

Abstract
Here we report on three new species of ornithuromorph birds from the Lower Cretaceous Xiagou Formation in the Changma Basin of Gansu Province, northwestern China: Yumenornis huangi gen. et sp. nov., Changmaornis houi gen. et sp. nov., and Jiuquanornis niui gen. et sp. nov.. The last of these is based on a previously published but unnamed specimen: GSGM-05-CM-021. Although incomplete, the specimens can be clearly distinguished from each other and from Gansus yumenensis Hou and Liu, 1984. Phylogenetic analysis resolves the three new taxa as basal ornithuromorphs. This study reveals previously unrecognized ornithuromorph diversity in the Changma avifauna, which is largely dominated by Gansus but with at least three other ornithuromorphs. Body mass estimates demonstrate that enantiornithines were much smaller than ornithuromorphs in the Changma avifauna. In addition, Changma enantiornithines preserve long and recurved pedal unguals, suggesting an arboreal lifestyle; in contrast, Changma ornithuromorphs tend to show terrestrial or even aquatic adaptions. Similar differences in body mass and ecology are also observed in the Jehol avifauna in northeastern China, suggesting niche partitioning between these two clades developed early in their evolutionary history.
Introduction
In the last decade, nearly one hundred fossil birds have been discovered from the Lower Cretaceous Xiagou Formation in the Changma Basin of Gansu Province in northwestern China. Most of these are referred to Gansus yumenensis, the first Mesozoic fossil bird from China [1-4], although additional diversity has slowly accumulated; a second unnamed ornithuromorph bird [5], the enantiornithine Qiliania graffini [6], and three other unnamed enantiornithine specimens [7-9] have also been described.
The second unnamed ornithuromorph specimen, GSGM-05-CM-021, consists of an isolated sternum, furcula, and sternal ribs [5]. Its sternum is distinct from that of Gansus, representing a basal non-ornithurine member of Ornithuromorpha similar to Archaeorhynchus from the Jiufotang Formation of Liaoning Province in northeastern China [5,10]. Despite this growing diversity, all other taxa are represented by isolated specimens and Gansus is by far the dominant taxon in the Changma avifauna.
Here we describe two new ornithuromorph specimens from the Xiagou Formation in the Changma Basin. Based on comparison with GSGM-05-CM-021 [5,10], we also erect a new species for this specimen. We describe the morphology of the new specimens and discuss the significance of these taxa in understanding the Changma avifauna.
Methods
Anatomical nomenclature primarily follows Baumel and Witmer [11]. English equivalents are used for osteological terminology while Latin is maintained for muscles. All measurements were taken with calipers to the nearest 0.1 mm.
The two new specimens, GSGM-06-CM-013 and GSGM-08-CM-002, and the previously described GSGM-05-CM-021 (Gansu Geological Museum) were discovered in the Xiagou Formation near Changma in Gansu Province of northwestern China. Precise locality information is available to qualified researchers upon request. The issuing authority is the Gansu Geological Museum, represented by Da-Qing Li (co-author). All necessary permits were obtained for the described study, which complied with all relevant regulations.
A phylogenetic analysis was performed using the O’Connor and Zhou [12] dataset, with the addition of Yumenornis, Changmaornis, and Jiuquanornis [5] (File S1). Neornithes was represented by Anas platyrhynchos and Gallus gallus; Dromaeosauridae was scored as the outgroup. The dataset consists of 245 characters (31 ordered; all characters weighted equally) evaluated for 63 taxa; the matrix was analyzed with TNT [13], applying a heuristic search retaining the single shortest tree from every 1000, followed by an additional round of tree bisection reconnection (TBR) branch swapping.
Nomenclatural Acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix "http://zoobank.org/". The LSID for this publication is: urn:lsid:zoobank.org:pub:DC6A3D74-B8B7-41BA-A6BF-7BD48D6CD3C7. The electronic edition of this work was published in a journal with an ISSN, and has been archived and is available from the following digital repositories: PubMed Central, LOCKSS.
Results
Locality and Horizon
In the vicinity of the Changma Township, Yumen City, Jiuquan area, northwestern Gansu Province, China; Lower Cretaceous (early Aptian), Xiagou Formation [7-9,14] (Figure 1).
Figure 1. Geographic location of the Changma Basin (indicated by star), Gansu Province, People’s Republic of China.

Systematic Paleontology
Aves Linnaeus, 1758 [15]
Pygostylia Chiappe, 2002 [16]
Ornithothoraces Chiappe, 1995 [17]
Ornithuromorpha Chiappe, 2002[16]
Yumenornis huangi gen. et sp. nov.
urn:lsid:zoobank.org:act:EEC6E8A1-318F-491B-BD62-EFF58C4DBFC0
Holotype
Gansu Geological Museum (GSGM)-06-CM-013, partial sternum, complete right coracoid and scapula, partial furcula, and a complete, articulated right forelimb (Figure 2).
Figure 2. Photograph of Yumenornis huangi gen. et sp. nov., GSGM-06-CM-013 (A) and line drawing (B).

Abbreviations: a, acromion; amc, alular metacarpal; bt, bicipital tubercle; co, coracoid; cp, craniolateral process; df, distal fossa; ff, fragment of the furcula; fit, fragment of the intermediate trabecula; hu, humerus; lp, lateral process; lt, lateral trabecula; mc II, metacarpal II; mc III, metacarpal III; md, manual digits; sc, scapula; st, sternum; str, sternal ribs; ra, radius; ul, ulna; ule, ulnare; vt, ventral tubercle.
Etymology
The generic name “Yumen” is derived from the name of the city near the fossil locality. The specific name is dedicated to Mr. Zhao-Chu Huang of the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) for his long-lasting support to several generations of IVPP staff.
Diagnosis
Yumenornis huangi is distinguished from other known ornithuromorphs by the following unique combination of characters: sternum with angular rostral margin (~90°), lateral (zyphoid) processes, and robust, distally expanded lateral trabeculae; radius with deep distal fossa; ratio of length of manus relative to humerus 1.1. Yumenornis huangi can be separated from Gansus by the following morphological differences: the rostral margin of the sternum is sharper than that of Gansus (90° vs. 140° in Gansus); the lateral trabeculae are short and caudally expanded, contrasting with the unexpanded trabeculae in Gansus.
