THC had no statistically significant effect on the potency or maximum percentage of displacement with which unmodified CBDV BDS displaced [3H]CP55940 from mouse brain membranes (Figure 5F and Table 3). Taken together, these results suggest that both Δ9-THC and Δ9-THCV together in the unmodified CBDV BDS accounts for its ability to displace [3H]CP55940 with greater potency than purified CBDV from specific binding sites in membranes from both MF1 whole brain and human CB1-CHO cells.
Figure 5.

Radioligand binding properties of CBDV BDSs. Displacement of [3H]CP55940 by pure CBDV and unmodified CBDV BDS from (A) specific binding sites on MF1 mouse brain membranes and (B) hCB1-CHO cell membranes. Displacement of [3H]CP55940 by (C) unmodified CBDV BDS and modified CBDV BDS, (D) unmodified CBDV BDS and BDS-pCB plus pure CBDV, (E) unmodified CBDV BDS and BDS-pCB plus purified Δ9-THCV, and (F) unmodified CBDV BDS and BDS-pCB plus purified Δ9-THC from specific binding sites on MF1 mouse brain membranes. Symbols represent mean values ± SEM. Modified CBDV BDS lacks both Δ9-THC and Δ9-THCV. n = 4 in all cases.
Table 2.
Displacement of [3H]CP55940 by (A) pure CBDV and unmodified CBDV BDS, and (B) unmodified CBDV BDS and modified CBDV BDS from specific binding sites on MF1 brain membranes
| Experiment | Compound | Ki (nM) (95% CL) | Maximum % displacement (95% CL) | n |
|---|---|---|---|---|
| A* | Pure CBDV | No plateau | – | 4 |
| A* | Unmodified CBDV BDS | 127.6 (52.5 and 310.2) | 113.6 (98.2 and 129.0) | 4 |
| B | Unmodified CBDV BDS | 722.6 (403.7 and 1293) | 98.6 (83.8 and 113.4) | 4 |
| B | Modified CBDV BDS | 1010 (223.5 and 4564) | 55.5 (33.4 and 77.6) | 4 |
Mean Ki and maximum % displacement values for pure CBDV and unmodified CBDV BDS in hCB1-CHO cell membranes were not significantly different from those obtained in MF1 whole mouse brain membranes (see Results and Figure 5A,B).
CL, confidence limit.
Table 3.
Displacement of [3H]CP55940 by (A) unmodified CBDV BDS and BDS-pCB + pure CBDV, (B) unmodified CBDV BDS and BDS-pCB + pure THCV, and (C) unmodified CBDV BDS and BDS-pCB + pure THC from specific binding sites on MF1 brain membranes
| Experiment | Compound | Ki (nM) (95% CL) | Maximum % displacement (95% CL) | n |
|---|---|---|---|---|
| A | Unmodified CBDV BDS | 300.8 (159.2 and 568.5) | 89.2 (78.7 and 99.7) | 4 |
| A | BDS-pCB + pure CBDV | 2151 (643.9 and 7184) | 80.2 (51.8 and 108.6) | 4 |
| B | Unmodified CBDV BDS | 363.9 (141.4 and 936.5) | 85.8 (70.4 and 101.3) | 4 |
| B | BDS-pCB + pure THCV | 960.0 (203.8 and 4523) | 73.0 (44.6 and 101.3) | 4 |
| C | Unmodified CBDV BDS | 604.5 (252.5 and 1447) | 99.5 (79.7 and 119.3) | 4 |
| C | BDS-pCB + pure THC | 861.7 (195.1 and 3805) | 86.5 (57.4 and 115.6) | 4 |
CL, confidence limit.
