

Abstract
This study aims to investigate the neuroprotective effect of the rhizome of Gastrodia elata (GE) aqueous extract on beta-amyloid(Aβ)-induced toxicity in vivo and in vitro. Transgenic Drosophila mutants with Aβ-induced neurodegeneration in pan-neuron and ommatidia were used to determine the efficacy of GE. The antiapoptotic and antioxidative mechanisms of GE were also studied in Aβ-treated pheochromocytoma (PC12) cells. In vivo studies demonstrated that GE (5 mg/g Drosophila media)-treated Drosophila possessed a longer lifespan, better locomotor function, and less-degenerated ommatidia when compared with the Aβ-expressing control (all P < 0.05). In vitro studies illustrated that GE increased the cell viability of Aβ-treated PC12 cells in dose-dependent manner, probably through attenuation of Aβ-induced oxidative and apoptotic stress. GE also significantly upregulated the enzymatic activities of catalase, superoxide dismutase, and glutathione peroxidase, leading to the decrease of reactive oxidation species production and apoptotic marker caspase-3 activity. In conclusion, our current data presented the first evidence that the aqueous extract of GE was capable of reducing the Aβ-induced neurodegeneration in Drosophila, possibly through inhibition of apoptosis and reduction of oxidative stress. GE aqueous extract could be developed as a promising herbal agent for neuroprotection and novel adjuvant therapies for Alzheimer's disease.
1. Introduction
Beta-amyloid (Aβ) protein plays a central role in Alzheimer's disease (AD). Although the exact mechanism of the disease is unknown, the devastating effect of beta-amyloid is quite clear. The protein would self-aggregate into a plaque [1], which lead to the generation of reactive oxygen species, disruption of membrane potential, and increased vulnerability to excitotoxicity, and eventually cause neuronal death [2] and related cognitive defects [3]. Recent report postulated an increasing prevalence of dementia all over the world, from 36 million in 2010 to 66 million by 2030, with majority of AD [4]. Nowadays, AD threatens our aging population with the possible loss of memory and cognitive functions and leads to increasingly heavy health care burden to our future economy. Despite advances in medical interventions, Alzheimer's disease is fatal, and presently, there is no cure. Due to the complexity of pathology, AD is not very responsive to current western medications [5, 6]. Increasing attentions have turned to the conventional medicinal herbs, which are multitargeting, to search for a novel way of AD treatment [7, 8].
Drosophila melanogaster was recently developed as a model organism for drug/herbal screening for neurodegenerative diseases. It provides several unique features such as highly stable and fully-known genetics, highly conserved disease pathways, high-throughput, and very low comparative costs [9]. Most of the genes implicated in human AD pathogenesis have Drosophila homologs, including amyloid precursor protein (APP), γ-secretase, and tau [10]. However, there are some dissimilarities, such as the absence of β-secretase, which cause a defect in endogenous production of Aβ42 [11]. In this study, the Drosophila models that overexpress human Aβ42 would be used. The neurodegeneration would result in reduced lifespan, reduced locomotor activity, histological change to the neuronal structure, and eye degeneration [10, 12]. These pathological phenotypes could be observed within a few weeks, much faster than the development of these phenotypes in transgenic mice [13]. Therefore, application of Drosophila as model of AD provides excellent tools for performing drug/herb screens to identify small molecules/herbal formula that can suppress the toxicity associated with Aβ accumulation.
There is a long history of the use of medicinal herbs in the treatment of neurological disorders, like convulsion, stroke, and epilepsy, that is, Poria cocos, Polygala tenuifolia, Uncaria rhynchophylla, Ginkgo biloba, and Lycium barbarum [8, 14]. Modern pharmacological studies revealed that Ginkgo biloba possessed neuroprotective effects towards D-galactose [15], beta-amyloid [16], and ischemia-induced neuronal death [17]. Uncaria rhynchophylla also prevented D-galactose [18], beta-amyloid [19], 6-hydroxydopamine [20], and kainic acid-induced neurotoxicity [21]. Similar neuroprotective effects were found in other commonly used herbs in China [22–25]. Rhizome of Gastrodia elata (Tianma, GE) is also one of the commonly used traditional Chinese medicines. Many studies have been performed to evaluate the neuroprotective effects of GE and its biologically active ingredients against different kinds of neuronal damages. The nonpolar extract of GE inhibited the 1-Methyl-4-phenylpyridinium and glutamate-induced apoptosis in neuronal cells [26, 27]. Additionally, the nonpolar extract of GE protected mice and rat against kainic acid [28] and aluminum chloride-induced neuronal damages [29]. Its active ingredient, gastrodin, has been shown to possess a protective effect against hypoxia injury on neurons [30]. Other active compounds, hydroxybenzyl alcohol and vanillin, could ameliorate ischemic cerebral injury in rats [31], and prevent ischemic death of hippocampal neuronal in gerbils [32], respectively. Recently, an in vitro study indicated that the aqueous extract of GE enhanced proteolytic processing of APP towards the noncytotoxic nonamyloidogenic pathway [33]. Previously, studies revealed that APP processing affected the production of Aβ, which strongly correlated to the neuronal degeneration in AD pathology [34]. Mishra et al. demonstrated that that GE was able to inhibit β-site APP-cleaving enzyme 1 activity and promote α-secretase activity [33]. The inhibition of β-site APP-cleaving enzyme 1 reduces the cleavage of APP into Aβ [35], and the activation of α-secretase increases the cleavage of APP into soluble-APP-α [36, 37].
Although the nonpolar extract of GE was found to have various neuroprotective effects, extraction of GE with water is the traditional way of preparing Chinese medicine for human consumption. The active ingredient content of aqueous extract and nonpolar extract is theoretically different, which aqueous extract should have a higher content of hydrophilic gastrodin and polysaccharides and a lower content of less hydrophilic ingredients, such as hydroxybenzaldehyde, hydroxybenzyl alcohol, vanillin, and vanillyl alcohol. According to the Chinese pharmacopeia 2010, aqueous extract of GE is a traditional Chinese medicine that is widely used for treatment of convulsive disorders, headache, dizziness, and vertigo [38]. However, there is a lack of scientific evidence to support these medical claims. Based on these previous studies and the traditional use of GE, we hypothesize that aqueous extract of GE may also be effective in protecting neurons against beta-amyloid-induced neuronal death. Moreover, there is a lack of information relating to the in vivo neuroprotective effects of GE aqueous extract. In hope of finding an extract which could modulate APP cleavage and reduce neurotoxic effect from beta-amyloid, in the present study, we aimed to investigate the neuroprotective effects of GE on beta-amyloid-induced neurodegeneration in Drosophila and its related mechanisms using pheochromocytoma (PC12) cells. The mechanism of the neuroprotective effects of GE on the downstream pathway after the cleavage of APP to Aβ were studied, including the reactive oxygen species production and the activity of the antioxidative enzyme. The apoptosis caused by Aβ was also determined by propidium iodide (PI)/Annexin V staining and confirmed by caspase-3 activity assay.
2. Materials and Methods
2.1. Materials
Aβ 25–35 peptide, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), and caspase-3 assay kit were purchased from Sigma-Aldrich (St Louis, MO, USA). RPMI medium 1640, fetal bovine serum (FBS), horse serum (HS), and 2′,7′-dichlorodihydrofluorescein diacetate (H2DCFDA) were obtained from Invitrogen (Carlsbad, CA, USA). Annexin V-FITC and propidium iodide (PI) were obtained from BD Biosciences (San Jose, CA, USA). Superoxide dismutase and glutathione peroxidase assay kits were from Cayman Chemical (Ann Arbor, MI, USA). Catalase fluorometric detection kit was obtained from Enzo Life Sciences (Farmingdale, NY, USA). Formula 4–24 instant Drosophila medium was obtained from Carolina Biological Supply Company (Burlington, NC, USA).
2.2. Herbal Materials and Extraction
The raw herbs of the rhizome of Gastrodia elata were purchased from Chinese herbal stores in the Guangdong province in Mainland China. It was chemically authenticated using thin layer chromatography in accordance to the Chinese Pharmacopoeia 2010 and deposited in the museum of the Institute of Chinese Medicine, the Chinese University of Hong Kong, with voucher specimen number of 2010-3294. For extraction, the raw herbs Tianma were firstly washed with tap water to remove any contaminants. They were then cut into small pieces. The herbs were soaked with 10-fold of water (v/w) for 1 h, followed by extraction at 100°C for 1 h. Subsequent extractions were carried out with 10-fold of water (v/w) for another 1 h. The extracts were combined and concentrated under reduced pressure to give dry Tianma powdered extract. Ultimately, 48.90 g of the aqueous extract was obtained from 100.00 g of raw GE herb. The content of gastrodin was determined to be 2.2% w/w using high performance liquid chromatography, according to the method listed in the Chinese Pharmacopeia 2010 [38], which is higher than the requirement of 0.2% w/w.
