

Abstract
Study Objectives:
Sleep deprivation and daytime somnolence impair numerous aspects of physical, cognitive, and memory performance. However, most studies examining the effect of somnolence on brain function focus on acute sleep restriction in young adults. We examine the relationship between chronic daytime somnolence and connectivity in six brain networks in both young and elderly subjects using stimulus-free resting-state functional magnetic resonance imaging.
Design:
Cross-sectional.
Setting:
Outpatient research at the Massachusetts General Hospital.
Participants:
Young (n = 27) and elderly (n = 84) healthy, cognitively normal volunteers.
Interventions:
None.
Measurements and Results:
Compared with young subjects, cognitively normal elderly adults report less daytime somnolence on the Epworth Sleepiness Scale (ESS) (P = 0.019) and display reduced default mode network (DMN) connectivity (P = 0.004). Across all subjects, increasing daytime sleepiness was associated with decreasing functional connectivity in the DMN (P = 0.003, partial r of ESS = -0.29). There was no difference in the slope of this relationship between young adults and elderly subjects. No other cortical networks were correlated with daytime sleepiness. Daytime sleepiness and DMN connectivity were not related to sex, brain structure, or body mass index.
Conclusions:
These findings suggest that daytime sleepiness is associated with impaired connectivity of the DMN in a manner that is distinct from the effects of aging. This association is important to consider in any study using DMN connectivity as a biomarker. Additionally, these results may help identify those subjects at risk for future memory decline.
Citation:
Ward AM; McLaren DG; Schultz AP; Chhatwal J; Boot BP; Hedden T; Sperling RA. Daytime sleepiness is associated with decreased default mode network connectivity in both young and cognitively intact elderly subjects. SLEEP 2013;36(11):1609-1615.
Keywords: Epworth Sleepiness Scale, functional connectivity, neuroimaging, magnetic resonance imaging, sleep
INTRODUCTION
Chronic sleep deprivation is a burgeoning public health concern, with as many as one-third of Americans showing signs of sleep deprivation—a number that is rising.1,2 Chronic sleep deprivation and sleep restriction lead to significant neurological and psychiatric comorbidities that are known to be detrimental to cardiovascular health,3 safety,4,5 and quality of life.6,7 Sleep deprivation has also been linked to declines in memory performance.8–12 Although the relationship between nighttime sleep deprivation and daytime sleepiness is complex,13 excessive daytime sleepiness increases risk of cognitive impairment and decline in elderly subjects at risk for dementia.14–17 Additionally, young adults and cognitively normal elderly adults exhibit differential sensitivity to sleep deprivation. Younger adults show more cognitive impairment with sleep deprivation than do elderly adults,18 although sleep deprivation in older adults also leads to poorer memory performance.19,20 However, it remains unclear the specific effects of chronic sleepiness on the waking brain.
Resting-state functional connectivity magnetic resonance imaging (fcMRI) identifies large-scale brain networks by identifying correlated intrinsic activity.21–23 These networks overlap with known anatomical connections.24,25 One of these networks, the default mode network (DMN), is strongly implicated in internal modes of cognition.26 Although further work is necessary to separate the relative effects on episodic and working memory, the strength of correlated activity within the DMN appears closely tied to memory performance.27–29 Decreased DMN connectivity is observed in amnestic mild cognitive impairment30,31 and Alzheimer disease dementia.30,32 Group-level fcMRI can be used to predict Alzheimer disease progression33 and to differentiate subjects with mild cognitive impairment who are particularly likely to progress to dementia,34 underscoring the potential clinical utility of DMN fcMRI.
DMN connectivity is significantly decreased in subjects who are deeply asleep35 or lightly sedated,36,37 as well as in subjects who are descending into sleep.38,39 Additionally, sleep-deprived subjects also show significantly decreased DMN connectivity.40,41 These results suggest that decreased DMN connectivity during the daytime may reflect a more “sleep-like” state in the brain. This may also indicate a neural mechanism by which disordered sleep can lead to decreased memory performance.
Building on the relationship between sleep deprivation and impaired memory and the rich literature linking decreased DMN fcMRI to mnemonic dysfunction, here we examine whether self-reported daytime somnolence affects DMN fcMRI. To address the effect of daytime somnolence (rather than acute sleep deprivation) on functional connectivity and memory, we focused on self-report of daytime somnolence using the Epworth Sleepiness Scale (ESS). We examined both young and cognitively normal older adults to determine if DMN fcMRI is differentially related to daytime sleepiness in these different age groups.
METHODS
Participants
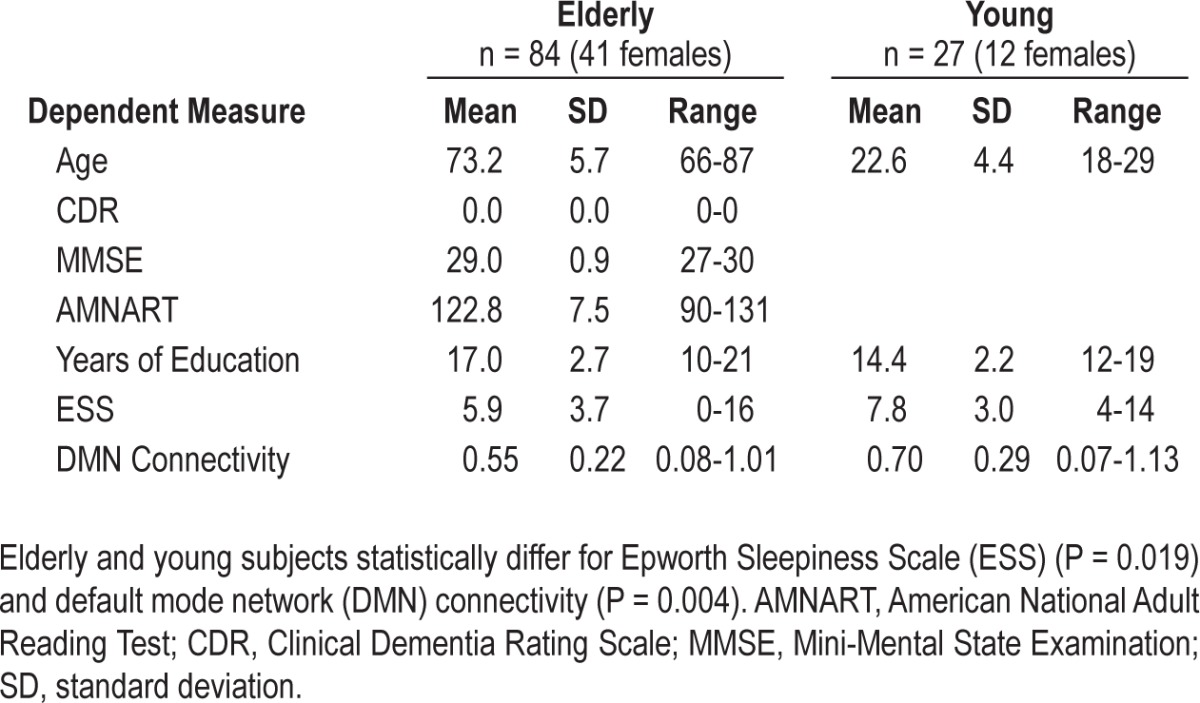
Eighty-five clinically normal elderly subjects (age 66-87 y) and 27 young adults (age 18-29 y) participated in the study (Table 1). All subjects were native English speakers and had normal or corrected-to-normal vision; had no history of sleep, psychiatric, or neurological disorders; and had no history of head trauma. Elderly inclusion criteria included Mini-Mental State Examination (MMSE) scores of ≥ 27, a Clinical Dementia Rating of 0, and scores within one standard deviation of age-adjusted norms for Logical Memory IIa. Subjects with a confirmed diagnosis of a sleep disorder and subjects using continuous positive airway pressure were excluded. The elderly adults are a subset of the longitudinal Harvard Aging Brain study participants. Informed consent was obtained in accordance with the guidelines and procedures governed by the institutional review board of the Massachusetts General Hospital (Boston, MA).
