

Abstract
For centuries mankind has stored its knowledge on paper, a remarkable biomaterial made of natural cellulose fibers. However, spontaneous cellulose degradation phenomena weaken and discolorate paper over time. The detailed knowledge of products arising from cellulose degradation is essential in understanding deterioration pathways and in improving durability of cultural heritage. In this study, for the first time, products of cellulose degradation were individually detected in solid paper samples by means of an extremely powerful proton HR-MAS NMR set-up, in combination to a wise use of both ancient and, as reference, artificially aged paper samples. Carboxylic acids, in addition to more complex dicarboxylic and hydroxy-carboxylic acids, were found in all samples studied. Since these products can catalyze further degradation, their knowledge is fundamental to improve conservation strategies of historical documents. Furthermore, the identification of compounds used in ancient production techniques, also suggests for artifacts dating, authentication and provenance.
Cellulose is the most abundant biopolymer on Earth1 and has served mankind for thousands of years as an indispensable material for human civilization, such as clothing, housing and as writing medium. Cellulose is produced in a natural way at an annual rate of 1011–1012 tonnes. It is produced either in a highly pure form, as for example in the cotton plants or by some strains of bacteria2, or combined with lignin and with other polysaccharides, so-called hemicelluloses in the cell wall of woody plants1,3.
Since the Egyptian papyri, cellulose has played an important role in human culture because it has been widely used as convenient vehicle for the acquisition, storage and dissemination of human knowledge. Paper, in particular, was invented in China in the 2nd century A.D. and, independently, in Mesoamerica in the 7th century A.D.. The art craft of making paper spread from the Far East to the Western World in the Middle Ages, and for centuries, a growing number of cultural resources have been accumulating in archives, libraries and museums, all over the world4.
Paper is produced from a dilute suspension of cellulose fibers in water, that are then drained through a sieve, pressed and dried, to obtain a sheet composed of a network of randomly interwoven fibers. The paper composition varies depending on the production period and technology employed. In the Middle Ages in Europe paper was made up of pure cellulose fibers (>90% in weight) from cotton, linen or hemp, usually obtained from rags with the addition of animal glue as a sizing agent4,5.
Cellulose is a linear homopolymer composed of β-D-glucopyranose units (C6H10O5)n, which are linked together by β-(1,4)-glycosidic bonds up to form chains with n from ≈102 to ≈104 elements. The cellulose chains have a strong tendency to aggregate into highly ordered structural entities through an extended network of both intra and intermolecular hydrogen bonds. As a consequence, a hierarchical arrangement is formed, from elemental fibrils through microfibrils up to fibers, whose diameter ranges from around 1 to around 10 μm1. Elemental fibrils, basic components of the cellulose supramolecular structure, include assembly of highly ordered (crystalline) domains and of disordered (amorphous-like) regions6. Crystalline domains constitute from around 60 to 70%, of total cellulose material depending on its origin and history.
The occurrence of a system of pores and voids, non-uniform in sizes and shapes, is complementary to the molecular architecture of cellulose fibers. A quantitative analysis on their size distribution reveals that pores diameters are approximately 1 nm when they are fully confined in the amorphous regions of cellulose elemental fibrils, otherwise their diameter can reach hundreds of nanometers within the cellulose fibers. Arrangement and contribution of pores and voids within the polymeric structure is of great importance, since they strongly influence reactivity of cellulose (due to water and external agents accessibility) as well as mechanical properties of fibers7,8,9.
Isolated cellulose possesses the most thermodynamically stable conformation amongst polysaccharides that allows its excellent stability over time10. Natural aging of paper-based materials can be accelerated by internal components and external agents often acting in a synergic manner11,12. Internal components include lignin, rosin and alum, whereas external agents comprise humidity, air, visible and/or ultraviolet radiation, pollutants, or microorganisms. On macroscopic scale, paper degradation is revealed by the weakening of mechanical properties of sheets and by widespread or localized discoloration. From a microscopic perspective paper degradation can be seen as the combination of three most important processes: i) recrystallization, ii) acid hydrolysis of β-D-(l,4)-glycosidic bonds, and iii) oxidation of the β-D-glucopyranose units; processes which, however, occur in different time scales1,11,13.
Acid hydrolysis, more profound in modern papers, mainly affects the mechanical properties of sheets, by shortening the cellulose chains. The cleavage of the glycosidic bond is catalyzed either by the hydronium cations (H3O+), formed through acidic hydrolysis of internal components, such as alum, or by the action of cellulolytic enzymes1,12.
In contrast to acid hydrolysis, oxidation of cellulose, running through the radical mechanism initiated by active oxygen species, is a complex process with many possible routes comprising numerous parallel and consecutive steps of reaction. Oxidative degradation usually starts with the conversion of single β-D-glucopyranose units to a fairly unstable oxidized derivative which induces ring opening and/or chain cleavage1,13. Destabilization of chair conformation of β-D-glucopyranose rings, accompanied by the occurrence of carbonyl groups (including chromophores14), is due to a change in the hybridization of carbon atoms composing the ring from sp3 to sp2. This in turn changes the charge distribution on oxygen atoms in the glucopyranose oxidized form which further facilitates depolymerization10.
Depolymerization, through hydrolysis of glycosidic bonds can be profoundly enhanced by acidic oxidation products such as carboxylic groups embedded into cellulose polymer and, also, by light products of degradation such as volatile compounds15. Overall, these degradation processes finally result in the formation of a great number of low molecular mass-products, such as glucopyranose, or several other organic compounds, such as acetic and formic acids16,17, oligomers of different composition (also hydrocarbons) and furfural15,18.
