

Abstract
The primary goal of this review is to highlight current research and theories describing the neurobiological basis of math (MD), reading (RD), and comorbid math and reading disability (MD+RD). We first describe the unique brain and cognitive processes involved in acquisition of math and reading skills, emphasizing similarities and differences in each domain. Next we review functional imaging studies of MD and RD in children, integrating relevant theories from experimental psychology and cognitive neuroscience to characterize the functional neuroanatomy of cognitive dysfunction in MD and RD. We then review recent research on the anatomical correlates of MD and RD. Converging evidence from morphometry and tractography studies are presented to highlight distinct patterns of white matter pathways which are disrupted in MD and RD. Finally, we examine how the intersection of MD and RD provides a unique opportunity to clarify the unique and shared brain systems which adversely impact learning and skill acquisition in MD and RD, and point out important areas for future work on comorbid learning disabilities.
Keywords: comorbidity, neurobiological, learning disabilities
Understanding the neural bases of reading and math disabilities has been a topic of great interest to clinical and basic researchers for many years. Recent advances in functional and structural neuroimaging have provided a window into a more comprehensive understanding of impaired brain function underlying math and reading disabilities (MD and RD, respectively). Converging behavioral findings suggest that MD and RD originate from unique cognitive mechanisms. In the case of RD, it is widely thought that a phonological deficit precludes normal reading acquisition (S. E. Shaywitz & Shaywitz, 2005) whereas MD is thought to originate from a core deficit in manipulation of quantity (Price, Holloway, Rasanen, Vesterinen, & Ansari, 2007). However, multiple cognitive processes are shared between reading and mathematics, including the representation and retrieval of symbolic information, attention, working memory, and cognitive control. Therefore, impairments in any one of these domain-general skills could conceivably play an important role in both pure and comorbid conditions—a topic that we address at length below.
Here our central goals are to (a) provide a concise review of extant findings describing the neural bases of MD and RD in relation to prominent cognitive theories of each disorder, (b) examine the extent to which these developmental disorders share common neural substrates, and (c) provide an integrated synthesis of these domain-specific results as they pertain to comorbidity of MD and RD (MD+RD). Specifically, we examine results from the functional magnetic resonance imaging (fMRI) literature, which describe abnormal brain activity in MD and RD populations when performing specific tasks related to reading and mathematics, as well as structural MRI, which describes impairments in the integrity of gray and white matter of the brain associated with MD and RD. A limitation of the body of learning disabilities (LD) work is that the literatures describing MD and RD have evolved largely independent of one another. Influential models of each disorder have, therefore, failed to account for the high levels of co-occurrence of these two disorders in many children and adults with LD. We argue that a more thorough understanding of the similarities and differences in the brain bases of these disorders is necessary not only for understanding comorbidity for these disorders but also for providing critical new information to aid in characterizing each of these disorders individually. We continue by highlighting areas of future research on comorbid MD and RD with the goal of better understanding the neurocognitive processes underlying subtypes of LDs.
A major effort of this work is to consolidate domain-specific knowledge from the MD and RD literatures as a means to build a framework for considering the neuroscientific basis of comorbid MD+RD. The motivation for this effort is that there is a dearth of research describing the neurobiological foundation of MD+RD, and an important step for progressing and promoting future research in this area is to generate a number of testable hypotheses regarding potential brain bases for this comorbidity. To this end, we propose a theoretical model to describe differential pathways for comorbidity, which includes three possible routes to comorbidity: (a) an additive model in which comorbidiy reflects the cumulative effects of impairment that characterize MD and RD, (b) a verbally mediated model in which comorbidity originates in impairments in phonological systems associated with RD language impairments, and (c) a domain-general model in which the comorbid condition originates from domain-general impairments in brain systems, notably the prefrontal cortex, that support both math and reading (Figure 1)—a topic that we address at length below. At the end of this review we expand on this model, including specifics of the neural loci associated with each of these pathways to comorbidity.
Figure 1.

Neurobiological pathways underlying math and reading disability (MD+RD) comorbidity. Schematic illustration of putative neurobiological pathways underlying MD+RD. (A) The “additive” pathway for comorbidity posits that pure RD and MD originate from unique cognitive and brain impairments, and the comorbid (“combined”) condition is the additive effect of impairment to brain systems that characterize MD and RD. (B) The “verbally mediated” pathway for comorbidity arises from the close link between symbolic mapping, fact retrieval, and word-based problem solving on one hand and verbally mediated processes on the other. In this case, comorbidity originates in impairments to phonological systems associated with RD. (C) The “domain-general” pathway for comorbidity originates from attention, working memory, and sequencing impairments such as those mediated by the prefrontal cortex (VLPFC and DLPFC). Again, this model states that pure RD and MD originate from unique cognitive and brain impairments. DLPFC = dorsolateral PFC; FG = fusiform gyrus; IPS = intraparietal sulcus; MTG = middle temporal gyrus; PFC = prefrontal cortex; SPL = superior parietal lobule; VLPFC = ventrolateral PFC.
Neurobiology of MD
Overview
The overarching goals of the MD (which includes developmental dyscalculia) sections of the review are to highlight the primary perceptual and cognitive processes involved in developing numerical abilities, to introduce the main cognitive theories that explain MD, and to connect theory with important neurobiological aspects of atypical development in MD. We review central theories of MD, both those that are domain-specific processes involving nonsymbolic and symbolic representations of quantity as well as domain-general processes such as working memory and attention. We specify the core brain areas and functional circuits involved in numerical cognition and examine how they are affected in MD, during tasks that range in complexity from basic symbolic and nonsymbolic quantity representation to complex problem solving. We then examine evidence for anatomical deficits in MD, from the perspective that anatomical studies provide an opportunity to examine core neurobiological deficits without confounds associated with task difficulty.