Table 1.
Demographic information
Neuropsychological Measures
All subjects completed the ESS42 within 50 days of the MRI. The ESS assesses the likelihood of dozing (rated from 0-3) in eight different scenarios. The ESS has been shown to remain largely stable over time across several populations.43,44 However, subjects with obstructive sleep apnea who started CPAP treatment showed a marked decrease in ESS.43 The ESS has been found to be internally consistent in elderly men44 and elderly women.45 Using a cutoff score of 10, the ESS is 93.5% sensitive and 100% specific for diagnosis of excessive daytime sleepiness.46 Elderly participants also completed the Digit Symbol Substitution Test, the Rey Auditory Verbal Learning Test, digits span backward, the Free and Cued Selective Reminding Test, the Trail Making Test parts A and B, and the Letter Number Sequencing Test.
MRI Data Acquisition and Processing
Subjects underwent an MRI on a Siemens Trio Tim 3.0 Tesla scanner (Siemens Medical Systems, Erlangen, Germany) equipped with a 12-channel phased-array head coil. Head motion was restrained with extendable foam-padded clamps and a memory-foam head coil insert. Resting-state data were acquired in two separate, sequentially acquired scans using gradient-echo echo-planar imaging sensitive to blood oxygenation level-dependent contrast.47,48 Subjects were instructed to keep their eyes open, lie still, and remain awake. A fixation cross was projected onto a screen positioned at the head of the magnet bore and viewed with a mirror attached to the head coil. Forty-seven interleaved axial slices aligned parallel to the anterior-/posterior-commissure plane provided whole brain coverage with the following parameters: repetition time (TR) = 3,000 ms, echo time (TE) = 30 ms, flip angle = 85°, field-of-view (FOV) = 216 × 216 mm, matrix = 72 × 72, and 3 × 3 × 3 mm voxels. One hundred twenty-four volumes were acquired in each scan. T1-weighted structural images were acquired with a multiecho magnetization-prepared rapid gradient echo (MEMP-RAGE) sagittal-oriented sequence49 with the following parameters: TR = 2,200 ms; inversion time (TI) = 1100 ms; TE = 1.54 ms, 3.36 ms, 5.18 ms, and 7ms; flip angle = 7°; FOV = 230 × 230 mm; matrix size = 192 × 192; NEX = 1; and 1.2 × 1.2 × 1.2 mm voxels. A root-mean-square image was produced from all four echoes and was used in the structural analyses.
Structural Preprocessing
MEMP-RAGE data were processed with SPM8 (http://www.fil.ion.ucl.ac.uk/spm/; version r4290) using the following supervised voxel-based morphometry (VBM) protocol: (1) T1-weighted images were manually translated and/or rotated to match the rotation and origin of the Montreal Neurological Institute (MNI) Template; (2) the translated/rotated T1-weighted images were segmented using the “New Segment” tool to produce rigidly aligned gray matter and white matter segments; (3) the rigidly aligned segments were normalized using a fast diffeomorphic image registration algorithm (DARTEL),50 which included estimating the flow fields, warping the images to MNI space, and modulating the images; and (5) then the images were smoothed with a 6-mm full width at half maximum (FWHM) gaussian kernel.
Structural Analysis
To examine the effect of sleepiness on brain structure, we conducted a series of VBM analyses: (1) whole-brain regressions using the VBM-derived amounts of gray matter as the dependent variables, (2) small-volume correction in the ventromedial prefrontal cortex (vmPFC), and (3) small-volume correction in the DMN regions of interest (ROIs). These analyses were conducted using multiple regression models in SPM8 where the volume for each subject was entered as the dependent variable, with ESS as the independent variable, while controlling for age, sex, and total gray matter. Because age contributes significantly to changes in morphometry, young and elderly subjects were analyzed in separate models. Second, previous work has shown a specific effect of ESS on structure in the vmPFC using small-volume correction.51 To assess this in our data, we performed a small-volume correction using 10-mm spheres centered on MNI coordinates [-12 26 -27] and [12 18 -23] as the small volume. Small-volume correction reduces the number of comparisons performed by performing an analysis on a restricted volume, thereby lowering the criterion for significance. Third, we performed a small-volume correction using our DMN ROIs as the small volume. The whole-brain and small-volume correction significance testing were corrected for multiple comparisons using false discovery rate correction using q < 0.05.52
Finally, we used the amount of gray matter within the functional connectivity ROIs as a covariate in our main analyses. This covariate was calculated by extracting the relative amount of gray matter in each ROI for each subject. These values were averaged to create an overall metric of ROI gray matter volume.
fcMRI Preprocessing
Resting-state functional data were preprocessed using SPM8 and MATLAB (v7.11; MathWorks, Natick, MA). The first four volumes of each run were discarded to allow for T1-equilibration effects. Each run was slice-time corrected, realigned to the first volume of the run with INRIAlign (http://www-sop.inria.fr/epidaure/software/INRIAlign/)53,54 using a rigid-body rotation, normalized to the MNI echo-planar imaging template, and smoothed with a 6-mm FWHM gaussian kernel. After preprocessing, a series of regressors from the resting-state data were entered into a multiple regression analysis. These were: (1) motion parameters as estimated by the six realignment parameters, (2) average signal from a deep white matter mask, (3) average signal from a ventricle mask, (4) average signal from the whole brain mask, and (5) the first derivative of each of these nuisance regressors. The residuals from this model were then linearly detrended and low-pass filtered with a second-order Butterworth filter with a frequency cutoff of 0.08 Hz.22,55 These runs were then concatenated for all analyses.
fcMRI First-level Analysis
Three a priori ROIs representing major nodal regions within the DMN were chosen for examination using a method described previously.56 Each ROI represents an 8-mm sphere centered on a peak voxel in an established node of the default-mode network: the posterior cingulate cortex (PCC) MNI [0, -53, 26], medial prefrontal cortex (mPFC) MNI [0, 52, -6], and inferior parietal lobule (IPL) MNI [-48, -62, 36; 46, -62, 32]. The IPL spheres were combined into a single ROI. Time series data were extracted from every voxel within each ROI, and an arithmetic mean across time was calculated. Pairwise correlations between individual nodes of the DMN were calculated. Next, these pairwise nodal functional connectivity values were normalized using Fisher r-to-z transformation.57 We also computed a summary measure of DMN connectivity by averaging the z-values. Identical methods were used to determine the network health for five other prominent brain networks using previously published spherical ROIs: the dorsal attention, ventral attention, frontal-parietal control, visual, and motor networks58 (see Table S1 for ROI locations). All connectivity data were analyzed using R (version 2.15.0, http://www.cran.r-project.org).