2.3. Drosophila Strains
Drosophila strains used in this study were Oregon-R-C (OR) (#5), w 1118 (#3605), and elav-GAL4 C155 (#458) (Bloomington Drosophila Stock Center, Department of Biology, Indiana University, Bloomington, IN, USA). UAS-Aβ42/CyO and GMR-Aβ42 K52 ; GMR-Aβ42 K53 heterozygous were gifts from Dr. M. Konsolaki (Rutgers University, USA). OR is a wild type Drosophila. w 1118 is a white-eye mutant with a deletion in the sex-linked white gene. Elav-GAL4 C155 is a mutant with an embryonic lethal abnormal vision (elav)-GAL4 insert on the X chromosome. UAS-Aβ42/CyO is a mutant with an UAS-Aβ42 insert and a Curly of Oster (CyO) balancer on the 2nd chromosome. GMR-Aβ42 K52; GMR-Aβ42 K53 heterozygous is a mutant with 2 copies of Glass Multiple Reporter (GMR)-Aβ42 inserts on the 3rd chromosome.
For longevity and climbing assay, genotypes of Drosophila used in this study were as follows: control: elav-GAL4 c155 /Y, Aβ42: elav-GAL4 c155 /Y; UAS-Aβ42/+; +/+. Elav-GAL4 C155 line was crossed with w 1118 line to produce control. Elav-GAL4 C155 line was crossed with UAS-Aβ42/CyO to produce elav-GAL4 c155 /Y; UAS-Aβ42/+; +/+. The genotypes of newly hatched Drosophila are different between male and female. The genotype of the male offspring is elav-GAL4 c155 /Y; UAS-Aβ42/+; +/+, while that of female offspring is elav-GAL4 c155 /w; UAS-Aβ42/+; +/+. The existence of the wild type gene in par with our elav-GAL4 c155 promoter would half the overall expression of the transgene [39]. In order to minimize the error due to genetic difference, male was chosen in the present study. For the psuedopupil assay, Drosophila genotypes were as follows: Control: OR, Aβ42: GMR- Aβ42 K52 ; GMR- Aβ42 K53 heterozygotes.
2.4. Effect of GE on Longevity of Aβ Expressing Drosophila
Genetic crosses were performed in the vials containing the diet with treatments. The normal control, which did not express Aβ, was maintained on the normal diet. The Aβ expressing control and the positive control were maintained on the normal diet and diet containing 10 mmol donepezil/g of Drosophila media, respectively, whereas the two GE groups were fed with diets containing 1 or 5 mg GE/g of Drosophila media, respectively. Newly hatched male Drosophila in each group was transferred to a new vial (30 Drosophila per vial), continued with their respective treatments, and incubated at 25°C. Dead Drosophila were counted on day 1 and 5 in a 7-day cycle, and the remaining live Drosophila were transferred to a new vial containing the same diet. The feeding lasted for 65 days. One hundred and fifty Drosophila were tested for each group.
2.5. Climbing Assay
Locomotor function of Drosophila was measured according to the climbing assay as previously reported by Lee et al. [40] with slight modifications. In brief, 30 male Drosophila were placed at the bottom of a 15 mL falcon tube and were given 10 s to climb up the tube. At the end of each trial, the number of Drosophila that climbed up to a vertical distance of 8 cm or above was recorded. Drosophila were tested on day 1 and 5 in a 7-day cycle. Each trial was performed three times. One hundred and fifty Drosophila were tested for each group.
2.6. Pseudopupil Assay
The control and Aβ42 Drosophila were treated with the same treatments as described above. Drosophila heads were examined under a light microscope (Olympus CX31; Olympus, Tokyo, Japan) as described previously [41]. Briefly, the compound eye of 5 days old Drosophila was viewed under microscope in a dark field. There were eight photoreceptors in each ommatidium, and seven of them were visible. Each photoreceptor projected a darkly staining rod, the rhabdomere, into the center of the ommatidium. Under the microscope, the rhabdomeres appeared as bright spots and rhabdomeres in each ommatidium were counted. In the control group, 7 rhabdomeres could be observed in each ommatidium. One hundred ommatidia were observed from 5 to 10 eyes, and the average rhabdomeres count per ommatidium was calculated. Three trials were conducted for each group.
2.7. Cell Culture and Drug Treatment
PC12 rat pheochromocytoma cells were obtained from American Type Culture Collection (Manassas, VA, USA) and maintained in RPMI medium 1640 supplemented with 10% (v/v) heat-inactivated HS and 5% (v/v) FBS at 37°C under 95% air/5% CO2. Cells were utilized for experiments during exponential growth.
Aβ 25–35 was dissolved in sterile distilled water at a concentration of 1.0 mM as a stock solution and preaggregated at 37°C for 7 days prior to use. Confluent cells were trypsinized, counted, and seeded on poly-L-lysine-coated 6-well culture plates at a density of 3 × 105 cells/well and incubated for 24 h. After that, cells were treated with various concentrations of GE and 20 μM of aggregated Aβ 25–35 for 48 h.
2.8. Cell Viability Assay
Cell viability was determined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT) assay. Briefly, the cells were plated on poly-L-lysine-coated 96-well culture plates at the density of 1 × 104 cells/well and incubated for 24 h. After that, the medium was replaced with fresh medium, and the cells were incubated with Aβ 25–35 (1 μM) in the presence or absence of aqueous extract of GE (250–1000 μg/mL) for 48 h. Thereafter, cells were incubated with 30 μL of MTT solution (final concentration, 1.5 mg/mL) for 4 h. The supernatant was then removed and 100 μL of dimethyl sulfoxide was added to dissolve the formazan crystal. Plates were shaken for 10 min and optical density was determined with a microplate reader at 540 nm. The optical density of control cell was 100% viability.
2.9. Flow Cytometric Detection of Apoptosis
Apoptotic cells were quantified by Annexin V-FITC and PI staining by flow cytometry. Briefly, the treated cells were trypsinized and centrifuged at 450 ×g at 25°C for 5 min. The pellet was washed twice with ice cold PBS and resuspended with Annexin V binding buffer. Annexin V-FITC and PI were added according to manufacturer's instruction and incubated in dark at room temperature for 15 min. 300 μL of binding buffer was added to each sample. The stained cells were analyzed by fluorescence-activated cell sorter (FACS). Ten thousands events were analyzed per sample.
2.10. Measurement of Apoptosis
The treated cells were collected and washed twice with ice cold PBS. PC12 cells were lysed in cell lysis buffer (50 mM Tris-HCl pH 7.4, 2 mM MgCl2, 0.1% Triton X-100) with 2 freeze/thaw cycles. The supernatant was collected after centrifugation at 15,000 ×g for 3 min; after that, the total protein concentration was determined by the bicinchoninic acid (BCA) assay, using bovine serum albumin (BSA) as a standard. The samples were then applied to caspase-3 activity assays, according to manufacturer's instructions. The activities were normalized using the total protein concentrations.
2.11. Measurement of Reactive Oxygen Species (ROS) Production
The 2,7-dichlorodihydrofluorescein diacetate (H2DCF-DA) method was used to measure intracellular ROS production. H2DCF-DA can pass through the cell membrane and oxidized by ROS to form the fluorochrome 2′,7′dichlorofluorescein (DCF). Therefore, H2DCF-DA was widely used to reflect the intracellular ROS content [42–44]. The treated cells were collected, washed twice with ice cold PBS, and incubated with H2DCF-DA (20 μM) in the dark at 37°C for 15 min. Then cells were washed once with PBS and harvested for fluorescence-activated cell sorter (FACS) analysis. Ten thousands events were analyzed per sample.
2.12. Measurement of the Antioxidative Enzyme Activities
The treated cells were collected and washed twice with ice cold PBS. The cells were lysed in cell lysis buffer (50 mM Tris-HCl pH 7.4, 2 mM MgCl2, 0.1% Triton X-100) with 2 freeze/thaw cycles. The supernatant was collected after centrifugation at 14,000 ×g for 3 min, after that the total protein concentration was determined by the bicinchoninic acid (BCA) assay, using bovine serum albumin (BSA) as a standard. The samples were then applied to antioxidative enzyme activity assays, including glutathione peroxidase (GPx), superoxide dismutase (SOD), and catalase (CAT), according to manufacturer's instructions. The activities were normalized using the total protein concentrations.
2.13. Statistical Analysis
Multiple group comparisons were performed using one-way analysis of variance (ANOVA) followed by Dunnett's test to detect intergroup differences. Comparisons for survival assay were performed using Log-Rank analysis and chi-square comparison.
All statistical analyses were performed using GraphPad Prism version 5.0 for Windows (GraphPad Software Inc., California, USA). The data were expressed as mean ± standard deviation (SD). A value of P < 0.05 was considered statistically significant.