One of the main problems in studying the degradation of ancient artifacts is that the environmental conditions to which the samples were subjected throughout their history are unknown. However, an artificial high-temperature ageing allows constant monitoring of environmental conditions even though it cannot perfectly mimic a slow natural degradation process, which provides useful information for ancient paper11. Therefore, to understand the role of the different aging factors we used model paper samples that were artificially aged.
The study of light products of paper degradation formed during natural aging is necessary to understand degradation pathways and to achieve further progress in preservation and conservation of cellulosic materials19. Recognition and description of light products (degradation markers) can also be used to evaluate the paper degradation stage - important from the point of view of paper conservators15.
The light products of paper degradation are usually analysed by means of different experimental methods like gas or liquid chromatography15, electrophoresis16, mass spectrometry16,17, X-ray and Neutron Scattering6,7,8 and Nuclear Magnetic Resonance (NMR)18; techniques which sometimes involve special multistep sample preparation protocols. In order to simplify studies which would allow focusing on a reduced number of species, an extraction procedure can be applied to degraded cellulolytic materials. Extraction processes were used to separate specific classes of chemical compounds, such as low molar mass aliphatic organic acids, and aromatic (phenolic and furanic), or hydroxybenzoquinones17,18,19. The procedures hence require an a priori selection of the target compounds after the paper degradation.
Another method of sampling, which is non-destructive in character, is based on collecting the volatile organic compounds (VOCs) emitted by paper during its degradation, which are believed to be responsible for the characteristic smell of old books. The formation of VOCs with low molecular mass has been known since the middle of the 20th century. Recent advances in analytical methods, such as gas chromatography-mass spectrometry, allowed for the experimental identification of several compounds emitted during paper degradation, e.g. furfural and long-chain hydrocarbons10,20,21.
If analyses of precious paper-based artifacts are considered in terms of assessing their degradation state, fully non-destructive methods have to be applied.
Among different spectroscopic methods, high resolution NMR is a well-suited technique to study complex materials like: soft matter (polymers, polyelectrolites, gels, etc.), biological samples (cells, DNA, proteins, etc.) intact tissues, organs, food specimens, membranes and interfaces at or out of the equilibrium. In this frame 1H High Resolution Magic Angle Spinning (HR-MAS) represents a recent and extremely powerful NMR method for the investigation of micro quantities of semi-solid compounds without the need of time-consuming separation procedures. It is the technique particularly suited for the study of complex materials, like soft matter and biological samples. Specifically, it allows the characterization of metabolic phenotypes or metabolite profiles of cells, intact tissues, organs, food specimens, hydrated protein and DNA and others, under both normal and altered conditions22,23,24. For this purpose 1H HR-MAS takes also advantage of most of the classical one and two dimensional homonuclear and heteronuclear experiments23,25.
Furthermore, it enables the investigation of the effect of hydrogen bonding that is of fundamental importance during paper degradation. Hence, we take advantage of this sensitive technique to study light degradation products in ancient and artificially aged paper. In addition, NMR two dimensional (2D) experiments like COrrelation SpectroscopY (COSY) and TOtal Correlation SpectroscopY (TOCSY) can be used for the same target.
In reference to this work the difference between natural and artificial paper degradation is described just in terms of degradation products. Specifically, for the first time, the light degradation products of cellulose were individually detected in solid paper samples both artificially and naturally aged. In Fig. 1 we show the 1H HR-MAS NMR spectrum of a sample produced in Milan (Italy) in 1430 from which we are able to identify the main products of degradation of paper over the centuries. As an example Fig. 1 shows the characteristic NMR peaks that evolve directly from the hydrolytic and oxidative mechanisms leading to glycosidic bond scission of cellulose. In addition, we were able to identify an extremely interesting bio-marker in form of choline (and other amino-compounds such as phenylalanine and trimethylammine) whose presence gives evidence of ancient production methods, which employed glue from animal connective tissues (i.e. denatured collagen) to improve paper writing quality4. For example, the presence of choline which is easily found in fish tissues, provides a possible marker to address the provenance of ancient artifacts.
Figure 1. 1H HR-MAS NMR spectrum of ancient sample A3 (green line).

The chemical structures of main degradation products of cellulose and additives used in the ancient production techniques, as revealed in this study, are also shown.
The aim of our work is to demonstrate the opportunities of 1H HR-MAS NMR technique to reveal the main cellulose degradation products in ancient and artificially aged paper on site without the need of a priori extraction from the solid. This approach is hoped to open perspectives for the molecular understanding of the degradation processes of cultural heritage supported on cellulose, for their conservation and preservation. Finally, the identification of compounds used in ancient production techniques, can shed light on artifacts provenance and authentication.
Results
The 1H HR-MAS NMR spectra of D2O-hydrated samples are reported in Fig. 2: ancient samples A1, B1 and A3 in panel A, model samples P2, P2C9024 and P2C9048 in panel B. At the bottom of both panels the spectrum of cellobiose, the repeating fragment of cellulose, dissolved in D2O, is reported as a reference.
Figure 2. 1H HR-MAS NMR spectra of ancient samples A1 (red line), B1 (brown line) and A3 (dark green line) (panel A); modern samples: not aged P2 (black line), and artificially aged P2C9024 (grey line) and P2C9048 (light green line) (Panel B); cellobiose dissolved in D2O (dark blue line at the bottom of both panels).