fcMRI Second-level Analysis: Main Model
Our main model analyzes overall DMN connectivity as the dependent variable. The amount of gray matter within the ROIs is entered as a covariate to control for the potentially confounding effects of age and cortical atrophy on connectivity. Because we include young and elderly subjects in the same model, age group is entered as a covariate. Age and the variable of interest, ESS, are included as parametric covariates, and allowed to interact with age group to control for the potential differing relationships between age and ESS on connectivity within young and elderly subjects.
fcMRI Second-Level Analysis: Selection of Covariates
First, we confirmed the normality of the ESS scores using the Shapiro-Wilk test (W = 0.984, P = 0.215). Next, we examined ESS scores and functional network connectivity for sex effects. None were found, so sex was not included as a covariate in any subsequent models. We then examined the data for any relationship with body mass index (BMI) to assess the potential contribution of undiagnosed sleep apnea. BMI was not significantly correlated with ESS or DMN connectivity. Because BMI was unrelated to ESS, we excluded it from the model. We investigated the effects of lag time between ESS and MRI acquisition because subjects did not have their MRI and ESS data acquisitions on the same day. Lag was not significantly correlated with ESS or DMN connectivity, nor did it significantly alter our subsequent analyses. Thus, we excluded it from the model. Because ESS scores have been shown to be stable over a period of 5 mo, this result was not surprising.43 Next, we examined our DMN connectivity and sleepiness data for age group effects. Both of these models allowed age to interact with age group to control for any within-group effects of age. The DMN connectivity model also controlled for gray matter volume to eliminate the possibility of morphometry significantly contributing to the results. Age group was the only significant predictor in both models. Although both DMN connectivity and sleepiness were significantly different between young and elderly subjects (Table 1), neither the (age group)*age nor the (age group)*ESS interactions were significant in our main model, indicating a similar relationship between ESS and DMN connectivity in both young and elderly. However, we include these variables in our model because they do not significantly alter the relationship between ESS and DMN connectivity.
Neuropsychological Models
Finally, to examine the effects of daytime somnolence and DMN connectivity on cognitive function in our elderly sample, we examined the relationship between these measures and neuropsychological measures of cognitive performance. These analyses were conducted using a standard general linear model controlling for age.
RESULTS
Default-Mode Network Connectivity Is Different between Young and Elderly Subjects
As expected, and consistent with previous work,29,59–61 the young group had significantly greater mean DMN connectivity than the elderly group while controlling for age and gray matter volume (P = 0.004; Table 1).
Epworth Sleepiness Scale Scores Are Different between Young and Elderly Subjects
Our young subject group had significantly higher ESS scores than our elderly group while controlling for age (P = 0.019; Table 1). This finding is consistent with previous work that indicates that elderly adults have a reduced propensity for daytime sleep.62–64
Increased ESS Scores Are Associated with Decreased DMN Connectivity
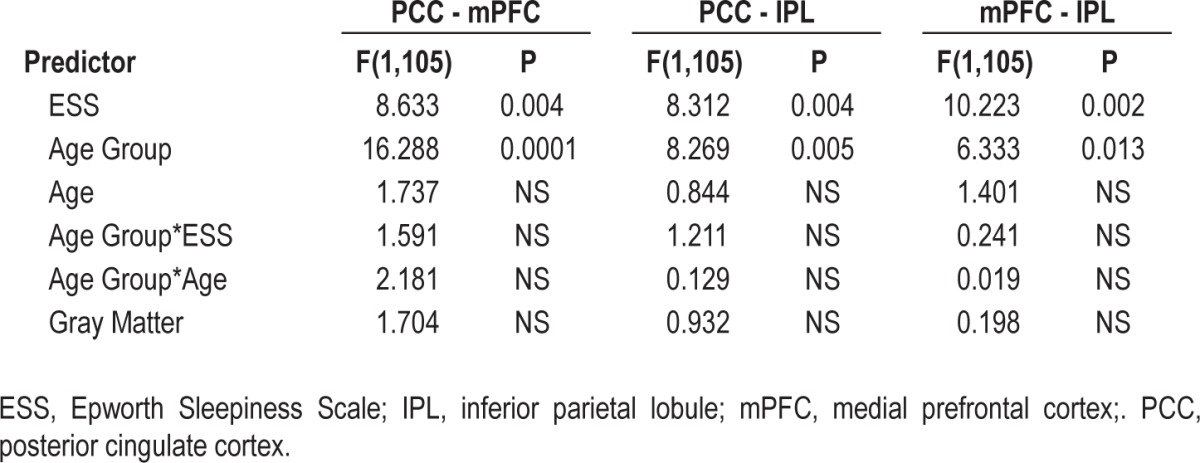
Grouping together older and younger subjects, but including age group as a factor, we observed that increasing ESS was significantly related to decreasing DMN functional connectivity (F(1,105) = 8.983, P = 0.003 partial r of ESS = -0.29; Figure 1, Table 2). Only ESS and age group were significantly related to DMN connectivity. Importantly, the age group by ESS interaction was not significant, indicating a similar relationship between ESS and DMN connectivity in both young and elderly subjects. These relationships were preserved when examining the pairwise ROI correlations individually (Table 3) as well as in a multivariate analysis of variance using all pairwise ROI correlations as dependent variables.
Figure 1.
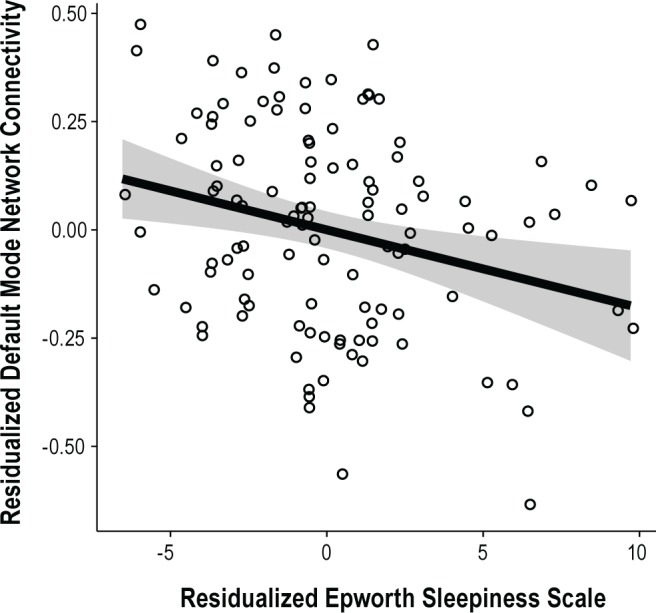
Residualized default mode network connectivity plotted against residualized Epworth Sleepiness Scale score (P = 0.003, partial r = -0.29).
Table 2.