3. Results
3.1. Tianma Prolonged the Lifespan and Improved Locomotor Abilities of Aβ-Expressing Drosophila
In the present study, we evaluated the neuroprotective effect of aqueous extract of GE, using Drosophila AD model. Before performing the experiments, we evaluated the effect of 5 and 50 mg GE extract/g of Drosophila media on food intake of Drosophila. Both GE treatments did not affect the food intake of Drosophila (data not shown), which ensured no experimental differences were due to the alteration of feeding behavior. For lifespan experiment, Aβ42 Drosophila showed a reduction of median and maximum lifespan by 17 days and 32 days when compared with control, respectively. Both GE treatments significantly improved the survival of Drosophila (Figure 1(a)). At 1 mg GE extract/g of Drosophila media, median and maximum lifespan were increased by 4 days (12.0%) and 4 days, respectively (P < 0.001 for mean increases). At 5 mg GE extract/g of Drosophila media, median and maximum lifespan were increased by 7 days (26.9%) and 7 days, respectively (P < 0.001 for mean increases).
Figure 1.
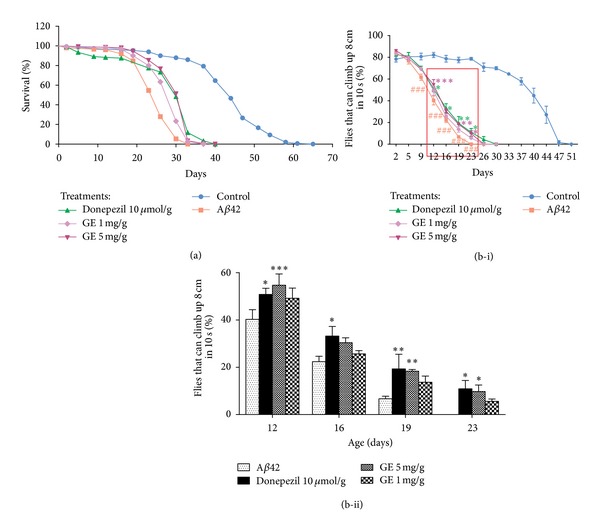
Intake of GE increases the (a) lifespan and (b-i) locomotor activity of Aβ-expressing Drosophila. The lifespan of Aβ42 group (squares in red) is shorter than the control group (circles in blue), while GE (asterisks and triangles in purple) or donepezil (triangles in green) treatments delay the mortality of the Drosophila. (b-ii) is an amplification of the region from days 12 to 23 showing the differences among the Aβ42 group and the treatment groups. The percentage of Drosophila climbing up 8 cm in 10 seconds was increased by GE or donepezil treatments when compared with Aβ42 group. Results are the means ± SEM from five independent crosses. ### P < 0.001 relative to control; *P < 0.05, **P < 0.01, ***P < 0.001 relative to Aβ42 Drosophila by one-way ANOVA for locomotor activity. Log-Rank analysis and chi-square comparison were applied to the survival data and P < 0.001 was obtained when comparing Aβ42 Drosophila and Donepezil 10 μmol/g or GE 1 mg/g or 5 mg/g treated ones (n = 150).
For locomotor abilities determination, Aβ42 Drosophila showed significant impaired locomotion from age of day 9 onwards (Figure 1(b-i)). GE-treated flies showed an improvement in locomotor activity from age of days 12 to 23. At day 12, 19, and 23, 5 mg GE extract/g of Drosophila media resulted in a 14.4%, 11.6%, and 9.74% improvement in locomotion, respectively (P < 0.001, P < 0.01, P < 0.05) (Figure 1(b-ii)) when compared with the Aβ42 Drosophila without GE treatment.
3.2. Tianma Rescued Neurodegeneration in Ommatidia of Aβ-Expressing Drosophila
We analyzed the effect of Aβ42 on degeneration of retinal tissue of Drosophila, which were mainly neurons. Aβ42 Drosophila contained significantly more degenerating rhabdomeres, compared with OregonR. The number of degenerated rhabdomeres was 3.82 ± 0.09. Aβ42 Drosophila treated with GE (1 and 5 mg/g of Drosophila media) had significantly rescued rhabdomere in each ommatidium, with an increase of 0.49 and 0.97 rhabdomere count per ommatidium, respectively (Figure 2), which reflected a preventive effect of GE on neurodegeneration. The preventive effect was comparable to donepezil medication (10 μmol/g of Drosophila media), in which there was an increase of 0.78 rhabdomere count per ommatidium than the Aβ42 Drosophila.
Figure 2.
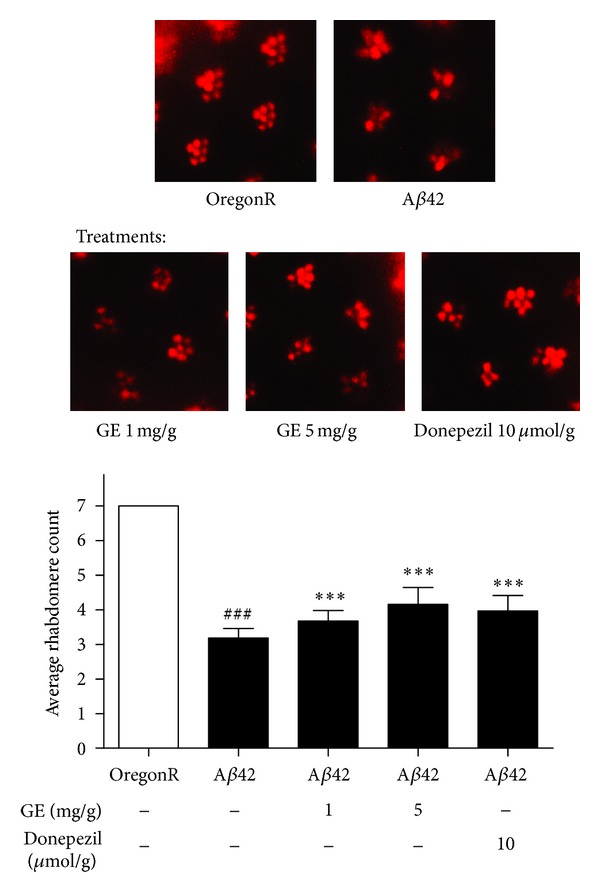
Rhabdomere count in the pseudopupil assay. Regular array of 7 ommatidia (bright red spots) was observed in OregonR eyes. Degeneration of ommatidia was observed in the Aβ42 group, while the degeneration is improved by GE or donepezil treatments. ### P < 0.001 relative to OregonR; ***P < 0.001 relative to Aβ42 Drosophila with no treatment by one-way ANOVA. Results are the means ± SEM from 3 independent crosses. One hundred ommatidia were observed from 10 eyes of 5 Drosophila from each group in each trial.
3.3. Tianma Reduced Aβ-Induced Cytotoxicity in PC12 Cells and Prevented Aβ-Induced Apoptosis
Exposure of PC12 cells to aggregated Aβ 25–35 (20 μM) for 48 h caused significant cytotoxicity. Concentration of GE in the range from 125 to 1000 μg/mL was identified to be non-toxic to PC12 cells by MTT assay (data not shown). The high concentration of GE used is also correlated to its high extraction yield in water (48.9%), compared with the yield of less than a few percent in extraction by nonpolar solvents. Our results demonstrated that GE imposed significant protective effect against Aβ 25–35-induced damage in a dose dependent manner, with the maximum effect observed at 1000 μg/mL (Figure 3). Therefore, concentration of GE in the range from 250 to 1000 μg/mL was selected for the further apoptosis study. In this regard, we investigated the effect of GE on Aβ 25–35-induced apoptosis using Annexin V-FITC and PI staining. Early apoptotic (PI: negative, Annexin V: positive) cells and late apoptotic (PI and Annexin V: positive) cells were quantified by flow cytometry. For the control group treated with Aβ 25–35 only, the normalized percentages of early and late apoptosis induced by Aβ 25–35 were 14.1 ± 3.5% and 2.6 ± 0.3%, respectively. For the treatment groups, the percentage of early and late apoptosis induced by Aβ 25–35 with treatment of GE were 9.7 ± 2.4% and 0.9 ± 0.6% for 250 μg/mL, 8.2 ± 0.3% and 0.4 ± 0.7% for 500 μg/mL, and 3.1 ± 3.1% and 0.1 ± 0.1% for 1000 μg/mL (Figure 4(b)). The results suggested that GE can reduce Aβ 25–35-induced apoptosis dose dependently. To further confirm the antiapoptotic effects of GE against Aβ 25–35-induced toxicity, the activity of crucial mediator of apoptosis caspase-3 was assessed. Caspase-3 activity was increased by 31.8 ± 13.4% with Aβ 25–35 treatment, and the increase in activity was attenuated dose dependently with treatment of GE (Figure 4(c)). At 1000 μg/mL of GE, the caspase-3 activation was totally abolished and reverted to the normal activity level of the PC12 cells without Aβ 25–35 treatment.
Figure 3.