Discussion
Here, for the first time, we demonstrate that cannabis extracts rich in CBDV and CBD can exert significant anticonvulsant effects. Importantly, these effects were observed in three separate models of seizure across two species. Both the modified and the unmodified CBDV BDSs had broadly similar efficacies to purified CBDV as well as CBDV and CBD in combination. Thereafter, we determined that CBDV and CBD act together in an additive manner against audiogenic seizures. While the presence of Δ9-THC and Δ9-THCV in the extracts did not affect anticonvulsant profile, their presence adversely affected motor function and was responsible for the displacement of CB1 cannabinoid receptor ligands in radioligand binding assays. Additionally, this is the first study to examine CBDV binding at the CB1 cannabinoid receptor where affinity was found to be very limited.
Anticonvulsant effects of CBDV BDSs
The modified CBDV BDS was investigated first as Δ9-THC at sufficiently high doses can induce psychoactive effects via the CB1 cannabinoid receptor, an undesirable clinical side effect. Although Δ9-THCV, a neutral CB1 cannabinoid receptor antagonist, can exert limited anticonvulsant effects, the clinical profile of such compounds remains unclear but is likely to be distinct from that of inverse agonists, for example, rimonabant (Pertwee, 2005).
Modified CBDV BDS dose-dependently reduced seizure severity and mortality, clearly demonstrating that the modified CBDV BDS was anticonvulsant. When purified CBDV and the modified CBDV BDS were compared, both reduced seizure severity and mortality; CBDV affected severity at a lower dose than modified CBDV BDS, but doses suppressing mortality were comparable. Subsequently, instead of comparing by absolute weight to the principal pCB, we compared by CBDV and CBD content to see if co-administered CBDV and CBD further reduced seizure severity, a broadly similar anticonvulsant effect resulted. In addition, modified CBDV BDS significantly reduced mortality at a lower dose than purified pCBs, a possible benefit of the BDS over purified pCBs. However, in Experiments 1.1–1.3 and 2.1, the highest doses of both the modified and the unmodified CBDV BDSs did not appear as efficacious as the preceding dose. While a definitive cause for this remains to be determined, this effect was not due to any significant pro-convulsant activity of the non-cannabinoid fraction (see Experiment 1.4). However, the complex nature (∼400 discrete non-cannabinoid constituents) of standardized cannabis extracts (Elsohly and Slade, 2005) means that concentrations of some constituents that could affect cannabinoid pharmacokinetics might only appear at higher doses to produce the differences seen.
Overall, these results indicate that CBDV BDS could be efficacious in the treatment of generalized seizures (Löscher, 2011), and its anticonvulsant activity against PTZ-induced seizures justifies further investigation of its utility against absence seizures using absence epilepsy models such as the GAERS and WAG/Rij rats (Coenen et al., 1992; Marescaux and Vergnes, 1995; Hosford and Wang, 1997).
Both unmodified and modified CBDV BDS reduced seizure severity when compared directly, with little appreciable difference in efficacy in the PTZ model. However, when the static beam task was employed to evaluate any motor side effects of the modified and unmodified CBDV BDSs, a drug-induced increase in the number of animals failing the task was seen. This effect was observed for all doses of unmodified CBDV BDS, but only the highest dose of modified CBDV BDS, suggesting that Δ9-THC and/or Δ9-THCV were responsible for a significant proportion of the observed motor deficits. The majority of unmodified CBDV BDS-treated animals failed due to the task's 5 min time limit being exceeded as animals remained stationary. This is consistent with previous reports on the effects of Δ9-THC on motor function (Järbe et al., 2002) and further suggests that the Δ9-THCV present in the unmodified CBDV BDS may have been insufficient to overcome Δ9-THC-mediated CB1 cannabinoid receptor partial agonism. In contrast to the effects on static beam performance, neither CBDV BDS elicited grip strength deficits, a test for muscle relaxation (Nevins et al., 1993) and, putatively, functional neurotoxicity (Sed et al., 2008). Many currently available AEDs produce notable side effects in people with epilepsy, including motor function deficits (Schachter, 2007), which reduce quality of life. These deficits are often also seen in non-clinical species (Löscher, 2011), potentially reducing the observed seizure severity in models of seizure (Hill et al., 2012a).