The spectrum of the not aged model paper sample P2 (black line in Fig. 2 B), made of very pure cotton cellulose, represents a moderately simple case where only two broad bands are evident, one centered at 1.30 ppm and another much broader at 1.54 ppm. Secondary bands at 0.86 and a smoother one around 2.16 ppm are also observed. To look at these ensembles in more detail the horizontally expanded spectra are shown in Fig. 3 (from 3 to 0.5 ppm), in Fig. 4 (from 4.5 to 3 ppm) and in Fig. 5 (from 5 to 0.8 ppm).
Figure 3. Enlarged view of 1H HR-MAS NMR spectra shown in Fig. 2.

Ancient samples A1 (red line), B1 (brown line) and A3 (dark green line) (panel A); modern samples P2 (black line), P2C9024 (grey line) and P2C9048 (light green line) (Panel B); acetic acid (black line), hexanoic acid (dark green line), pyruvic acid (cyan line) and sebacic acid (dark pink line) 1H HR-MAS NMR spectra in D2O, from28, are plotted for comparison at the bottom of both panels.
Figure 4. Enlarged view of 1H HR-MAS NMR spectra shown in Fig. 2.

Ancient samples A1 (red line), B1 (brown line) and A3 (dark green line) (panel A); modern samples P2 (black line), P2C9024 (grey line) and P2C9048 (light green line) (Panel B); cellulose from28 (dark blue line), and cellobiose (purple line) 1H HR-MAS NMR spectra in D2O are plotted for comparison at the bottom of both panels.
Figure 5. Enlarged view of 1H HR-MAS NMR spectra shown in Fig. 2.

Ancient samples A1 (red line), B1 (brown line) and A3 (dark green line) (panel A); modern samples P2 (black line), P2C9024 (grey line) and P2C9048 (light green line) (Panel B); the weighted linear combination of aminoacids 1H HR-MAS NMR spectra (see text for details) obtained from on-line NMR databases28,29 (blue line) is plotted for comparison at the bottom of both panels. The smaller presence of collagen NMR features in A3 is due to the intense degradation of this sample14. As a consequence, collagen underwent heavy chemical and biological degradation processes which have strongly reduced its original amount in the paper.
The detected resonances in NMR spectra were assigned by means of the well established NMR Suite Professional version 7.1 (Chenomx, Alberta, Canada)26,27, 2D homonuclear NMR experiments and most reliable online NMR databases based on experimentally acquired spectra: the online Human Metabolome Database28 and Biological Magnetic Resonance Data Bank29 (see Methods for details).
The simplest possible interpretation of the observed ensembles involves carboxylic acids as products of cellulose oxidation. From a chemical point of view, because cellulose units are composed of 6 carbon atoms bonded in a glucopyranose ring, a degradation product resulting from oxidation coupled with ring opening, should be the hexanoic acid (C5H11COOH). Just for a visual comparison, the hexanoic acid 1H NMR spectrum in D2O (taken from the online Human Metabolome Database28) is reported at the bottom of Fig. 3. The spectrum of hexanoic acid shows four peaks (of different multiplicity) centered at 0.86, 1.30, 1.54 and 2.16 ppm. However, other carboxylic acids of lower molecular mass can also occur with similar resonances as, for instance, butyric acid (4 carbon atoms) which shows bands at 0.86, 1.54 and 2.16 ppm28.
Besides NMR pattern typical of carboxylic acids, an evident shoulder is present in the spectrum of sample P2 centered around 3.4 ppm, superimposed with several low intensity peaks (Fig. 4 B). The chemical shift of this shoulder drops into the range of the characteristic frequencies of cellobiose, that is between 3 and 4 ppm (Fig. 2, dark blue line).
These results show that non-negligible amount of carboxylic acids is present even in the not artificially aged sample (P2) made of pure cellulose (Fig. 2, black line). The fact that the characteristic bands of carboxylic acids are more intense than those of cellulose is due to higher sensitivity of 1H HR-MAS NMR to amorphous regions in comparison with crystalline regions of cellulose (prevailing in samples). It is noteworthy that degradation processes take place within amorphous regions, which makes HR-MAS technique, especially, useful in studying degradation processes and recognizing degradation products in ancient samples.
As the paper ages (by applying high temperature and humidity), sharp peak components increase whereas the broad resonances decrease (Fig. 2 B, grey and green lines). This tendency is probably due to the fact that the paper is being hydrated during the aging process, enabling the spectra to reveal the signals of mobile cellulose-like species (3–4 ppm) and light products of cellulose degradation (0–3 ppm). Indeed, water is a known cellulose plasticizer. In fact, upon artificial aging of P2 samples, as the cellulose structure swells, making the cellulose fibers more accessible to water and oxygen, the increased molecular mobility makes evident new features in the 1H NMR spectra. For moderately aged sample P2C9024, even though peaks at 0.86, 1.30, 1.54 and 2.16 ppm are still discernible, new relevant features appear. Firstly, sharp peaks at 1.91 and 8.45 ppm (Fig. 2 B) can be attributed to the simplest carboxylic acids, acetic and formic acids, respectively28. Furthermore, a sharp peak at about 2.40 ppm can be attributed to dicarboxylic acids with 4 carbon atoms such as succinic and pyruvic acids (Fig. 3 B)28. By means of Chenomx package even minor compounds were safely identified, as for example, methyl malonic acid (a C-methylated derivative of malonic acid), which shows peaks at about 1.12 and 3.07 ppm (Fig. 3 B and Fig. 4 B). The presence of these acidic compounds reinforces the hypothesis that the degradation processes are initially oxidative in nature16.