Description
The sternum is preserved in dorsal view, lacking the left half (Figure 2). The rostral margin defines an angle of approximately 90°, similar to Jianchangornis microdonta, Yanornis martini (Figure 3H), and the unnamed ornithuromorph GSGM-05-CM-021 (Jiuquanornis niui) (Figure 3D) also from Changma [5,18,19], but contrasting with the greater angle in Gansus (Figure 3E) (approximately 140°; GSGM-06-CM-011). A small notch is present on the midline of the rostral margin, indicating the coracoids were separated mediolaterally; but this notch may be due to preservation. The coracoidal sulci are well developed as relatively deep cranial grooves. The dorsal surface is concave, as in most flying neornithines. The right craniolateral process, although possibly flattened during diagenesis, projects craniolaterally, and defines a sharp angle of nearly 65°, similar to that of Yixianornis grabaui (Figure 3G), Yanornis martini [19,20], Songlingornis linghensis (Figure 3I) [12,21], and GSGM-05-CM-021 [5]. In Gansus (GSGM-06-CM-011) the craniolateral process defines a wider angle of approximately 100° and situates more cranially than that of Yumenornis. Four sternal ribs are preserved in articulation with the lateral margin, which is less than those in Gansus (five costal facets; GSGM-04-CM-020) and GSGM-05-CM-021 (potentially five costal facets) [5]. A rounded lateral (zyphoid) process is present prominently on the lateral margin, close to the cranial end of the lateral trabecula, similar to that of Yixianornis [20]. The lateral processes in Songlingornis and Gansus are more quadrangular, while in GSGM-05-CM-021 they are small and triangular. The lateral trabecula is short and robust, with a large fan-shaped caudal expansion, similar to that in Yixianornis, Yanornis, and Songlingornis; in Gansus, the lateral trabeculae are shorter, thinner, and slightly curved medially, without a caudal expansion (Figure 3E). The intermediate trabecula is damaged, with only a fragment remaining; it is thinner than the lateral trabecula and appears to curve medially, similar to Ambiortus dementjevi [22], Gansus, Songlingornis, Yanornis, and Yixianornis. Whether this process contacted the xiphial region defining a fenestra, as in Gansus, Songlingornis, Yanornis, and Yixianornis, cannot be determined. The preserved morphology of the sternum of Yumenornis is most similar to that of Yixianornis and Songlingornis; all three taxa have sharp craniolateral processes, large and rounded lateral processes, and caudal expansions of the lateral trabeculae.
Figure 3. Comparison of sterna of Yumenornis huangi (C) and Jiuquanornis niui (D) with other Early Cretaceous ornithuromorphs.

A, Longicrusavis houi; B, Hongshanornis longicresta; E, Gansus yumenensis; F, Archaeorhynchus spathula; G, Yixianornis grabaui; H, Yanornis martini; I, Songlingornis linghensis. Dashed lines in C represent estimated reconstruction.
The right coracoid is strut-like, preserved in dorsal view; however, it is partially covered proximally by the scapula and distally by the sternum (Figure 2). The rounded acrocoracoid process appears to be hooked medially, although it is somewhat destroyed. The procoracoid process is well-developed and projected medially. The body of the coracoid bears a large, shallow, and triangular fossa for the attachment of the m. sternocoracoidei impression, as in other Early Cretaceous ornithuromorphs (e.g. Gansus, Yanornis, and Jianchangornis). The medial and lateral margins are approximately the same length, and are both strongly concave, forming an expanded sternal margin, as in Yanornis and Yixianornis. In Gansus, the lateral margin is more strongly concave than the medial margin. The sternal margin is covered by the sternum, and the presence of a lateral process, like that present in most ornithuromorphs, cannot be determined. The right scapula is completely preserved but slightly crushed proximally, obscuring most details of the articular surfaces. The corpus is curved and slightly tapered distally. The acromion process is short, only extending slightly beyond the coracoidal tubercle, contrasting with the longer process in Gansus. A delicate, curved fragment of bone is interpreted as part of the right furcular ramus.
The right humerus is completely preserved in caudal view, although covered by the scapula proximoventrally and badly crushed distally (Figure 2). The humeral head appears globose: it is proximally and caudally convex, and projects more proximally than the deltopectoral crest, like most ornithuromorphs. The small ventral tubercle is weakly separated from the humeral head by a shallow capital incision; it is weakly excavated ventrally by a shallow pneumotricipital fossa. The deltopectoral crest extends for 44% the total length of the humerus, similar to that of Gansus (43%, GSGM-05-CM-014); the width of the crest is slightly less than that of the shaft (wider than in Yanornis but narrower than in Gansus and hongshanornithids) [23]. The crest tapers into the shaft distally, as in most Early Cretaceous ornithuromorphs. The ratio of the width of the midshaft to the total length of humerus in Yumenornis is 0.1 (Figure 4A), much larger than those observed in specimens of Gansus GSGM-05-CM-014 (0.07) and GSGM-IG-04-CM-004 (0.06) (Figure 4B); the robust humerus resembles those of Yanornis (0.1, IVPP V13278) and Yixianornis (0.1, IVPP V13631) from The Jehol Group (Figure 4C, D).
Figure 4. Comparison of forelimbs of Yumenornis huangi (A) and other Early Cretaceous ornithuromorphs.
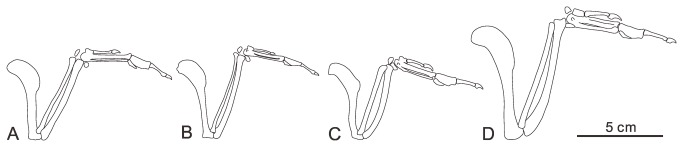
B, Gansus yumenensis; C, Yixianornis grabaui; D, Yanornis martini.
The ulna and radius are both severely damaged (Figure 2). The ulna is longer than the humerus and only slightly bowed, similar to the condition in Gansus, but unlike the relatively short and strongly bowed ulnae of Yixianornis and Yanornis (Figure 4). Proximally, the olecranon process is short and rounded. The bicipital tubercle is larger and more rounded than that of Gansus. An extremely deep fossa is present on the lateral surface of the distal end of the radius, nearly perforating the bone; this fossa appears more likely to be formed naturally than the result of preparation. Only the ulnare is preserved; it bears a shallow, round fossa on the possibly dorsal ramus, resembling that of Gansus. Both the ulna and radius are more robust than those in Gansus (Table 1), while similar to those of Yixianornis, respectively (IVPP V13631; Figure 4) [20].