As with behavioral studies, it should be noted that our understanding of the underlying neurobiology of MD is inherently limited by criteria used to define the disorder, which tend to vary considerably with respect to diagnostic measures, cutoff scores, criteria for comorbidity, exclusion criteria for study participants, and composition of control groups. Despite these limitations, neuroimaging studies have provided novel insights into the nature of brain and cognitive processes disrupted in MD.
Cognitive theories of MD
Four hypotheses grounded in cognitive theory have been proposed to characterize core deficits of MD. One hypothesis describes MD as a core deficit in processing quantity (Butterworth, Varma, & Laurillard, 2011) and number sense (Piazza et al., 2010; Wilson & Dehaene, 2007), the inability to make judgments about quantity and to reason with symbolic representations of quantity (Butterworth et al., 2011). Consistent with this hypothesis, it has been demonstrated that, compared to typically developing (TD) children, children with MD have lower than expected abilities in quantity estimation (Mazzocco, Feigenson, & Halberda, 2011; Piazza et al., 2010) and abnormal magnitude representations (Ashkenazi, Mark-Zigdon, & Henik, 2009; Mussolin, Mejias, & Noël, 2010).
The second hypothesis describes MD as a weakness in automatically mapping symbols to their internal magnitude representations, reflecting a specific impairment in symbolic processing that does not affect nonsymbolic processing (Rousselle & Noël, 2007; Rubinsten & Henik, 2005). In line with this hypothesis, individuals with MD have difficulties in the comparison of numbers but not the length of sticks (Rousselle & Noël, 2007) and have weakness in the automatic association between number and quantities but intact automatic association between size and quantities (Rubinsten & Henik, 2005).
The third hypothesis posits MD as a domain-general phenomenon involving working memory (Rotzer et al., 2009; Toll, Van der Ven, Kroesbergen, & Van Luit, 2011) and attention (Ashkenazi & Henik, 2010a, 2010b), rather than as a specific deficit in number processing. Consistent with the domain-general hypothesis, individuals with MD often have deficits in the use of developmentally appropriate arithmetic procedures, impairments that have been attributed to weaknesses in working memory rather than to specific deficits in number sense (Geary, 2004). In further support of a domain-general hypothesis, a meta-analysis examining 28 studies of children with MD indicated that the MD groups consistently demonstrated visuospatial and verbal working memory deficits (Swanson, Howard, & Saez, 2006).
A fourth “hybrid” hypothesis explains MD as an impairment both in representing and manipulating numerical magnitude on an internal number line and in working memory and attention, cognitive processes that are not specific to number processing. This hypothesis further states that deficits in any of these nonmathematical processes should be reflected not only in impaired math skills but also in poor reading and learning abilities. Of importance, this hypothesis predicts subgroups of MD that present isolated difficulties in number sense as well as difficulties with word problems. Consistent with this model, it has been suggested that two thirds of children with MD have comorbid MD+RD, whereas one third have “pure” MD (von Aster & Shalev, 2007). This hypothesis is most useful when considering comorbid conditions that represent an important and significant aspect of MD.
General math circuit
Multiple distributed brain structures are known to be important for mathematical cognition (Table 1, Figure 2). Converging evidence from studies in infants (Feigenson, Dehaene, & Spelke, 2004; Xu & Carey, 1996; Xu & Spelke, 2000), preschool children (Cantlon, Brannon, Carter, & Pelphrey, 2006), and adults (Ansari, 2008; Rosenberg-Lee, Chang, Young, Wu, & Menon, 2011), as well as nonhuman primates (Cantlon & Brannon, 2006), indicates that the representation of approximate quantities is supported by the bilateral intraparietal sulcus (IPS) in the dorsal aspects of the posterior parietal cortex (PPC). For this analysis, the IPS was divided to distinct subdivisions: hIP2 on the anterior lateral bank of the IPS, hIP1, which is posterior to hIP2, and hIP3, which is posterior and medial to both hIP1 and hIP3 (Uddin et al., 2010; Figure 2). In addition to the dorsal PPC, the ventral occipitotemporal cortex (VOT) also plays an important, though underappreciated, role in number processing. A recent meta-analysis found that within the VOT, the left fusiform gyrus (FG; Table 1) is consistently activated across a wide range of numerical tasks (Arsalidou & Taylor, 2011), consistent with its hypothesized role in processing orthographic structure (Binder, Medler, Westbury, Liebenthal, & Buchanan, 2006). Beyond this, more complex calculation abilities place demands on multiple cognitive systems involving visuospatial working memory and executive control functions (Menon, Rivera, White, Glover, & Reiss, 2000). In such situations, multiple prefrontal cortex (PFC) areas are also consistently activated during numerical processing. This includes the bilateral inferior frontal gyrus (IFG), which is engaged by tasks that involve effortful maintenance and retrieval, and the bilateral middle frontal gyrus (MFG), which is engaged by tasks that require manipulation of information in working memory during multistage calculation tasks (Menon et al., 2000; Menon, Mackenzie, Rivera, & Reiss, 2002; Zago et al., 2008). Finally, the bilateral anterior cingulate cortex, which is involved in error and conflict monitoring (Kerns et al., 2004), has also been found to be consistently activated across several numerical and arithmetic tasks in both children (Davis et al., 2009; De Smedt & Boets, 2010; Rivera, Reiss, Eckert, & Menon, 2005) and adults (Arsalidou & Taylor, 2011). In addition, canonical “language” areas, including the left temporoparietal cortex (encompassing the angular gyrus, supramarginal gyrus, and posterior temporal cortex) and the left IFG (Table 1), are engaged during retrieval of well-rehearsed arithmetic facts (e.g., multiplication tables; Dehaene, Piazza, Pinel, & Cohen, 2003; Delazer et al., 2003; Prado et al., 2011; Rosenberg-Lee, Chang, et al., 2011).