Results of full analysis of covariance model with default mode network connectivity as dependent, and Epworth Sleepiness Scale as variable of interest
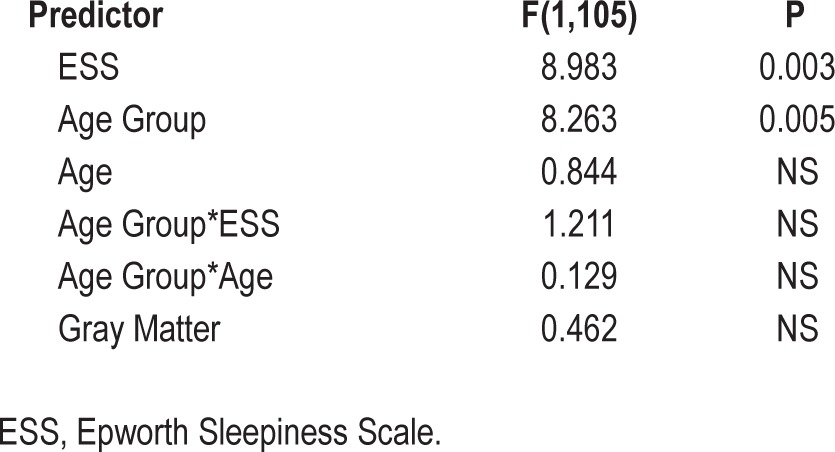
Table 3.
Relationship between default-mode network pairwise correlations as dependent and Epworth Sleepiness Scale as variable of interest
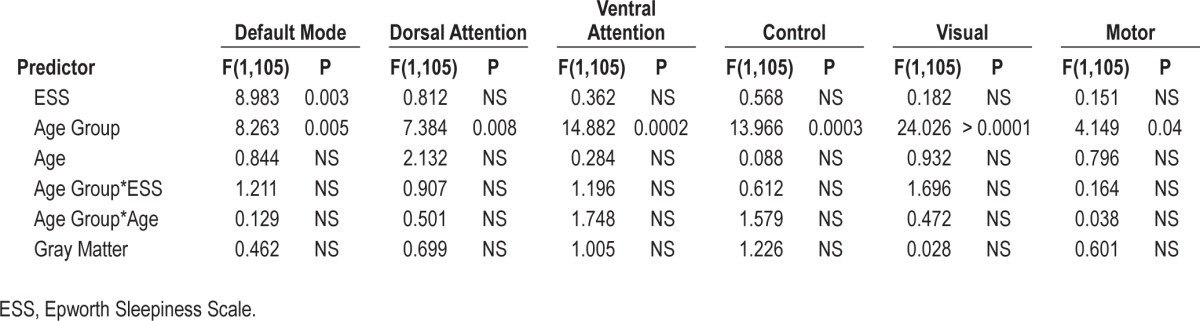
To determine whether decreased fcMRI is specific to the DMN or representative of widespread changes in functional connectivity in subjects with high ESS scores, we performed functional connectivity analyses in five other well-established brain networks: the dorsal attention, ventral attention, frontal-parietal control, visual, and motor networks.58 ESS scores were not significantly correlated with functional connectivity in any of these other networks (Table 4), suggesting that daytime somnolence-related decreases in network connectivity are specific to the DMN. Additionally, the lack of association between daytime somnolence and functional connectivity in the primary sensory networks suggests that the changes in connectivity within the DMN are not attributable to differences in the global signal properties of the brain (i.e., does not represent a systematic alteration in measurement bias), but instead reflect specific changes within the DMN.
Table 4.
Relationship between Epworth Sleepiness Scale and network connectivity for six major cortical networks.
Neither ESS Scores nor DMN Connectivity Are Associated with Differences in Brain Structure
Previous work has indicated that increasing daytime somnolence may have an effect on brain structure in young subjects.51 To test this possibility in our sample, we analyzed the relationship between ESS scores and VBM-derived gray matter at the whole-brain level. This model assessed every voxel in the brain as a dependent variable, covarying for age and ESS score. Because age contributes significantly to changes in morphometry, young and elderly subjects were analyzed in separate models. However, no voxels survived false discovery rate correction (q < 0.05) at the whole-brain level for either positive or negative correlations with ESS in either group. We used small-volume correction to perform a directed volumetric analysis of the ventromedial prefrontal cortex (which had previously been noted to covary with ESS)51 but we found no significant relationships in this region. Finally, because our functional connectivity analysis focuses on DMN regions, we also performed a small-volume correction focusing on these regions. Again, no voxels survived false discovery rate correction.
To determine if DMN connectivity has an independent relationship from ESS on brain structure, we repeated the aforementioned structural analysis, substituting our DMN connectivity metric for the ESS measure. Once again, no voxels survived false discovery rate correction, indicating no relationship between network connectivity and brain structure in our sample. These results combined suggest that changes in daytime somnolence and functional connectivity are not linked to alterations in brain structure.
Increased ESS Scores Are Not Associated with Cognitive Deficits
Although the neuropsychological testing within the elderly cohort spanned several cognitive domains, we found no relationship between any of the cognitive tests (Digit Symbol Substitution, Rey Auditory Verbal Learning Test, digits span backwards, Free and Cued Selective Reminding, Trail Making Part B minus Part A, and letter number span) and ESS. Additionally, within this sample, there was no relationship between the DMN connectivity metric and any of the neuropsycho-logical test scores. None of the pairwise ROI measures were related to the neuropsychological test scores.
DISCUSSION
Daytime somnolence, as measured by the ESS, is associated with a reduction in DMN functional connectivity. This effect is found in both young and elderly subjects and the slope of the relationship is the same between these age groups. The relationship is not accounted for by changes in brain structure (as measured by VBM) and is specific to the DMN. An association between somnolence and connectivity is not seen in five other well-established functional networks.
Previous work has indicated reduced DMN connectivity in subjects who are descending into sleep,38 are deeply asleep,35 are lightly sedated,36,37 have acute sleep deprivation,40,41 or who have slept poorly for a single night.65 Our results extend these findings: chronic daytime somnolence is similarly associated with reduced DMN connectivity. These results suggest that sleepiness, or a sleep-like state, correlate with decreased DMN connectivity, and that this disconnection occurs in both acute and chronic somnolence. This decrease in DMN connectivity may reflect a “local sleep” phenomenon whereby parts of the brain enter a sleep state during consciousness.66
Consistent with prior studies,29,59–61,67 we found that older subjects have lower DMN connectivity than younger subjects, regardless of ESS score. This replicates the finding that some decreases in DMN connectivity are related to aging in the absence of pathological cognitive decline.60,68 However, despite the fact that older individuals showed lower DMN fcMRI at baseline, the relationship between somnolence and DMN fcMRI was similar in both young and elderly cohorts. Thus, it is likely that the relationship between ESS and DMN fcMRI is distinct from the effect of normal aging. The observation that increasing ESS scores correlate with decreasing DMN fcMRI in young subjects suggests that this mechanism is unlikely to be neurodegenerative, a conclusion bolstered by the lack of a relationship between ESS and brain structure in older individuals. Additionally, we found that our elderly sample reported less daytime somnolence than our young sample, consistent with previous work,62–64 again suggesting that the relationship of age to DMN connectivity is distinct from the relationship between somnolence and DMN connectivity. However, data relating somnolence to DMN connectivity in nonselected aged populations are needed to confirm this hypothesis.