Table 1. Measurements (mm) of forelimbs of Gansus yumenensis (CAGS-IG-04-CM-004, GSGM-05-CM-014) and Yumenornis huangi gen. et sp. nov. (GSGM-06-CM-013).
| Measurement | Specimen |
||
|---|---|---|---|
| CAGS-IG-04-CM-004 | GSGM-05-CM-014 | GSGM-06-CM-013 | |
| Humerus length | 48.0 | 47.8 | 49.9 |
| Ulna length | 49.1 | 51.1 | 52.9 |
| Ulna midshaft width | 2.9 | 3.0 | 3.6 |
| Radius length | 46.7 | 48.9 | 49.7 |
| Radius midshaft width | 1.6 | 1.7 | 2.0 |
| Carpometacarpus length | 23.2 | 23.7 | 27.0 |
| Metacarpal II width | 1.8 | 1.4 | 2.0 |
| Metacarpal III width | 0.8 | 0.7 | 0.9 |
| Phalanx I-1 length | 8.8 | 9.6 | 10.9 |
| Phalanx I-2 length | 3.9 | 3.6e | 5.4 |
| Phalanx II-1 length | 9.7 | — | 12.1 |
| Phalanx II-2 length | 9.0 | — | 11.5 |
| Phalanx II-3 length | 3.2 | 2.8 | 4.2 |
| Phalanx III-1 length | — | — | 6.7 |
| Phalanx III-2 length | — | 5.8 | 6.3 |
e Estimated value.
The right carpometacarpus is preserved in dorsal view; the proximal end is heavily damaged and the degree of fusion is unclear (Figure 2). The alular metacarpal is short and nearly rectangular; the cranial half is convex, developed into a small extensor process that only slightly surpasses the distal articular facet for phalanx I-1in cranial extent, similar to those in Gansus and Yixianornis, but less developed than that of Apsaravis ukhaana [24]. Metacarpals II and III are both straight. Metacarpal II is much wider than metacarpal III, which is very thin. Their distal ends appear not to be fused or are only partially fused, suggesting the specimen is a subadult; metacarpal II extends distally slightly farther than metacarpal III, and both the distal ends are expanded. The two metacarpals define a narrow intermetacarpal space that is more than twice the width of metacarpal III (narrower in Gansus; Figure 4).
The manual digits are preserved completely in articulation (Figure 2). Phalanx I-1 is slightly bowed and tapered distally, but expanded at the articular surface. Phalanx I-2 is a weakly recurved claw. This digit is very short, ending well proximal to the distal end of metacarpal II, as in other ornithuromorphs. Phalanx II-1 is craniocaudally expanded to nearly twice the width of metacarpal II. Phalanx II-2 is approximately the same length as II-1, but is thin, slightly bowed, and tapered medially so that the proximal and distal ends are both expanded relative to the shaft. Phalanx II-3 is an ungual, smaller than that of the alular digit, bearing a shallow extensor groove along its length. Phalanx III-1 is relatively slender, slightly less than half the length of II-1 and less than one third the width. It is wedge-shaped and slightly tapered distally, with a small, pointed, caudally directed tubercle. All the manual phalanges of Yumenornis are proportionately longer and more robust than those of Gansus (Table 1), but equal to those of Yixianornis (IVPP V 13631; Figure 4) [20].
Changmaornis houi gen. et sp. nov.
urn:lsid:zoobank.org:act:8E3453BC-A004-4383-BC3C-A889F0F9E2B9
Holotype
GSGM-08-CM-002, two thoracic vertebrae, a thoracic rib, partial synsacrum, a nearly complete right ischium and pubis, right distal tibiotarsus, and a complete, articulated right foot (Figure 5).
Figure 5. Photograph of Changmaornis houi gen. et sp. nov., GSGM-08-CM-002 (A) and line drawing (B).

Abbreviations: cp, costal process; il, ilium; is, ischium; mec, medial epicondyle; pu, pubis; syn, synsacrum; tb, tibiotarsus; tm, tarsometatarsus; tr, thoracic rib; tv, thoracic vertebra; I- IV, pedal digit I- IV. Numbers 1-11 in B indicate the number of the sacral vertebrae.
Etymology
The generic name “Changma” is derived from the name of the town where the fossil locality belongs to. The specific name is in honor of Professor Lian-Hai Hou, who named Gansus yumenensis.
Diagnosis
Changmaornis houi is distinguished from other known ornithuromorphs by the following unique combination of characters: synsacrum composed of at least 11 sacral vertebrae with elongate distal transverse processes; ischium with dorsal process; distal half of the pubis compressed mediolaterally; metatarsal I J-shaped; distal margin of metatarsal II trochlea does not reach the proximal margin of the metatarsal III trochlea; pedal digit III longest in foot; ratio of pedal digit III to tibiotarsus 0.82; robust and blunt pedal unguals with poorly developed flexor tubercles. Changmaornis houi can be distinguished from Gansus by the following morphological differences: pedal digit III is the longest and shorter than tarsometatarsus, while in Gansus pedal digit IV is the longest and longer than tarsometatarsus; pedal unguals lack well-developed flexor tubercles, contrasting with the pointed flexor tubercles in Gansus.
Description
A relatively complete thoracic vertebra is preserved in articulation with another completely damaged thoracic (Figure 5). The spinous process is rectangular, with a distinct ridge forming the dorsal margin. The vertebral centrum is laterally excavated by a deep, broad fossa, similar to those of other basal ornithuromorphs (e.g. Gansus, Yixianornis, Yanornis, and Hongshanornis longicresta [25]). The length of the centrum is nearly twice its width. The articular surfaces appear to be amphicoelous. The proximal half of an isolated thoracic rib is preserved. It bears a round costal tubercle on the proximal end. The synsacrum is preserved in ventral view, crushed so that the cranial and caudal ends are unclear. The two cranialmost sacral vertebrae bear laterodorsally directed costal processes. The synsacrum is formed by at least 11 completely fused vertebrae based on the number of preserved transverse processes (Figure 5B). In most ornithuromorphs, the synsacrum is composed of at least nine vertebrae; Yixianornis and Yanornis both have nine fused sacral vertebrae [19,20], while the more derived Gansus [2], Apsaravis [24], and Ichthyornis [26] possess 10 fused sacrals. The basalmost ornithuromorph, Archaeorhynchus spathula, possesses only seven to eight fused sacral vertebrae [10,12,27], although this may potentially be ontogenetic given that all known specimens are subadult [10]. In enantiornithines, the synsacrum is composed of seven or eight vertebrae [12,28]. The transverse processes of the sacral vertebrae are much longer than those of Gansus [2]; in the first few vertebrae they are laterocaudally directed, becoming more caudally directed and elongate in the last four synsacral vertebrae, similar to Yanornis [19].
Of the two ilia, only the proximal half of the right side and a fragment of the left are preserved, although these do not provide many anatomical details; the lateral margins appear to be deeply concave in ventral view (Figure 5). The right ischium and pubis are not preserved in articulation and are missing their proximal ends. The dorsal process of the ischium is large and obtusely triangular, as in Gansus and other ornithuromorphs, but is more proximally located than in Yixianornis. The distal end of the ischium tapers to its terminus, as in Gansus and Yixianornis. The pubis is medially curved and mediolaterally compressed along its distal half, contrary to the uncompressed pubis in Gansus. The distal end of the pubis does not expand ventrally although a dorsal expansion may have been present.
