

Abstract
Lymphatic filariasis affects nearly 120 million people worldwide and mass preventive chemotherapy is currently used as a strategy to control this infection. This has substantially reduced the incidence of the infection in several parts of the world. However, a prophylactic vaccine would be more effective in preventing future infections and will supplement the mass chemotherapy efforts. Unfortunately, there is no licensed vaccine available currently to prevent this infection. Molecules expressed on the surface of the parasite are potential candidates for vaccine development as they are exposed to the host immune system. In this study we show that the large extracellular loop of tetraspanin (TSP LEL), a protein expressed on the cuticle of Brugia malayi and Wuchereria bancrofti is a potential vaccine candidate. Our results showed that BmTSP LEL is expressed on the surface of B. malayi infective third stage larvae (L3) and sera from human subjects who are putatively immune to lymphatic filariasis carry high titer of IgG1 and IgG3 antibodies against BmTSP LEL and WbTSP LEL. We also showed that these antibodies in the sera of human subjects can participate in the killing of B. malayi L3 in an antibody dependent cell-mediated cytotoxicity mechanism. Vaccination trials in mice showed that close to 64% protection were achieved against challenge infections with B. malayi L3. Immunized animals showed high titer of anti-WbTSP LEL IgG1, IgG2a and IgG2b antibodies in the sera and IFN-γ secreting cells in the spleen. Onchocerca volvulus another filarial parasite also expresses TSP LEL. Cross-reactivity studies showed that IgG1 antibody in the sera of endemic normal subjects, recognize OvTSP LEL. Similarly, anti-OvTSP LEL antibodies in the sera of subjects who are immune to O. volvulus were also shown to cross-react with rWbTSP LEL and rBmTSP LEL. These findings thus suggested that rTSP LEL can be developed as a potential vaccine candidate against multiple filarial infections.
Introduction
Lymphatic filariasis (LF) affects 120 million people in 72 countries and more than 1.2 billion people are at the risk of acquiring the infections [1]. The disease is transmitted by mosquitoes carrying any of three parasitic nematodes: Wuchereria bancrofti, Brugia malayi, and Brugia timori. In the tropical and sub tropical regions of the world filarial infections are a major public health and socio-economic problem [2,3]. Mass drug administration is currently used as an approach to control this infection in endemic areas [4]. However, an effective prophylactic vaccine against lymphatic filariasis coupled with effective vector control measures can supplement the mass drug administration approach and speed up the effort to eliminate this infection in the endemic regions. Several potential vaccine candidates have been identified and evaluated from different laboratories [5-9], yet there is no effective vaccine available against this disease. This may be largely due to the complex life cycle stages of the parasites and the varied immune responses developed by the host to each antigen.
Lymphatic filarial parasites are notorious for evading the host immune responses by molecular mimicry and active modulation of the host immune responses [10]. Tetraspanin (TSP) family of proteins is one among the parasite antigens that plays an important role in host immunomodulation. TSPs are membrane-spanning proteins containing four transmembrane domains, three short intracellular domains, and two extracellular loops, EC1 and EC2 [11]. TSPs are abundantly expressed on the surface or body wall of several helminth parasites including S. mansoni [12], S. japonicum [13] and in the nematode C. elegans [14]. TSP is believed to play important roles in signal transduction, cell proliferation, adhesion, migration, fusion and host-parasite interactions [15]. Among the various TSPs reported, TSP-2 identified from Schistosoma mansoni (SmTSP-2) is shown to be involved in the formation of tegument [13] which is critically important for the parasite’s survival in the host [16]. Since TSPs are expressed on the surface, they are easy targets for immune attack by host antibodies and cells [17]. Especially, the extracellular loops of TSP are more amenable to the host immune system. Given its critical role in the formation of tegument and survival of the parasite in the host, TSP has been a target for developing vaccine against S. mansoni [18-20]. Especially, the large extracellular loop (LEL) of SmTSP-2 was shown to be highly effective as a vaccine immunogen conferring high levels of protection in mice against challenge infections [12]. Previously we reported the presence of SmTSP-2 homologue in B. malayi parasites [21]. Since TSP-LEL appears to be a good vaccine candidate, in the present study we cloned tsp lel from lymphatic filarial parasites (W. bancrofti and B. malayi). We then expressed the recombinant B. malayi TSP LEL (rBmTSP LEL) and W. bancrofti TSP LEL (rWbTSP LEL) proteins and evaluated its vaccine potential in human and in mice.
Developing a single global vaccine that is effective against multiple infections is an attractive strategy for endemic regions where mixed infections are common. Administering vaccines separately for diseases require an increased number of visits, labor, side effects, and costs not to mention the various regulatory approvals needed to develop separate new vaccines. Generation of broadly protective “universal” vaccines restricted to species or groups of closely related pathogens or even cross-family or -kingdom vaccines might be useful to overcome some of the barriers in vaccine development [22]. Very few studies have attempted to identify these cross-species protective antigens. Among the eight filarial species that parasitize humans, lymphatic filariasis caused by W. bancrofti and B. malayi and onchocerciasis caused by Onchocerca volvulus are more common and occur as co-infection in several endemic regions [23,24]. These three parasites are also the major targets for vaccine development by a number of laboratories [23,25-27]. In this report we show that homologue of TSP-2 is expressed in O. volvulus and show significant sequence similarity to BmTSP-2 and WbTSP-2. Therefore, in this study we have also attempted to evaluate the cross-protection potential of rTSP LEL from LF parasites against onchocerciasis.
Materials and Methods
Ethics Statement
Use of human subjects in this study was approved by the Institutional Review Board (IRB) of the University of Illinois, College of Medicine at Rockford; the New York Blood Center’s IRB and by an NIH accredited Institutional Review Board of the Medical Research Council Kumba, Cameroon and the IRB committee of Mahatma Gandhi Institute of Medical Sciences, Sevagram, Maharashtra, India. Informed written consent in native language was obtained from all the subjects before collecting the samples. Humane use of animals was performed in this study according to the guidelines for the care and use of laboratory animals and with the rules formulated under the Animal Welfare Act by the U.S. Department of Agriculture. The protocol was approved by The IACUC Committee of the University of Illinois, College of Medicine at Rockford and performed at a facility accredited by AAALAC and USDA.
Parasites
Brugia malayi infective third stage larvae (L3) were obtained from the NIAID/NIH Filariasis Research Reagent Resource Center (FR3) at the University of Georgia, Athens, GA.
Human sera Samples
Blood samples were collected after taking informed consent from clinically diagnosed filarial patients and from healthy adult individuals residing in Sevagram and surrounding villages in Maharashtra State, which are non-coastal endemic areas for nocturnally periodic W. bancrofti infection. Samples were also collected from volunteers who live in areas that are non endemic for filariasis. Parasitological examination of all individuals was done by detection of microfilariae in night blood smears. The presence of mf was further confirmed by membrane (Millipore-5m filters) filtration of 1.0 ml of heparinized venous blood [28]. The presence of circulating antigen was detected using a WbSXP based enzyme-linked immunosorbent assay (ELISA) [29]. About 10 ml of blood samples (5 ml in heparinized tubes) were collected from the following clinical groups of subjects (a) Endemic normal (EN) subjects, these are individuals who are asymptomatic and non-microfilaraemic residing in Sevagram (b) Asymptomtic microfilaraemic subjects (MF) were classified based on the presence of circulating mf in blood. (c) Chronic Pathology (CP) patients include those subjects who exhibit lymph edema and other chronic clinical symptoms of filariasis. These CP patients did not carry circulating mf and (d) Non-endemic normal (NEN) subjects are healthy individuals who live in a non-endemic country and have no circulating parasites or antibodies in their sera and show no evidence of any clinical lymphatic filarial disease. Sera were separated from these blood samples and were stored at - 80°C until use. PBMC proliferation was performed using freshly collected blood samples from EN, MF and CP individuals.
Sera samples from onchocerciasis immune (PI) and onchocerciasis infected individuals (INF) were collected from residents of 5 villages around Kumba, Cameroon, a hyperendemic area after obtaining the informed consent by signing or thumb printing a consent form after reading it or after the content of the form was read and/or explained to them. These villages were Marumba I, Marumba II, Boa Bakundu, Bombanda, and Bombele. All participants were born or had resided for more than 10 years in the villages. The standard skin snip test for detection of microfilariae (mf) was performed on each subject and clinical symptoms of onchocerciasis were recorded. The averages of the mf counts of four skin snips taken from each individual were used in estimating the individual skin mf densities. None of the subjects had received ivermectin treatment prior to the collection of blood. Serum samples of 20 putatively immune (PI) subjects of onchocerciasis were studied. These individuals had no signs or history of onchocerciasis and were parasitologically negative during at least a two-year follow-up survey employing the standard skin snip test and a polymerase chain reaction (PCR) assay. Age, sex matched infected individuals were used for comparison as described before [30,31] .