Sleep loss and fragmented sleep lead to decreases in many aspects of neurocognitive performance,8,11,20,69,70 and chronic mild sleep deprivation is associated with adverse effects on reaction times, memory, cognition, and general health.8,10,11,71–73 However, our elderly sample was selected on the basis of their intact cognitive test scores. This truncated the range on the cognitive measures, thereby decreasing the likelihood of significant cross-sectional associations with sleepiness. Furthermore, the ESS is a short self-reported questionnaire, and may therefore be relatively insensitive to incremental differences between subjects with similar levels of sleepiness. Finally, recent work indicates that daytime sleepiness is related to longitudinal change in neuropsychological scores as opposed to being directly related to baseline measures.14–17
Coupled with literature tying DMN fcMRI with mnemonic function,74–76 the current results raise the intriguing possibility that changes in DMN connectivity attributable to somnolence may be a mechanism by which excessive daytime somnolence (whether acute or chronic) leads to declines in memory function. Future studies aimed at assessing the relationship of nighttime sleep quality (as measured by actigraphy or polysomnography) to daytime sleep and DMN fcMRI will help to clarify the interdependent relationship between sleep structure and quality, propensity for daytime sleep, and DMN connectivity.
Additionally, this study was conducted without the benefit of simultaneous electroencephalographic recording. Consequently, we cannot guarantee that our subjects did fall asleep during the resting state scans. However, it is unlikely that this potential confound is present in our paradigm because (1) the ESS is not correlated with sleep latency in an elderly population,13 (2) our fMRI scanning is performed during the mornings, early in the MRI session, and with eyes open, and (3) is divided into two shorter-duration (6-12 min) scans with interruptions that require subject response between scans. Finally, we performed detailed analyses of head movement to determine if subjects with high sleepiness scores moved more during the scans (suggesting hypnic myoclonia) or moved less (suggesting sleep atonia) but found no association with ESS scores (see supplemental materials for details). It is important to note that this confound potentially affects any study examining resting-state connectivity.
Finally, given the development of DMN fcMRI as a noninvasive biomarker in mild cognitive impairment and Alzheimer disease,77,78 the current results suggest that ESS or other measures of somnolence or sleep quality may be useful additions to functional connectivity imaging studies in neurodegenerative disease. First, it appears that the effects of daytime sleepiness on DMN connectivity are dissociable from the effects of aging on DMN connectivity. Second, recent reports suggest that disruptions of sleep throughout the life span correlate with poorer cognition later in life,79 excessive daytime sleepiness increases an elderly individual's risk of cognitive decline,14–17 and sleep quality may be associated with beta-amyloid accumulation.80 Further, it will be important to assess the possibility that sleepiness may mediate the changes in DMN connectivity that are observable during the conversion from cognitive normality to mild cognitive impairment and Alzheimer disease. Finally, consideration of the effects of excessive daytime somnolence on DMN fcMRI may help to explain the variability seen in neurodegenerative studies of DMN fcMRI, and to potentially identify subjects at risk for cognitive decline.
DISCLOSURE STATEMENT
This was not an industry supported study. The authors have indicated no financial conflicts of interest. This work was supported by NIA AG027435, P01AG036694, P50 AG005134, K24 AG035007 to Dr. Sperling.
ACKNOWLEDGMENTS
The authors gratefully acknowledge: the Martinos Center for Biomedical Imaging staff, including Mary Foley and Larry White; Sarah Wigman and Caroline Sullivan for assistance with scanning; Dorene Rentz, Gad Marshall, Rebecca Amariglio, and Scott McGinnis for assistance with subject evaluations; and the participation of the research subjects.
SUPPLEMENTAL MATERIAL
SUPPLEMENTAL METHODS
Although simultaneous electroencephalographic data were not available on individual subjects, we conducted an analysis on movement during our resting state acquisition to assess the relative likelihood that the patients with excessive daytime sleepiness (Epworth Sleepiness Scale ≥ 10) were sleeping, as opposed to sleepy, in the scanner.
To investigate decreased muscle tone due to stage 1 nonrapid eye movement sleep (NREM), we calculated the euclidian distance that the subject moved between each frame. Importantly, these distances are calculated frame-to-frame (i.e., euclidian distance from the frame before) as opposed to average deviation from the mean image. This allows us to avoid overemphasizing the effects of drift, or of a single large movement.
To investigate hypnic myoclonia due to subjects entering and exiting stage 1 NREM sleep, we calculated the number of frames where movement from the frame before exceeded 0.75 mm or 1.5 degrees, or both.1 Because the relationship between movement and sleepiness was unlikely to be linear, we divided our subjects into three groups: not sleepy (ESS ≤ 3, n = 23), middle (4 ≤ ESS ≥ 9, n = 68), and excessive daytime sleepiness (ESS ≥ 10, n = 21). This design allows us to focus on mean differences, which allows us to examine differences without presupposing the relationships between these means. However, we also examined linear models of both mean movement and count of large-amplitude movements, as well as a linear model containing both measures of movement and their continuous interaction. We conducted models controlling for age because elderly subjects have been shown to move more than younger subjects,1 but this predictor was not significant in our group. We also conducted the analyses without age in the model to prevent the possible age/movement collinearity from suppressing any sleep/movement relationships. These models were nearly identical to models including age.
SUPPLEMENTAL RESULTS
We present a series of analyses that indicate on the group level that we are recording sleepiness as opposed to sleep.
First, the resting-state data were collected using an eyes-open paradigm. Eyes-open resting state exhibits stronger correlations within the default mode network (DMN) than does eyes-closed resting state,1 possibly due to fewer lapses into stage 1 NREM sleep or due to a heightened state of vigilance. Additionally, the resting-state data were acquired in two sequential runs. Instructions were given before each run, and each required a response with a button press before each run was initiated. This, too, limits the opportunity for sleep.
Second, previous work has indicated that ESS ratings are not related to quantification of sleep latency in a population of elderly community-dwelling adults.2 That is, ESS scores are not related to how quickly elderly subjects actually fall asleep. Thus, patients with higher ESS scores may not necessarily be biased to fall asleep in the scanner more quickly.
Third, we analyzed subject movement during our resting-state scans. We hypothesize that subjects who may have fallen asleep during the scan will fit into one of the following groups: (1) lower average movement due to decreased muscle tone, or (2) above-average large amplitude sharp movements due to hypnic myoclonia.3
Other network seed locations
None of the groups significantly differ for mean movement (Figure S1). The mean movement for the high ESS group is arithmetically, but not statistically, greater than the low group, indicating that it is unlikely that subjects in the high group have sleep-related muscle atonia in comparison with the low group. There is also no linear effect of ESS on mean movement (r = -0.064, P = 0.854). However, it is possible that mean movement may be contaminated by hypnic jerks. Therefore, we repeated the aforementioned analyses using a count of repetition times exceeding threshold. Again, we see no significant differences (Figure S1B). Again, running counter to the hypotheses of high ESS subjects falling asleep in the scanner, the low sleepiness group contains the highest absolute values of large movements, as well as an arithmetically higher mean. There is also no linear relationship (r = 0.071, P = 0.696). Finally, no predictors were significant in a model including cutoff volumes, mean movement, and the continuous interaction of these two predictors, controlling for age (model r2 = 0.008, P = 0.586). Combined, these results suggest that sleepier subjects are not exhibiting movement tendencies of subjects who are falling asleep. Although these data alone are insufficient to absolutely claim that the subjects are awake, they strongly implicate wakefulness during the scanning session.
Movement by sleepiness group. (A) Mean movement as euclidian distance moved from one repetition time (TR) to the next plotted by sleepiness group. None of the differences between group means are significant. (B) Number of volumes where TR-to-TR movement exceeded 0.75 mm or rotation exceeded 1.5 degrees. Again, none of the differences between group means are significant.