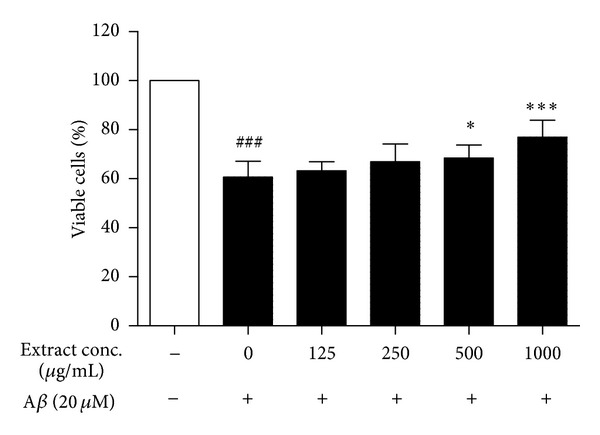
Protective effect of GE on Aβ-induced cytotoxicity in PC12 cells. Effect of 48 h treatment of GE extract on the viability of PC12 cells was determined by MTT assay. Results are the means ± SD from three separate experiments. ### P < 0.001 relative to control; *P < 0.05, ***P < 0.001 relative to Aβ treatment only by one-way ANOVA.
Figure 4.
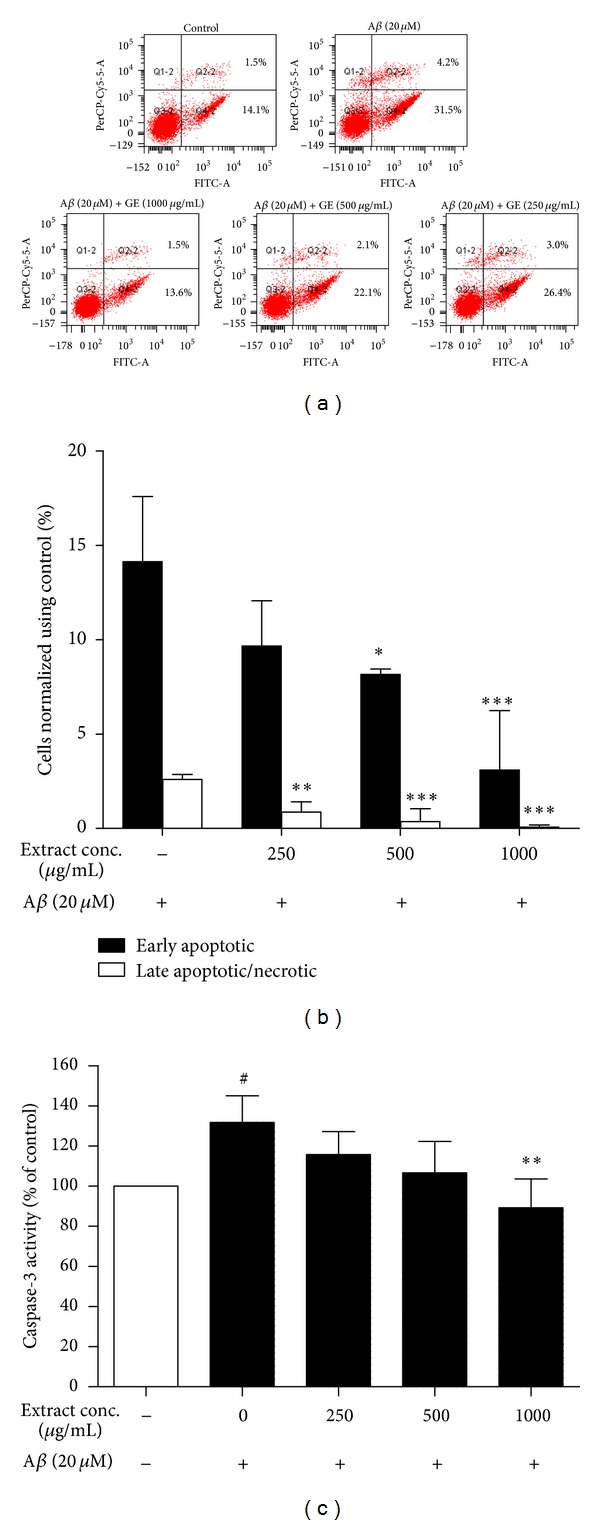
Antiapoptotic effect of GE on Aβ-induced cytotoxicity in PC12 cells. (a) Representative plots for the flow cytometric analysis. (b) GE extract reduced Aβ-induced apoptosis in flow cytometric analysis. The fluorescence intensity was measured after PC12 cells were exposed to 20 μM Aβ for 48 h, followed by incubation with Annexin V-FITC and PI for 15 min. (c) 48 h treatment of GE extract attenuated Aβ-induced activation of caspase-3. Results are the means ± SD from three separate experiments. # P < 0.05 relative to control; *P < 0.05, **P < 0.01, ***P < 0.001 relative to Aβ treatment only by one-way ANOVA.
3.4. Tianma Prevented Aβ-Induced Oxidative Stress
Figure 5(a) shows that 20 μM Aβ 25–35 elevated the production of ROS from 100% to 145.2 ± 16.3%, whereas the fluorescence intensity in GE-treated groups decreased significantly (110.9 ± 7.5%, 103.7 ± 23.1%, and 99.0 ± 15.1%, resp.). The decrease of fluorescence by GE reflected the reduction of ROS content, which possibly caused by the activation of antioxidative enzymes.
Figure 5.
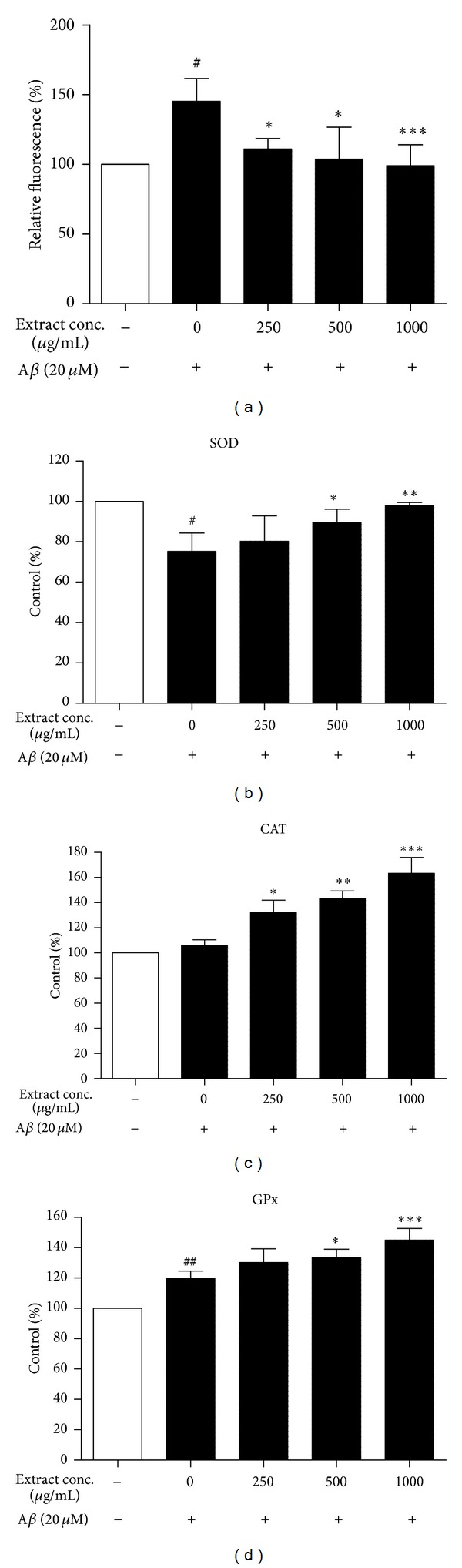
Antioxidative effect of GE on Aβ-induced cytotoxicity in PC12 cells. (a) GE extract reduced Aβ-induced oxidative stress in flow cytometric analysis of DCF positive cells. The fluorescence intensity of DCF was measured after PC12 cells were exposed to 20 μM Aβ for 48 h, followed by 20 μM H2DCF-DA for 15 min. 48 h treatment of GE extract increased the activities of antioxidative enzymes (b) superoxide dismutase (c) catalase and (d) glutathione peroxidase in 20 μM Aβ-treated cells. Results are the means ± SD from three separate experiments. # P < 0.05, ## P < 0.01 relative to control; *P < 0.05, **P < 0.01, ***P < 0.001 relative to Aβ treatment only by one-way ANOVA.