Cloning of tsp from three filarial parasites
Transcripts encoding TSP LEL proteins were amplified from W. bancrofti, B. malayi and O. volvulus L3 cDNA libraries. Primers were designed from the genebank sequence (Accession number EF397425.1) of B. malayi tetraspanin LEL domain. The same set of primers were used to amplify tsp lel from the cDNA libraries of all the three parasites; Forward primer sequence with BamHI restriction sites 5′-CGGGATCCCGGCAAGGATCAATTTAAAA-3′ and Reverse primer sequence with EcoRI 5′-CGGAATTCTCAATCTTTTTGAGATGAAT-3′. PCR was performed with denaturation at 95°C, annealing at 50°C, and extension at 75°C. Five different PCR clones from each parasite species were sent out for sequencing at the DNA sequencing facility at UIC, Chicago. The results showed no difference in the sequences between the PCR clones. A multiple sequence alignment of helminth TSP LEL was then performed using ClustalW.
Expression and purification of rTSP LEL domain
tsp lel amplified from L3 cDNA libraries (W. bancrofti, B. malayi and O. volvulus) were cloned into pRSETA expression vector and transformed into BL21(DE3) pLysS E. coli expression host. E. coli cultures with pRSETA tsp plasmid was inoculated in 500ml of LB broth and incubated for 3 hours until the log phase of bacterial growth was reached (absorbance OD 0.6 at 600nm). 1 mM of isopropyl-beta-D-thiogalactopyranoside [IPTG] was added to the culture to induce the proteins expressions and incubated for an additional 3 hours. Recombinant WbTSP LEL, BmTSP LEL and OvTSP LEL expressed as His-tag proteins were purified using IMAC column. SDS PAGE was run to check the purity of the recombinant proteins. Immunoblot was performed by probing with anti penta-His antibodies (Qiagen, Valencia, CA). Endotoxin was removed from the recombinant proteins using High Capacity Endotoxin removal resin (Thermo Fisher Scientific, Rockford, IL). Levels of endotoxin in the final recombinant protein preparation were confirmed by LAL assay (Gen Script, Piscataway, NJ) and were found to be below 4 EU/ml.
Immunization of mice
Six weeks old male Balb/c mice purchased from Charles River laboratory (Wilmington, MA) were divided into three groups with five mice in each group. Mice were immunized four times subcutaneously (s.c) with 15μg of recombinant tetraspanin proteins (rWbTSP LEL or rBmTSP LEL) in 100μl volume combined with 50μl of Imject alum (Thermo Fisher Scientific) given at 2 weeks interval. Control group of animals received only alum. Blood was collected from the retro-orbital space of each mouse two weeks after the final dose of immunization. This experiment was repeated two times.
Immunofluorescence assay to evaluate the presence of TSP LEL on the surface of B. malayi L3
Presence of TSP LEL on the surface of infective larvae of B. malayi was analyzed by an immunofluorescence whole mount assay [32]. About 5-10 L3 were incubated with 1% BSA, 0.1% Triton X-100 in phosphate buffer saline (PBS) for two hours at 4°C under agitation in the microcentrifuge tube. Pooled mouse anti-BmTSP LEL sera (optimum dilution was found to be 1:25) diluted in 100µl volumes of blocking buffer was added to L3 and incubated at room temperature for 2 h. After washing with PBS, the parasites were incubated at RT for 1 h with anti-mouse IgG conjugated with fluorescein isothiocyanate (FITC, purchased from Thermo Fisher Scientific) at 1:1000 dilutions. After washing with PBS, the parasites were placed on to a glass slide and mounted with fluorophore mounting medium. After placing a cover slip, the edges were sealed and observed under a fluorescence microscope (Olympus, Center valley, PA) using 10X and 60X objective lens. Sera from mice immunized with alum or anti-mouse IgG FITC alone were used as negative controls (data not shown).
Analysis of total anti-TSP LEL IgG antibodies in the sera samples
IgG antibodies against rWbTSP LEL, rBmTSP LEL or rOvTSP LEL were evaluated in the sera of different filarial groups of subjects by indirect ELISA [33]. Briefly, wells of a 96 well microtiter ELISA plates were coated with 100 ng/well of recombinant proteins (rWbTSP LEL, rBmTSP LEL or rOvTSP LEL) in 0.05M carbonate-bicarbonate buffer, pH 9.6. After blocking the wells with 3% BSA in PBS-Tween 20 (PBS-T, 0.05%), 100µl of sera samples (1:100 diluted in PBS-T) from different groups of subjects with filarial infections (MF, CP, EN and NEN) were added to each well. Horse radish peroxidase-labeled Goat anti-human IgG was used as the secondary antibody. The color was developed using OPD substrate and absorbance was read at 450 nm in the ELISA reader (BioRad, Hercules, CA). Antigen-specific isotype of IgG antibodies present in the sera of different filarial groups were evaluated by indirect ELISA [34]. Biotinylated anti-human IgG1 (1:1000), IgG2 (1:15000), IgG3 (1:1000) and IgG4 (1:15000) isotypes (Sigma Aldrich, St. Louis, MO) were used as the secondary antibodies. Levels of anti-TSP LEL IgG and IgG isotype antibodies (Biolegend, San Diego, CA) were similarly measured in the sera of mice immunized with rTSP LEL proteins.
Levels of cross-reactive IgG1 and IgG3 antibodies against WbTSP LEL was measured in the sera of O. volvulus putatively immune (PI) or O. volvulus infected individuals (INF) using an indirect ELISA described by MacDonald et al., [31]. Briefly, wells of a microtiter Immunolon 2 plates (Dynex, Chantilly, VA), were coated with 1 µg/ml rWbTSP LEL diluted in 0.05M carbonate-bicarbonate buffer, (pH 9.6) overnight at 4°C. Non-specific sites were blocked with 3% BSA in PBS-T for 2 h at 37°C. After washing the plates six times with PBS-T, individual serum samples that were pre-cleared with E. coli lysate (400 μg/ml, Promega, Madison, WI) and diluted to 1:100 in binding buffer (1% non-fat milk in PBS-T) were added to each well in duplicate and incubated for 1 h at 37°C. Pre-clarification using E. coli lysate was performed as described previously [30] to remove any cross-reacting E. coli specific antibodies in the sera samples that may interfere in our assay since all our recombinant proteins were prepared in E. coli. Monoclonal antibodies (1:1000 dilution) against different human IgG subclasses (Hybridoma Reagent Laboratory, Kingsville, MD) was used as the secondary antibodies and horseradish peroxidase-conjugated goat anti-mouse IgG (H+L) (KPL, Gaithersburg, MD, 1:8000, incubated for 60 min at room temperature) was used as the detection antibodies. Absorbance (OD450) was read after stopping the reaction with an equal volume of 1M H2SO4 using an Emax ELISA reader (Molecular Devices, Sunnyvale, CA).
We also depleted anti-WbTSP LEL IgG antibodies from the sera of EN subjects by passing the sera through a cobalt IMAC resin column coupled to rWbTSP LEL [35]. Briefly, 1 mg of his-tagged rWbTSP LEL was coupled to 2 ml bed volume of IMAC resin for 2 hrs at 37°C. After washing the resin once with 10 ml of PBS (pH.8), 200 ml of pooled sera was added and incubated overnight at 4°C. After incubation, the resin mixture was centrifuged for 2 min at 750 rpm and the supernatant were collected. Total IgG was removed by incubating sera samples with Protein A gel beads (Thermo Fisher Scientific). Depletion of rWbTSP LEL specific antibodies in the sera samples was confirmed by an indirect ELISA as described above.
Antibody dependent cell mediated cytotoxicity (ADCC) assay using human and mouse sera
In vitro ADCC assay was performed using both human and mice sera as described previously [34]. Briefly, ten L3 of B. malayi were incubated with 2 x 105 peripheral blood mononuclear cells (PBMC) collected from normal healthy subjects, 50 µl of pooled EN sera samples and 50 µl of RPMI 1640 media in a 96 well culture plate (Thermo Fisher Scientific). After 48 h of incubation at 37°C and 5% CO2, the larval viability was determined under a light microscope (400 X). Viable larvae were actively moving, coiled and translucent. Dead larvae were flaccid, transparent, damaged and had clumps of cells attached to them. ADCC was estimated as percent larval death calculated using the following formula: Number of dead larvae ÷ Total number of larvae x 100. ADCC assay was also performed using sera samples from rTSP LEL immunized mice. 2 x 105 peritoneal exudates cells (PEC) from normal mice were used as effector cells in these assays.