The distal end of the right tibiotarsus is preserved in medial view, in articulation with the completely preserved right tarsometatarsus and pedal phalanges (Figure 5). A medial epicondyle is developed on the medial surface of the distal tibiotarsus. Two tubercles are developed on the cranial surface of the distal end of the tibiotarsus. One is subrounded, located midway between the medial and lateral condyles, and bounded by a shallow fossa. The other is developed proximal to the medial condyle, forming a cranially projecting, proximodistally elongate ridge. The shape and position of the two tubercles resemble the paired ridges present in Apsaravis, which are interpreted as comparable to the tubercles for attachment of the extensor retinaculum in Neornithes [24]. However, three tubercles are present cranially on the distal end of the tibiotarsus in Gansus (GSGM-05-CM-014): the distalmost tubercle is subtriangular; the next tubercle is smaller, and located proximal and lateral to the first; the proximalmost one is medially located, forming an elongate ridge similar to the second one just described for Changmaornis.
As in Gansus, the tarsometatarsus is straight and completely fused (Figure 5). The dorsal surface of the proximal end is somewhat crushed; a large prominence without grooves or ridges projects weakly on the plantar surface of the proximal end, interpreted as a primitive hypotarsus, similar to that in Gansus and other basal ornithuromorphs (e.g. Yixianornis and Yanornis) [20,29]. The proximodorsal surface bears a deep dorsal infracotylar fossa, and the tubercle for the attachment of the m. tibialis cranialis is located on the mediodistal margin of this fossa [11]. Metatarsal IV is buried in the matrix; only metatarsals I, II, and III are visible. The short metatarsal I is nearly “J”-shaped in medial view, articulating with the medial surface of metatarsal II proximal to the trochlea, similar to the condition in Gansus. A small collateral ligament pit is present on the medial surface of the trochlea of metatarsal I. Metatarsal II does not extend distally to the base of the metatarsal III trochlea, and is displaced plantarly; the proximal position of metatarsal II trochlea strongly resembles that in Gansus. The trochlear surface of metatarsal II is partially ginglymous, while metatarsal III is fully ginglymous. Both metatarsals bear a pit for the collateral ligament on the medial surface of the trochlea.
The pedal phalanges are well preserved (Figure 5). Digit I is completely reversed, as in Gansus. The first phalanx in each digit is proportionately shorter than that of Gansus (CAGS-IG-04-CM-008 and GSGM-05-CM-014; Table 2). The nonungual phalanges of digits II-IV are also shorter than those in Gansus (Table 1; Figure 6C). Digit III is the longest, as in most ornithuromorphs (e.g. Schizooura, Yanornis, and Yixianornis; Figure 6) including neornithines [10,19,20,23,25], although the digit is shorter than the tarsometatarsus; the ratio of the length of digit III (including the ungual) relative to the tarsometatarsus is approximately 0.82, similar to that of Schizooura (0.80, IVPP V16861; Figure 6B) [30]. This clearly distinguishes this specimen from Gansus (CAGS-IG-04-CM-008), in which digit IV is the longest and the ratio of the length of digit III (including the ungual) to the tarsometatarsus is nearly 1.21.
Table 2. Measurements (mm) of hind limbs of Gansus yumenensis (CAGS-IG-04-CM-008, GSGM-05-CM-014) and Changma houi gen. et sp. nov. (GSGM-08-CM-002).
| Measurement | Specimen |
||
|---|---|---|---|
| CAGS-IG-04-CM-008 | GSGM-05-CM-014 | GSGM-08-CM-002 | |
| Tarsometatarsus length | 31.5 | 40 | 36.9 |
| Pedal phalanx I-1 length | 8.1 | 7.3 | 7.4 |
| Pedal phalanx I-2 length | 3.7 | 4.1 | 3.9 |
| Pedal phalanx II-1 length | 13.7 | 15.1 | 10 |
| Pedal phalanx II-2 length | 10.4 | 12.9 | 9.9 |
| Pedal phalanx II-3 length | 4.4 | 4.6 | 4.8 |
| Pedal phalanx III-1 length | 13.6 | 13.5 | 11.4 |
| Pedal phalanx III-2 length | 8.5 | 12.2 | 7.4 |
| Pedal phalanx III-3 length | 7.1 | 9.0 | 7.3 |
| Pedal phalanx III-4 length | 4.3 | 4.6 | 4.2 |
| Pedal phalanx IV-1 length | 11.5e | 12.0 | 8.5 |
| Pedal phalanx IV-2 length | 8.3 | 9.7 | 6.3 |
| Pedal phalanx IV-3 length | 7.5 | 8.7 | 4.9 |
| Pedal phalanx IV-4 length | 7.2 | 9.3 | 4.9 |
| Pedal phalanx IV-5 length | 3.5 | 3.7 | 3.6 |
e Estimated value.
Figure 6. Comparison of pedes of Changmaornis houi (A) and other Early Cretaceous ornithuromorphs.

B, Schizooura lii; C, Gansus yumenensis; D, Yixianornis grabaui; E, Yanornis martini.
The unguals are all small but robust (Figure 5); ungual IV is the shortest, and II and III are the longest. Relatively deep extensor grooves (compared to those in Gansus) run the entire length of the medial and lateral surfaces of all the unguals, resembling Yixianornis (Figure 6D), rather than restricted to the distal portion as in Gansus (CAGS-IG-04-CM-008). Flexor tubercles are nearly absent; the proximal ends have plantarly projected, weak and rounded tubercles that can barely be distinguished from the proximoventral corner of the articular cotyla.
Jiuquanornis niui gen. et sp. nov.
urn:lsid:zoobank.org:act:DF96E3B8-4A15-403D-85BC-EE46517858E6
Holotype
GSGM-05-CM-021, a complete, associated sternum and furcula, with sternal ribs and possible thoracic rib fragments (Figure 7) [5].
Figure 7. Photograph of Jiuquanornis niu gen. et sp. nov., GSGM-05-CM-021.

Etymology
The generic name “Jiuquan” is derived from the name of the city near the fossil locality. The specific name is dedicated to Professor Shao-Wu Niu, for his contribution to geological research in the Changma Basin.