SUPPLEMENTAL REFERENCES
- 1.Van Dijk KRA, Sabuncu MR, Buckner RL. The influence of head motion on intrinsic functional connectivity MRI. Neuroimage. 2012;59:431–8. doi: 10.1016/j.neuroimage.2011.07.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Buysse DJ, Hall ML, Strollo PJ, et al. Relationships between the Pittsburgh Sleep Quality Index (PSQI), Epworth Sleepiness Scale (ESS), and clinical/polysomnographic measures in a community sample. J Clin Sleep Med. 2008;4:563–71. [PMC free article] [PubMed] [Google Scholar]
- 3.Carskadon MA, Dement WC. Monitoring and staging human sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and practice of sleep meedicine. ed 5. St. Louis: Elsevier Saunders; 2011. [Google Scholar]
- 4.Yeo BTT, Krienen FM, Sepulcre J, et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 2011;106:1125–65. doi: 10.1152/jn.00338.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Biswal B, Yetkin FZ, Haughton VM, Hyde JS. Functional connectivity in the motor cortex of resting human brain using echo-planar MRI. Magn Reson Med. 1995;34:537–41. doi: 10.1002/mrm.1910340409. [DOI] [PubMed] [Google Scholar]
REFERENCES
- 1.Webb WB, Agnew HW. Are we chronically sleep deprived? Bull Psychonomic Soc. 1975;6:47–8. [Google Scholar]
- 2.Bonnet MH, Arand DL. We are chronically sleep deprived. Sleep. 1995;18:908–11. doi: 10.1093/sleep/18.10.908. [DOI] [PubMed] [Google Scholar]
- 3.Gottlieb DJ, Redline S, Nieto FJ, et al. Association of usual sleep duration with hypertension: the Sleep Heart Health Study. Sleep. 2006;29:1009–14. doi: 10.1093/sleep/29.8.1009. [DOI] [PubMed] [Google Scholar]
- 4.Van Dongen HPA, Maislin G, Mullington JM, Dinges DF. The cumulative cost of additional wakefulness: dose-response effects on neurobehavioral functions and sleep physiology from chronic sleep restriction and total sleep deprivation. Sleep. 2003;26:117–26. doi: 10.1093/sleep/26.2.117. [DOI] [PubMed] [Google Scholar]
- 5.Folkard S, Lombardi DA, Tucker PT. Shiftwork: safety, sleepiness and sleep. Ind Health. 2005;43:20–3. doi: 10.2486/indhealth.43.20. [DOI] [PubMed] [Google Scholar]
- 6.Flemons WW, Tsai W. Quality of life consequences of sleep-disordered breathing. J Allergy Clin Immunol. 1997;99:S750–6. doi: 10.1016/s0091-6749(97)70123-4. [DOI] [PubMed] [Google Scholar]
- 7.Singh NA, Clements KM, Fiatarone MA. A randomized controlled trial of the effect of exercise on sleep. Sleep. 1997;20:95–101. doi: 10.1093/sleep/20.2.95. [DOI] [PubMed] [Google Scholar]
- 8.Harrison Y, Horne JA. Sleep loss and temporal memory. Q J Exp Psychol A. 2000;53:271–9. doi: 10.1080/713755870. [DOI] [PubMed] [Google Scholar]
- 9.Jones K, Harrison Y. Frontal lobe function, sleep loss and fragmented sleep. Sleep Med Rev. 2001;5:463–75. doi: 10.1053/smrv.2001.0203. [DOI] [PubMed] [Google Scholar]
- 10.Chee MWL, Choo WC. Functional imaging of working memory after 24 hr of total sleep deprivation. J Neurosci. 2004;24:4560–7. doi: 10.1523/JNEUROSCI.0007-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Goel N, Rao H, Durmer JS, Dinges DF. Neurocognitive consequences of sleep deprivation. Semin Neurol. 2009;29:320–39. doi: 10.1055/s-0029-1237117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Basner M, Rao H, Goel N, Dinges DF. Sleep deprivation and neurobehavioral dynamics. Curr Opin Neurobiol. 2013:1–10. doi: 10.1016/j.conb.2013.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Buysse DJ, Hall ML, Strollo PJ, et al. Relationships between the Pittsburgh Sleep Quality Index (PSQI), Epworth Sleepiness Scale (ESS), and clinical/ polysomnographic measures in a community sample. J Clin Sleep Med. 2008;4:563–71. [PMC free article] [PubMed] [Google Scholar]
- 14.Foley D, Monjan A, Masaki K, et al. Daytime sleepiness is associated with 3-year incident dementia and cognitive decline in older Japanese-American men. J Am Geriatr Soc. 2001;49:1628–32. doi: 10.1046/j.1532-5415.2001.t01-1-49271.x. [DOI] [PubMed] [Google Scholar]
- 15.Jelicic M, Bosma H, Ponds RWHM, Van Boxtel MPJ, Houx PJ, Jolles J. Subjective sleep problems in later life as predictors of cognitive decline. Report from the Maastricht Ageing Study (MAAS) Int J Geriatr Psychiatry. 2002;17:73–7. doi: 10.1002/gps.529. [DOI] [PubMed] [Google Scholar]
- 16.Ohayon MM, Vecchierini M-F. Daytime sleepiness and cognitive impairment in the elderly population. Arch Intern Med. 2002;162:201–8. doi: 10.1001/archinte.162.2.201. [DOI] [PubMed] [Google Scholar]
- 17.Ohayon MM, Vecchierini M-F. Normative sleep data, cognitive function and daily living activities in older adults in the community. Sleep. 2005;28:981–9. [PubMed] [Google Scholar]
- 18.Duffy JF, Willson HJ, Wang W, Czeisler CA. Healthy older adults better tolerate sleep deprivation than young adults. J Am Geriatr Soc. 2009;57:1245–51. doi: 10.1111/j.1532-5415.2009.02303.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Blagrove M, Alexander C, Horne JA. The effects of chronic sleep reduction on the performance of cognitive tasks sensitive to sleep deprivation. Appl Cognitive Psych. 1995;9:21–40. [Google Scholar]
- 20.Durmer JS, Dinges DF. Neurocognitive consequences of sleep deprivation. Semin Neurol. 2005;25:117–29. doi: 10.1055/s-2005-867080. [DOI] [PubMed] [Google Scholar]
- 21.Greicius MD, Krasnow B, Reiss AL, Menon V. Functional connectivity in the resting brain: a network analysis of the default mode hypothesis. Proc Natl Acad Sci U S A. 2003;100:253–8. doi: 10.1073/pnas.0135058100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci U S A. 2005;102:9673–8. doi: 10.1073/pnas.0504136102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.van Dijk KRA, Hedden T, Venkataraman A, Evans KC, Lazar SW, Buckner RL. Intrinsic functional connectivity as a tool for human connectomics: theory, properties, and optimization. J Neurophysiol. 2010;103:297–321. doi: 10.1152/jn.00783.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.De Luca M, Beckmann CF, De Stefano N, Matthews PM, Smith SM. fMRI resting state networks define distinct modes of long-distance interactions in the human brain. Neuroimage. 2006;29:1359–67. doi: 10.1016/j.neuroimage.2005.08.035. [DOI] [PubMed] [Google Scholar]
- 25.Greicius MD, Supekar K, Menon V, Dougherty RF. Resting-state functional connectivity reflects structural connectivity in the default mode network. Cereb Cortex. 2009;19:72–8. doi: 10.1093/cercor/bhn059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Buckner RL, Andrews-Hanna JR, Schacter DL. The brain's default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci. 2008;1124:1–38. doi: 10.1196/annals.1440.011. [DOI] [PubMed] [Google Scholar]