Analysis of cross reactivity between lymphatic filarial TSP LELs and Onchocerca TSP LEL
Cross reactivity of anti-WbTSP LEL IgG antibodies against OvTSP LEL was evaluated using an indirect ELISA as described above. The cross reactivity was also evaluated between BmTSP LEL, WbTSP LEL and OvTSP LEL using an immunoblot analysis. Briefly, nitrocellulose membrane blotted with the each of the recombinant proteins was probed with sera collected from mice immunized with rBmTSP LEL or rWbTSP LEL for 1hr at room temperature (RT). After washing with PBST, membranes were incubated for 1hr at room temperature with a secondary goat anti-mouse IgG antibodies conjugated to HRP (1:5000, Thermo Fisher). Color was developed using diamino benzidine (Thermo Fisher Scientific) substrate.
Analysis of vaccine induced protection in mice
Two weeks after the final vaccination we performed the in vivo challenge experiments by surgically implanting 20 B. malayi L3 in a micropore chamber in the peritoneal cavity of each mouse [36,37]. After 48 hours, mice were sacrificed and the chambers were surgically removed and examined under the microscope to determine the larval viability. Percent larval death was determined as described before [36,37].
Antigen specific proliferation assay
Antigen specific proliferation of mouse spleen cells were evaluated as described previously [34]. Briefly, 1 x 105 splenocytes were stimulated with (a) 1 µg/ml of rWbTSP LEL or 1 µg/ml of rBmTSP LEL, (b) 1 µg/ml of a non-specific recombinant protein (recombinant S. mansoni G-binding factor, rSmGBF), (c) 1 µg/ml of ConA or (d) medium alone (unstimulated) in triplicate wells for 72hrs at 37°C. Stimulation index (S.I) was calculated from the unstimulated controls. For cytokine analysis, splenocytes were stimulated with 1 µg/ml of rWbTSP LEL and medium alone (unstimulated) for 72 hrs.
Antigen specific proliferation of PBMC was also determined similarly as described previously [34]. Briefly, PBMC were isolated from heparinized venous blood of study subjects using histopaque 1077 column (Sigma). Isolated PBMC were stimulated with (a) 1 µg/ml of rWbTSP LEL, (b) 1 µg/ml of ConA (positive control), or (c) medium alone (unstimulated controls). PBMC were stimulated in triplicate wells at 37°C in 5% CO2 for 72h. Cell proliferation was measured using an MTT assay (Cell Titer 96 R aqueous non-radioactive cell proliferation assay, Promega, Madison, WI). Stimulation indices (S.I) were calculated as above. For cytokine analysis, PBMC were stimulated with 1 µg/ml of rWbTSP LEL and medium alone (unstimulated) for 72 hrs.
Analysis of cytokine levels in the culture supernatants of the antigen specific cells
After 72hrs of stimulation with the antigens, culture supernatants of splenocytes were collected and the levels of secreted cytokines (IL-1, IL-4, IL-5, IL-10, IFN-γ, TNF-α) were measured using multianalyte ELISA array kit according to manufacturer’s instructions (SA biosciences, Valencia, CA). Cytokine values in the culture supernatant of cells incubated with rSmGBF or media alone was used as the background value. For human assays, levels of Interferon-γ (IFN-γ), Interleukin-4 (IL-4) and Interleukin- 10 (IL-10) were estimated in the culture supernatants of PBMC 72 hrs after incubation using human cytokine ELISA kits (E-biosciences, San Diego, CA) as per the manufacturer’s instruction. Sensitivity and minimum detection limits of the kits were 0.05 - 0.1 pg/ml. Concentration of each cytokine in the culture supernatant was plotted from a standard curve using recombinant rIFN-γ, rIL-4 or rIL-10 and the data was expressed as pg/ml.
Statistical analysis
GraphPad Prism version 5.0 (GraphPad Software, San Diego, CA) was used to analyze the data. Comparisons between two individual data points were made using a Student’s t-test. For multiple comparisons, one way ANOVA was used along with the Tukey–Kramer and/or Dunnet’s post-test wherever appropriate. For cytokine proliferation two-way ANOVA was used with Bonferroni post test. A probability (P) value of <0.05 was considered statistically significant.
Results
BmTSP LEL is expressed on the surface of B. malayi L3.
Recombinant WbTSP LEL, rBmTSP LEL and rOvTSP LEL with his tag had same molecular size of 12 kDa and appeared as a single band in the SDS PAGE gel. The predicted size of BmTSP LEL is 8 kDa plus his-tag resulted in 12 kDa. Subsequently we immunized mice with rBmTSP LEL and generated polyclonal antibodies. TSP is one of the relatively few integral membrane proteins that are consistently found in the tegument of S. mansoni [13], where they are believed to play an important role in cellular interactions and help maintain the integrity of tegument membrane [16,38]. Therefore, we first wanted to determine if BmTSP LEL is expressed on the surface of the third stage larvae (L3) of B. malayi. Our results showed significant fluorescence uniformly present throughout the cuticle of L3 (Figure 1A). These findings suggested that BmTSP LEL is expressed on the surface of L3. Incubation with control sera or secondary antibodies alone did not show any fluorescence on the surface of L3 (data not shown).
Figure 1. Characterizations of TSP LEL.

A. BmTSP LEL is expressed on the cuticle of B. malayi L3. Presence of BmTSP LEL on the surface of B. malayi L3 was demonstrated by staining L3 with mouse anti-BmTSP LEL antibodies followed by FITC conjugated anti-mouse IgG (1,2). Green fluorescence denotes regions where antibody was bound. No fluorescence found when L3 was stained with control negative sera (3). Scale bar is 11µm. B. Multiple sequence alignment of Tetraspanin was performed using clustalW online tools. Sequence alignment showed that LEL domain of WbTSP is 100% similar to BmTSP and 97% similar to OvTSP. Arrow showing only two amino acids (C35-Y35 /D48 -G48) of OvTSP LEL was different from BmTSP LEL and/or WbTSP LEL. C. Immunoblot was performed by probing anti-His tag, anti-BmTSPLEL or anti-WbTSPLEL antibodies. Both anti-WbTSPLEL and anti-BmTSP LEL antibodies cross reacted with rOvTSP LEL proteins.
Anti-Bm and Wb TSP LEL cross react with Ov TSP LEL.
Sequence alignment of WbTSP LEL showed 100% and 97% homology with BmTSP LEL and OvTSP LEL respectively (Figure 1B). This was further confirmed by comparing the sequence of TSP LEL with published WbTSP (Accession No: WUBG_09786) sequence at the NCBI site. This analysis showed 10aa differences in the LEL regions (data not shown). Polymorphisms have been reported in the SmTSP-2 gene as well [39]. Thus, we believe that these differences are due to polymorphisms in the TSP proteins, rather than to species differences. Subsequently, we also performed an immunoblot analysis to check the cross reactivity of anti-BmTSP LEL and anti-WbTSP LEL antibodies with rOvTSP LEL. Results showed that anti–BmTSP LEL and anti-WbTSP LEL antibodies cross react with rOvTSP LEL (Figure 1C).
Both LF human and mice sera carry cytophillic antibodies against TSP LEL
Expression of TSP LEL on the surface of B. malayi makes the antigen easily available to the host immune system. To determine if the sera from putatively immune and infected individuals carry antibodies against filarial TSP LEL, we first screened for the presence of anti-TSP LEL antibodies in the sera of different subjects (EN, CP, MF and NEN) using an indirect ELISA. Our results show that 14 out of 15 putatively immune endemic normal (EN) individuals carry circulating IgG antibodies against rWbTSP LEL (Figure 2) proteins. No significant levels of IgG were observed in the sera samples of MF, CP and NEN individuals. Positive reactions in the EN sera were determined as readings above the cut off value of mean + 3SD of NEN OD values. We also analyzed the isotype of anti-WbTSP LEL IgG antibodies in these sera samples. Our results showed that anti-TSP LEL IgG1 were predominant in the sera of EN individuals (Figure 3) compared to sera from other filarial groups (MF and CP). High levels of IgG3 antibodies were found both in EN and CP individuals. IgG2 and IgG4 isotypes did not show any significant differences between the filarial groups. Anti-TSP LEL IgG antibodies were not detectable in the sera of NEN individuals. Similar antibody responses were observed against rBmTSP LEL (data not shown).
Figure 2. TSP LEL specific IgG antibodies in the sera of LF human subjects.