Diagnosis
Jiuquanornis niui is distinguished from other known ornithuromorphs by the following unique combination of characters: U-shaped furcula without a hypocleidium; short imperforate body of sternum; small lateral processes on sternum; lateral trabeculae distally expanded medially; elongate intermediate trabeculae, equal to lateral trabeculae in distal extent; V-shaped xiphoid formed by short, fused medial trabeculae [5]. The sternum of Jiuquanornis niui can be clearly differentiated from that of Gansus by the long lateral and intermediate caudal trabeculae, V-shaped xiphoid, and absence of caudal fenestrae.
Comments
Jiuquanornis niui was first described as an unnamed ornithuromorph by You et al. [5]. Where comparison allows, the specimen is morphologically distinct from all other described ornithuromorphs, and thus we erect a new taxon. The sternum of Jiuquanornis strongly resembles that of Archaeorhynchus: both have long lateral and intermediate caudal trabeculae separated by deep incisures, which distinctly separate them from other basal ornithuromorphs; the trabeculae extend well past the caudal midline margin in both taxa. However, the two taxa differ from each other in the following features of the sternum: the rostral margin in Jiuquanornis defines an angle of approximately 100°, constituting a sharp craniomedial apex [5], while the rostral margin in Archaeorhynchus is more rounded, defining a greater angle of nearly 110°; the craniolateral processes in Jiuquanornis are smaller and more pointed than those of Archaeorhynchus; Jiuquanornis possesses a tapered, V-shaped xiphoid process, while the xiphial region in Archaeorhynchus is very short, lacking a distinct process (Figure 3F) [10]. The furcula of Jiuquanornis is similar to that of Archaeorhynchus and other basal ornithuromorphs (e.g. Gansus, Jianchangornis, and Yanornis); it is U-shaped with a low interclavicular angle of approximately 36°, and lacks a hypocleidium (Figure 7).
Phylogenetic Analysis
The analysis produced 5040 most parsimonious trees (MPTs) of 842 steps. The MPTs differed mainly in the placement of Zhongjianornis, Chaoyangia, and the three new Changma taxa within Ornithuromorpha. The strict consensus of these trees supports Yumenornis, Changmaornis, and Jiuquanornis as basal ornithuromorphs (Figure 8).
Figure 8. Strict consensus cladogram illustrating the phylogenetic positions of Yumenornis huangi, Changmaornis houi, and Jiuquanornis niui.

Tree length: 842 steps, consistency index = 0.387, retention index = 0.670.
The relationships among long-tailed birds, basal pygostylians, and enantiornithines concur with the analysis of O’Connor and Zhou [12]. Archaeorhynchus is resolved as the basalmost ornithuromorph; Zhongjianornis, Chaoyangia, Patagopteryx, Jianchangornis, Schizooura, and Vorona form a polytomy with the clade formed by more derived taxa, as in previous analyses [12,18,20,27,31]. However, the addition of the Changma specimens has reduced resolution in the relationships of more derived ornithuromorphs [12,29]: Yumenornis, Changmaornis, Jiuquanornis, Gansus, Yixianornis, Hollanda, Hongshanornis, Apsaravis, Longicrusavis, Ambiortus, and Yanornis + Songlingornis form a large polytomy that includes Ornithurae.
Although Yumenornis, Changmaornis, and Jiuquanornis are confirmed to be members of Ornithuromorpha, the interrelationships of these three taxa and their positions relative to other ornithuromorphs remain vague. The phylogenetic hypothesis is weakly supported, as indicated by the low consistency index (0.387) and retention index (0.670); this may result from the lack of data concerning most characters in the largely incomplete new specimens (Yumenornis, 81% incomplete; Changmaornis, 89% incomplete; Jiuquanornis, 94% incomplete; File S1) and the fragmentary or poorly preserved nature of some problem taxa (e.g. Chaoyangia, 91% incomplete; Zhongjianornis, 63% incomplete) [12].
Discussion
Yumenornis is confirmed to be an ornithuromorph bird by the presence of an elongate and imperforate sternal body, a strut-like coracoid with a well-developed procoracoid process, a curved scapula, a prominently convex humeral head, and a craniocaudally expanded first phalanx of manual digit II. Changmaornis is also referable to Ornithuromorpha by the presence of a synsacrum composed of at least 11 sacral vertebrae, a completely fused tarsometatarsus, and the plantar displacement of the proximal end of metatarsal III relative to metatarsals II and IV.
Yumenornis can be clearly differentiated from Gansus and Jiuquanornis based on sternal morphology alone: the rostral margin of the sternum defines an angle of approximately 90°, contrasting with the greater angle in Gansus; the lateral process is larger and more rounded than that of Jiuquanornis; the sternal body is comparatively much longer than that of Jiuquanornis; and the lateral trabecula is short with a large, fan-shaped caudal expansion, contrasting with the long, asymmetrically expanded lateral trabecula in Jiuquanornis or the unexpanded strap-like trabecula in Gansus (Figure 3). The sternum of Yumenornis can be also distinguished from other basal ornithuromorphs by the unique combination of a relatively straight rostral margin with a low intersecting angle (more rounded and with a greater angle in Archaeorhynchus and hongshanornthids), a sharp craniolateral process (more obtuse in Archaeorhynchus, Jianchangornis, Hongshanornis, and Yixianornis), a large and rounded lateral process (more quadrangular in hongshanornthids and Songlingornis; smaller in Archaeorhynchus; absent in Jianchangornis and Yanornis), and a short lateral trabecula with caudal expansion (strap-like in hongshanornthids) (Figure 3). The new specimen is also more robust than previously described Changma ornithuromorphs; the ratio of the width of the midshaft to the total length of the humerus is more than 50% greater than in Gansus. Yixianornis and Songlingornis both possess similar sterna to Yumenornis. All three taxa have well developed craniolateral processes, lateral processes, and the short, robust lateral trabeculae all bear large caudal expansions, although there are slight differences in the shape of the expansion compared of Yumenornis (potentially preservational). In addition, the length of the forelimb of Yumenornis (Table 1) is approximately equal to that of Yixianornis (IVPP V13631) [20], and both taxa possess robust humeri (the ratio of the midshaft width to the length of the humerus is 0.1 in both of the taxa) (Figure 4). These similarities may suggest a close relationship between these taxa.
Changmaornis can be separated from Gansus by several morphological differences: the transverse processes of the synsacrum are longer than those of Gansus; the distal half of the pubis in Changmaornis is compressed mediolaterally, whereas it is rod-like in Gansus; two tubercles on the cranial surface of the distal end of the tibiotarsus contrasting with three tubercles in Gansus; pedal digit III is the longest and shorter than the tarsometatarsus (pedal digit IV is longest in Gansus, and both digit III and IV are longer than the tarsometatarsus); the grooves on the lateral and medial surfaces of the unguals are deep and run nearly the entire length of the surface (shallow and distally restricted in Gansus); and well-developed flexor tubercles are absent (pointed flexor tubercles in Gansus). The tarsometatarsus of Changmaornis is unique from other basal ornithuromorphs, although it somewhat resembles that of Gansus. The metatarsal II trochlea is displaced plantarly and does not overlap with the metatarsal III trochlea in medial view, similar to that of Gansus, whereas in other basal ornithuromorphs (e.g. hongshanornthids, Yixianornis, Yanornis, and Apsaravis) the metatarsal II trochlea is only slightly displaced plantarly; in Changmaornis this trochlea does not reach the proximal margin of the metatarsal III trochlea, and thus is relatively even shorter than observed in Gansus and other Early Cretaceous ornithuromorphs.