- 27.Spreng RN, Mar RA, Kim ASN. The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: a quantitative meta-analysis. J Cogn Neurosci. 2009;21:489–510. doi: 10.1162/jocn.2008.21029. [DOI] [PubMed] [Google Scholar]
- 28.Anticevic A, Repovs G, Shulman GL, Barch DM. When less is more: TPJ and default network deactivation during encoding predicts working memory performance. Neuroimage. 2010;49:2638–48. doi: 10.1016/j.neuroimage.2009.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.He J, Carmichael O, Fletcher E, et al. Influence of functional connectivity and structural MRI measures on episodic memory. Neurobiol Aging. 2012 doi: 10.1016/j.neurobiolaging.2011.12.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Greicius MD, Srivastava G, Reiss AL, Menon V. Default-mode network activity distinguishes Alzheimer's disease from healthy aging: evidence from functional MRI. Proc Natl Acad Sci U S A. 2004;101:4637–42. doi: 10.1073/pnas.0308627101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sorg C, Riedl V, Mühlau M, et al. Selective changes of resting-state networks in individuals at risk for Alzheimer's disease. Proc Natl Acad Sci U S A. 2007;104:18760–5. doi: 10.1073/pnas.0708803104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brier MR, Thomas JB, Snyder AZ, et al. Loss of intranetwork and internetwork resting state functional connections with Alzheimer's disease progression. J Neurosci. 2012;32:8890–9. doi: 10.1523/JNEUROSCI.5698-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Damoiseaux JS, Prater KE, Miller BL, Greicius MD. Functional connectivity tracks clinical deterioration in Alzheimer's disease. Neurobiol Aging. 2012;33:828.e19–30. doi: 10.1016/j.neurobiolaging.2011.06.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Petrella JR, Sheldon FC, Prince SE, Calhoun VD, Doraiswamy PM. Default mode network connectivity in stable vs progressive mild cognitive impairment. Neurology. 2011;76:511–7. doi: 10.1212/WNL.0b013e31820af94e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Horovitz SG, Braun AR, Carr WS, et al. Decoupling of the brain's default mode network during deep sleep. Proc Natl Acad Sci U S A. 2009;106:11376–81. doi: 10.1073/pnas.0901435106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Greicius MD, Kiviniemi V, Tervonen O, et al. Persistent default-mode network connectivity during light sedation. Hum Brain Mapp. 2008;29:839–47. doi: 10.1002/hbm.20537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Horovitz SG, Fukunaga M, de Zwart JA, et al. Low frequency BOLD fluctuations during resting wakefulness and light sleep: a simultaneous EEG-fMRI study. Hum Brain Mapp. 2008;29:671–82. doi: 10.1002/hbm.20428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Larson-Prior LJ, Zempel JM, Nolan TS, Prior FW, Snyder AZ, Raichle ME. Cortical network functional connectivity in the descent to sleep. Proc Natl Acad Sci U S A. 2009;106:4489–94. doi: 10.1073/pnas.0900924106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sämann PG, Wehrle R, Hoehn D, et al. Development of the brain's default mode network from wakefulness to slow wave sleep. Cereb Cortex. 2011;21:2082–93. doi: 10.1093/cercor/bhq295. [DOI] [PubMed] [Google Scholar]
- 40.Sämann PG, Tully C, Spoormaker VI, et al. Increased sleep pressure reduces resting state functional connectivity. MAGMA. 2010:375–89. doi: 10.1007/s10334-010-0213-z. [DOI] [PubMed] [Google Scholar]
- 41.De Havas JA, Parimal S, Soon CS, Chee MWL. Sleep deprivation reduces default mode network connectivity and anti-correlation during rest and task performance. Neuroimage. 2012;59:1745–51. doi: 10.1016/j.neuroimage.2011.08.026. [DOI] [PubMed] [Google Scholar]
- 42.Johns MW. A new method for measuring daytime sleepiness: the Epworth sleepiness scale. Sleep. 1991;14:540–5. doi: 10.1093/sleep/14.6.540. [DOI] [PubMed] [Google Scholar]
- 43.Johns MW. Reliability and factor analysis of the Epworth Sleepiness Scale. Sleep. 1992;15:376–81. doi: 10.1093/sleep/15.4.376. [DOI] [PubMed] [Google Scholar]
- 44.Spira AP, Beaudreau SA, Stone KL, et al. Reliability and validity of the Pittsburgh Sleep Quality Index and the Epworth Sleepiness Scale in older men. J Gerontol A Biol Sci Med Sci. 2012;67:433–9. doi: 10.1093/gerona/glr172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Beaudreau SA, Spira AP, Stewart A, et al. Validation of the Pittsburgh Sleep Quality Index and the Epworth Sleepiness Scale in older black and white women. Sleep medicine. 2012;13:36–42. doi: 10.1016/j.sleep.2011.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Johns MW. Sensitivity and specificity of the multiple sleep latency test (MSLT), the maintenance of wakefulness test and the epworth sleepiness scale: failure of the MSLT as a gold standard. J Sleep Res. 2000;9:5–11. doi: 10.1046/j.1365-2869.2000.00177.x. [DOI] [PubMed] [Google Scholar]
- 47.Kwong KK, Belliveau JW, Chesler DA, et al. Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc Natl Acad Sci U S A. 1992;89:5675–9. doi: 10.1073/pnas.89.12.5675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ogawa S, Tank DW, Menon R, et al. Intrinsic signal changes accompanying sensory stimulation: functional brain mapping with magnetic resonance imaging. Proc Natl Acad Sci U S A. 1992;89:5951–5. doi: 10.1073/pnas.89.13.5951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.van der Kouwe AJW, Benner T, Salat DH, Fischl B. Brain morphometry with multiecho MPRAGE. Neuroimage. 2008;40:559–69. doi: 10.1016/j.neuroimage.2007.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ashburner J. A fast diffeomorphic image registration algorithm. Neuroimage. 2007;38:95–113. doi: 10.1016/j.neuroimage.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 51.Killgore WDS, Schwab ZJ, Kipman M, DelDonno SR, Weber M. Voxel-based morphometric gray matter correlates of daytime sleepiness. Neurosci Lett. 2012;518:10–3. doi: 10.1016/j.neulet.2012.04.029. [DOI] [PubMed] [Google Scholar]
- 52.Genovese CR, Lazar NA, Nichols T. Thresholding of statistical maps in functional neuroimaging using the false discovery rate. Neuroimage. 2002;15:870–8. doi: 10.1006/nimg.2001.1037. [DOI] [PubMed] [Google Scholar]
- 53.Freire L, Mangin JF. Motion correction algorithms may create spurious brain activations in the absence of subject motion. Neuroimage. 2001;14:709–22. doi: 10.1006/nimg.2001.0869. [DOI] [PubMed] [Google Scholar]
- 54.Freire L, Roche A, Mangin J. What is the best similarity measure for motion correction in fMRI time series? IEEE Trans Med Imaging. 2002;21:470–84. doi: 10.1109/TMI.2002.1009383. [DOI] [PubMed] [Google Scholar]