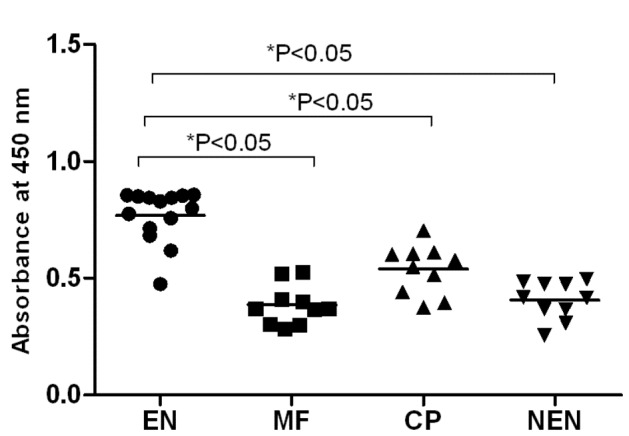
Levels of total IgG antibodies against rWbTSP LEL in the sera of Endemic normal (EN), Microfilaremic (MF), Chronic pathology (CP) and Non-endemic normal (NEN) subjects were measured using an indirect ELISA. 15 sera samples (diluted 1:100) were evaluated from EN, and 10 samples from MF, CP and NEN individuals. Each data point represents sera sample from a single individual. Horizontal lines represent geometric mean value. Data is represented as scatter plot where each dot represents absorbance of individual sera. Significant (*P<0.05) IgG antibodies in EN individuals compared to other groups (One way ANOVA along with Tukey-Kramer post statistics test was used).
Figure 3. TSP LEL specific IgG isotype of antibodies in the sera of human.
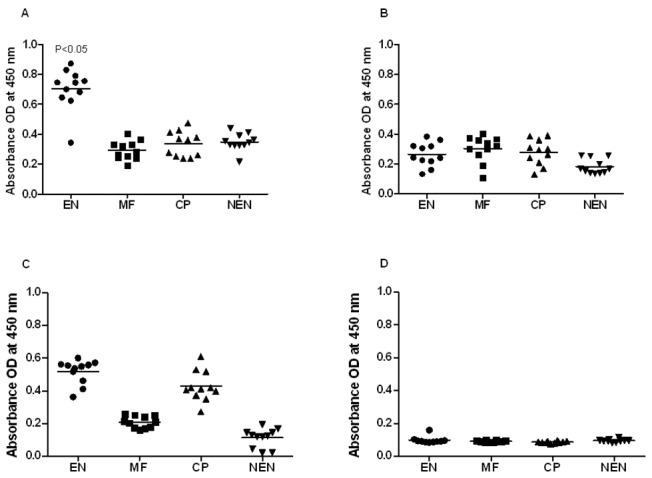
Isotype of IgG antibodies A) IgG1, B) IgG2, C) IgG3 and D) IgG4 against rWbTSP LEL were measured in the sera of putatively immune individuals (n=10) using an indirect ELISA. Each data point represents sera sample from a single individual. Horizontal lines represent geometric mean value. Data is represented as scatter plot where each dot represents absorbance of individual sera. Significant *(P<0.05) levels of isotype antibodies in EN individuals compared to other groups (One way ANOVA along with Tukey-Kramer post statistics test was used).
Similar results were obtained when we analyzed sera samples from mice immunized with rWbTSP LEL (Figure 4). Immunized mice showed significantly high titers of anti-WbTSP LEL IgG antibodies in their sera (Figure 4A). Subsequent isotype analysis showed that the anti-WbTSP LEL antibodies were predominantly of IgG1, followed by IgG2a and IgG2b isotypes (Figure 4B). Control animals did not show any IgG specific antibodies against rWbTSP LEL. Similar results were observed when rBmTSP LEL was used (data not shown)
Figure 4. TSP LEL specific IgG antibodies in the sera of mice.
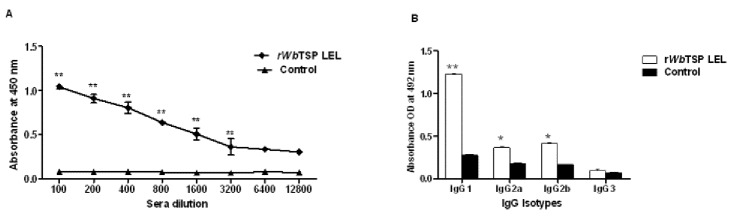
Mice were immunized four times at two weeks interval using 15 µg of rWbTSP LEL combined with alum adjuvant. A) Titer of anti-TSP LEL IgG antibodies. Approximately, 100 ng of recombinant proteins (100 ng /100µl /well) were coated onto the wells of an ELISA plate and bound serum IgG was detected using an HRP-labeled anti-mouse IgG secondary antibody. Each data point indicates mean ±S.D value from five animals. B). Isotype of anti-TSP LEL IgG antibodies in the sera of mice. Isotypes specific ELISA was performed as described in the methods section. Bars represent mean ±SD from five mice per group. ** Significant (P<0.001) and * (P<0.05) compared to control groups (Student’s t-test).
Anti-TSP LEL antibodies participated in the killing of B. malayi L3 in an ADCC mechanism
Results presented above show that sera of EN subjects and immunized mice have significantly high titer of anti-TSP LEL IgG antibodies. IgG antibodies can bind to FcγRI and FcγRII receptors expressed on several inflammatory cells such as macrophages, neutrophils, eosinophils and participate in an ADCC mechanism [40]. Therefore, in this study we evaluated if anti-TSP LEL antibodies in the sera of EN subjects and immunized mice can participate in the killing of B. malayi L3 in vitro in an ADCC assay. Our results show that pooled sera samples from EN subjects were able to kill 65% of B. malayi L3 in an ADCC assay. Depletion of anti-WbTSP-LEL antibodies from the sera of EN subjects resulted in the reduction of the killing ability of the sera by 2/3. When we depleted total IgG from the sera of EN subjects the killing ability was reduced by 85%. Incubating the larvae with cells alone or sera alone killed only <5% of L3 (Figure 5A). A similar ADCC assay performed using sera samples from immunized mice showed that ~67% of B. malayi L3 were killed when immune sera was used, whereas, only 5% of L3 were killed when sera samples from control animals were used (Figure 5B). These findings suggest that the anti-TSP LEL antibodies play a critical role in the killing of B. malayi L3 in vitro.
Figure 5. Sera from EN subjects and mice immunized with rWbTSP LEL participated in the killing of B. malayi L3 in an ADCC mechanism.
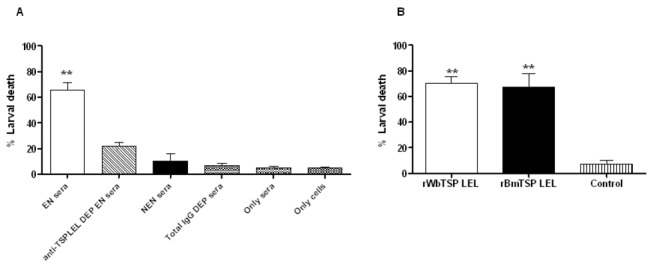
A) 50 µl of pooled EN sera (n = 10) samples or sera samples depleted off anti-TSP LEL antibodies were incubated with 2x105 peripheral blood mononuclear cells from normal healthy individuals and 10 B. malayi L3 for 48 hrs at 37°C. B) 50 µl of pooled sera (n = 5) samples from rWbTSP LEL immunized or rBmTSP LEL immunized mice were incubated with 2x105 peritoneal exudates cells from naive mice and 10 B. malayi L3 for 48 hrs at 37°C. Larval viability was evaluated at the end of the incubation. Values represent mean ± SD of three wells. Significant larval death ** (P<0.001) compare to controls analyzed by Student’s t-test.
Antibodies in the sera of EN subjects can recognize rOvTSP LEL.
Another important finding in this study was that the IgG antibodies in the sera of EN subjects could recognize rOvTSP LEL. Further analysis showed that the rOvTSP LEL recognizing antibodies in the sera of EN subjects were of IgG1 and IgG3 isotype. This suggested that potentially the same antibodies may be recognizing both rWbTSP LEL and rOvTSP LEL. This was not surprising given the significant sequence similarities between OvTSP LEL and WbTSP LEL.
We then tested the reverse cross reactivity, that is, if antibodies in the sera of Ov-PI subjects (these are PI subjects from an O. volvulus endemic area in Cameroon) will react with rWbTSP LEL. Our results show that IgG1 specific antibodies in the sera of Ov-PI individuals recognized rWbTSP LEL proteins (Figure 6A). Levels of anti-WbTSP LEL IgG3 antibodies were not significantly different in the sera of Ov-PI subjects and O. volvulus infected patients (Figure 6B). These findings suggested the presence of cross reactive IgG1 antibodies in the sera of LF-EN and Ov-PI subjects.
Figure 6. Sera from individuals who are putatively immune against O. volvulus infections (PI) has antibodies that cross react with rWbTSP LEL.