The activities of antioxidative enzymes (SOD, CAT, and GPx) in untreated PC12 cells and in those treated with 20 μM Aβ 25–35 alone or with GE together are presented in Figures 5(b)–5(d). Activity of SOD was decreased by 24.75 ± 9.07% in the cells exposed to 20 μM Aβ 25–35 (Figure 5(b)). Exposure to 20 μM Aβ 25–35 did not significantly affect the activity of CAT (Figure 5(c)) and induced a 19.70 ± 4.87% increase in activity of GPx (Figure 5(d)). With 1000 μg/mL of GE treatment, the activity of SOD was reverted to the normal activity level of the PC12 cells without Aβ 25–35 treatment, while the activity of CAT was enhanced by 63.30 ± 12.58% compared with the normal control. Moreover, 1000 μg/mL of GE further increase the activity of GPx to 45.00 ± 7.71% higher than the normal control. Overall, treatment with different doses of GE significantly and dose-dependently enhanced the activities of SOD, CAT, and GPx (Figures 5(b)–5(d)).
4. Discussion
In the present study, we have presented the first evidence that the aqueous extract of GE could significantly ameliorate the adverse morphological changes from Aβ protein in Drosophila, as indicated by improving locomotor abilities, prolonging the lifespan, and rescuing neurodegeneration in ommatidia in Aβ-expressing Drosophila. In vitro experiments showed that Aβ-treated cultures exhibited characteristic features of ROS production, apoptosis, and cell death in PC12 cells. GE aqueous extract attenuated Aβ-induced cytotoxicity effectively, probably through increasing the activities of antioxidative enzymes so as to reduce overall oxidative stress and subsequently inhibiting Aβ-induced apoptosis.
Two Drosophila lines were used to overexpress different levels of Aβ, one was using GMR promoter and one was using GAL4-UAS system. GMR promoter element directs the expression of the protein at the eye imaginal disc. The advantage of expressing only in the eye is that flies producing a highly toxic protein may still be viable. Rapid and severe degeneration of the ommatidia (eyes of Drosophila) was achieved due to the presence of two copies of gene encoding for Aβ in our Drosophila with GMR promoter [10]. For the latter one, the GAL4-UAS system is more complex. Tissue-specific expression of the UAS-Aβ42 is achieved by crossing the transgenic Drosophila with driver lines that control tissue-specific expression GAL4, which would bind with UAS to activate gene transcription. Elav-GAL4 is a commonly used pan-neuronal driver that directs the expression of transgene throughout the brain, neuronal system, and retina of the Drosophila [45]. The advantage of this model is that the lethal gene can be carried in the parents without affecting their viability and fecundity. In this study, UAS-Aβ42 would be crossed with elav C155 -GAL4 to express Aβ42 in the brain and the whole neuronal system and gradually accumulate to induce the degenerative phenotypes, such as the pathological morphologies and behavioral changes, in weeks. Therefore, to identify the efficacy against Aβ toxicity, the rationale of the current assays aimed to see whether GE aqueous extract can rescue retinal degeneration, locomotion and climbing deficits, and increase the lifespan of the flies, restoration of normal activity.
Using these two Drosophila models, the in vivo effects of GE aqueous extract in Alzheimer's disease were studied. Firstly, we found that GE aqueous extract reduced the neurotoxic effect of Aβ to ommatidia. The degree of degeneration of ommatidia reflected the extent of neurodegeneration [46], based on the fact that photoreceptors were neurons in nature. Overexpression of Aβ causes plaque formation and neuronal degeneration, which was responsible for the eye morphological changes [10, 12]. The intake of GE extract reduced the adverse effect of Aβ-associated plaque formation and rescued the eye phenotype. Similar findings were observed in the other Drosophila model with systemic pan-neuronal Aβ42 expression. GE aqueous extract significantly prolonged the lifespan and improved locomotor dysfunction of the flies. We also found that the beneficial effects of GE were comparable with the medicine donepezil. Hong et al. recently reported that Chinese traditional medicinal prescription SuHeXiang Wan improved the longevity and locomotor ability using the same Drosophila model system [47]. The results of the current study suggested that GE aqueous extract confers a therapeutic potential to AD-like pathology of Aβ42 overexpressing in different Drosophila models. In our Drosophila model, the AD-like pathology was caused by the neurotoxic Aβ produced, secreted to and aggregated in the extracellular matrix [48], yet there was no previous report on the effect of Aβ on apoptosis and oxidative stress on Drosophila. Previous reports revealed the direct proportional relationship between the manifestations of neuronal dysfunction in Drosophila, such as locomotor deficits and reduced lifespan, and aggregation rate of the Aβ, which provide evidence that the aggregated Aβ is the primary determinant of the pathological behavior in the Drosophila system [49]. To study pathogenic mechanisms, we have developed in vitro model that recapitulate many of the signature events in Aβ neurotoxicity including the accumulation extracellular aggregated Aβ, leading to apoptotic events and formation of reactive oxygen species. Basing on previous studies that were using PC12 cells as a platform to express and study the action mechanisms of Drosophila proteins [50], we postulate that the PC12 cells would be able to mimic the cellular environment of the Drosophila. Moreover, there was a well-established platform using PC12 cells and Drosophila toscreen and validate aggregation inhibitors of polyglutamine, which resulted in neurodegeneration [51, 52]. The platform suggested that PC12 cells and Drosophila would have correlation in neurodegeneration mechanisms. Therefore, PC12 cell line was used to explain the in vivo effects in the present study.
For the in vitro mechanistic studies, PC12 cell line, which is originated from transplantable rat adrenal pheochromocytoma, was used. Due to their similarity with sympathetic neurons and their reversible differentiation response to nerve growth factor [53], PC12 cells were widely used in the study of neuronal differentiation [54], neuronal function [55], and neurodegeneration [56, 57]. Aβ-induced cytotoxicity on PC12 cell line is widely used to study the AD-related neurodegeneration [58]. In the present study, we adopted this cell line and found that GE possessed protective effect against Aβ-induced cell death in MTT assay. Previously, Kim et al. had also demonstrated that the ethyl ether fraction of GE was able to protect Aβ-induced IMR-32 neuroblastoma cell death [59]. However, the content of active ingredients in the ethyl ether extract was expected to be different from the aqueous extract. Although the dose of extract used in the study was as low as 10 μg/mL, the extraction yield of the extract was only 1.12%. When comparing with the present study of extraction yield of 48.90%, the dose was equivalent to 420.61 μg/mL in the present study, which is similar to the present dose of 500 μg/mL. Moreover, the study only demonstrated the protective effect using MTT assay, but lacked further elucidation of any protective mechanisms. A complete picture from the in vivo effect to the downstream neuroprotective mechanisms was yet to be provided, and the present study was the novel one targeting this. Extensive evidence shows that neuron cell death in AD is mediated by apoptosis [60, 61]. For instance, postmortem analysis of AD brain shows that there is DNA fragmentation in neurons and glia of hippocampus and cortex as detected by TdT-mediated dUTP nick end labeling [62]. It was also found that the extracellular accumulation of Aβ, which triggers the intracellular formation of neurofibrillary tangles [63], leads to the loss of cholinergic neurons [64]. Hence, a common theory believed that the pathological neuronal loss in AD is through apoptosis, which may be caused by Aβ accumulation and cytotoxicity [2, 34]. In order to elucidate the possible mechanisms of the neuroprotective effect, the antiapoptotic effects of GE were determined by flow cytometry using PI/Annexin V staining method and caspase-3 activity assay. Our PI/Annexin V data demonstrated that GE could strongly attenuate not only the early stage but also the late stage of apoptosis/necrosis induced by Aβ. Besides, we also found that GE could suppress Aβ-induced caspase-3 activity, which provided further evidence in antiapoptosis.
Although the exact underlying mechanism leading to Aβ-induced apoptosis was not well understood, oxidative stress caused by the Aβ plaque was widely believed to seriously impair various cellular function and play an important role in apoptosis [65, 66]. Therefore, reducing reactive oxygen species (ROS) production was a promising approach to inhibit Aβ-induced apoptosis. It has been previously reported that the nonpolar fractions of GE and its active constituents could inhibit ROS generation [32, 67]. In this study, we found that the aqueous extract of GE also possessed strong antioxidative action, which decreased the H2DCF-DA-labeled ROS accumulation in PC12 cells. Antioxidative action can be mediated by 2 mechanisms: activation of antioxidative enzymes and direct free radical scavenging [68]. Antioxidative enzymes, including superoxide dismutase and catalase, convert superoxides, a strong ROS, to hydrogen peroxide and then to water. Glutathione peroxidase catalyzes the reaction of glutathione and hydrogen peroxide, which is a crucial endogenous antioxidative mechanism, to water [69]. In the present study, although Aβ did not affect the activity of CAT, the impairment of the upstream SOD would cause the accumulation of superoxides. On the other hand, the upregulation of GPx by Aβ was possibly a response to the increased ROS and facilitated the action of glutathione. Due to the fact that CAT and GPx could not breakdown superoxides, the accumulation of superoxides may be the explanation for the observed oxidative stress after the Aβ treatment. Our results also demonstrated that GE up-regulated the activity of SOD, CAT, and GPx during Aβ-insult. The activity of SOD was retained, which resume the breakdown of superoxides to hydrogen peroxide. The up-regulation of CAT and GPx can promote the clearance of ROS, and that partially explained the antioxidative action for GE. Other studies also demonstrated that both nonpolar and polar fractions of GE have hydroxyl radical scavenging activity and reduce lipid peroxidation [67, 70, 71]. Its active constituents, including vanillyl alcohol, vanillin, hydroxybenzyl alcohol, and hydroxybenzaldehyde, were found to be potent antioxidants [32, 72]. These compounds can be found in aqueous extract of GE [73, 74].