Jiuquanornis is confirmed to be a member of Ornithuromorpha by its U-shaped furcula lacking a hypocleidium [5]. During the evolution of Aves, the number of fused vertebrae forming the synsacrum increases in more derived taxa [10,12,24]. A synsacrum composed of at least 11 fused vertebrae suggests that Changmaornis is a more derived ornithuromorph than Gansus, Yixianornis, and Yanornis. Although unsupported by the phylogenetic analysis, we suggest that Changmaornis may also be more derived than Yumenornis. The close relationship of Jiuquanornis and Archaeorhynchus based on numerous similarities of their sterna suggests that Jiuquanornis is the most basal member among the four Changma ornithuromorph taxa and potentially is basal within Ornithuromorpha.
The holotype material of Jiuquanornis was originally considered too incomplete to justify the erection of a new taxon [5]; however, most Cretaceous fossil bird specimens from other regions in the world that have been used to erect new species over the past few years are even more fragmentary: the enantiornithine Bauxitornis mindszentyae from Hungary is based on a right tarsometatarsus [32]; the enantiornithine Intiornis inexpectatus from Argentina is based on a right foot [33]; and the ornithuromorph Alamitornis minutus also from Argentina is based on the proximal third of a left humerus [34]. The three specimens from Changma are all more complete than these species and can be distinguished morphologically from other Mesozoic ornithuromorphs, thus we justify erecting a new taxon for each specimen.
Numerous fossils belonging to Ornithuromorpha and Enantiornithes have been recovered from the Aptian (Lower Cretaceous) Xiagou Formation in the Changma Basin, although no basal, non-ornithothoracine pygostylians have ever been found. Gansus yumenensis was the first avian species recovered from the Changma Basin [1], resolved as an advanced non-ornithurine ornithuromorph or basal ornithurine [2,5,10,12,29,35]. With the addition of Yumenornis, Changmaornis, and Jiuquanornis, a total of four named ornithuromorph taxa have been reported now [1,2,5].
Enantiornithines also form an important component of the Changma avifauna, with one named taxon, Qiliania graffini, and three unnamed taxa [6-9]. Although being less numerous than ornithuromorphs in overall quantity of specimens, enantiornithines formed a greater portion of the taxonomic diversity until the discovery of the three new ornithuromorphs described here [5,6,8,9]. Thus, the hypothesis that the Changma avifauna, which is younger than the Jehol avifauna, may represent an initial stage in the domination of Ornithuromorpha over Enantiornithes both in numbers and taxonomic diversity, is supported here [2,5].
Body mass in Changma birds
In order to further explore morphological differences between the specimens described here and Gansus, as well as between Changma ornithuromorphs and enantiornithines, we estimated body mass for five specimens of Gansus, the holotype specimens of Yumenornis and Changmaornis, and six enantiornithine specimens (including two specimens of Qiliania, two reported unnamed specimens, and two undescribed specimens). We estimated the body mass based on Liu et al. [36]; however, since no limb bones are preserved in the holotype of Jiuquanornis, this taxon could not be included. The selected specimens of Gansus are the most complete and best preserved; however, some of them still lack forelimb or hind limb elements. The humeral length and the tibiotarsal width were measured (Table 3). According to Liu et al. [36], humeral length gives a better estimate of body mass for arboreal birds, while tibiotarsal width is more appropriate for cursorial birds. The width was measured as close to the midpoint as possible in all the measured tibiotarsi.
Table 3. Estimated body masses of fossil birds from Changma.
| Species | Humeral length (mm) | Tibiotarsal width (mm) | Estimated weight (g) |
|---|---|---|---|
| Ornithuromorphs | |||
| Gansus yumenensis | |||
| CAGS-IG-04-CM-001 | ? | 2.5 | ?/124.8 |
| CAGS-IG-04-CM-002 | ? | 2.7 | ?/147.4 |
| CAGS-IG-04-CM-003 | 48.5 | 3.0 | 144.2/185.0 |
| CAGS-IG-04-CM-004 | 48.0 | ? | 141.7/? |
| GSGM-05-CM-014 | 47.8 | 2.7 | 140.6/147.4 |
| Mean weight | 142.2/142.9 | ||
| Yumenornis huangi | 49.9 | ? | 151.5/? |
| Changmaornis houi | ? | 2.9 | ?/172.0 |
| Enantiornithines | |||
| Qiliania graffini | |||
| GSGM-05-CM-006 | ? | 1.3 | ?/30.4 |
| GSGM-04-CM-006 | ? | 1.31 | ?/30.9 |
| Mean weight | ?/30.7 | ||
| CAGS-IG-04-CM-007 | ? | 1.7 | ?/54.3 |
| CAGS-IG-02-0901 | 47.7 | ? | 140.1/? |
| GSGM-05-CM-068 | ? | 1.9 | ?/69.0 |
| GSGM-05-CM-012 | ? | 2.0 | ?/77.0 |
Ornithuromorphs show morphological indicators of a terrestrial lifestyle, while the inferred ecology of basal birds and enantiornithines trends more closely towards arboreal [36-38]. Gansus is interpreted as amphibious, potentially a shallow-diving bird [2,4,39], suggesting tibiotarsal width would be a more accurate indicator of body mass. However, the body mass estimates based on each of the two measures are nearly the same, averaging 142.2 g (humeral length) and 142.9 g (tibiotarsal width). Changmaornis has a smaller foot than Gansus, but potentially it has a slightly greater body mass than most specimens of Gansus even though the two ornithuromorphs may have been similar in size. Although lacking hind limb material, Yumenornis exhibits a more robust humerus than that of Gansus, and it is estimated to have a greater body mass than Gansus. Yumenornis is estimated to be larger than both Gansus and Changmaornis (Table 3).
The enantiornithine specimens from Changma that preserve hind limb material all possess long and highly curved pedal unguals, indicative of an arboreal lifestyle [40]. Unfortunately, the absence of preserved forelimb material prevents the most accurate body mass estimates for these specimens [36]. Body mass estimates for Changma enantiornithines are much lower than those for ornithuromorphs (Table 3), consistent with the small and delicate morphology of the preserved material. A similar trend is observed in the Jehol avifauna [36].