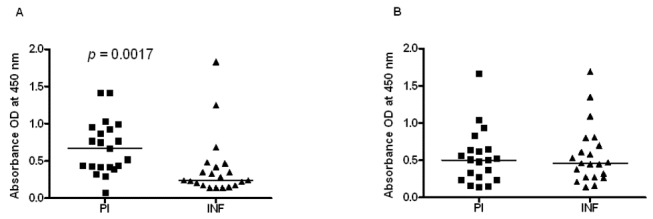
Isotype of IgG antibody A) IgG1 and B) IgG3 reactivity was measured in the sera of putatively immune individuals (PI) and O. volvulus infected individuals (INF) against rWbTSP LEL. Each data point represents sera sample from a single individual (n=20). Horizontal lines represent median value. Data is represented as scatter plot where each dot represents absorbance of individual sera. PI sera showing significant (P<0.001) cross reactivity of IgG1 antibodies compared to INF individuals. The OD of NEN (NYC individuals) was always below OD 0.1. Significant (P<0.001) levels of isotype antibodies in PI compared to INF group (Student’s t-test).
Vaccination of mice with rTSP LEL conferred protection against a challenge infection.
Vaccine potential of rBmTSP LEL or rWbTSP LEL was confirmed in the mouse model by challenging with 20 B. malayi L3 in a micropore chamber. Mice vaccinated with rBmTSP LEL and/or rWbTSP LEL gave 65% and 67% protection respectively against B. malayi L3 challenge (Table 1).
Table 1. Vaccine induced protection in mice.
| Animal groups | Number of L3 recovered Mean ± S.D | Number of Dead L3 Mean ± S.D | % Larval death Mean ± S.D |
|---|---|---|---|
| Trial-I | |||
| Alum (Control) | 16±1.34 | 1.4±0.543 | 8.27±2.592 |
| rWbTSPLEL | 18±2.70 | 11.6±2.073 | 63.2±7.505** |
| rBmTSP LEL | 16±2.70 | 10.6±0.894 | 65.44±6.898** |
| Trial-II | |||
| Alum (Control) | 19.2±2.38 | 2.2±0.83 | 2.2±0.83 |
| rWbTSPLEL | 15±0.707 | 9.75±1.14 | 64.28±9.49** |
| rBmTSP LEL | 17.25±2.79 | 10.25±1.67 | 60.33±3.46** |
In vivo micropore chamber assay was performed by surgically implanting 20 B. malayi L3 into the peritoneal cavity of each mouse. 48 hrs after implantation, chambers were removed and larval viability and larval death was determined. Values are mean ± SD. N = 5. Data presented here is from one of two similar experiments showing comparable results.
significant larval death (P < 0.001) compared to control mice groups analyzed by one way ANOVA followed by Dunnett’s post ANOVA test.
rTSP LEL specific IFN-γ secreting cells were present in the PBMC of EN subjects and in vaccinated mice.
PBMC from putatively immune EN subjects and spleen cells from vaccinated mice proliferated significantly in response to rWbTSP LEL stimulation suggesting the presence of antigen-specific memory cell population in both human and mice (Figure 7A and 7B). The stimulation indices were 1 fold and 2 folds higher compared to NEN individuals and control mice respectively. PBMC from MF and CP individuals did not show any significant proliferations. Similarly, spleen cells from control mice also did not show any significant proliferation.
Figure 7. Proliferation of PBMC from human subjects and spleen cells from vaccinated mice following stimulation with rWbTSP LEL.
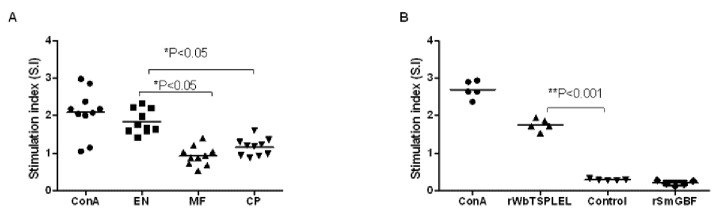
A. PBMC from Endemic Normal (EN), Microfilaraemic (MF) and Chronic Pathology (CP) groups were stimulated with rWbTSP LEL. Concanavalin A (ConA) was used as a positive control. Significantly high S.I. *(P<0.05) for cells from EN compared to cells collected from other groups, analyzed by two way ANOVA with Bonferonni test. B. Splenocytes from mice immunized with rWbTSP LEL and alum proliferated significantly to rWbTSP LEL stimulation. ConA was used as a positive control. Non-specific recombinant protein (recombinant S. mansoni G-binding factor, rSmGBF) was used as a negative control. Each data point represents stimulation index (S.I) of individual human sample (n = 10) and mice (n=5). Significantly high S.I. **(P<0.001) for cells from vaccinated animals compared to control cells analyzed by student’s t-test.
Analysis of the levels of secreted cytokines in the culture supernatants of PBMC stimulated with rWbTSP LEL showed that significantly high levels of IFN-γ (168 pg/ml) (P<0.001) was secreted by PBMC from EN subjects, whereas, PBMC from MF individuals secreted predominantly IL-10 (210 pg/ml) (Figure 8A). There were no significant differences in the levels of IL-4 in the culture supernatants of PBMC from different groups. Splenocytes from mice immunized with rWbTSP LEL secreted significant (P<0.05) amounts of IFN-γ (550 pg/ml) and IL-10 (300 pg/ml) in response to rWbTSP LEL (Figure 8B). However, secreted levels of IL-1, TNF-α, IL-4 and IL-5 were not significantly different in the culture supernatants of splenocytes compared to control.
Figure 8. Cytokine responses of rWbTSP LEL responding cells in the PBMC of human subjects and in the spleen of vaccinated mice.
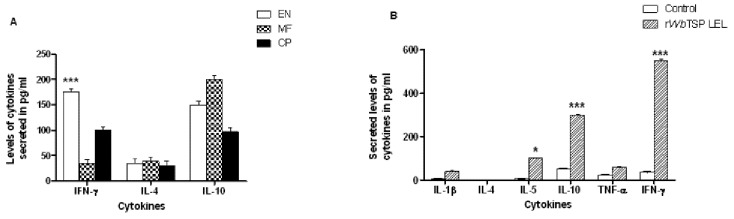
Cytokines (pg/ml) in the culture supernatants of human PBMC and mice spelnocytes were measured using an ELISA. A) Results show that significant level of IFN-γ is secreted by PBMCs from EN subjects in response to rWbTSP LEL. Significant ***(P<0.001) compared to other two groups (CP and MF) analyzed by One way ANOVA along with Tukey-Kramer post statistics test. B). IFN-γ level was prominent in the culture supernatants of spleen cells from rWbTSP LEL immunized mice compared to controls. Levels of secreted IL-10 were also high in the culture supernatants of EN PBMC and in the spleen cell culture supernatants of immunized mice. The data shown are normalized with the unstimulated controls. Significant cytokine levels ***(P<0.001) *(P<0.05) compared to control groups analyzed by Student’s t -test. Data is from one of two similar experiments showing comparable results.
Discussion
Lymphatic filariasis is the second leading cause of disability in the world [41], and is thus considered a major obstacle to economic development in the developing countries where this disease is endemic. Mass drug administration sphere headed by the World Health Organization is making substantial headway in reducing the incidence of this infection in several parts of the world [4]. Additional strategies such as vector control measures and an effective vaccine to prevent lymphatic filariasis will substantially supplement the effort towards elimination of this disease. Several previous studies have shown the feasibility of developing a vaccine against lymphatic filariasis and a number of potential antigens were identified as suitable candidates for vaccine against this infection in experimental animals [5, 7-9, 42]. In this study we show that the large extracellular loop of tetraspanin (TSP LEL) is a potential vaccine candidate against lymphatic filariasis. We also show that TSP LEL is present in several filarial parasites and antibodies generated against TSP LEL from one filarial parasite can cross-react with TSP LEL antigen from another filarial parasite suggesting a broader potential for developing TSP LEL as a common vaccine against several filarial parasites.
Molecules expressed on the surface of the parasite tend to modulate or down regulate host immune responses so the parasite can successfully establish infection in its host and/or escape host detection [43]. Targeting these parasite surface molecules have been used as a control strategy to reverse the host immunomodulation and allow the host immune system to recognize the parasite and eliminate them form the host [12]. Previous studies showed that TSP is expressed on the surface of helminthes [44,45]. Surface staining with anti-TSP LEL antibodies confirm that BmTSP LEL is also expressed on the surface of B. malayi L3. We have already reported that BmTSP is present in B. malayi L3, adults and Mf stages [21]. Thus, all stages of the parasite may potentially express TSP LEL suggesting an important role for this protein in these parasites. Targeting TSP by developing a vaccine against TSP LEL afforded substantial protection against primary alveolar echinococcosis [46] and S. mansoni [12]. These reports suggested that TSP LEL is an attractive target for developing a vaccine against helminthes.