Table 1.
Neurobiology of MD and RD: Summary of Key Findings With Selected References.
| Area/location: coronal horizontal sagittal | Brain region/function | MD | RD |
|---|---|---|---|
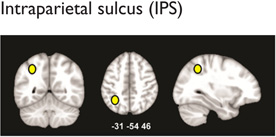 |
↓ Right IPS: nonsymbolic processing (Price, Holloway, Rasanen, Vesterinen, & Ansari, 2007) | ✓ | |
| ↓ Right IPS: symbolic comparison (Mussolin, De Volder, et al., 2010) | ✓ | ||
| ↓ Right IPS: arithmetic (Ashkenazi, Rosenberg-Lee, Tenison, & Menon, 2012) | ✓ | ||
| ↓ Right IPS: (reduced gray matter volume; Rotzer et al., 2008; Rykhlevskaia, Uddin, Kondos, & Menon, 2009) | ✓ | ||
| ↑Left IPS: non-symbolic processing (Kaufmann et al., 2009) | ✓ | ||
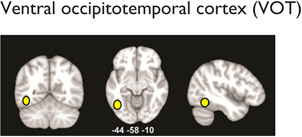 |
↓ Right VOT: nonsymbolic number comparison (Price et al., 2007) | ✓ | |
| ↓Bilateral VOT: arithmetic problem solving (Ashkenazi et al., 2012) | ✓ | ||
| ↓ Left VOT: reduced gray matter volumes (Silani et al., 2005) | ✓ | ||
| ↓ Left VOT: in prereaders with family history (Raschle, Chang, & Gaab, 2011) | ✓ | ||
| ↓ Left VOT: (VWFA) visual processing and word identification (Schlaggar & McCandliss, 2007) | ✓ | ||
| ↓ Left VOT: (cortical circuitry in occipitotemporal regions) learning to read, mapping between orthography and phonology (Dehaene & Cohen, 2011; Maisog, Einbinder, Flowers, Turkeltaub, & Eden, 2008; Richlan, Kronbichler, & Wimmer, 2011; Schlaggar & McCandliss, 2007; Wandell, Rauschecker, & Yeatman, 2012) | ✓ | ||
| ↓ Left VOT: connectivity to left inferior frontal and IPL (van der Mark et al., 2011) | ✓ | ||
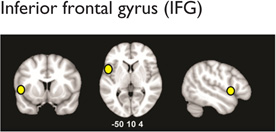 |
↓ Left IFG: arithmetic problem solving in children(Ashkenazi et al., 2012) | ✓ | |
| ↑ Bilateral IFG: possible compensatory mechanism: age-related increases in activation (B. A. Shaywitz et al., 2002), treatment response (Farris et al., 2011), and reading outcome prediction (Hoeft et al., 2011) | ✓ | ||
| ↓Left IFG (Maisog et al., 2008; Richlan, Kronbichler, & Wimmer, 2009) | ✓ | ||
| ↓Left IFG reduced gray matter volumes (Eckert et al., 2003; Laycock at al., 2008) | ✓ | ||
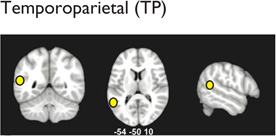 |
↓Left STG and MTG: phonological/processing speech sounds (Golestani, Molko, Dehaene, LeBihan, & Pallier, 2007) | ✓ | |
| ↓Left TP: IPL and posterior STG, phonological storage/retrieval (Vigneau et al., 2006) | ✓ | ||
| ↓Left TP: impaired connectivity between angular gyrus and posterior language system (Horwitz, Rumsey, & Donohue, 1998) | ✓ | ||
| ↓Bilateral TP: reduced gray matter (Hoeft et al., 2007; Richlan, Kronbichler, & Wimmer, in press) | ✓ | ||
| ↓Bilateral TP: reduced gray matter/surface area in prereaders with greater (maternal) history of RD (Black et al., 2012; Raschle et al., 2011) and activation (Raschle et al., 2011) | ✓ | ||
| ↑ Right TP: possible compensatory mechanism (Ligges, Ungureanu, Ligges, Blanz, & Witte, 2010) | ✓ |
Note. IPL = inferior parietal lobule; MD = math disability; MTG = middle temporal gyrus; RD = reading disability; STG = superior temporal gyrus; VWFA = visual word form area.
Figure 2.

Core math-related functional circuits. The intra-parietal sulcus (IPS) is a critical processing region in the brain for math and magnitude-related functions, and fronto-parietal circuits have been identified using resting-state functional connectivity analysis of the IPS. (A) IPS region-of -interests (ROIs) derived from cytoarchitectonic maps for the three sub-divisions of the IPS: hIP2 is the lateral and anterior sub-division of the IPS (blue); hIP1 is the sub-division located posterior to hIP2 (green); hIP3 is the posterior sub-division of the IPS (red). (B) Functional connectivity maps associated with hIP1, hIP2, and hIP3. The color code represents voxels correlated with each source ROI. The IPS has significant connectivity with distributed frontal (MFG and PMC) and parietal (SPL, and SMG) cortical regions in both hemispheres. Additional functional circuits associated with the ventral-occipital temporal cortex are not shown, but see Figure 3. Adapted from Uddin et al., 2010. hIP: human intra parietal; MFG: Middle frontal gyrus, PMC: PreMotor cortex, SMG: Supramarginal gyrus, SPL: Superior parietal lobule.