Cannabinoid and non-cannabinoid interactions in BDS
Cannabis-based BDSs have been reported to possess different pharmacological activity to their principal pCB constituent, where the presence of the non-principal pCBs and pCB-free fraction can enhance or decrease activity in some in vitro assays (De Petrocellis et al., 2011). Given this a priori knowledge and because the results obtained from our comparisons of CBDV BDS and purified pCB effects in seizure models suggested possible advantages of CBDV BDSs, we also investigated the actions and interactions of the pCB and pCB-free components of CBDV BDSs. Importantly, we showed that the BDS-pCB exerted no significant effect upon seizure severity in the PTZ model of seizure. In order to investigate interactions between CBDV and CBD, we employed the mouse audiogenic seizure model. Prior to the isobolographic study, we confirmed a dose-dependent anticonvulsant effect of the modified and unmodified CBDV BDSs in the audiogenic model comparable to that in PTZ. A core temperature below 34.5°C in DBA/2 mice can reduce seizure incidence in the audiogenic model (Essman and Sudak, 1964); however, significant reductions in seizure severity were seen in animals with body temperatures higher than this threshold, indicating that anticonvulsant effects were not primarily due to hypothermia. Using the isobolographic study design, we demonstrated an additive anticonvulsant effect of purified CBDV and CBD co-administration; there were no deleterious effects from co-administering these pCBs.
We have previously reported that administration of ≤200 mg·kg−1 CBDV did not produce anticonvulsant effects in the pilocarpine model (Hill et al., 2012a) except in highly powered experiments (n = 60 CBDV-treated animals), and that CBD did not affect convulsion severity in this model despite reduced tonic–clonic seizure incidence (Jones et al., 2012). In contrast, here we have demonstrated that ≥116 mg·kg−1 CBDV and ≥27 mg·kg−1 CBD co-administered as unmodified CBDV BDS or a combination of purified pCBs significantly reduced convulsion severity, demonstrating a clear advantage to combinatorial use of CBDV and CBD to treat temporal lobe convulsions in this model.
CB1 cannabinoid receptor binding of CBDV BDSs
In MF1 mouse whole brain membranes, we found that unmodified CBDV BDS showed greater affinity for CB1 cannabinoid receptors than purified CBDV, with mean Ki and maximum percentage displacement values not significantly different from those obtained using CHO cells over-expressing the human CB1 cannabinoid receptor. Purified CBDV had little affinity for CB1 cannabinoid receptors, with only the highest concentration tested (10 μM) exhibiting an effect. Our results suggest that the unmodified CBDV BDS's higher potency is due to its ability to bind to CB1 receptors, and not other GPCRs present in the brain. Unmodified CBDV BDS contains a significant percentage of CBD, Δ9-THC and Δ9-THCV. Although we and others have reported that CBD binds to CB1 receptors with only very weak affinity (mean Ki values in the micromolar range; Pertwee, 2008), both Δ9-THC and Δ9-THCV are potent CB1 cannabinoid receptor ligands (mean Ki values in the nanomolar range; Pertwee, 2008). Interestingly we found that (i) although the modified CBDV BDS's affinity was not significantly affected by the removal of Δ9-THC and Δ9-THCV, its maximum percentage displacement was significantly lower than that of unmodified CBDV BDS; (ii) the