As aqueous extract of GE was widely and traditionally used in Chinese medicine as a supplement in diet and an herbal medicine [75], and the further development of GE as novel non-toxic preventive/treatment interventions for life-threatening neurodegenerative diseases, such as AD, is possible. In order to confirm our current findings, further investigation of the neuroprotective effect of GE to mammalian AD model is necessary. Since the traditional way of consuming Chinese herbs is to be taken orally, the gastrointestinal metabolic ingredients of GE are the final effective elements. However, very limited information were found regarding the pharmacokinetic data of GE aqueous extract, except that gastrodin was known to be metabolized to p-hydroxybenzyl alcohol [76], and both gastrodin and p-hydroxybenzyl alcohol possess significant free radical scavenging and memory consolidation effects [77, 78]. Hence, further pharmacokinetic studies are required to understand the post metabolism ingredients of GE. Besides, further investigation is needed to determine the clinical efficacy and safety of GE in human subjects because the presence of blood-brain barrier (BBB) may block those beneficial active ingredients from the brain. Although BBB exists in Drosophila and serves the function of blocking the passage of ions and small molecules [79, 80], the Drosophila's BBB is morphologically different from the mammalian one [81, 82]. Nevertheless, some previous studies demonstrated that intravenous administration of gastrodin and hydroxybenzyl alcohol were able to pass through BBB in rats [83, 84]. However, the pharmacokinetics of GE in the human brain is yet to be investigated.
5. Conclusions
In conclusion, the present study demonstrated the novel use of aqueous extract of GE against Aβ-induced neurodegeneration in Drosophila. Its effect is mediated through the increasing activity of antioxidative enzymes and reducing oxidative stress in cells, together with the inhibition of caspase-3, leading to the attenuation of apoptosis. Based on these findings, we suggest developing GE aqueous extract as a potential therapeutic intervention for neurodegenerative diseases, such as Alzheimer's disease.
Conflict of Interests
None of the authors has any conflict of interests regarding this study.
Acknowledgment
This project was supported by Ming Lai Foundation.
References
- 1.Schnabel J. Amyloid: little proteins, big clues. Nature. 2011;475(7355):S12–S14. doi: 10.1038/475S12a. [DOI] [PubMed] [Google Scholar]
- 2.Mattson MP. Pathways towards and away from Alzheimer’s disease. Nature. 2004;430(7000):631–639. doi: 10.1038/nature02621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hardy J, Selkoe DJ. The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science. 2002;297(5580):353–356. doi: 10.1126/science.1072994. [DOI] [PubMed] [Google Scholar]
- 4.Alzheimer's Disease International. World Alzheimer Report. London, UK: Alzheimer's Disease International; [Google Scholar]
- 5.Gravitz L. Drugs: a tangled web of targets. Nature. 2011;475(7355):S9–S11. doi: 10.1038/475S9a. [DOI] [PubMed] [Google Scholar]
- 6.Francis PT, Palmer AM, Snape M, Wilcock GK. The cholinergic hypothesis of Alzheimer’s disease: a review of progress. Journal of Neurology Neurosurgery and Psychiatry. 1999;66(2):137–147. doi: 10.1136/jnnp.66.2.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fu LM, Li JT. A systematic review of single chinese herbs for Alzheimer's disease treatment. Evidence-Based Complementary and Alternative Medicine. 2011;2011:8 pages. doi: 10.1093/ecam/nep136.640284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Howes MJR, Houghton PJ. Ethnobotanical treatment strategies against alzheimer’s disease. Current Alzheimer Research. 2012;9(1):67–85. doi: 10.2174/156720512799015046. [DOI] [PubMed] [Google Scholar]
- 9.Greenspan RJ. Fly Pushing: The Theory and Practice of Drosophila Genetics. 2nd edition. Cold Spring Harbor, NY, USA: Cold Spring Harbor Laboratory Press; 2004. [Google Scholar]
- 10.Finelli A, Kelkar A, Song HJ, Yang H, Konsolaki M. A model for studying Alzheimer’s Aβ42-induced toxicity in Drosophila melanogaster. Molecular and Cellular Neuroscience. 2004;26(3):365–375. doi: 10.1016/j.mcn.2004.03.001. [DOI] [PubMed] [Google Scholar]
- 11.Moloney A, Sattelle DB, Lomas DA, Crowther DC. Alzheimer’s disease: insights from Drosophila melanogaster models. Trends in Biochemical Sciences. 2010;35(4):228–235. doi: 10.1016/j.tibs.2009.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Iijima K, Chiang HC, Hearn SA, et al. Aβ42 mutants with different aggregation profiles induce distinct pathologies in Drosophila. PLoS ONE. 2008;3(2) doi: 10.1371/journal.pone.0001703.e1703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Paris D, Ganey NJ, Laporte V, et al. Reduction of β-amyloid pathology by celastrol in a transgenic mouse model of Alzheimer’s disease. Journal of Neuroinflammation. 2010;7(article 17) doi: 10.1186/1742-2094-7-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.May BH, Lit M, Xue CC, et al. Herbal medicine for dementia: a systematic review. Phytotherapy Research. 2009;23(4):447–459. doi: 10.1002/ptr.2656. [DOI] [PubMed] [Google Scholar]
- 15.Wang N, Chen X, Geng D, Huang H, Zhou H. Ginkgo biloba leaf extract improves the cognitive abilities of rats with D-galactose induced dementia. Journal of Biomedical Research. 2013;27(1):29–36. doi: 10.7555/JBR.27.20120047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shi C, Zhao L, Zhu B, et al. Protective effects of Ginkgo biloba extract (EGb761) and its constituents quercetin and ginkgolide B against β-amyloid peptide-induced toxicity in SH-SY5Y cells. Chemico-Biological Interactions. 2009;181(1):115–123. doi: 10.1016/j.cbi.2009.05.010. [DOI] [PubMed] [Google Scholar]
- 17.Spinnewyn B, Blavet N, Clostre F. Effects of Ginkgo biloba extract on a cerebral ischaemia model in gerbils. Presse Medicale. 1986;15(31):1511–1515. [PubMed] [Google Scholar]
- 18.Xian YF, Lin ZX, Zhao M, Mao QQ, Ip SP, Che CT. Uncaria rhynchophylla ameliorates cognitive deficits induced by D-galactose in mice. Planta Medica. 2011;77(18):1977–1983. doi: 10.1055/s-0031-1280125. [DOI] [PubMed] [Google Scholar]
- 19.Xian YF, Lin ZX, Mao QQ, et al. Bioassay-guided isolation of neuroprotective compounds from Uncaria rhynchophylla against beta-amyloid-induced neurotoxicity. Evidence-Based Complementary and Alternative Medicine. 2012;2012:8 pages. doi: 10.1155/2012/802625.802625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Shim JS, Kim HG, Ju MS, Choi JG, Jeong SY, Oh MS. Effects of the hook of Uncaria rhynchophylla on neurotoxicity in the 6-hydroxydopamine model of Parkinson’s disease. Journal of Ethnopharmacology. 2009;126(2):361–365. doi: 10.1016/j.jep.2009.08.023. [DOI] [PubMed] [Google Scholar]
- 21.Hsieh CL, Liu CH, Lin YW, Tang NY, Liu HJ. Neuroprotective effect of Uncaria rhynchophylla in Kainic acid-induced epileptic seizures by modulating hippocampal mossy fiber sprouting, neuron survival, astrocyte proliferation, and S100b expression. Evidence-based Complementary and Alternative Medicine. 2012;2012 doi: 10.1155/2012/194790.194790 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lin Z, Gu J, Xiu J, Mi T, Dong J, Tiwari JK. Traditional chinese medicine for senile dementia. Evidence-Based Complementary and Alternative Medicine. 2012;2012:13 pages. doi: 10.1155/2012/692621.692621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhu KY, Mao QQ, Ip SP, et al. A standardized chinese herbal decoction, kai-xin-san, restores decreased levels of neurotransmitters and neurotrophic factors in the brain of chronic stress-induced depressive rats. Evidence-Based Complementary and Alternative Medicine. 2012;2012:13 pages. doi: 10.1155/2012/149256.149256 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Naito R, Tohda C. Characterization of anti-neurodegenerative effects of Polygala tenuifolia in Aβ(25-35)-treated cortical neurons. Biological and Pharmaceutical Bulletin. 2006;29(9):1892–1896. doi: 10.1248/bpb.29.1892. [DOI] [PubMed] [Google Scholar]