Our previous studies showed that differential recognition of protective filarial antigens by endemic normal individuals can be a useful and rigorous approach to identify vaccine candidates against filarial infections [47]. Several previous studies demonstrate the presence of high titer of antibodies against key antigens in the sera of putatively immune endemic normal individuals [48,49]. In vitro ADCC assays confirm that these antibodies can participate in the killing of infective larvae of filarial parasites. Presence of these host protective antibodies in EN subjects, but not in the infected individuals (MF, CP) potentially suggests a strong correlation for selecting the candidate antigen that elicited the protective antibodies for further testing as a vaccine candidate. Based on this paradigm, in this study we first analyzed if anti-TSP LEL antibodies are uniquely present in EN subjects. Our results confirm the presence of high titer of anti-TSP LEL IgG antibodies in the putative immune EN subjects. These anti-TSP LEL IgG antibodies were found to be primarily cytophilic antibodies (IgG1 and IgG3). Similar antibody responses (IgG1 and IgG3) were demonstrated against SmTSP-2 LEL as well in individuals who are putatively resistant against schistosomiasis [12]. These findings suggest that parasite TSP LEL in general induces similar antibody responses in human. No detectable levels of rWbTSP LEL specific IgE antibodies were present in the sera of EN or in the sera of subjects who have filarial infection (MF and CP) (data not shown). This suggests that if rWbTSP LEL were to be developed as a vaccine for human use, potential allergic reactions will be minimal.
Immunization of mice with rBmTSP LEL or rWbTSP LEL resulted in the generation of anti-TSP LEL IgG1, IgG2a and IgG2b antibodies similar to that reported in mice vaccinated with Sm-TSP-2 [12]. These findings also suggested that parasite TSP LELs potentially induces similar types of antibody responses in the mouse model. Immune human subjects carry anti-TSP LEL IgG1 and IgG3 antibodies which is equivalent to IgG2a and IgG2b in mouse.
IgG1 and IgG3 antibodies can mediate antibody-dependent cellular cytotoxicity (ADCC) [50] and phagocytosis [51] by a variety of effector cells that express FcγRI and FcγRIII. Elevated levels of these cytophilic antibodies against rWbTSP LEL in putatively immune EN subjects and immunized mice suggested that these antibodies may participate in the killing of the parasite via ADCC mechanism. In fact, our results confirm that the anti-WbTSP LEL antibodies in LF-EN subjects and immunized mice have the ability to participate in the killing of B. malayi L3 through an ADCC mechanism. When we depleted the anti-WbTSP LEL antibodies from the sera samples, ability of the sera samples to participate in the killing B. malayi L3 was substantially decreased suggesting a key role for the anti-WbTSP LEL antibodies in the killing mechanism. These findings suggested that the anti-WbTSP LEL antibodies may have an important role in the defense against this parasitic infection in human.
Subsequently we evaluated the vaccine potential of rWbTSP LEL in the mouse model using a micropore chamber challenge approach. Although this is not truly a challenge model, given the difficulty in performing immunological analysis in jirds and due to prohibitive cost for the primate trials, the mouse micropore challenge system was used as a surrogate to challenge studies. Nevertheless, the data was interpreted with caution. Our results showed that immunization with rBmTSP LEL or rWbTSP LEL conferred 65% larval killing in this model. Compared to other vaccine proteins (BmHSP12.6, BmGST, BmVAL, BmTPX-2) tested in our laboratory [25][33] [35][42], rBmTSP LEL or rWbTSP LEL appears to be a good vaccine candidate against B. malayi. We also observed several cells attached firmly to the dead larvae in the micropore chambers. We did not identify the morphology of these cells; however, based on our ADCC results, we believe that these are possibly the effector cells. Protection conferred by rBmTSP LEL or rWbTSPLEL is also comparable to the protection reported for SmTSP-2 (57%–64%) in mice [12]. Similarly, vaccination trials in cestode infections also suggest that TSP is an attractive vaccine candidate against helminthes [46].
Evaluation of the immune correlates of rBmTSP LEL-induced protection showed that antigen specific memory cells are present in putatively immune LF-EN subjects and in vaccinated mice. Analysis of the cytokines secreted by these antigen-specific proliferating cells revealed that both PBMC from immune individuals and vaccinated mice splenocytes secreted IFN-γ and IL-10 in response to rTSP LEL stimulation. These findings were similar to those reported after vaccination of mice with SmTSP-2 [52]. Presence of antigen-specific IFN-γ secreting cells together with IL-10 secreting cells in the spleen suggest a balanced Th1/Th2 immune responses that may be important for regulating the inflammation caused by the host-protective mechanism in the filarial infections or for preventing the development of highly polarized Th1 responses [53,54]. Increase in the levels of IFN-γ suggests the activation of macrophages that may be important in the ADCC mediated killing of parasites observed in our studies [35] and others [55].
Another important observation in our studies is the finding that the TSP LEL sequences from different filarial parasites (B. malayi, W. bancrofti, O. volvulus) share sequence identity. This was reflected in the anti-TSP LEL antibody cross reactivity observed in the sera of putatively immune individuals for lymphatic filariasis and onchocerciasis. Because of the significance sequence identity, filarial TSP LEL can be used as a cross-protective vaccine antigen against both lymphatic filariasis and possibly O. volvulus infections in human. This observation has broader implications for the control of both the infections, especially, in parts of Africa where both these infections occur as co-infections [4]. However, challenge studies in animals are required to confirm this. Recently there is significant interest in identifying cross-protective vaccine antigens that are effective against multiple closely-related pathogens in an effort to reduce the cost of manufacturing and development pipelines [56].
In summary, we show that TSP LEL is expressed on the surface of B. malayi L3 and are thus easily accessible to the host immune system. The fact that putatively immune individuals carry IgG1 and IgG3 antibodies against WbTSP LEL and these antibodies participate in the killing of L3 in an ADCC assay confirm the potential of this antigen as a vaccine candidate against lymphatic filariasis. Our vaccination trials in the mouse model confirm this notion. Significant cross-reactivity of the anti-WbTSP LEL IgG1 antibodies in the sera of individuals who are protected against both lymphatic filarial and onchocerciasis infections suggest that WbTSP LEL could be developed as a potential vaccine candidate against onchocerciasis. These are only preliminary observations additional trials are needed to confirm the vaccine potential of rTSP LEL against filarial infections.
Acknowledgments
We would like to thank John Javeherian for the technical assistance with the animal experiments. B. malayi third stage infective larvae (L3) were obtained from NIH/NIAID filariasis research reagent resource center, College of Veterinary Medicine, University of Georgia, Athens, GA. We also thank Dr. Sara Lustigman and Nancy Tricoche of the Lindsley F. Kimball Research Institute, New York Blood Center, NY, for performing the ELISA with the repository sera from Cameroon, which was collected from participating individuals living in the villages around Kumba (Marumba I, Marumba II, Boa Bakundu, Bombanda, and Bombele).
Funding Statement
This study was supported by funding from National Institutes of Health through grant RO1 AI064745. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Michael E, Nutman TB (2000) Lymphatic filariasis. Tropical Medicine: Science and Practice London: Imperial College Press vol. 1: 41-82.