Apart from carboxylic acid signals, other features are also evident in the spectrum of P2C9024: a broad peak at 1.99 ppm, which seems also present but less intense in P2, and several intense sharp peaks covering the frequency range from 3.20 to 4.52 ppm and at about 5.2 ppm. These well resolved peaks are assigned to cellobiose (bottom of Fig. 2) and their strong increase in P2C9024 indicates an enhanced presence of water within the cellulose fibers. This is accompanied by the cleavage of intra-molecular hydrogen bonding in cellulose, which at this stage of degradation makes more degraded cellulose samples more accessible to oxygen active species and thus prone to further oxidation. An increase of cellobiose-like fragments can be accounted for by fractionation of cellulose polymer induced by hydrolysis and oxidation3,8.
In the spectrum of the most artificially aged P2C9048 sample (Fig. 2 B, green line) the previously described peaks are essentially present with stronger intensity. However, all features are superimposed to a much flatter background than that observed for P2 and P2C9024. In particular, the large resonance centered at about 1.99 ppm is now reduced to an evident but low intensity peak. This could be explained in terms of consumption of amorphous regions of cellulose fibers due to progressive degradation: they become disrupted and cellulose polymers are transformed into degradation products like carboxylic and dicarboxylic acids. In this way resonances of poorly mobile compounds that were broad become narrower as the compounds become more mobile in degraded cellulose plasticized by water. This finding is in agreement with previously observed results which describe increasing recrystallization of cellulose upon ageing30.
All compounds detected in modern artificially aged samples are also found in ancient paper samples A1, B1 and A3, although the bands assigned to them show slightly different shapes and intensities (Fig. 2 A, Fig. 3 A, Fig. 4 A). 1H NMR spectra of ancient samples A1, B1 and A3 (Fig. 2 A) are characterized by a flat background similar to that observed for the most degraded modern sample P2C9048. This confirms that degradation processes affect amorphous regions which become disrupted, then more mobile, so providing well resolved sharp peaks in 1H NMR spectra. This can also be inferred from the sharpening of the peak centered at about 1.99 ppm.
In ancient samples, the characteristics peaks of cellobiose (bottom of Fig. 2) are less evident. Most intense peaks are likely due to hydroxy-carboxylic acids, such as glucaric and lactic acids. Glucaric acid is derived from the oxidation of D-glupyranose, which can be also oxidized to pyruvic acid and finally transformed in lactic acid31. Both these compounds are identified in our samples. In particular, the cross-peaks of lactic acid are also very intense in the COSY and TOCSY spectra (Fig. 6 and 7, respectively). In fact, lactic acid cross-peaks are well evident in both 2D spectra at about 1.3 and 4.1 ppm. In the following cross-peaks will be indicated within brackets, such as (1.3, 4.1) ppm.
Figure 6. COSY spectrum of ancient paper A3.

In the graph are shown the compounds that origin the most intense signals.
Figure 7. TOCSY spectrum of ancient paper A3.

In the graph are shown the compounds that origin the most intense signals.
Interestingly, ancient samples show bands which could be assigned to several amino-acids. In particular, the peak at about 1.46 ppm can be assigned to alanine (Ala) whereas the broad one at about 1.6–1.75 ppm to arginine (Arg). Other peaks from 1.96 to 2.20 ppm and from 2.30 to 2.44 ppm are assigned to proline (Pro), glutamic acid (Glu) and hydroxyproline (Hyp). Characteristic peaks of phenylalanine can be also identified at about 3.1, 3.3, 4.05 and around 7.3 ppm, some of them originate evident TOCSY cross-peaks at (3.1, 3.3) and (3.1, 4.05) ppm (Fig. 7).
The presence of these amino-acids gives evidence of ancient production methods, which employed glue from animal connective tissues (gelatin, i.e. denatured collagen) to improve paper writing quality4. Collagen consists of a triple chain of amino-acids, the most abundant of which are glycine (Gly), proline (Pro) and alanine (Ala).
For a visual comparison between spectra of ancient paper and collagen, we used a weighted linear combination, I, of single spectra (in aqueous solution) of most abundant amino-acids in collagen (Gly, Pro, Ala, Hyp, Glu, Asp and Arg), obtained from on-line NMR databases28,29 (bottom of Fig. 5):
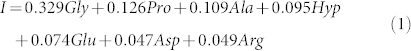 |
Contributions of spectra of amino-acids have been chosen accordingly to the average amino-acid composition for fish and mammal skin, and considering only those present at least at 4.9 wt%32. The agreement between spectra of ancient paper and collagen seems better for A1 and B1 samples. This can be explained by the fact that A3 sample was wet in the past and, as a result, it is presently characterized by an intense level of degradation14. As a consequence, collagen underwent heavy chemical and biological degradation processes which have strongly reduced its original amount in the paper, as can be observed nowadays by means of our NMR approach. Furthermore, it should be noted that the used linear combination could not well account for the type of collagen employed to size the specific ancient papers. Finally, the collagen peak positions and shapes could be significantly modified because gelatin in ancient samples is in solid form and could be degraded.