addition of Δ9-THC or Δ9-THCV to the BDS-pCB had no statistically significant effect on the ability of this BDS to displace [3H]CP55940 from mouse brain membranes compared with unmodified CBDV-BDS. In contrast (Figure 5D), the BDS-pCB plus CBDV mixture displaced [3H]CP55940 with significantly less potency than unmodified CBDV-BDS, and is consistent with our finding that purified CBDV binds to CB1 cannabinoid receptors with only very weak affinity (limited effect and no plateau was reached at the highest concentration tested). Together, these in vitro results suggest that the presence of both Δ9-THC and Δ9-THCV accounts for the ability of unmodified CBDV BDS to displace [3H]CP55940 with higher affinity than purified CBDV. Furthermore, as modified and unmodified CBDV BDS display a similar anticonvulsant action in vivo, the binding data suggest that the anticonvulsant mechanism of action of the CBDV BDSs are not CB1 cannabinoid receptor mediated. Despite CBD inhibiting ion channel function (e.g. T-type Ca2+ channels; Ross et al., 2008) and increasing inhibitory activity at some GPCRs (e.g. 5-HT1A; Russo et al., 2005), the specific molecular target(s) underlying CBD's anticonvulsant effects remain unconfirmed (Hill et al., 2012b). While limited structural changes can profoundly affect cannabinoid pharmacology (e.g. Δ9-THC and Δ9-THCV at CB1 cannabinoid receptors), it is not unreasonable to suggest that CBDV may share some pharmacological properties with CBD. CBD and CBDV have agonist effects at TRPA1, TRPV1 and TRPV2 receptors, and antagonistic properties at TRPM8 (De Petrocellis et al., 2011). Although TRPV1 expression has been reported in the brain (Tóth et al., 2005), as with other TRP receptors to which CBDV can bind, its functional role in health and disease is unclear. CBDV has also been reported to inhibit diacylglycerol lipase-α (De Petrocellis et al., 2011), an enzyme responsible for the synthesis of the endocannabinoid, 2-arachidonoylglycerol, and to stimulate fibroblast formation via a mechanism that can be functionally blocked by concurrent CB2 cannabinoid receptor antagonism (Scutt and Williamson, 2007). However, no evidence yet exists to link any of these mechanisms to CBDV's anticonvulsant activity.
Conclusions
In conclusion, these results demonstrate a strong CB1 cannabinoid receptor-independent anticonvulsant action of both modified and unmodified CBDV BDSs in three models of seizure, across two species. While unmodified CBDV BDS negatively affected performance in one motor function task, this was in stark contrast to the modified CBDV BDS that was well tolerated at anticonvulsant doses. This indicates that both modified and unmodified CBDV BDSs have potential to treat generalized or temporal lobe seizures with the modified CBDV BDS being best tolerated.
Acknowledgments
The authors thank GW Pharmaceuticals and Otsuka Pharmaceuticals for research sponsorship and provision of cannabinoids, and Mrs. Lesley A. Stevenson for technical support. We are also grateful to Professor Stephen Wright for critical comments on the manuscript.
Glossary
- AED
antiepileptic drug
- BDS
botanical drug substance
- CBD
cannabidiol
- CBDV
cannabidivarin
- pCB
phytocannabinoid
- PTZ
pentylenetetrazole
- Δ9-THC
Δ9-tetrahydrocannabinol
- Δ9-THCV
Δ9-tetrahydrocannabivarin
Conflict of Interest
The work reported was funded by grants to BJW, CMW and RGP from GW Pharmaceuticals and Otsuka Pharmaceuticals. MD is a GW Pharmaceuticals employee.