Although there are similarities in brain areas engaged by numerical tasks in adults and children, there are key differences as well. In particular, with experience and learning, there is decreased dependence on the PFC and greater reliance on multiple regions of the PPC, including the left IPS (Ansari & Dhital, 2006; Cantlon et al., 2006; Rivera et al., 2005). Furthermore, even the seemingly brief one-year interval spanning Grades 2 and 3 is characterized by significant task-related changes in brain response and connectivity, a finding that suggests that pooling data across wide age ranges and grades can miss important neurodevelopmental changes (Rosenberg-Lee, Barth, & Menon, 2011). Specifically, over the long term, there is a shift from more controlled and effortful to more automatic processing of both numerical magnitude and arithmetic problem solving (Ansari & Dhital, 2006; Cantlon et al., 2009; Rivera et al., 2005); however, recent evidence suggests that over the short term there is a process of continued refinement of brain responses. Moreover, as demonstrated by recent multivariate analyses, similar levels of signal level cannot be conflated with similar types of information processing (Blair, Rosenberg-Lee, Tsang, Schwartz, & Menon, 2011).
Functional neuroimaging studies of number processing deficits in MD
One prominent theory of MD describes it as a core deficit in processing numerical quantity (Butterworth et al., 2011). Several neuroimaging studies of MD have therefore focused on tasks involving symbolic and nonsymbolic number comparison with small and large numerical distance (Cohen Kadosh et al., 2007; Kaufmann, Vogel, Starke, Kremser, & Schocke, 2009; Kucian, Loenneker, Martin, & von Aster, 2011; Mussolin, De Volder, et al., 2010; Price et al., 2007). The choice of these tasks is based on the landmark behavioral research of Moyer and Landauer (1967), who showed that adults are significantly slower and more error prone when they compare numbers with a smaller distance (e.g., 1 and 2) compared to a larger distance (e.g., 1 and 7; Moyer & Landauer, 1967). In the context of neurobiological studies, this allows for the contrast of two task conditions that are closely matched on several perceptual, cognitive, and response factors, differing only in the complexity of magnitude judgments.
A number of brain regions have been implicated in math-related deficits in children and adults with MD. The IPS is particularly sensitive to manipulations of numerical distance in both children and adults. Aberrant IPS responses to numerical distance have been found in several studies of MD (Figure 3; Cohen Kadosh et al., 2007; Kaufmann, Vogel, Starke, Kremser, & Schocke, 2009; Mussolin, De Volder, et al., 2010; Price et al., 2007). Specifically, reduced activation of the right IPS has been shown for nonsymbolic (Figure 3A; Price et al., 2007) as well as symbolic number comparison tasks (Figure 3B; Mussolin, De Volder, et al., 2010). Decreased IPS activity has also been shown in MD during a numerical ordering task (Kucian, Grond, et al., 2011). Despite this consistency, a few studies have also found evidence for increased IPS activity, suggesting compensatory mechanisms in children with MD (Kaufmann, Vogel, Starke, Kremser, Schocke, & Wood, 2009). Finally, novel support for impaired right IPS function in MD has also been provided by a transcranial magnetic stimulation study. In this study, neural function in the right IPS was temporarily disrupted by transcranial magnetic stimulation in normal adult participants, which caused a temporary magnitude processing deficit similar to those seen in individuals with dyscalculia (Cohen Kadosh et al., 2007).
Figure 3.

Aberrant functional brain responses during numercial tasks in MD. Brain regions that show decreased activation in MD relative to TD during (A) non-symbolic comparsion (Price et al. 2007), (B) symbolic comparison (Mussolin et al. 2010), and (C) an arithmetic problem solving task (Ashkenazi et al. 2012). Task paradigms are presented on the left in panels A, B, and C, with numerically “easy” tasks on top and numerically “difficult” tasks on the bottom. All studies in panels A through C found reduced activity in children with MD related to the difficulty of the tasks. (D) Summary of anatomical locations of brain regions highlighted in panels A through C. FG: Fusiform gyrus, IFG: Inferior frontal gyrus, IPS: Intraparietal sulcus, LOC: lateral occipital cortex; MFG: Middle frontal gyrus, MTG: Middle temporal gyrus, SPL: Superior parietal lobule.
In addition to decreased activity in the IPS, individuals with MD also show atypical activity in VOT, superior parietal lobule (SPL), and PFC during basic number comparisons tasks (Table 1; Figure 3B). In a study involving symbolic number comparison, children with MD showed decreased activation for numerical distance in the right MFG and the right middle cingulate gyrus (Mussolin, De Volder, et al., 2010). Children with MD also show reduced numerical distance effects in the left FG and left MFG (Price et al., 2007) during nonsymbolic number comparisons (Figure 3A). However, a third study that parametrically varied the numerical distance effect during nonsymbolic comparison found evidence for compensatory activity in the right FG and bilateral supplementary motor area, but not the PPC, in the MD group (Kucian, Loenneker, Martin, & von Aster, 2011). Despite the small number of comparison studies to date, these results indicate that children with MD show abnormal activity in multiple brain areas that consistently extends beyond the IPS into multiple PFC and inferior-temporal regions. In studies that focus on numerical distance effects, with appropriate control tasks, findings are most consistently in support of deficits in the IPS, FG, and the MFG. Given the foundational importance of basic number sense in MD, further studies are needed to further clarify the direction of deficits and their relation to poor performance and skill acquisition. A further issue is that overemphasis on the IPS has distracted from a proper consideration of the role of other brain regions in MD. Given that the IPS has strong interconnectivity with the MFG and premotor cortex (PMC; Figure 2), the contribution of these and other brain regions to impaired number processing in MD warrants further investigation.