- 55.Vincent JL, Snyder AZ, Fox MD, et al. Coherent spontaneous activity identifies a hippocampal-parietal memory network. J Neurophysiol. 2006;96:3517–31. doi: 10.1152/jn.00048.2006. [DOI] [PubMed] [Google Scholar]
- 56.Ward AM, Schultz AP, Huijbers W, et al. The parahippocampal gyrus links the default-mode cortical network with the medial temporal lobe memory system. Hum Brain Mapp. 2013. Feb 13, [Epub ahead of print]. http://dx.doi.org/10.1002/hbm.22234. [DOI] [PMC free article] [PubMed]
- 57.Zar JH. Biological analysis. Upper Saddle River, NJ: Prentice-Hall; 1996. [Google Scholar]
- 58.Yeo BTT, Krienen FM, Sepulcre J, et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol. 2011;106:1125–65. doi: 10.1152/jn.00338.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bluhm RL, Osuch EA, Lanius RA, et al. Default mode network connectivity: effects of age, sex, and analytic approach. Neuroreport. 2008;19:887–91. doi: 10.1097/WNR.0b013e328300ebbf. [DOI] [PubMed] [Google Scholar]
- 60.Damoiseaux JS, Beckmann CF, Arigita EJS, et al. Reduced resting-state brain activity in the “default network” in normal aging. Cereb Cortex. 2008;18:1856–64. doi: 10.1093/cercor/bhm207. [DOI] [PubMed] [Google Scholar]
- 61.Grady CL, Protzner AB, Kovacevic N, et al. A multivariate analysis of age-related differences in default mode and task-positive networks across multiple cognitive domains. Cereb Cortex. 2010;20:1432–47. doi: 10.1093/cercor/bhp207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Carrier J, Land S, Buysse DJ, Kupfer DJ, Monk TH. The effects of age and gender on sleep EEG power spectral density in the middle years of life (ages 20-60 years old) Psychophysiology. 2001;38:232–42. [PubMed] [Google Scholar]
- 63.Dijk D-J, Groeger JA, Stanley N, Deacon S. Age-related reduction in daytime sleep propensity and nocturnal slow wave sleep. Sleep. 2010;33:211–23. doi: 10.1093/sleep/33.2.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pace-Schott EF, Spencer RMC. Age-related changes in the cognitive function of sleep. Prog Brain Res. 2011;191:75–89. doi: 10.1016/B978-0-444-53752-2.00012-6. [DOI] [PubMed] [Google Scholar]
- 65.Killgore WDS, Schwab ZJ, Weiner MR. Self-reported nocturnal sleep duration is associated with next-day resting state functional connectivity. Neuroreport. 2012;23:741–5. doi: 10.1097/WNR.0b013e3283565056. [DOI] [PubMed] [Google Scholar]
- 66.Vyazovskiy VV, Olcese U, Hanlon EC, Nir Y, Cirelli C, Tononi G. Local sleep in awake rats. Nature. 2011;472:443–7. doi: 10.1038/nature10009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Voss MW, Erickson KI, Prakash RS, et al. Functional connectivity: a source of variance in the association between cardiorespiratory fitness and cognition? Neuropsychologia. 2010;48:1394–406. doi: 10.1016/j.neuropsychologia.2010.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Andrews-Hanna JR, Snyder AZ, Vincent JL, et al. Disruption of large-scale brain systems in advanced aging. Neuron. 2007;56:924–35. doi: 10.1016/j.neuron.2007.10.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Fishbein W. Disruptive effects of rapid eye movement sleep deprivation on long-term memory. Physiol Behav. 1971;6:279–82. doi: 10.1016/0031-9384(71)90155-7. [DOI] [PubMed] [Google Scholar]
- 70.Richardson GS, Carskadon MA, Flagg W, Van den Hoed J, Dement WC, Mitler MM. Excessive daytime sleepiness in man: Multiple sleep latency measurement in narcoleptic and control subjects. Electroencephalogr Clin Neurophysiol. 1978;45:621–7. doi: 10.1016/0013-4694(78)90162-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Belenky G, Wesensten NJ, Thorne DR, et al. Patterns of performance degradation and restoration during sleep restriction and subsequent recovery: a sleep dose-response study. J Sleep Res. 2003;12:1–12. doi: 10.1046/j.1365-2869.2003.00337.x. [DOI] [PubMed] [Google Scholar]
- 72.Drake CL, Roehrs TA, Burduvali E, Bonahoom A, Rosekind M, Roth T. Effects of rapid versus slow accumulation of eight hours of sleep loss. Psychophysiology. 2001;38:979–87. doi: 10.1111/1469-8986.3860979. [DOI] [PubMed] [Google Scholar]
- 73.Smith CT, Conway JM, Rose GM. Brief paradoxical sleep deprivation impairs reference, but not working, memory in the radial arm maze task. Neurobiol Learn Mem. 1998;69:211–7. doi: 10.1006/nlme.1997.3809. [DOI] [PubMed] [Google Scholar]
- 74.Esposito F, Aragri A, Latorre V, et al. Does the default-mode functional connectivity of the brain correlate with working-memory performances? Arch Ital Biol. 2009;147:11–20. [PubMed] [Google Scholar]
- 75.Sorg C, Riedl V, Perneczky R, Kurz A, Wohlschlager AM. Impact of Alzheimer's Disease on the Functional Connectivity of Spontaneous Brain Activity. Curr Alzheimer Res. 2009;6:541–53. doi: 10.2174/156720509790147106. [DOI] [PubMed] [Google Scholar]
- 76.Wang L, Laviolette PS, O'Keefe K, et al. Intrinsic connectivity between the hippocampus and posteromedial cortex predicts memory performance in cognitively intact older individuals. Neuroimage. 2010;51:910–7. doi: 10.1016/j.neuroimage.2010.02.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Zhang D, Raichle ME. Disease and the brain's dark energy. Nat Rev Neurol. 2010;6:15–28. doi: 10.1038/nrneurol.2009.198. [DOI] [PubMed] [Google Scholar]
- 78.Sperling RA, Aisen PS, Beckett LA, et al. Toward defining the preclinical stages of Alzheimer's disease: Recommendations from the National Institute on Aging and the Alzheimer's Association workgroup. Alzheimer's and Dementia. 2011:1–13. doi: 10.1016/j.jalz.2011.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Tworoger SS, Lee S, Schernhammer ES, Grodstein F. The association of self-reported sleep duration, difficulty sleeping, and snoring with cognitive function in older women. Alzheimer Dis Assoc Disord. 2006;20:41–8. doi: 10.1097/01.wad.0000201850.52707.80. [DOI] [PubMed] [Google Scholar]
- 80.Ju YE, McLeland JS, Toedebusch CD, et al. Sleep quality and preclinical Alzheimer disease. JAMA Neurol. 2013;70:587–93. doi: 10.1001/jamaneurol.2013.2334. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Other network seed locations
Movement by sleepiness group. (A) Mean movement as euclidian distance moved from one repetition time (TR) to the next plotted by sleepiness group. None of the differences between group means are significant. (B) Number of volumes where TR-to-TR movement exceeded 0.75 mm or rotation exceeded 1.5 degrees. Again, none of the differences between group means are significant.