References
- Alexander SPH, Mathie A, Peters JA. Guide to receptors and channels (GRAC) Br J Pharmacol. 2011;164:S1–S324. doi: 10.1111/j.1476-5381.2011.01649_1.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BNF. British National Formulary. 62nd edn. London: British Medical Association and Royal Pharmaceutical Society of Great Britain; 2011. [Google Scholar]
- Cheng YC, Prusoff WH. Relationship between the inhibition constant (KI) and the concentration of inhibitor which causes 50 percent inhibition (IC50) of an enzymatic reaction. Biochem Pharmacol. 1973;22:3099–3108. doi: 10.1016/0006-2952(73)90196-2. [DOI] [PubMed] [Google Scholar]
- Chesher G, Jackson D. Anticonvulsant effects of cannabinoids in mice: drug interactions within cannabinoids and cannabinoid interactions with phenytoin. Psychopharmacology (Berl) 1974;37:255–264. doi: 10.1007/BF00421539. [DOI] [PubMed] [Google Scholar]
- Coenen A, Drinkenburg W, Inoue M, Van Luijtelaar E. Genetic models of absence epilepsy, with emphasis on the WAG/Rij strain of rats. Epilepsy Res. 1992;12:75–86. doi: 10.1016/0920-1211(92)90029-s. [DOI] [PubMed] [Google Scholar]
- Consroe P, Benedito MAC, Leite JR, Carlini EA, Mechoulam R. Effects of cannabidiol on behavioral seizures caused by convulsant drugs or current in mice. Eur J Pharmacol. 1982;83:293–298. doi: 10.1016/0014-2999(82)90264-3. [DOI] [PubMed] [Google Scholar]
- Corcoran ME, McCaughran JJA, Wada JA. Acute antiepileptic effects of Δ9-tetrahydrocannabinol in rats with kindled seizures. Exp Neurol. 1973;40:471–483. doi: 10.1016/0014-4886(73)90088-5. [DOI] [PubMed] [Google Scholar]
- Cunha JM, Carlini EA, Pereira AE, Ramos OL, Pimentel C, Gagliardi R, et al. Chronic administration of cannabidiol to healthy volunteers and epileptic patients. Pharmacology. 1980;21:175–185. doi: 10.1159/000137430. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Ligresti A, Moriello AS, Allarà M, Bisogno T, Petrosino S, et al. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol. 2011;163:1479–1494. doi: 10.1111/j.1476-5381.2010.01166.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deiana S, Watanabe A, Yamasaki Y, Amada N, Arthur M, Fleming S, et al. Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Δ 9-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive–compulsive behaviour. Psychopharmacology (Berl) 2012;219:859–873. doi: 10.1007/s00213-011-2415-0. [DOI] [PubMed] [Google Scholar]
- Elsohly MA, Slade D. Chemical constituents of marijuana: the complex mixture of natural cannabinoids. Life Sci. 2005;78:539–548. doi: 10.1016/j.lfs.2005.09.011. [DOI] [PubMed] [Google Scholar]
- Essman WB, Sudak FN. Audiogenic seizure in genetically susceptible mice: relation of hypothermia to onset and susceptibility. Exp Neurol. 1964;9:228–235. doi: 10.1016/0014-4886(64)90019-6. [DOI] [PubMed] [Google Scholar]
- Farrimond JA, Hill AJ, Jones NA, Stephens GJ, Whalley BJ, Williams CM. A cost-effective high-throughput digital system for observation and acquisition of animal behavioral data. Behav Res Methods. 2009;41:446–451. doi: 10.3758/BRM.41.2.446. [DOI] [PubMed] [Google Scholar]
- FDA. 2004. Guidance for industry: botanical drug products [Online]. Available at: http://www.fda.gov/downloads/Drugs/GuidanceComplianceRegulatoryInformation/Guidances/ucm070491.pdf (accessed 13 August 2013)