Functional neuroimaging studies of arithmetic processing deficits in MD
Poor fluency in retrieval of arithmetic facts is one of the most prominent difficulties in children with MD (Geary, 2004). TD children, typically during the second and third grades, show a rapid shift in the distribution of strategies toward greater use of direct retrieval in solving simple arithmetic problems (Ashcraft & Fierman, 1982; Carpenter & Moser, 1984; Geary, 1994; Siegler, 1996). In contrast, children with MD demonstrate poorer performance and continue to use less mature strategies, such as finger counting, to solve arithmetic problems (Geary, 1993; Shalev, Auerbach, Manor, & Gross-Tsur, 2000). Only a handful of brain imaging studies has examined the neural basis of arithmetic deficits in MD. Similar to the emphasis placed on the IPS for basic numerical processing, aberrant activity in this region has also been implicated as the primary deficit underlying atypical development of arithmetic problem solving in individuals with MD (Butterworth et al., 2011).
Three studies in children with MD have all reported atypical functional activation in IPS and PMC/MFG regions of the PFC during arithmetic problem solving (Ashkenazi, Rosenberg-Lee, Tenison, & Menon, 2012; Davis et al., 2009; Kucian et al., 2006), although the direction of effects has not always been consistent. Kucian and olleagues (2006) examined 18 children with MD from third and sixth grades and found no group differences between MDs and TDs at the whole-brain level. Region-of-interest analyses, however, revealed reduced brain activity in the right IPS, IFG, and MFG. These differences were found for approximate, but not exact, addition problems (Kucian et al., 2006). In contrast, Davis and colleagues (2009) used these same tasks and found increased PPC and PFC activation during both exact and approximate addition in third grade children with MD (Davis et al., 2009). Ashkenazi and colleagues examined univariate and multivariate brain responses to addition problems that differed only in arithmetic complexity in a group of 7- to 9-year-old (grades 2 and 3) children with MD (Ashkenazi et al., 2012). They found that although the TD group showed strong modulation of brain responses with increasing arithmetic complexity, children with MD failed to show such modulation (Figure 3C). Children with MD showed significantly reduced activation compared to TD children in the IPS and SPL in the dorsal PPC, supramarginal gyrus in the ventral PPC, and bilateral MFG as a function of arithmetic complexity. In addition to univariate analyses, they also examined multivoxel activation patterns using representational similarity analysis, which is a multivariate approach for investigating the relationship between stimulus representation and neural activity (Norman, Polyn, Detre, & Haxby, 2006). It examines the spatial pattern of multivoxel brain activity in a specific region of interest across task conditions, independent of overall differences in signal level. They used similarity analysis to probe the similarity of spatial activation patterns between different types of arithmetic problems and provided complementary information about problem representation in children with MD. Critically, multivariate analyses revealed that brain response patterns to complex and simple problems were less differentiated in the MD group in bilateral IPS, independent of overall differences in signal level. Taken together, these results show that children with MD not only underactivate key brain regions implicated in mathematical cognition but also fail to generate distinct neural responses and representations for different arithmetic problems.
Morphometry and tractography in MD
One potential reason for discrepancies in neuroimaging studies of MD is that the profile of functional deficits in mathematical task processing varies with the level of task difficulty and type of operation performed. A complementary approach to the systematic investigation of MD is to examine whether there are neuroanatomical and structural brain differences relative to well-matched TD controls. White matter and gray matter integrity are crucial for nearly all higher cognitive operations (Johansen-Berg & Behrens, 2006), and systematic identification of anatomical deficits can provide more concrete evidence for core neurobiological deficits in MD. Here we discuss findings from studies of brain morphometry, which use structural neuroanatomical data to describe local differences in gray and white matter density and volume and tractography, which describes the integrity of white matter tracts that connect distal and proximal brain regions (Rykhlevskaia, Uddin, Kondos, & Menon, 2009).
In one voxel-based morphometric study, 12 children with MD (mean age of 9 years) were compared to IQ-matched controls, and it was shown that those with MD had decreased gray matter volume in the right IPS, left IFG, bilateral MFG, and anterior cingulate cortex, and also showed white matter volume reductions in right parahippocampal gyrus (Rotzer et al., 2008). Rykhlevskaia and colleagues examined gray and white matter in a group of 7- to 9-year-old children with MD, in comparison with children matched on age, gender, intelligence, reading abilities, and working memory capacity (Figure 4; Rykhlevskaia et al., 2009). They found multiple loci of structural abnormalities in children with MD, including reduced gray matter volume in the right IPS, FG, parahippocampal gyrus, and right anterior temporal cortex and bilateral hippocampus (Figure 4A). Furthermore, deficiencies in right hemisphere micro-structure and long-range white matter projection fibers linking the right FG with temporoparietal cortex were also deficient in MD (Figure 4B). Analysis of fiber tracts also points to deficits in multiple long-range white matter tracts including the inferior fronto-occipital fasciculus and inferior and superior longitudinal fasciculus (Figure 4C). Although this neuroanatomical evidence is still emerging, these results point to macro- and micro-structural abnormalities in right hemisphere temporal-parietal white matter, and pathways associated with it, as key neuroanatomical correlates of MD (Molko et al., 2003; Rotzer et al., 2008; Rykhlevskaia et al., 2009).
Figure 4.