The proximal position of the metatarsal II trochlea and elongate proximal phalanges of the pedal digits of Changmaornis are similar to that of Gansus, Hesperornis, loons, and grebes [2], thus it is possible that Changmaornis was also aquatic. The holotype specimens of Yumenornis and Jiuquanornis are too incomplete to determine their ecology. However, the potential close relationship between Yumenornis and Yixianornis suggests Yumenornis may have had a more terrestrial lifestyle. The morphology of the sternum in Jiuquanornis, with short body and long trabeculae bordering deep incisures, convergently occurs in terrestrial, largely cursorial galliforms and tinamids [5,41]. Only the discovery of more complete materials can clarify these ecological inferences. Comparatively, Changma enantiornithines are considered arboreal. Similar ecological differences are also observed between Enantiornithes and Ornithuromorpha in other regions, especially in The Jehol Group [10,24,36-38,42,43]. Coupled with differences in body mass between these clades, we suggest niche partitioning had strongly developed between Enantiornithes and Ornithuromorpha by the Early Cretaceous.
Supporting Information
Character states for Yumenornis huangi, Changmaornis houi, and Jiuquanornis niui used in the phylogenetic analysis in this study.
(DOC)
Acknowledgments
We thank the personnel of the former Fossil Research and Development Center, Third Geology and Mineral Resources Exploration Academy, Gansu Provincial Bureau of Geo-Exploration and Mineral Development (Lanzhou, China) for discovering and preparing the specimens. We also thank Dr. Andrew Farke for handling this manuscript, and Drs. Gareth Dyke and Federico Agnolin for careful and critical comments.
Funding Statement
This research was supported by the Hundred Talents Project of the Chinese Academy of Sciences, the National Natural Science Foundation of China (40672007,41072019), and the Department of Land and Resources of Gansu Province to LDQ. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hou L, Liu Z (1984) A new fossil bird from Lower Cretaceous of Gansu and early evolution of birds. Sci Sin B 27: 1296-1301. [Google Scholar]
- 2. You HL, Lamanna MC, Harris JD, Chiappe LM, O'Connor J et al. (2006) A nearly modern amphibious bird from the Early Cretaceous of northwestern China. Science 312: 1640-1643. doi: 10.1126/science.1126377. PubMed: 16778053. [DOI] [PubMed] [Google Scholar]
- 3. Ji S, Ji Q, You H, Lü J, Yuan C (2006) Webbed foot of an Early Cretaceous ornithurine bird Gansus from China. Geological Bulletins China 25: 1296-1298. [Google Scholar]
- 4. Li Y, Zhang Y, Zhou Z, Li Z, Liu D et al. (2011) New material of Gansus and a discussion on its habit. Vertebrata PalAsiatica 49: 435-445. [Google Scholar]
- 5. You H, Atterholt J, O'Connor JK, Harris JD, Lamanna MC et al. (2010) A second Cretaceous Ornithuromorph bird from the Changma Basin, Gansu Province, northwestern China. Acta Palaeontol Pol 55: 617-625. doi: 10.4202/app.2009.0095. [DOI] [Google Scholar]
- 6. Ji S, Atterholt J, O'Connor JK, Lamanna MC, Harris JD et al. (2011) A new, three-dimensionally preserved enantiornithine bird (Aves: Ornithothoraces) from Gansu Province, north-western China. Zool J Linn Soc 162: 201-219. doi: 10.1111/j.1096-3642.2010.00671.x. [DOI] [Google Scholar]
- 7. You H, O' Connor JK, Chiappe LM, Ji Q (2005) A new fossil bird from the Early Cretaceous of Gansu Province, northwestern China. Hist Biol 17: 7-14. doi: 10.1080/08912960500284851. [DOI] [Google Scholar]
- 8. Harris JD, Lamanna MC, You H, Ji S, Ji Q (2006) A second enantiornithean (Aves: Ornithothoraces) wing from the Early Cretaceous Xiagou Formation near Changma, Gansu Province, People's Republic of China. Can J Earth Sci 43: 547-554. doi: 10.1139/e06-007. [DOI] [Google Scholar]
- 9. Lamanna MC, Li D, Harris JD, Atterholt J, You H (2006) Exceptionally-preserved Enantiornithes (Aves: Ornithothoraces) from the Early Cretaceous of northwestern China. Acta Palaeontol Pol 51: 423-434. [Google Scholar]
- 10. Zhou S, Zhou Z, O' Connor JK (2013) Anatomy of the basal ornithuromorph bird Archaeorhynchus spathula from the Early Cretaceous of Liaoning, China. J Vertebr Paleontol 33: 142-152. [Google Scholar]
- 11. Baumel JJ, Witmer LM (1993) Osteologia. In: Baumel JJ, King AS, Breazile JE, Evans HE, Berge JCV. Handbook of Avian Anatomy: Nomina Anatomica Avium, second Edition. Cambridge: Nuttall Ornithological Club; pp. 45-132. [Google Scholar]
- 12. O’Connor JK, Zhou Z (2012) A redescription of Chaoyangia beishanensis (Aves) and a comprehensive phylogeny of Mesozoic birds. J Syst Palaeontol: 1-18. [Google Scholar]
- 13. Goloboff PA, Farris JS, Nixon KC (2008) TNT, a free program for phylogenetic analysis. Cladistics 24: 774-786. doi: 10.1111/j.1096-0031.2008.00217.x. [DOI] [Google Scholar]
- 14. Suarez BM, Ludvigson AG, González AL, Al-Suwaidi HA, You H (2013) Stable isotope chemostratigraphy in lacustrine strata of the Xiagou Formation, Gansu Province, NW China. In: Bojar AV, Melinte-Dobrinescu MC, Smit J. Isotopic Studies in Cretaceous Research Geological Society, London: Special Publications; , 382. First published online 10.1144/sp382.1. [DOI] [Google Scholar]
- 15. Linnaeus C (1758) Systems Naturae. 10th edition; Salvii Laurentii Holmiae. 824 p [Google Scholar]
- 16. Chiappe LM (2002) Basal bird phylogeny problems and solutions. In: Chiappe LM, Witmer LM. Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. [Google Scholar]
- 17. Chiappe LM (1995) The phylogenetic position of the Cretaceous birds of Argentina: Enantiornithes and Patagopteryx deferrariisi . In: Peters DS. Proceedings of the 3rd Symposium of the Society of Avian Paleontology and Evolution. Courier Forschungsinstitut; Senckenberg: 181,Frankfurt am Main: . pp. 55-63 [Google Scholar]