- 2. Lahariya C, Tomar SS (2011) How endemic countries can accelerate lymphatic filariasis elimination? an analytical review to identify strategic and programmatic interventions. J Vector Borne Dis 48: 1-6. PubMed: 21406730. [PubMed] [Google Scholar]
- 3. Patel V, Ramasundarahettige C, Vijayakumar L, Thakur JS, Gajalakshmi V et al. (2012) Suicide mortality in india: A nationally representative survey. Lancet 379: 2343-2351. doi: 10.1016/S0140-6736(12)60606-0. PubMed: 22726517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Hotez PJ (2009) Mass drug administration and integrated control for the world's high-prevalence neglected tropical diseases. Clin Pharmacol Ther 85: 659-664. doi: 10.1038/clpt.2009.16. PubMed: 19322166; Retrieved onpublished at whilst December year 1111 from 10.1038/clpt.2009.16 [DOI] [PubMed] [Google Scholar]
- 5. Madhumathi J, Prince PR, Anugraha G, Kiran P, Rao DN et al. (2010) Identification and characterization of nematode specific protective epitopes of Brugia malayi TRX towards development of synthetic vaccine construct for lymphatic filariasis. Vaccine 28: 5038-5048. doi: 10.1016/j.vaccine.2010.05.012. PubMed: 20653106. [DOI] [PubMed] [Google Scholar]
- 6. Murray J, Gregory WF, Gomez-Escobar N, Atmadja AK, Maizels RM (2001) Expression and immune recognition of Brugia malayi VAL-1, a homologue of vespid venom allergens and Ancylostoma secreted proteins. Mol Biochem Parasitol 118: 89-96. doi: 10.1016/S0166-6851(01)00374-7. PubMed: 11704277. [DOI] [PubMed] [Google Scholar]
- 7. Pokharel DR, Rai R, Nandakumar K, Reddy MV, Rathaur S (2006) Vaccination with Setaria cervi 175 kDa collagenase induces high level of protection against Brugia malayi infection in jirds. Vaccine 24: 6208-6215. doi: 10.1016/j.vaccine.2006.05.103. PubMed: 16870314. [DOI] [PubMed] [Google Scholar]
- 8. Sahoo MK, Sisodia BS, Dixit S, Joseph SK, Gaur RL et al. (2009) Immunization with inflammatory proteome of Brugia malayi adult worm induces a Th1/Th2-immune response and confers protection against the filarial infection. Vaccine 27: 4263-4271. doi: 10.1016/j.vaccine.2009.05.015. PubMed: 19450648. [DOI] [PubMed] [Google Scholar]
- 9. Vedi S, Dangi A, Hajela K, Misra-Bhattacharya S (2008) Vaccination with 73kDa recombinant heavy chain myosin generates high level of protection against Brugia malayi challenge in jird and Mastomys models. Vaccine 26: 5997-6005. doi: 10.1016/j.vaccine.2008.08.073. PubMed: 18817835. [DOI] [PubMed] [Google Scholar]
- 10. Schmid-Hempel P (2009) Immune defence, parasite evasion strategies and their relevance for 'macroscopic phenomena' such as virulence. Philos Trans R Soc Lond B Biol Sci 364: 85-98. doi: 10.1098/rstb.2008.0157. PubMed: 18930879; Retrieved onpublished at whilst December year 1111 from 10.1098/rstb.2008.0157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Maecker HT, Todd SC, Levy S (1997) The tetraspanin superfamily: Molecular facilitators. FASEB J 11: 428-442. PubMed: 9194523. [PubMed] [Google Scholar]
- 12. Tran MH, Pearson MS, Bethony JM, Smyth DJ, Jones MK et al. (2006) Tetraspanins on the surface of Schistosoma mansoni are protective antigens against schistosomiasis. Nat Med 12: 835-840. doi: 10.1038/nm1430. PubMed: 16783371. [DOI] [PubMed] [Google Scholar]
- 13. Tran MH, Freitas TC, Cooper L, Gaze S, Gatton ML et al. (2010) Suppression of mRNAs encoding tegument tetraspanins from Schistosoma mansoni results in impaired tegument turnover. PLOS Pathog 6: e1000840. doi: 10.1371/journal.ppat.1000840. PubMed: 20419145; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.ppat.1000840 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Moribe H, Yochem J, Yamada H, Tabuse Y, Fujimoto T et al. (2004) Tetraspanin protein (TSP-15) is required for epidermal integrity in Caenorhabditis elegans . J Cell Sci 117: 5209-5220. doi: 10.1242/jcs.01403. PubMed: 15454573. [DOI] [PubMed] [Google Scholar]
- 15. Hemler ME (2003) Tetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomain. Annu Rev Cell Dev Biol 19: 397-422. doi: 10.1146/annurev.cellbio.19.111301.153609. PubMed: 14570575. [DOI] [PubMed] [Google Scholar]
- 16. Jones MK, Gobert GN, Zhang L, Sunderland P, McManus DP (2004) The cytoskeleton and motor proteins of human schistosomes and their roles in surface maintenance and host-parasite interactions. Bioessays 26: 752-765. doi: 10.1002/bies.20058. PubMed: 15221857. [DOI] [PubMed] [Google Scholar]
- 17. Jones EL, Demaria MC, Wright MD (2011) Tetraspanins in cellular immunity. Biochem Soc Trans 39: 506-511. doi: 10.1042/BST0390506. PubMed: 21428929; Retrieved onpublished at whilst December year 1111 from 10.1042/BST0390506 [DOI] [PubMed] [Google Scholar]
- 18. Braschi S, Wilson RA (2006) Proteins exposed at the adult schistosome surface revealed by biotinylation. Mol Cell Proteomics 5: 347-356. doi: 10.1074/mcp.M500287-MCP200. PubMed: 16269422. [DOI] [PubMed] [Google Scholar]
- 19. Da’dara AA, Skelly PJ, Wang MM, Harn DA (2001) Immunization with plasmid DNA encoding the integral membrane protein, Sm23, elicits a protective immune response against schistosome infection in mice. Vaccine 20: 359-369. doi: 10.1016/S0264-410X(01)00374-7. PubMed: 11672898. [DOI] [PubMed] [Google Scholar]
- 20. Da'dara AA, Li YS, Xiong T, Zhou J, Williams GM et al. (2008) DNA-based vaccines protect against zoonotic schistosomiasis in water buffalo. Vaccine 26: 3617-3625. doi: 10.1016/j.vaccine.2008.04.080. PubMed: 18524429; Retrieved onpublished at whilst December year 1111 from 10.1016/j.vaccine.2008.04.080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Gnanasekar M, Anand SB, Ramaswamy K (2008) Identification and cloning of a novel tetraspanin (TSP) homologue from Brugia malayi . DNA Seq 19: 151-156. doi: 10.1080/10425170701517614. PubMed: 17852334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Cassone A, Rappuoli R (2010) Universal vaccines: Shifting to one for many. mBio 1: ([MedlinePgn:]) doi: 10.1128/mBio.00042-10. PubMed: 20689748. Retrieved onpublished at whilst December year 1111 from 10.1128/mBio.00042-10; Retrieved onpublished at whilst December year 1111 from 10.1128/mBio.00042-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Babayan SA, Allen JE, Taylor DW (2012) Future prospects and challenges of vaccines against filariasis. Parasite Immunol 34: 243-253. doi: 10.1111/j.1365-3024.2011.01350.x. PubMed: 22150082; Retrieved onpublished at whilst December year 1111 from 10.1111/j.1365-3024.2011.01350.x [DOI] [PubMed] [Google Scholar]
- 24. Taylor MJ, Hoerauf A, Bockarie M (2010) Lymphatic filariasis and onchocerciasis. Lancet 376: 1175-1185. doi: 10.1016/S0140-6736(10)60586-7. PubMed: 20739055; Retrieved onpublished at whilst December year 1111 from 10.1016/S0140-6736(10)60586-7 [DOI] [PubMed] [Google Scholar]
- 25. Anand SB, Rajagopal V, Kaliraj P (2012) Brugia malayi thioredoxin peroxidase as a potential vaccine candidate antigen for lymphatic filariasis. Appl Biochem Biotechnol 167: 1351-1364. doi: 10.1007/s12010-012-9643-6. PubMed: 22528648; Retrieved onpublished at whilst December year 1111 from 10.1007/s12010-012-9643-6 [DOI] [PubMed] [Google Scholar]
- 26. Makepeace BL, Jensen SA, Laney SJ, Nfon CK, Njongmeta LM et al. (2009) Immunisation with a multivalent, subunit vaccine reduces patent infection in a natural bovine model of onchocerciasis during intense field exposure. PLoS Negl Trop. Drosophila Inf Serv 3: e544. doi: 10.1371/journal.pntd.0000544; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.pntd.0000544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Wang J, Tricoche N, Du L, Hunter M, Zhan B et al. (2012) The adjuvanticity of an O. volvulus-derived rOv-ASP-1 protein in mice using sequential vaccinations and in non-human primates. PLOS ONE 7: e37019. doi: 10.1371/journal.pone.0037019. PubMed: 22615877; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.pone.0037019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Steel C, Guinea A, Ottesen EA (1996) Evidence for protective immunity to bancroftian filariasis in the cook islands. J Infect Dis 174: 598-605. doi: 10.1093/infdis/174.3.598. PubMed: 8769619. [DOI] [PubMed] [Google Scholar]
- 29. Rao KV, Eswaran M, Ravi V, Gnanasekhar B, Narayanan RB et al. (2000) The Wuchereria bancrofti orthologue of Brugia malayi SXP1 and the diagnosis of bancroftian filariasis. Mol Biochem Parasitol 107: 71-80. doi: 10.1016/S0166-6851(99)00231-5. PubMed: 10717303. [DOI] [PubMed] [Google Scholar]