Neuroanatomical substrates of MD. Neuroanatomical studies have shown that children with MD have reduced (A) grey matter volume in bilateral SPL, right IPS, lingual and fusiform gyri, MTL, and hippocampus, (B) white matter volume in right temporal-parietal cortex, and (C) lower fiber density in inferior longitudinal fasciculus, inferior fronto-occipital fasciculus, and caudal forceps major compared to typically developing children. The right panel of (C) shows reduced connectivity in children with MD for long-range white matter projection fibers that link the right fusiform gyrus with temporal-parietal areas (Adapted from Rykhlevskaia et al. 2009). IPS: Intraparietal sulcus, MTL: Middle temporal lobule, SPL: Superior parietal lobule. VBM: Voxel-based morphometry
Summary
Converging evidence from both structural and multiple functional imaging studies, involving a wide range of numerical tasks, points to the right IPS as a major locus of deficits in children and adults with MD. However, the findings reviewed above sum to highlight the importance of several additional brain areas and suggest that the model of IPS deficiency as the unitary source for deficits in MD is a gross oversimplification. Although the role of brain regions beyond the IPS has often been deemphasized in many previous studies, it is now increasingly evident that individuals with MD show deficits in a distributed, but interconnected, set of brain regions that includes the bilateral IPS and FG in posterior brain regions and the PMC/MFG regions of the PFC. Network analysis further suggests the possibility of multiple dysfunctional circuits arising from a core white matter deficit and leads to the testable hypothesis that MD may, at its core, be a disconnection syndrome. In TD children, the role of these areas varies according to the developmental stage and the nature of the numerical task performed, with more mature function characterized by decreased engagement of the PFC and increased engagement of posterior brain areas. The extent to which children with MD show systematic variation in patterns of brain responses and connectivity as a function of age, instruction, and intervention is at present unclear and is an important topic of ongoing research.
Neurobiology of RD
Overview
The overarching goals of the RD (also referred to as developmental dyslexia) sections of the review are to highlight the primary cognitive tasks and skills required for fluent reading, to ground the discussion in theory, and to connect theory to important neurobiological aspects of both typical and atypical reading trajectories. We begin by defining RD and then describe both the primary methods for diagnosis and RD prevalence rates. We move to a review of the neurobiological basis of typical reading, addressing both the general reading circuit and specific components of reading including orthographic, phonological, and semantic processing. We then review central theories of RD, including domain-general (such as attention) and domain-specific (such as phonological processing) models, and map the various theories onto neuroimaging evidence. We conclude with a review of neuroimaging findings in RD, pointing out overlap and distinctions with brain areas implicated in MD (Table 1). It should be noted, however, that the review of both theory and the underlying neurobiology of RD is inherently circumscribed for the following reasons. First, there are variable diagnostic criteria for RD, in terms of both specific assessment measures and the differing usage of cutoff scores for diagnosis. Furthermore, RD is examined in multiple countries and language settings, and languages vary greatly along a spectrum based on the extent to which orthography maps clearly onto phonology. This transparency – opaqueness spectrum may influence the manifestation of RD (e.g., accuracy vs. processing speed). Studies also vary in their exclusion criteria for comorbidity (and RD is often concomitant with other disorders such as MD and attention-deficit/ hyperactivity disorder) and are often guided by or seek to evaluate different theories of RD.
Cognitive theories of RD
Skilled reading relies on the integration of orthographic (understanding the written form of language), phonological (knowledge of sound structure), and semantic processing (being able to elicit meaning from oral or written words). There are several preconditions to fluent reading, including accurate vision, speech understanding, and efficiency in both hearing and manipulating speech sounds. Next, children need to learn to match orthography (written symbols) and phonology; deficiency in single word reading beyond what is expected because of cognitive capability is called RD.
Though many factors can account for individual differences in reading ability in childhood, it is widely hypothesized that RD is neurobiological in origin (Lyon, Shaywitz, & Shaywitz, 2003) and reflects an underlying weakness in language processing that is specific to phonological awareness, defined as the ability to identify and manipulate phonemes (basic sound structures; Wagner & Torgesen, 1987). Phonological awareness is associated with later reading skills such as orthographic awareness and comprehension (Torgesen, Wagner, Rashotte, Burgess, & Hecht, 1997), and the theory that RD is fundamentally rooted in a deficit in phonological processing (as opposed to, for example, solely visual deficits) is referred to as the phonological core deficit model (Stanovich, 1994). Naming speed deficits, however, are also evident in individuals with RD (Bowers, Steffy, & Tate, 1988). Although there is ample behavioral evidence supporting a deficit in naming speed in RD, some researchers contend that the phonological processing deficit is independent of naming speed (Wolf, 1991), leading to the double-deficit hypothesis (Wolf & Bowers, 1999). According to the double-deficit hypothesis, individuals with deficits in both naming speed and phonological processing have more severe deficits in reading than those with single deficits (i.e., impaired naming speed or phonological processing) or without any deficits (Wolf & Bowers, 1999). In fact, there are many domain-specific hypotheses based on phonological awareness deficits, including the rapid auditory processing hypothesis, which suggests a causal relationship between impaired auditory temporal processing for rapid acoustic stimuli and the ability to accurately discriminate speech sounds (Tallal, 2004).
Another hypothesis, called the temporal sampling framework, places impaired phonology at the heart of RD and links the sensory and phonological deficits found in the disorder to neurobiological models of speech processing (Poeppel, Idsardi, & van Wassenhove, 2008). According to the theory, the primary deficit of RD is impaired processing of slower temporal modulations in speech (Abrams, Nicol, Zecker, & Kraus, 2009), at relatively low levels of the auditory system (Goswami, 2011). Still other theorists point out the importance of the role of both visual processing and visual attention in reading, and the potential impact on visual perception of graphemes, its translation to phonemes (and thereby phonemic awareness), and possible impairment in RD (Vidyasagar & Pammer, 2010). The magnocellular (or M-deficit) hypothesis suggests that for individuals with RD the magnocellular layers of the lateral geniculate nucleus of thalamus develop abnormally, leading to both binocular vision instability and reduced motion sensitivity, both of which impair visual perception (R. Laycock & Crewther, 2008; Stein, 2001). In essence, letters may seem to move, which negatively affects orthographic processing (Stein, 2001). More recent hypotheses extend this to other sensory domains such as to audition.