It is also interesting to note that formic acid is a final degradation product of glycine (the most abundant amino-acid in collagen). This observation can justify the weak intensity of the characteristic glycine peak at about 3.54 ppm, and consequently the stronger intensity of the formic acid peak at about 8.45 ppm with respect to the modern samples made of pure cellulose (Fig. 2 A).
To summarize briefly, except for cellulose oxidation, as clearly shown in the case of modern samples, carboxylic acids could also originate from collagen degradation, even if it is not simple to discriminate the origin of these compounds in ancient samples.
The intense five peaks between 3.19 and 3.30 ppm are very well resolved in the most degraded ancient sample A3 (Fig. 4 A). In samples A1 and B1 only 3 less resolved peaks appear in the same region of chemical shift at about 3.21, 3.23 and 3.26 ppm. In this region amines derivatives such as trimethylamine may appear. In addition, an isolated sharp peak at about 2.81 ppm can be attributed to trimethylamine which together with other amines, may evolve as a degradation product of plant and animal origin. Particularly, it can be found in animal tissue, especially in fish. Moreover, the sharp peak at about 3.26 ppm can be assigned to trimethylamine oxide, whereas that at about 3.2 ppm to carnitine. Another compound of the same class is choline, a precursor of trimethylamine, which displays peaks at about 3.1, 3.5 and 4.1 ppm.
COSY and TOCSY analyses confirm and improve the identification of these compounds (Fig. 6 and 7, respectively). The COSY cross-peak at (1.54, 2.16) ppm in sample A3 confirms the presence of hexanoic and adipic acids, whereas that at (2.31, 2.48) ppm indicates the presence of 3-hydroxy butyric acid. Cross-peaks in the chemical shift region between 3.5 and 3.9 ppm are due to mobile cellulose-like signals. These last cross-peaks are also present in the TOCSY spectrum (Fig. 7), even if their number is higher and the ppm range is extended due to the different mechanism of magnetization transfer (see Methods section).
In the TOCSY spectrum the cross-peaks at about (1.30, 1.54) and (1.30, 2.16) ppm confirm the presence of hexanoic, adipic and other carboxylic and dicarboxylic acids, like sebacic acid reported at the bottom of Fig. 3. In addition, the cross-peaks at about (0.7, 1.1) ppm are the signature of the presence of fatty acids such as oleanolic acid, a compound widely present in plants. Cross-peaks at about (2.9, 3.1) ppm can be assigned to phenyl-lactic acid, which is, together with phenethylamine, a degradation product of phenylalanine.
Some cross-peaks remain unassigned as those at (1.15, 1.31) and (3.1, 3.55) ppm, however, since their chemical shift falls in the same spectral region as carboxylic- and amino-acids, respectively, they are probably due to compounds with a similar chemical structure that can derive from both cellulose and collagen degradation. In summary, the overall results suggest that our approach is able to furnish fine details about the production process of ancient paper, as for example the use of collagen for paper sizing during production, which could be very useful for historical and provenance studies.
Discussion
Ancient paper documents, even if made of durable cotton, linen or hemp cellulose fibers, are subjected to degradation during their life span. Degradation phenomena are due to acid hydrolysis, oxidation or recrystallization of cellulose polymers. However, little is known about the products of these processes and their possible risk factors for ancient documents preservation.
In this work, by means of an extremely powerful proton HR-MAS NMR set-up, for the first time, the presence of low molecular mass compounds was detected in solid fibers of cellulose. This was possible thorough comparison of ancient and artificially aged model paper samples, in combination with the knowledge of water-cellulose interaction. The experimental approach has been optimized for the analysis of paper based artifacts and was applied to deepen our understanding of degradation phenomena that occur within them. Degradation products were assigned by means of Chenomx NMR Suite, aided by correlation spectroscopy methods, such as COSY and TOCSY, and the use of NMR online databases (see Methods section).
First of all our results evidence the presence of carboxylic acids in all not aged and artificially aged model samples made of pure cellulose as well as in naturally aged samples of ancient paper. The most evident acidic compound which was found in NMR spectra was assigned to hexanoic acid. This can be accounted for the ring opening of β-D-glucopyranose, followed by oxidation. When degradation is enhanced in paper by heating at a temperature of 90°C in the presence of humidity, a larger set of different acidic products, including low molecular mass aliphatic acids, evolves over time. It has been proved that common compounds in all degraded samples are the formic and the acetic acid. More complex compounds than simple carboxylic acids, such as dicarboxylic and hydroxy-carboxylic acids, are also present. This finding shows that even at a moderate artificial aging β-D-glucopyranose units are broken to give products in form of acidic compounds.
Furthermore, the presence of the simplest carboxylic acids, acetic and formic acids, together with the cellobiose-like fragments in modern aged samples, is an indication of coupling between hydrolytic and oxidative degradation routes.
Overall, a greater number of degradation products can be identified in ancient samples which constitute some sort of their fingerprint. Interestingly, degradation phenomena in paper materials are driven by complex reactions that take place during centuries even at ambient conditions. This is extremely important for the improvement of conservation strategies, which should always use relatively low conservation temperature, low relative humidities and, possibly, adopt conservation environment with reduced oxygen content.
Our technique was also able to detect the presence of amino-acids in all ancient samples. Amino-acids are indicative of the presence of gelatin, a material which was usually used in the production of European paper in the Middle Ages as sizing compound. Another interesting outcome is the detection of impurities typical of specific animal tissues, such as choline, which can be found in fish collagen. This result opens the way to detailed studies of paper artifacts dating and authentication.