- Formukong EA, Evans AT, Evans FJ. Inhibition of the cataleptic effect of tetrahydrocannabinol by other constituents of Cannabis sativa L. J Pharm Pharmacol. 1988;40:132–134. doi: 10.1111/j.2042-7158.1988.tb05198.x. [DOI] [PubMed] [Google Scholar]
- Fried P, McIntyre D. Electrical and behavioral attenuation of the anti-convulsant properties of 9-THC following chronic administrations. Psychopharmacology (Berl) 1973;31:215–227. doi: 10.1007/BF00422512. [DOI] [PubMed] [Google Scholar]
- Hermann BP, Seidenberg M, Bell B, Woodard A, Rutecki P, Sheth R. Comorbid psychiatric symptoms in temporal lobe epilepsy: association with chronicity of epilepsy and impact on quality of life. Epilepsy Behav. 2000;1:184–190. doi: 10.1006/ebeh.2000.0066. [DOI] [PubMed] [Google Scholar]
- Hill A, Mercier M, Hill T, Glyn S, Jones N, Yamasaki Y, et al. Cannabidivarin is anticonvulsant in mouse and rat in vitro and in seizure models. Br J Pharmacol. 2012a;167:1629–1642. doi: 10.1111/j.1476-5381.2012.02207.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill AJ, Weston SE, Jones NA, Smith I, Bevan SA, Williamson EM, et al. Δ9-Tetrahydrocannabivarin suppresses in vitro epileptiform and in vivo seizure activity in adult rats. Epilepsia. 2010;51:1522–1532. doi: 10.1111/j.1528-1167.2010.02523.x. [DOI] [PubMed] [Google Scholar]
- Hill AJ, Williams CM, Whalley BJ, Stephens GJ. Phytocannabinoids as novel therapeutic agents in CNS disorders. Pharmacol Ther. 2012b;133:79–97. doi: 10.1016/j.pharmthera.2011.09.002. [DOI] [PubMed] [Google Scholar]
- Hitiris N, Mohanraj R, Norrie J, Sills GJ, Brodie MJ. Predictors of pharmacoresistant epilepsy. Epilepsy Res. 2007;75:192–196. doi: 10.1016/j.eplepsyres.2007.06.003. [DOI] [PubMed] [Google Scholar]
- Hosford DA, Wang Y. Utility of the lethargic (lh/lh) mouse model of absence seizures in predicting the effects of lamotrigine, vigabatrin, tiagabine, gabapentin, and topiramate against human absence seizures. Epilepsia. 1997;38:408–414. doi: 10.1111/j.1528-1157.1997.tb01729.x. [DOI] [PubMed] [Google Scholar]
- Järbe TU, Andrzejewski ME, DiPatrizio NV. Interactions between the CB1 receptor agonist Δ9-THC and the CB1 receptor antagonist SR-141716 in rats: open-field revisited. Pharmacol Biochem Behav. 2002;73:911–919. doi: 10.1016/s0091-3057(02)00938-3. [DOI] [PubMed] [Google Scholar]
- Jensen LH, Petersen EN, Braestrup C. Audiogenic seizures in DBA/2 mice discriminate sensitively between low efficacy benzodazepine receptor agonists and inverse agonists. Life Sci. 1983;33:393–399. doi: 10.1016/s0024-3205(83)80014-9. [DOI] [PubMed] [Google Scholar]
- Jones NA, Hill AJ, Smith I, Bevan SA, Williams CM, Whalley BJ, et al. Cannabidiol displays antiepileptiform and antiseizure properties in vitro and in vivo. J Pharmacol Exp Ther. 2010;332:569–577. doi: 10.1124/jpet.109.159145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones NA, Glyn SE, Akiyama S, Hill TDM, Hill AJ, Weston SE, et al. Cannabidiol exerts anti-convulsant effects in animal models of temporal lobe and partial seizures. Seizure. 2012;21:344–352. doi: 10.1016/j.seizure.2012.03.001. [DOI] [PubMed] [Google Scholar]
- Kanner AM, Schachter SC, Barry JJ, Hersdorffer DC, Mula M, Trimble M, et al. Depression and epilepsy: epidemiologic and neurobiologic perspectives that may explain their high comorbid occurrence. Epilepsy Behav. 2012;24:156–168. doi: 10.1016/j.yebeh.2012.01.007. [DOI] [PubMed] [Google Scholar]