Although the aforementioned hypotheses are domain-specific, there are also hypotheses of RD that are more domain-general and thus may influence other cognitive impairments and disorders beyond RD. For example, one hypothesis highlights the role of working memory and attentional systems on fluent reading and suggests impairments in RD for word-form storage and processing, linked processing of orthographic and phonological information, and rapid and automatic attentional switching (i.e., executive function components; Berninger, Raskind, Richards, Abbott, & Stock, 2008; S. E. Shaywitz & Shaywitz, 2008) as with MD (Geary, 2004; Hecht, Torgesen, Wagner, & Rashotte, 2001; Rotzer et al., 2009). A more domain-general hypothesis (the anchoring hypothesis) suggests that the phonological, visual, and/or auditory impairments seen in RD may indicate a general impairment in perception, which may negatively affect short-term memory (Ahissar, 2007). Specifically, this hypothesis suggests that compared to nonimpaired readers, those with RD do not benefit from repeated incoming stimuli because they are unable to automatically attend to the “parameters” of speech. Another theory, called the cerebellar deficit theory, states that those with RD suffer from an impairment of automaticity, which negatively affects language function and reading (Nicolson & Fawcett, 2009).
General reading circuit
Fluent reading depends on the ability to recognize single words rapidly, and functional neuroimaging studies have helped to elucidate the brain regions and pathways important for skilled reading (Wandell, Rauschecker, & Yeatman, 2012). Focus on the orthographic (ventral) systems and the connection to the phonological (dorsal) systems including the left IFG and temporoparietal region (inferior parietal lobule [IPL] posterior superior temporal gyrus [pSTG]) are important for fluent reading (Schlaggar & McCandliss, 2007; Figures 5A, 5B). Extant findings also suggest that the circuits in the VOT cortex (of which the visual word form area [VWFA] and FG are key subdivisions), develop over time and these circuits’ response profiles change during the time that children learn to read (Wandell et al., 2012; Figures 5A, 5B). The visual orthographic regions are located in the extrastriate cortex, and bilateral regions are presumed to support processing of visual input into the VWFA (Schlaggar & McCandliss, 2007). It is important to note that activity in the VFWA may represent a bottom-up encoding of orthography of letter strings, yet may also be influenced by top-down attention processes related to the connection between phonological and semantic systems (Schlaggar & McCandliss, 2007). For fluent reading to occur, letters and sounds must be integrated. Specifically, mappings between orthography and phonology are required (such as letter sound integration), and studies suggest the importance of cortical circuitry involving the occipital and temporal regions, such as the left lateral occipitotemporal sulcus (VWFA), left superior temporal cortex, left ventral occipitotemporal, and left occipitoparietal cortex (Dehaene & Cohen, 2011; Schlaggar & McCandliss, 2007). Recent intrinsic functional connectivity studies, which identify correlated fMRI activity between spatially distributed brain regions measured during a period of rest (i.e., no task), further support the importance of these pathways and demonstrate increased coupling between FG and PPC as well as IFG (Broca’s) regions (Koyama et al., 2011; Figure 5B).
Figure 5.

Core reading-related functional circuits. (A) Surface rendering of 730 activation peaks reported in two meta-analyses of reading (Koyama et al., 2011; Vigneau et al., 2006). Each activation peak is color-coded according to its contrast category: phonology (blue), semantic (red), and syntax (green). Results of meta-analyses show a concentration of peaks in ventral occipito-temporal cortex (VOT), temporo-parietal region, inferior frontal cortex, and premotor/motor regions with overlapping representation for phonology, semantic and syntax processing. (B) Major reading-related functional circuit identified using resting-state functional connectivity with a seed (region of interest, ROI) placed in the fusiform gyrus (FG) within the VOT and Visual Word Form Area (VWFA). Connectivity from FG to the left inferior parietal lobule (IPL)/supramarginal gyrus (SMG) and pars opercularis (IFGop) of the left inferior frontal gyrus showed positive association with reading measures in typical adult readers (MNI coordinates derived from Koyama et al., 2011).
Functional neuroimaging findings in RD
A large number of functional neuroimaging studies of reading have been performed over the past decade in children and adults with RD. Recent meta-analyses of these studies point to a consistent pattern of deficits in a distributed left hemisphere network that includes the left VOT, STG, middle temporal gyrus (MTG), IPL (supramarginal gyrus), and IFG (Maisog, Einbinder, Flowers, Turkeltaub, & Eden, 2008; Richlan, Kronbichler, & Wimmer, 2009; Figure 6A and Table 1). Deficits in the VOT region encompassing the VWFA point to abnormal visual processing and automatic identification of words (Schlaggar & McCandliss, 2007). Atypical brain responses in the STG and MTG (Figure 6A), on the other hand, point to aberrant phonological processing of the sounds of spoken language as well as deficits in rapid processing of speech sounds (Golestani, Molko, Dehaene, LeBihan, & Pallier, 2007), whereas deficits in the temporoparietal cortex (including the IPL [SMG] and posterior STG; Figure 6A) point to impairments in phonological storage and retrieval (Vigneau et al., 2006). This pattern of deficits is consistent with perceptual and cognitive theories, as described in the “General Reading Circuit” section above, which suggest that impaired mapping of orthography onto the phonologic structure of the language system is a core feature of RD (Booth et al., 2002; Cao, Bitan, Chou, Burman, & Booth, 2006; Newman & Joanisse, 2011). Anomalous function in these areas likely compromises the phonological processes essential for learning to read, and this pattern of deficits appears to persist from childhood to adulthood in RD (Richlan et al., 2009).