Our results comply with previous studies carried out by using very different techniques such as gas or liquid chromatography15, electrophoresis16, mass spectrometry16,17, for the detection of carboxylic acid, such as acetic and formic acids. In addition, our approach clearly shows the evidence of more complex carboxylic acids and specific minor compounds.
In summary, the knowledge of the presence of carboxylic, dicarboxylic and hydroxy-carboxylic acids in ancient paper, could play a relevant role in the optimization of conservation procedures like those based on deacidification. Furthermore, the knowledge of specific degradation products, when associated with their formation rate, could also give hints concerning condition and also age estimation of ancient materials based on cellulose. This knowledge could be used to improve conservation, preservation and restoration of cultural heritage made of cellulose, and suggests for their dating, authentication and provenance.
Methods
Ancient samples used in this research consist of small pieces of paper, produced in the 15th century in European countries, that had already been used for other investigations7,14,33. Samples showed different degrees of degradation and discoloration. These fragments originated from end leaves, meaning to say the sheets normally used by book binders at the beginning and end of a book, as protection for the main text block. This feature allows for a fairly precise determination of the age of paper samples whilst the presence of watermarks allows for a precise identification of the mill where it was produced. All ancient paper used in the experiments carried no printing. As was the custom in ancient times our samples were made of cotton and linen cellulose fibers and, to improve the paper writing quality, they were probably sized with gelatin. Ancient samples are named A1 and B1 (both made in Perpignan, France, in 1413), in good and in intermediate conservation conditions respectively, and A3 (made in Milan, Italy, in 1430) in bad conservation conditions (within a water stain)14.
Model samples, obtained from Netherlands Organization for Applied Scientific Research (TNO), are made of unbleached cotton linters, containing very low inorganic ingredients (ash content < 0.005% in weight) and no additives or lignin, with high mean degree of polymerization. These samples are named P210,14. P2 samples were artificially aged in air in a climatic chamber at relative humidity of 59% and temperature of 90°C for 24 and 48 days. We named accordingly the aged samples P2C9024 and P2C9048, respectively. In this situation the assumed degradation factors are air and water vapor. These aging conditions were designed as a compromise in order to get measurable changes in 1H NMR spectra of model paper samples within a reasonable period of time. Thus at 90°C a moderate degradation of the cellulose polymers is expected to occur10,14.
As a reference cellobiose sample was also used (obtained from Merck Chemicals Ltd, United Kingdom). The 1D HR-MAS NMR spectrum of the sample dissolved in D2O (purity 98%) was acquired in the same way as paper samples for comparisons.
The advantage of HR-MAS with respect to the static NMR technique is to provide separated information on the presence of low mass molecules spread in solid cellulose materials. In fact, because of strong dipolar interactions that produce very large resonances (water and cellulose contributions), the static NMR technique cannot be used for a microscopic characterization of paper degradation processes34,35. Several studies performed by means of Cross Polarization Magic Angle Spinning (CP-MAS) were able to investigate the local variation of the amorphous and crystalline composition, by looking at 13C signals35,36. As confirmed by this study, the determination of cellulose degradation products may be better achieved by means of 1H detection that also permits most of the classical one and two dimensional homo- and heteronuclear experiments25.
Hydrogen one and two dimensional NMR experiments on ancient and modern cellulose samples, and on cellobiose, were conducted at ambient pressure by using a Bruker Avance spectrometer operating at 700 MHz 1H resonance frequency in the experimental configuration known as Magic Angle Spinning (MAS). By tilting samples of a precise angle (about 54.7°) with respect to the direction of the applied magnetic field, the Hamiltonian term corresponding to dipolar interactions vanishes and NMR peaks become narrower25. Furthermore, by spinning the sample holder (rotor) at the magic angle by few thousands of Hertz, line broadening effects due to susceptibility differences within the sample are removed resulting in high resolution quality spectra. The sample temperature was kept at 300 K and controlled by a cold N2 flow and a heating element, calibrated against the standard CH3OH reference (4% CH3OH in CD3OD) with an accuracy of 0.2 K.
Small rectangular pieces, 0.7 × 1.8 cm2, of about 10 mg in weight, of different papers were rolled up and carefully placed in the rotor. After placing the rolled samples in the rotor 30 μl of D2O were added. The rotor was then sealed (with the corresponding insert) and, in order to have a complete hydration of samples, approximately 30 minutes were waited. 1H HR-MAS measurements were conducted by spinning the rotor at 7000 Hz at the magic angle to increase the spectral resolution.
During samples spinning at a frequency of several thousands of Hertz, an high intensity centrifugal force acts on samples pushing them towards the rotor internal surface. However, due to the fact that paper is essentially a bi-dimensional material, rolling the samples and inserting them in form of small cylinders, causes no macroscopic mechanical damages. By just drying each sample before removing it from the rotor, we were able to recover its shape and appearance as it was before measurements. However, one has to consider that the hydration process could modify the original state of paper samples.
Paper samples were hydrated with D2O in order to enhance the molecular mobility and then the detection by the NMR technique of the cellulose components and degradation products, while keeping relatively small the signal due to water contained within cellulose pores and voids. The use of deuterated water as a solvent is expected to evidence polar compounds preferentially. A common feature of all spectra is a residual HOD peak at 4.7 ppm even if for all samples a presaturation pulse was executed.