- Karler R, Turkanis SA. Subacute cannabinoid treatment: anticonvulsant activity and withdrawal excitability in mice. Br J Pharmacol. 1980;68:479–484. doi: 10.1111/j.1476-5381.1980.tb14562.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilkenny C, Browne W, Cuthill IC, Emerson M, Altman DG. Animal research: reporting in vivo experiments: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1577–1579. doi: 10.1111/j.1476-5381.2010.00872.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonardi M, Ustun TB. The global burden of epilepsy. Epilepsia. 2002;43:21–25. doi: 10.1046/j.1528-1157.43.s.6.11.x. [DOI] [PubMed] [Google Scholar]
- Löscher W. Critical review of current animal models of seizures and epilepsy used in the discovery and development of new antiepileptic drugs. Seizure. 2011;20:359–368. doi: 10.1016/j.seizure.2011.01.003. [DOI] [PubMed] [Google Scholar]
- Marescaux C, Vergnes M. Genetic absence epilepsy in rats from Strasbourg (GAERS) Ital J Neurol Sci. 1995;16:113–118. doi: 10.1007/BF02229083. [DOI] [PubMed] [Google Scholar]
- McGrath J, Drummond G, McLachlan E, Kilkenny C, Wainwright C. Guidelines for reporting experiments involving animals: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1573–1576. doi: 10.1111/j.1476-5381.2010.00873.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McPartland JM, Russo EB. Cannabis and cannabis extracts. J Cannabis Ther. 2001;1:103–132. [Google Scholar]
- Nevins ME, Nash SA, Beardsley PM. Quantitative grip strength assessment as a means of evaluating muscle relaxation in mice. Psychopharmacology (Berl) 1993;110:92–96. doi: 10.1007/BF02246955. [DOI] [PubMed] [Google Scholar]
- Ortinski P, Meador KJ. Cognitive side effects of antiepileptic drugs. Epilepsy Behav. 2004;5:60–65. doi: 10.1016/j.yebeh.2003.11.008. [DOI] [PubMed] [Google Scholar]
- Pertwee R. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. Br J Pharmacol. 2008;153:199–215. doi: 10.1038/sj.bjp.0707442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG. Inverse agonism and neutral antagonism at cannabinoid CB1 receptors. Life Sci. 2005;76:1307–1324. doi: 10.1016/j.lfs.2004.10.025. [DOI] [PubMed] [Google Scholar]
- Perucca E, Beghi E, Dulac O, Shorvon S, Tomson T. Assessing risk to benefit ratio in antiepileptic drug therapy. Epilepsy Res. 2000;41:107–139. doi: 10.1016/s0920-1211(00)00124-8. [DOI] [PubMed] [Google Scholar]
- Ross HR, Napier I, Connor M. Inhibition of recombinant human T-type calcium channels by Delta9-tetrahydrocannabinol and cannabidiol. J Biol Chem. 2008;283:16124–16134. doi: 10.1074/jbc.M707104200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross RA, Brockie HC, Stevenson LA, Murphy VL, Templeton F, Makriyannis A, et al. Agonist-inverse agonist characterization at CB1 and CB2 cannabinoid receptors of L759633, L759656 and AM630. Br J Pharmacol. 1999;126:665–672. doi: 10.1038/sj.bjp.0702351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo E, Guy GW. A tale of two cannabinoids: the therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med Hypotheses. 2006;66:234–246. doi: 10.1016/j.mehy.2005.08.026. [DOI] [PubMed] [Google Scholar]
- Russo EB. Taming THC: potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br J Pharmacol. 2011;163:1344–1364. doi: 10.1111/j.1476-5381.2011.01238.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo EB, Burnett A, Hall B, Parker KK. Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochem Res. 2005;30:1037–1043. doi: 10.1007/s11064-005-6978-1. [DOI] [PubMed] [Google Scholar]