- 25.Rui C, Yuxiang L, Yinju H, et al. Protective effects of Lycium barbarum polysaccharide on neonatal rat primary cultured hippocampal neurons injured by oxygen-glucose deprivation and reperfusion. Journal of Molecular Histology. 2012;43(5):535–542. doi: 10.1007/s10735-012-9420-4. [DOI] [PubMed] [Google Scholar]
- 26.An H, Kim IS, Koppula S, et al. Protective effects of Gastrodia elata Blume on MPP+-induced cytotoxicity in human dopaminergic SH-SY5Y cells. Journal of Ethnopharmacology. 2010;130(2):290–298. doi: 10.1016/j.jep.2010.05.006. [DOI] [PubMed] [Google Scholar]
- 27.Huang ZB, Wu Z, Chen FK, Zou LB. The protective effects of phenolic constituents from Gastrodia elata on the cytotoxicity induced by KCl and glutamate. Archives of Pharmacal Research. 2006;29(11):963–968. doi: 10.1007/BF02969279. [DOI] [PubMed] [Google Scholar]
- 28.Kim HJ, Moon KD, Oh SY, Kim SP, Lee SR. Ether fraction of methanol extracts of Gastrodia elata, a traditional medicinal herb, protects against kainic acid-induced neuronal damage in the mouse hippocampus. Neuroscience Letters. 2001;314(1-2):65–68. doi: 10.1016/s0304-3940(01)02296-0. [DOI] [PubMed] [Google Scholar]
- 29.Shuchang H, Qiao N, Piye N, et al. Protective effects of gastrodia elata on aluminium-chloride-induced learning impairments and alterations of amino acid neurotransmitter release in adult rats. Restorative Neurology and Neuroscience. 2008;26(6):467–473. [PMC free article] [PubMed] [Google Scholar]
- 30.Xu X, Lu Y, Bie X. Protective effects of gastrodin on hypoxia-induced toxicity in primary cultures of rat cortical neurons. Planta Medica. 2007;73(7):650–654. doi: 10.1055/s-2007-981523. [DOI] [PubMed] [Google Scholar]
- 31.Yu SS, Zhao J, Lei SP, Lin XM, Wang LL, Zhao Y. 4-hydroxybenzyl alcohol ameliorates cerebral injury in rats by antioxidant action. Neurochemical Research. 2011;36(2):339–346. doi: 10.1007/s11064-010-0335-8. [DOI] [PubMed] [Google Scholar]
- 32.Kim HJ, Hwang IK, Won MH. Vanillin, 4-hydroxybenzyl aldehyde and 4-hydroxybenzyl alcohol prevent hippocampal CA1 cell death following global ischemia. Brain Research. 2007;1181(1):130–141. doi: 10.1016/j.brainres.2007.08.066. [DOI] [PubMed] [Google Scholar]
- 33.Mishra M, Huang J, Lee YY, et al. Gastrodia elata modulates amyloid precursor protein cleavage and cognitive functions in mice. BioScience Trends. 2011;5(3):129–138. doi: 10.5582/bst.2011.v5.3.129. [DOI] [PubMed] [Google Scholar]
- 34.Mattson MP. Cellular actions of β-amyloid precursor protein and its soluble and fibrillogenic derivatives. Physiological Reviews. 1997;77(4):1081–1132. doi: 10.1152/physrev.1997.77.4.1081. [DOI] [PubMed] [Google Scholar]
- 35.Vassar R, Bennett BD, Babu-Khan S, et al. β-Secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science. 1999;286(5440):735–741. doi: 10.1126/science.286.5440.735. [DOI] [PubMed] [Google Scholar]
- 36.Colciaghi F, Borroni B, Zimmermann M, et al. Amyloid precursor protein metabolism is regulated toward alpha-secretase pathway by Ginkgo biloba extracts. Neurobiology of Disease. 2004;16(2):454–460. doi: 10.1016/j.nbd.2004.03.011. [DOI] [PubMed] [Google Scholar]
- 37.Nistor M, Don M, Parekh M, et al. Alpha- and beta-secretase activity as a function of age and beta-amyloid in Down syndrome and normal brain. Neurobiology of Aging. 2007;28(10):1493–1506. doi: 10.1016/j.neurobiolaging.2006.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chinese Pharmacopoeia Commission. Pharmacopoeia of the People's Republic of China. Chemical Industry Press; 2010. [Google Scholar]
- 39.Warrick JM, Chan HYE, Gray-Board GL, Chai Y, Paulson HL, Bonini NM. Suppression of polyglutamine-mediated neurodegeneration in Drosophila by the molecular chaperone HSP70. Nature Genetics. 1999;23(4):425–428. doi: 10.1038/70532. [DOI] [PubMed] [Google Scholar]
- 40.Lee FKM, Wong AKY, Lee YW, Wan OW, Edwin Chan HY, Chung KKK. The role of ubiquitin linkages on α-synuclein induced-toxicity in a Drosophila model of Parkinson’s disease. Journal of Neurochemistry. 2009;110(1):208–219. doi: 10.1111/j.1471-4159.2009.06124.x. [DOI] [PubMed] [Google Scholar]
- 41.Wong SLA, Wing MC, Chan HYE. Sodium dodecyl sulfate-insoluble oligomers are involved in polyglutamine degeneration. FASEB Journal. 2008;22(9):3348–3357. doi: 10.1096/fj.07-103887. [DOI] [PubMed] [Google Scholar]
- 42.Xiao C, Li J, Dong X, et al. Anti-oxidative and TNF-α suppressive activities of puerarin derivative (4AC) in RAW264.7 cells and collagen-induced arthritic rats. European Journal of Pharmacology. 2011;666(1–3):242–250. doi: 10.1016/j.ejphar.2011.05.061. [DOI] [PubMed] [Google Scholar]
- 43.Choi RC, Jiang Z, Xie HQ, et al. Anti-oxidative effects of the biennial flower of Panax notoginseng against H2O2-induced cytotoxicity in cultured PC12 cells. Chinese Medicine. 2010;5(article 38) doi: 10.1186/1749-8546-5-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Boldyrev A, Bulygina E, Leinsoo T, Petrushanko I, Tsubone S, Abe H. Protection of neuronal cells against reactive oxygen species by carnosine and related compounds. Comparative Biochemistry and Physiology B. 2004;137(1):81–88. doi: 10.1016/j.cbpc.2003.10.008. [DOI] [PubMed] [Google Scholar]
- 45.Duffy JB. GAL4 system in Drosophila: a fly geneticist’s Swiss army knife. Genesis. 2002;34(1-2):1–15. doi: 10.1002/gene.10150. [DOI] [PubMed] [Google Scholar]
- 46.Jackson GR, Salecker I, Dong X, et al. Polyglutamine-expanded human huntingtin transgenes induce degeneration of Drosophila photoreceptor neurons. Neuron. 1998;21(3):633–642. doi: 10.1016/s0896-6273(00)80573-5. [DOI] [PubMed] [Google Scholar]
- 47.Hong YK, Park SH, Lee S, et al. Neuroprotective effect of SuHeXiang Wan in Drosophila models of Alzheimer’s disease. Journal of Ethnopharmacology. 2011;134(3):1028–1032. doi: 10.1016/j.jep.2011.02.012. [DOI] [PubMed] [Google Scholar]
- 48.Ling D, Song HJ, Garza D, Neufeld TP, Salvaterra PM. Abeta42-induced neurodegeneration via an age-dependent autophagic-lysosomal injury in Drosophila. PLoS ONE. 2009;4(1) doi: 10.1371/journal.pone.0004201.e4201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Luheshi LM, Tartaglia GG, Brorsson AC, et al. Systematic in vivo analysis of the intrinsic determinants of amyloid beta pathogenicity. PLoS Biology. 2007;5(11):p. e290. doi: 10.1371/journal.pbio.0050290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Van De Goor J, Kelly RB. Association of Drosophila cysteine string proteins with membranes. FEBS Letters. 1996;380(3):251–256. doi: 10.1016/0014-5793(96)00026-9. [DOI] [PubMed] [Google Scholar]