In our one-dimensional experiments we used 16000 points in the time domain and a spectral width of 20 ppm (~14 kHz). The repetition time was set to 3 s and the number of transients was 512. The duration of the hard pulse was of 7 μs with an attenuation of 3 dB. To reduce the residual water peak we use a standard presaturation pulse sequence from Bruker called zgpr. Every acquisition lasted 33 minutes. Free induction decays were multiplied by an exponential weighting function equivalent to a 1 Hz line, after which they were Fourier transformed from the time to the frequency domain and referenced to the tetramethylsilane (TMS) by means of Bruker Xwinnmr 3.5 software packages.
Individual metabolites were identified by means of NMR Suite Professional version 7.1 (Chenomx, Alberta, Canada). This is a well established software package based on an highly sophisticated targeted profiling technology which allows an easy deconvolution of complex NMR spectra into their individual components with good accuracy. The Chenomx reference libraries contain hundreds of fully searchable compound models that are automatically calibrated by adjusting compound line shapes, peak widths, and chemical shifts to better match the sample conditions. In order to confirm the chemical shift assignment performed by means of Chenomx we also carried out 2D homonuclear NMR experiments, and made use of most reliable online NMR databases based on experimentally acquired spectra: the online Human Metabolome Database28 and Biological Magnetic Resonance Data Bank29. The combination of these analyses increases the level of confidence of the performed chemical shift assignments. 1H-13C heteronuclear experiments were also attempted within this survey, however, no significant signal was detected with our set-up.
The 2D COSY and TOCSY experiments, by probing the homonuclear correlations, allow determining and characterizing the molecular structure. In particular, COSY cross-peaks correlate the chemical shifts of 1H nuclei which are J-coupled to one another. In TOCSY experiments, instead, cross-peaks are generated between all members of a coupled spin network by a net magnetization transfer from one spin to another even without direct coupling. Double-Quantum Filtered COSY using Pulsed Field Gradients (GRASP-DQF-COSY) experiments were performed by collecting 1024(F2) × 256(F1) complex data points of 32 scans each, zero filled to 512 along F1 dimension prior to Fourier transform. We used a spectral width of 3500 Hz, centered at 2 ppm to highlight the spectral region between -0.5 and 4.5 ppm, in both dimensions and a relaxation delay of 3 s. The same parameters were used for the TOCSY experiments (Bruker mlevphpr pulse sequence with presaturation) but with a spectral width of 7000 Hz (centered at 4.7 ppm) in both dimensions and a mixing time of 100 ms. Every acquisition lasted 130 minutes.
Author Contributions
C.C., D.M. and M.M. carried out the NMR measurements and performed data analysis. J.L. performed the artificial aging of paper samples. F.M. and L.P. supervised the research project. All authors discussed the results and reviewed this manuscript.
Acknowledgments
The research in Messina is supported by the Fondazione Frisone (Catania, Italy). All authors thank the Istituto Centrale per il Restauro e la Conservazione del Patrimonio Archivistico e Librario (Roma, Italy) and the Centro Italiano di Studi e Ricerche di Storia e Tecnologia della Carta ”Andrea F. Gasparinetti” Fabriano, Italy for their support. Dr. Pawel Wojcieszek is acknowledged for his close reading and correction of text in the English language.
References
- Klemm D., Philipp B., Heinze T., Heinze U. & Wagenknecht W. Comprehensive Cellulose Chemistry; Volume I: Fundamentals and Analytical Methods (WILEY-VCH Verlag GmbH, Weinheim, Germany 1998). [Google Scholar]
- Svensson A. et al. Bacterial cellulose as a potential scaffold for tissue engineering of cartilage. Biomaterials 26, 419–431 (2005). [DOI] [PubMed] [Google Scholar]
- Krassig H. A. Cellulose - Structure, Accessibility and Reactivity (Gordon and Breach Science Publisher, Singapore, 1993). [Google Scholar]
- Hunter D. Papermaking: The History and Technique of an Ancient Craft (Dover Publications, New York, 1978). [Google Scholar]
- Missori M., Righini M. & Dupont A.-L. Gelatine sizing and discoloration: A comparative study of optical spectra obtained from ancient and artificially aged modern papers. Opt. Commun. 263, 289–294 (2006). [Google Scholar]
- Nishiyama Y., Langan P. & Chanzy H. Crystal Structure and Hydrogen-Bonding System in Cellulose Iβ from Synchrotron X-ray and Neutron Fiber Diffraction. J. Am. Chem. Soc. 124, 9074–9082 (2002). [DOI] [PubMed] [Google Scholar]
- Missori M. et al. Modifications of the Mesoscopic Structure of Cellulose in Paper Degradation. Phys. Rev. Lett. 97, 238001 (2006). [DOI] [PubMed] [Google Scholar]
- De Spirito M. et al. Modifications in solvent clusters embedded along the fibers of a cellulose polymer network cause paper degradation. Phys. Rev. E 77, 041801 (2008). [DOI] [PubMed] [Google Scholar]
- Kato K. L. & Cameron R. E. A review of the relationship between thermally-accelerated ageing of paper and hornification. Cellulose 6, 23–40 (1999). [Google Scholar]