- Ryan D, Drysdale AJ, Pertwee RG, Platt B. Differential effects of cannabis extracts and pure plant cannabinoids on hippocampal neurones and glia. Neurosci Lett. 2006;408:236–241. doi: 10.1016/j.neulet.2006.09.008. [DOI] [PubMed] [Google Scholar]
- Sagredo O, Pazos MR, Satta V, Ramos JA, Pertwee RG, Fernández-Ruiz J. Neuroprotective effects of phytocannabinoid-based medicines in experimental models of Huntington's disease. J Neurosci Res. 2011;89:1509–1518. doi: 10.1002/jnr.22682. [DOI] [PubMed] [Google Scholar]
- Schachter SC. Currently available antiepileptic drugs. Neurother. 2007;4:4–11. doi: 10.1016/j.nurt.2006.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scutt A, Williamson EM. Cannabinoids stimulate fibroblastic colony formation by bone marrow cells indirectly via CB2 receptors. Calcif Tissue Int. 2007;80:50–59. doi: 10.1007/s00223-006-0171-7. [DOI] [PubMed] [Google Scholar]
- Sed J, Urdzíková L, Jendelová P, Syková E. Methods for behavioral testing of spinal cord injured rats. Neurosci Biobehav Rev. 2008;32:550–580. doi: 10.1016/j.neubiorev.2007.10.001. [DOI] [PubMed] [Google Scholar]
- Tallarida RJ. An overview of drug combination analysis with isobolograms. J Pharmacol Exp Ther. 2006;319:1–7. doi: 10.1124/jpet.106.104117. [DOI] [PubMed] [Google Scholar]
- Thomas A, Ross R, Saha B, Mahadevan A, Razdan R, Pertwee RG. 6″-azidohex-2″-yne-cannabidiol: a potential neutral, competitive cannabinoid CB1 receptor antagonist. Eur J Pharmacol. 2004;487:213–221. doi: 10.1016/j.ejphar.2004.01.023. [DOI] [PubMed] [Google Scholar]
- Tóth A, Boczán J, Kedei N, Lizanecz E, Bagi Z, Papp Z, et al. Expression and distribution of vanilloid receptor 1 (TRPV1) in the adult rat brain. Brain Res Mol Brain Res. 2005;135:162–168. doi: 10.1016/j.molbrainres.2004.12.003. [DOI] [PubMed] [Google Scholar]
- Valdeolivas S, Satta V, Pertwee RG, Ferna′ndez-Ruiz J, Sagredo O. Sativex-like combination of phytocannabinoids is neuroprotective in malonate-lesioned rats, an inflammatory model of Huntington's disease: role of CB1 and CB2 receptors. ACS Chem Neurosci. 2012;3:400–406. doi: 10.1021/cn200114w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallace MJ, Wiley JL, Martin BR, DeLorenzo RJ. Assessment of the role of CB1 receptors in cannabinoid anticonvulsant effects. Eur J Pharmacol. 2001;428:51–57. doi: 10.1016/s0014-2999(01)01243-2. [DOI] [PubMed] [Google Scholar]
- Wallace MJ, Blair RE, Falenski KW, Martin BR, DeLorenzo RJ. The endogenous cannabinoid system regulates seizure frequency and duration in a model of temporal lobe epilepsy. J Pharmacol Exp Ther. 2003;307:129–137. doi: 10.1124/jpet.103.051920. [DOI] [PubMed] [Google Scholar]
- Whalley BJ, Wilkinson JD, Williamson EM, Constanti A. A novel component of cannabis extract potentiates excitatory synaptic transmission in rat olfactory cortex in vitro. Neurosci Lett. 2004;365:58–63. doi: 10.1016/j.neulet.2004.04.044. [DOI] [PubMed] [Google Scholar]
- Wilkinson JD, Whalley BJ, Baker D, Pryce G, Constanti A, Gibbons S, et al. Medicinal cannabis: is 9–tetrahydrocannabinol necessary for all its effects? J Pharm Pharmacol. 2003;55:1687–1694. doi: 10.1211/0022357022304. [DOI] [PubMed] [Google Scholar]