- 18. Zhou Z, Zhang F, Li Z (2009) A new basal ornithurine bird (Jianchangornis microdonta gen. et sp. nov.) from the lower Cretaceous of China. Vertebrata PalAsiatica 47: 299-310. [Google Scholar]
- 19. Zhou Z, Zhang F (2001) Two new ornithurine birds from the Early Cretaceous of western Liaoning, China. Chin Sci Bull 46: 1-7. doi: 10.1007/BF02900665. [DOI] [Google Scholar]
- 20. Clarke JA, Zhou Z, Zhang F (2006) Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui . J Anat 208: 287-308. doi: 10.1111/j.1469-7580.2006.00534.x. PubMed: 16533313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Hou L (1997) Mesozoic Birds of China: Feng huang ku Bird; Park, Taiwan. [Google Scholar]
- 22. Kurochkin EN (1982) New order of birds from the Lower Cretaceous in Mongolia. Palaeontol J: 1982: 215-218. [Google Scholar]
- 23. O’Connor JK, Gao KQ, Chiappe LM (2010) A new ornithuromorph (Aves: Ornithothoraces) bird from the Jehol Group indicative of higher-level diversity. J Vertebr Paleontol 30: 311-321. doi: 10.1080/02724631003617498. [DOI] [Google Scholar]
- 24. Clarke JA, Norell MA (2002) The morphology and phylogenetic position of Apsaravis ukhaana from the Late Cretaceous of Mongolia. Am Museum Nov 3387: 1-46. doi: 10.1206/0003-0082(2002)387. [DOI] [Google Scholar]
- 25. Zhou Z, Zhang F (2005) Discovery of an ornithurine bird and its implication for Early Cretaceous avian radiation. Proc Natl Acad Sci U S A 102: 18998-19002. doi: 10.1073/pnas.0507106102. PubMed: 16344487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Clarke JA (2004) Morphology, phylogenetic taxonomy, and systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). Bull Am Museum Nat Hist 286: 1-179. doi: 10.1206/0003-0090(2004)286. [DOI] [Google Scholar]
- 27. Zhou Z, Zhang F (2006) A beaked basal ornithurine bird (Aves, Ornithurae) from the Lower Cretaceous of China. Zool Scripta 35: 363-373. doi: 10.1111/j.1463-6409.2006.00234.x. [DOI] [Google Scholar]
- 28. O' Connor JK (2009) A systematic review of Enantiornithes (Aves:Ornithothoraces). Los Angeles: University of Southern California. 600 pp. [Google Scholar]
- 29. O’Connor JK, Chiappe LM, Bell A (2011) Pre-modern birds: avian divergences in the Mesozoic. In: Dyke GD, Kaiser G. Living Dinosaurs: the Evolutionary History of Birds. New Jersey: J. Wiley & Sons. [Google Scholar]
- 30. Zhou Z, Clarke JA, Zhang F (2002) Archaeoraptor's better half: the other component of this infamous fossil forgery is identified as a fish-eating bird. Nature 420: 285. doi: 10.1038/420285a. PubMed: 12447431. [DOI] [PubMed] [Google Scholar]
- 31. Zhou S, Zhou Z, O' Connor JK (2012) A new toothless ornithurine bird (Schizooura lii gen. et sp. nov.) from the Lower Cretaceous of China. Vertebrata Palasiatica 50: 9-24. [Google Scholar]
- 32. Dyke GJ, Ősi A (2010) A review of Late Cretaceous fossil birds from Hungary. Geol J 45: 434-444. [Google Scholar]
- 33. Novas FE, Agnolín FL, Scanferla CA (2010) New enantiornithine bird (Aves, Ornithothoraces) from the Late Cretaceous of NW Argentina. Comp R Palévol 9: 499-503. [Google Scholar]
- 34. Agnolin FLM, Martinelli AG (2009) Fossil birds from the Late Cretaceous Los Alamitos Formation, Río Negro Province, Argentina. J South American Earth Sci 27: 42-49. doi: 10.1016/j.jsames.2008.09.003. [DOI] [Google Scholar]
- 35. O' Connor JK, Wang X, Chiappe LM, Gao C, Meng Q et al. (2009) Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species. J Vertebr Paleontol 29: 188-204. doi: 10.1080/02724634.2009.10010371. [DOI] [Google Scholar]
- 36. Liu D, Zhou Z, Zhang Y (2012) Mass estimate and evolutionary trend in Chinese Mesozoic fossil birds. Vertebrata PalAsiatica 50: 39-52. [Google Scholar]
- 37. Zhang Y, Tian X, Li Z (2008) Hindlimb bone proportions of Chinese Mesozoic birds and their implications for perching habits. Vertebrata PalAsiatica 46: 317-329. [Google Scholar]
- 38. Morschhauser E, Varricchio DJ, Gao C, Liu J, Wang X et al. (2009) Anatomy of the Early Cretaceous bird Rapaxavis pani, a new species from Liaoning Province, China. J Vertebr Paleontol 29: 545-554. doi: 10.1671/039.029.0210. [DOI] [Google Scholar]
- 39. Nudds RL, Atterholt J, Wang X, Dyke GJ, You H (2013) Locomotory abilities and habitat of the Cretaceous bird Gansus yumenensis inferred from limb length proportions. J Evol Biol 26: 150-154. doi: 10.1111/jeb.12036. PubMed: 23194019. [DOI] [PubMed] [Google Scholar]
- 40. Hopson J (2001) Ecomorphy of avian and nonavian theropod phalangeal proportions: implications for the arboreal versus terrestrial origin of bird flight. In: New Perspectives on the Origin and Evolution of Birds: Proceedings of the International Symposium in honor of John H. Ostrom. Peabody Museum of Natural History, Yale University; . pp. 211-235 [Google Scholar]
- 41. Feduccia A (1995) Explosive evolution in Tertiary birds and mammals. Science 267: 637-638. doi: 10.1126/science.267.5198.637. PubMed: 17745839. [DOI] [PubMed] [Google Scholar]
- 42. Zhou Z, Clarke J, Zhang F, Wings O (2004) Gastroliths in Yanornis: an indication of the earliest radical diet-switching and gizzard plasticity in the lineage leading to living birds? Naturwissenschaften 91: 571-574. doi: 10.1007/s00114-004-0567-z. PubMed: 15452699. [DOI] [PubMed] [Google Scholar]
- 43. Martin LD (1983) The origin of birds and of avian flight. In: Johnston RF. Current Ornithology Vol.4 New York: Plenum Press. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Character states for Yumenornis huangi, Changmaornis houi, and Jiuquanornis niui used in the phylogenetic analysis in this study.
(DOC)