- Lojewski T. et al. FTIR and UV/vis as methods for evaluation of oxidative degradation of model paper: DFT approach for carbonyl vibrations. Carbohydrate Polymers 82, 370–375 (2010). [Google Scholar]
- Zervos S. Natural and accelerated ageing of cellulose and paper: A literature review. Cellulose: Structure and Properties, Derivatives and Industrial Uses Lejeune, A. & Deprez, T. (eds.), 155–203 (Nova Publishing, New York, 2010). [Google Scholar]
- Sterflinger K. & Pinzari F. The revenge of time: fungal deterioration of cultural heritage with particular reference to books, paper and parchment. Environ. Microbiol. 14, 559–566 (2012). [DOI] [PubMed] [Google Scholar]
- Kolar J. Mechanism of Autoxidative degradation of cellulose. Restaurator 18, 163–217 (1997). [Google Scholar]
- Mosca Conte A. et al. Role of Cellulose Oxidation in the Yellowing of Ancient Paper. Phys. Rev. Lett. 108, 158301 (2012). [DOI] [PubMed] [Google Scholar]
- Lojewski T. et al. Furfural as a marker of cellulose degradation. A quantitative approach. Appl. Phys. A-Mater. 100, 873–884 (2010). [Google Scholar]
- Souguir Z. & Dupont A.-L. Formation of Brown Lines in Paper: Characterization of Cellulose Degradation at the Wet-Dry Interface. Biomacromolecules 9, 2546–2552 (2008). [DOI] [PubMed] [Google Scholar]
- Bogolitsyna A. et al. Determination of carbohydrate- and lignin-derived components in complex effluents from cellulose processing by capillary electrophoresis with electrospray ionization-mass spectrometric detection. J. Chromatogr. A 1218, 8561–8566 (2011). [DOI] [PubMed] [Google Scholar]
- Rosenau T. et al. Chromophores in cellulosics, VI. First isolation and identification of residual chromophores from aged cotton linters. Cellulose 18, 1623–1632 (2011). [Google Scholar]
- Dupont A.-L., Seemann A. & Lavédrine B. Capillary electrophoresis with electrospray ionisation-mass spectrometry for the characterisation of degradation products in aged papers. Talanta 89, 301–309 (2012). [DOI] [PubMed] [Google Scholar]
- Strlič M., Kralj Cigić I., Kolar J., de Bruin G. & Pihlar B. Non-Destructive Evaluation of Historical Paper Based on pH Estimation from VOC Emissions. Sensors 7, 3136–3145 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattuati-Derieux A., Bonnassies-Termes S. & Lavedrine B. Characterisation of compounds emitted during natural and artificial ageing of a book. Use of headspace-solid-phase microextraction/gas chromatography/mass spectrometry. J. Cult. Herit. 7, 123–133 (2006). [Google Scholar]
- Moestue S., Sitter B., Bathen T. F., Tessem M.-B. & Gribbestad S. I. HR MAS MR Spectroscopy in Metabolic Characterization of Cancer. Curr. Top. Med. Chem. 11, 2–26 (2011). [DOI] [PubMed] [Google Scholar]
- Beckonert O. et al. High-resolution magic-angle-spinning NMR spectroscopy for metabolic profiling of intact tissues. Nat. Protoc. 5, 1019–1032 (2010). [DOI] [PubMed] [Google Scholar]
- Corsaro C. & Mallamace D. A Nuclear Magnetic Resonance study of the reversible denaturation of hydrated lysozyme. Physica A 390, 2904–2908 (2011). [Google Scholar]
- Winter W. T. & Barnhart D. HR-MAS: The Other NMR Approach to Polysaccharide Solids. Polysaccharide Materials: Performance by Design. Edgar, K. J., Heinze, T. & Buchanan, C. M. (eds), 261–270 (American Chemical Society, Washington, DC, 2010). [Google Scholar]
- Behrends V. et al. Metabolite Profiling to Characterize Disease-related Bacteria: gluconate excretion by pseudomonas aeruginosa mutants and clinical isolates from cystic. J. Biol. Chem. 288, 15098–15109 (2013), 10.1074/jbc.M112.442814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahmed S. S. S. J. Systems biology in unruptured intracranial aneurysm: a metabolomics study in serum for the detection of biomarkers. Metabolomics 1–11 (2013), 10.1007/s11306-013-0551-8. [Google Scholar]
- Wishart D. S. et al. HMDB 3.0–The Human Metabolome Database in 2013. Nucleic Acids Res. 41, D801–7 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ulrich E. L. et al. BioMagResBank. Nucleic Acids Res. 36, D402–D408 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali M., Emsley A. M., Herman H. & Heywood R. J. Spectroscopic studies of the ageing of cellulosic paper. Polymer 42, 2893–2900 (2001). [Google Scholar]
- Lehninger Albert L., Nelson David L. & Cox Michael M. Principles of Biochemistry (5th ed.) (W. H. Freeman and Company, New York, NY, 2008). [Google Scholar]
- Szpak P. Fish bone chemistry and ultrastructure: implications for taphonomy and stable isotope analysis. J. Archaeol. Sci. 38, 3358–3372 (2011). [Google Scholar]
- Missori M., Righini M. & Selci S. Optical reflectance spectroscopy of ancient papers with discoloration or foxing. Opt. Commun. 231, 99 (2004). [Google Scholar]
- Topgard D. & Söderman O. Diffusion of Water Absorbed in Cellulose Fibers Studied with 1H-NMR. Langmuir 17, 2694–2702 (2001). [Google Scholar]
- Capitani D., Proietti N., Ziarelli F. & Segre A. L. NMR Study of Water-Filled Pores in One of the Most Widely Used Polymeric Material: The Paper. Macromolecules 35, 5536–5543 (2002). [Google Scholar]
- Federici C. et al. Ancient Paper and Its NMR Characterisation. Sci. Tech. Cult. Herit. 5, 37–47 (1996). [Google Scholar]