- 30. Cho-Ngwa F, Liu J, Lustigman S (2010) The Onchocerca volvulus cysteine proteinase inhibitor, ov-CPI-2, is a target of protective antibody response that increases with age. PLoS Negl Trop. Drosophila Inf Serv 4: e800. doi: 10.1371/journal.pntd.0000800; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.pntd.0000800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. MacDonald AJ, Turaga PS, Harmon-Brown C, Tierney TJ, Bennett KE et al. (2002) Differential cytokine and antibody responses to adult and larval stages of Onchocerca volvulus consistent with the development of concomitant immunity. Infect Immun 70: 2796-2804. doi: 10.1128/IAI.70.6.2796-2804.2002. PubMed: 12010965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Kurniawan-Atmadja A, Sartono E, Partono F, Yazdanbakhsh M, Maizels RM (1998) Antibody responses to filarial infective larvae are not dominated by the IgG4 isotype. Parasite Immunol 20: 9-17. doi: 10.1046/j.1365-3024.1998.t01-1-00118.x. PubMed: 9491413. [DOI] [PubMed] [Google Scholar]
- 33. Veerapathran A, Dakshinamoorthy G, Gnanasekar M, Reddy MV, Kalyanasundaram R (2009) Evaluation of Wuchereria bancrofti GST as a vaccine candidate for lymphatic filariasis. PLoS Negl Trop. Drosophila Inf Serv 3: e457. doi: 10.1371/journal.pntd.0000457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Dakshinamoorthy G, Samykutty AK, Munirathinam G, Shinde GB, Nutman T et al. (2012) Biochemical characterization and evaluation of a Brugia malayi small heat shock protein as a vaccine against lymphatic filariasis. PLOS ONE 7: e34077. doi: 10.1371/journal.pone.0034077. PubMed: 22496777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Dakshinamoorthy G, Samykutty AK, Munirathinam G, Reddy MV, Kalyanasundaram R (2013) Multivalent fusion protein vaccine for lymphatic filariasis. Vaccine 31: 1616-1622. doi: 10.1016/j.vaccine.2012.09.055. PubMed: 23036503; Retrieved onpublished at whilst December year 1111 from 10.1016/j.vaccine.2012.09.055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Abraham D, Grieve RB, Holy JM, Christensen BM (1989) Immunity to larval Brugia malayi in BALB/c mice: Protective immunity and inhibition of larval development. Am J Trop Med Hyg 40: 598-604. PubMed: 2662785. [DOI] [PubMed] [Google Scholar]
- 37. Samykutty A, Dakshinamoorthy G, Kalyanasundaram R (2010) Multivalent vaccine for lymphatic filariasis. Procedia Vaccinol 3: 12-18. doi: 10.1016/j.provac.2010.11.003. PubMed: 21709765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Levy S, Shoham T (2005) The tetraspanin web modulates immune-signalling complexes. Nat Rev Immunol 5: 136-148. doi: 10.1038/nri1548. PubMed: 15688041. [DOI] [PubMed] [Google Scholar]
- 39. Cupit PM, Steinauer ML, Tonnessen BW, Eric Agola L, Kinuthia JM et al. (2011) Polymorphism associated with the Schistosoma mansoni tetraspanin-2 gene. Int J Parasitol 41: 1249-1252. doi: 10.1016/j.ijpara.2011.07.007. PubMed: 21889508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Teillaud J (2001) Antibody-dependent cellular cytotoxicity (ADCC). In: Anonymous eLS. John Wiley & Sons, Ltd. Retrieved onpublished at whilst December year 1111 from . doi: 10.1002/9780470015902.a0000498.pub2. [DOI] [Google Scholar]
- 41. Zeldenryk LM, Gray M, Speare R, Gordon S, Melrose W (2011) The emerging story of disability associated with lymphatic filariasis: A critical review. PLoS Negl Trop. Drosophila Inf Serv 5: e1366. doi: 10.1371/journal.pntd.0001366; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.pntd.0001366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Anand SB, Gnanasekar M, Thangadurai M, Prabhu PR, Kaliraj P et al. (2007) Immune response studies with Wuchereria bancrofti vespid allergen homologue (WbVAH) in human lymphatic filariasis. Parasitol Res 101: 981-988. doi: 10.1007/s00436-007-0571-2. PubMed: 17558521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Perez-Sanchez R, Ramajo-Hernandez A, Ramajo-Martin V [!(surname)!] (2006) Proteomic analysis of the tegument and excretory-secretory products of adult Schistosoma bovis worms. Proteomics, 6 Suppl 2 Suppl 1: 6–15: S226-36 doi: 10.1002/pmic.200500420. PubMed: 17031799. [DOI] [PubMed] [Google Scholar]
- 44. Dang Z, Yagi K, Oku Y, Kouguchi H, Kajino K et al. (2012) A pilot study on developing mucosal vaccine against alveolar echinococcosis (AE) using recombinant tetraspanin 3: Vaccine efficacy and immunology. PLoS Negl Trop. Drosophila Inf Serv 6: e1570. doi: 10.1371/journal.pntd.0001570; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.pntd.0001570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Hemler ME (2001) Specific tetraspanin functions. J Cell Biol 155: 1103-1107. doi: 10.1083/jcb.200108061. PubMed: 11756464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Dang Z, Yagi K, Oku Y, Kouguchi H, Kajino K et al. (2009) Evaluation of Echinococcus multilocularis tetraspanins as vaccine candidates against primary alveolar echinococcosis. Vaccine 27: 7339-7345. doi: 10.1016/j.vaccine.2009.09.045. PubMed: 19782112; Retrieved onpublished at whilst December year 1111 from 10.1016/j.vaccine.2009.09.045 [DOI] [PubMed] [Google Scholar]
- 47. Freedman DO, Nutman TB, Ottesen EA (1989) Protective immunity in bancroftian filariasis selective recognition of a 43-kD larval stage antigen by infection-free individuals in an endemic area. J Clin Invest 83: 14-22-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Ravindran B, Satapathy AK, Sahoo PK, Mohanty MC (2003) Protective immunity in human lymphatic filariasis: Problems and prospects. Med Microbiol Immunol 192: 41-46. doi: 10.1007/s00430-002-0157-7. PubMed: 12592562. [DOI] [PubMed] [Google Scholar]
- 49. Stewart GR, Zhu Y, Parredes W, Tree TI, Guderian R et al. (1997) The novel cuticular collagen ovcol-1 of Onchocerca volvulus is preferentially recognized by immunoglobulin G3 from putatively immune individuals. Infect Immun 65: 164-170. PubMed: 8975907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Rozsnyay Z, Sármay G, Walker M, Maslanka K, Valasek Z et al. (1989) Distinctive role of IgG1 and IgG3 isotypes in Fc gammaR-mediated functions. Immunology 66: 491-498. PubMed: 2523858. [PMC free article] [PubMed] [Google Scholar]
- 51. Kipps TJ, Parham P, Punt J, Herzenberg LA (1985) Importance of immunoglobulin isotype in human antibody-dependent, cell-mediated cytotoxicity directed by murine monoclonal antibodies. J Exp Med 161: 1-17. doi: 10.1084/jem.161.1.1. PubMed: 3918141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Pearson MS, Pickering DA, McSorley HJ, Bethony JM, Tribolet L et al. (2012) Enhanced protective efficacy of a chimeric form of the schistosomiasis vaccine antigen Sm-TSP-2. PLoS Negl Trop. Drosophila Inf Serv 6: e1564. doi: 10.1371/journal.pntd.0001564; Retrieved onpublished at whilst December year 1111 from 10.1371/journal.pntd.0001564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Liu XS, Leerberg J, MacDonald K, Leggatt GR, Frazer IH (2009) IFN-gamma promotes generation of IL-10 secreting CD4+ T cells that suppress generation of CD8 responses in an antigen-experienced host. J Immunol 183: 51-58. doi: 10.4049/jimmunol.0802047. PubMed: 19535638; Retrieved onpublished at whilst December year 1111 from 10.4049/jimmunol.0802047 [DOI] [PubMed] [Google Scholar]
- 54. Dash Y, Ramesh M, Kalyanasundaram R, Munirathinam G, Shultz LD et al. (2011) Granuloma formation around filarial larvae triggered by host responses to an excretory/secretory antigen. Infect Immun 79: 838-845. doi: 10.1128/IAI.00128-10. PubMed: 21078849; Retrieved onpublished at whilst December year 1111 from 10.1128/IAI.00128-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Fan S, Fehr HG, Adams D (1991) Activation of macrophages for ADCC in vitro: Effects of IL-4, TNF, interferons-α/β, interferon-γ, and GM-CSF. Cell Immunol 135: 78-87. doi: 10.1016/0008-8749(91)90255-A. PubMed: 1708310. [DOI] [PubMed] [Google Scholar]
- 56. Hoerauf A, Pfarr K, Mand S, Debrah AY, Specht S (2011) Filariasis in Africa--treatment challenges and prospects. Clin Microbiol Infect 17: 977-985. doi: 10.1111/j.1469-0691.2011.03586.x. PubMed: 21722251; Retrieved onpublished at whilst December year 1111 from 10.1111/j.1469-0691.2011.03586.x [DOI] [PubMed] [Google Scholar]