- 51.Apostol BL, Kazantsev A, Raffioni S, et al. A cell-based assay for aggregation inhibitors as therapeutics of polyglutamine-repeat disease and validation in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(10):5950–5955. doi: 10.1073/pnas.2628045100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wang J, Pfleger CM, Friedman L, et al. Potential application of grape derived polyphenols in huntington’s disease. Translational Neuroscience. 2010;1(2):95–100. doi: 10.2478/v10134-010-0022-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Greene LA, Tischler AS. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proceedings of the National Academy of Sciences of the United States of America. 1976;73(7):2424–2428. doi: 10.1073/pnas.73.7.2424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Xu SL, Choi RC, Zhu KY, et al. Isorhamnetin, a flavonol aglycone from Ginkgo biloba L., induces neuronal differentiation of cultured PC12 cells: potentiating the effect of nerve growth factor. Evidence-Based Complementary and Alternative Medicine. 2012;2012:11 pages. doi: 10.1155/2012/278273.278273 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chuang MC, Lai HY, Annie Ho JA, Chen YY. Multifunctional microelectrode array (mMEA) chip for neural-electrical and neural-chemical interfaces: characterization of comb interdigitated electrode towards dopamine detection. Biosensors and Bioelectronics. 2013;41:602–607. doi: 10.1016/j.bios.2012.09.030. [DOI] [PubMed] [Google Scholar]
- 56.Wang AM, Miyata Y, Klinedinst S, et al. Activation of Hsp70 reduces neurotoxicity by promoting polyglutamine protein degradation. Nature Chemical Biology. 2013;9(2):112–118. doi: 10.1038/nchembio.1140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lee SB, Kim CK, Lee KH, Ahn JY. S-nitrosylation of B23/nucleophosmin by GAPDH protects cells from the SIAH1-GAPDH death cascade. Journal of Cell Biology. 2012;199(1):65–76. doi: 10.1083/jcb.201205015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jayaprakasam B, Padmanabhan K, Nair MG. Withanamides in Withania somnifera fruit protect PC-12 cells from β-amyloid responsible for Alzheimer’s disease. Phytotherapy Research. 2010;24(6):859–863. doi: 10.1002/ptr.3033. [DOI] [PubMed] [Google Scholar]
- 59.Kim HJ, Moon KD, Lee DS, Lee SH. Ethyl ether fraction of Gastrodia elata Blume protects amyloid β peptide-induced cell death. Journal of Ethnopharmacology. 2003;84(1):95–98. doi: 10.1016/s0378-8741(02)00290-8. [DOI] [PubMed] [Google Scholar]
- 60.Shimohama S. Apoptosis in Alzheimer’s disease: an update. Apoptosis. 2000;5(1):9–16. doi: 10.1023/a:1009625323388. [DOI] [PubMed] [Google Scholar]
- 61.Yuan J, Yankner BA. Apoptosis in the nervous system. Nature. 2000;407(6805):802–809. doi: 10.1038/35037739. [DOI] [PubMed] [Google Scholar]
- 62.Ankarcrona M, Winblad B. Biomarkers for apoptosis in Alzheimer’s disease. International Journal of Geriatric Psychiatry. 2005;20(2):101–105. doi: 10.1002/gps.1260. [DOI] [PubMed] [Google Scholar]
- 63.LaFerla FM, Green KN, Oddo S. Intracellular amyloid-β in Alzheimer’s disease. Nature Reviews Neuroscience. 2007;8(7):499–509. doi: 10.1038/nrn2168. [DOI] [PubMed] [Google Scholar]
- 64.Coyle JT, Price DL, DeLong MR. Alzheimer’s disease: a disorder of cortical cholinergic innervation. Science. 1983;219(4589):1184–1190. doi: 10.1126/science.6338589. [DOI] [PubMed] [Google Scholar]
- 65.Mattson MP, Pedersen WA, Duan W, Culmsee C, Camandola S. Cellular and molecular mechanisms underlying perturbed energy metabolism and neuronal degeneration in Alzheimer’s and Parkinson’s diseases. Annals of the New York Academy of Sciences. 1999;893:154–175. doi: 10.1111/j.1749-6632.1999.tb07824.x. [DOI] [PubMed] [Google Scholar]
- 66.Annunziato L, Amoroso S, Pannaccione A, et al. Apoptosis induced in neuronal cells by oxidative stress: role played by caspases and intracellular calcium ions. Toxicology Letters. 2003;139(2-3):125–133. doi: 10.1016/s0378-4274(02)00427-7. [DOI] [PubMed] [Google Scholar]
- 67.Jung TY, Suh SI, Lee H, et al. Protective effects of several components of Gastrodia elata on lipid peroxidation in gerbil brain homogenates. Phytotherapy Research. 2007;21(10):960–964. doi: 10.1002/ptr.2193. [DOI] [PubMed] [Google Scholar]
- 68.Butterfield DA, Lauderback CM. Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: potential causes and consequences involving amyloid β-peptide-associated free radical oxidative stress. Free Radical Biology and Medicine. 2002;32(11):1050–1060. doi: 10.1016/s0891-5849(02)00794-3. [DOI] [PubMed] [Google Scholar]
- 69.Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000;408(6809):239–247. doi: 10.1038/35041687. [DOI] [PubMed] [Google Scholar]
- 70.Shin EJ, Bach JH, Nguyen TTL, et al. Gastrodia elata Bl attenuates methamphetamine-induced Dopaminergic toxicity via inhibiting oxidative burdens. Current Neuropharmacology. 2011;9(1):118–121. doi: 10.2174/157015911795016967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ji YL, Young WJ, Hyo SK, Moon H, Sang SS, Chang JK. Anti-inflammatory action of phenolic compounds from Gastrodia elata root. Archives of Pharmacal Research. 2006;29(10):849–858. doi: 10.1007/BF02973905. [DOI] [PubMed] [Google Scholar]
- 72.Kim IS, Choi DK, Jung HJ. Neuroprotective effects of vanillyl alcohol in gastrodia elata blume through suppression of oxidative stress and anti-apoptotic activity in toxin-induced dopaminergic MN9D cells. Molecules. 2011;16(7):5349–5361. doi: 10.3390/molecules16075349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Teo CC, Tan SN, Yong JWH, Hew CS, Ong ES. Evaluation of the extraction efficiency of thermally labile bioactive compounds in Gastrodia elata Blume by pressurized hot water extraction and microwave-assisted extraction. Journal of Chromatography A. 2008;1182(1):34–40. doi: 10.1016/j.chroma.2008.01.011. [DOI] [PubMed] [Google Scholar]
- 74.Wang Y, Lin S, Chen M, et al. Chemical constituents from aqueous extract of Gastrodia elata. China Journal of Chinese Materia Medica. 2012;37(12):1775–1781. [PubMed] [Google Scholar]
- 75.Bulpitt CJ, Li Y, Bulpitt PF, Wang J. The use of orchids in Chinese medicine. Journal of the Royal Society of Medicine. 2007;100(12):558–563. doi: 10.1258/jrsm.100.12.558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wu CR, Hsieh MT, Huang SC, Peng WH, Chang YS, Chen CF. Effects of Gastrodia elata and its active constituents on scopolamine-induced amnesia in rats. Planta Medica. 1996;62(4):317–321. doi: 10.1055/s-2006-957892. [DOI] [PubMed] [Google Scholar]
- 77.Hsieh MT, Wu CR, Chen CF. Gastrodin and p-hydroxybenzyl alcohol facilitate memory consolidation and retrieval, but not acquisition, on the passive avoidance task in rats. Journal of Ethnopharmacology. 1997;56(1):45–54. doi: 10.1016/s0378-8741(96)01501-2. [DOI] [PubMed] [Google Scholar]
- 78.Wu CR, Hsieh MT, Liao J. p-Hydroxybenzyl alcohol attenuates learning deficits in the inhibitory avoidance task: involvement of serotonergic and dopaminergic systems. Chinese Journal of Physiology. 1996;39(4):265–273. [PubMed] [Google Scholar]
- 79.Juang JL, Carlson SD. Analog of vertebrate anionic sites in blood-brain interface of larval Drosophila. Cell and Tissue Research. 1994;277(1):87–95. doi: 10.1007/BF00303084. [DOI] [PubMed] [Google Scholar]
- 80.Stork T, Engelen D, Krudewig A, Silies M, Bainton RJ, Klämbt C. Organization and function of the blood-brain barrier in Drosophila. Journal of Neuroscience. 2008;28(3):587–597. doi: 10.1523/JNEUROSCI.4367-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Freeman MR, Doherty J. Glial cell biology in Drosophila and vertebrates. Trends in Neurosciences. 2006;29(2):82–90. doi: 10.1016/j.tins.2005.12.002. [DOI] [PubMed] [Google Scholar]
- 82.Daneman R, Barres BA. The blood-brain barrier—lessons from moody flies. Cell. 2005;123(1):9–12. doi: 10.1016/j.cell.2005.09.017. [DOI] [PubMed] [Google Scholar]
- 83.Wang Q, Chen G, Zeng S. Distribution and metabolism of gastrodin in rat brain. Journal of Pharmaceutical and Biomedical Analysis. 2008;46(2):399–404. doi: 10.1016/j.jpba.2007.10.017. [DOI] [PubMed] [Google Scholar]
- 84.Wang Q, Chen G, Zeng S. Pharmacokinetics of Gastrodin in rat plasma and CSF after i.n. and i.v. International Journal of Pharmaceutics. 2007;341(1-2):20–25. doi: 10.1016/j.ijpharm.2007.03.041. [DOI] [PubMed] [Google Scholar]