Figure 6.

Aberrant functional brain activation during reading tasks in RD. (A) Brain regions that show under- and over-activation during fMRI tasks in RD adults and children compared to controls are color-coded (MNI coordinates taken from (Maisog, Einbinder, Flowers, Turkeltaub, & Eden, 2008; Richlan, Kronbichler, & Wimmer, 2009)). (B) Cold colors indicate brain regions that show reduced activation during a phonological processing fMRI task in RD compared to chronological age-matched and reading level-matched typical readers (i.e., RD-specific regions). Warm colors indicate brain regions that show increased activation in RD compared to age-matched but not reading-matched controls, likely reflecting compensatory mechanisms (Hoeft et al., 2007). (C) Connectivity analysis using direct causal modeling showed left FG to IPL and left FG/IPL to IFG connectivity deficits in RD (Cao, Bitan, & Booth, 2008). FG: Fusiform gyrus, IFGop: Inferior frontal gyrus, pars opercularis, IFGtr: IFG pars triangularis, IPL: Inferior parietal lobule, SMG: Supramarginal gyrus, STG: Superior temporal gyrus, VOT: Ventral occipito-temporal cortex, VWFA: Visual word form area.
In addition to posterior brain areas, the IFG and insula have also been implicated in RD. In fluent reading, the left- hemisphere pars triangularis and opercularis of the IFG (IFGtr and IFGop, respectively) are important for articulation and naming (Fiez & Petersen, 1998). Results from fMRI studies in RD have been mixed and have shown both hypo- and hyper-activation in IFG. IFG has been implicated in compensatory mechanisms (Hoeft et al., 2007; Figure 6B), a hypothesis further supported by age-related increases in activation (Hoeft et al., 2007; B. A. Shaywitz et al., 2002), greater functional connectivity in treatment responders versus nonresponders (Farris et al., 2011), and a study in which reading outcomes predicted IFG activation in dyslexia (Hoeft et al., 2011). Meta-analyses, however, challenged these studies and reported hypo- rather than hyper-activation in the left IFG in RD (Maisog et al., 2008; Richlan et al., 2009), with hyperactivation only in the left (Richlan et al., 2009) or right insula (Maisog et al., 2008) rather than the IFG. More anatomically precise studies of the IFG and frontoinsular cortex are needed to clarify the differential role of these PFC regions in reading, along the lines of a recent study in numerical cognition (Supekar & Menon, 2012).
RD has also been considered a disconnection syndrome, a hypothesis first suggested by Geschwind (1965). Supporting this hypothesis, many neuroimaging studies have reported abnormal left frontoparietal (Cao, Bitan, & Booth, 2008; Quaglino et al., 2008), temporoparietal, and VOT networks in RD (Cao et al., 2008; Horwitz, Rumsey, & Donohue, 1998; van der Mark et al., 2011; Figure 6C). Compensatory networks have been identified in the IFG, and in right hemisphere regions homologous to the left posterior brain system (Ligges, Ungureanu, Ligges, Blanz, & Witte, 2010). Another characteristic that has been proposed is that individuals with RD have less efficient functional circuits (Casanova, El-Baz, Giedd, Rumsey, & Switala, 2010; Dhar, Been, Minderaa, & Althaus, 2010; Ligges et al., 2010; Vourkas et al., 2011). Preliminary evidence, however, suggests that abnormal connectivity in RD can normalize with appropriate intervention (Richards & Berninger, 2008), suggesting that these abnormalities may be state rather than trait markers.
Finally, electrophysiological evidence has provided complementary evidence for temporal processing deficits in RD. Notably, RD impairments in both rapid and slow auditory temporal processing have been reported, and these deficits may contribute to a wide range of perceptual and phonological impairments seen in RD (Abrams et al., 2009; Abrams, Nicol, Zecker, & Kraus, 2006).
Taken together, these results show that children with RD underactivate key brain regions and underrecruit key functional networks implicated in phonological, orthographic, and semantic processing. Of interest, meta-analyses suggest greatest evidence in the left VOT (Maisog et al., 2008; Richlan, Kronbichler, & Wimmer, 2011). Whether this is because of core deficits in RD or differences resulting from task, control task, and processing methods is still unclear. There is evidence for compensatory mechanisms, as reflected by overactivation, although the pathways involved are still unclear.
Morphometry and tractography in RD
Structural MRI studies generally support findings from fMRI studies (Jobard, Crivello, & Tzourio-Mazoyer, 2003), and show reduced gray matter volumes in left temporoparietal (Hoeft et al., 2007), VOT (Silani et al., 2005), left IFG, and cerebellum (Eckert et al., 2003; S. K. Laycock et al., 2008), though a recent meta-analysis that included a small number of studies found consistent RD-related deficits only in the bilateral STG and adjoining temporoparietal cortex (Richlan, Kronbichler, & Wimmer, in press; Figure 7A). Although the cerebellum is implicated in RD and in rapid naming, this region has been largely absent in fMRI studies, possibly because studies often exclude part or most of the cerebellum when scanning because of physical constraints. Although the majority of structural imaging studies have been conducted with adolescents and adults, evidence of anatomical abnormalities in prereaders, identified as at risk through examining family history, largely overlap and converge with the RD literature (Black et al., 2012; Raschle, Chang, & Gaab, 2011). Finally, diffusion tensor imaging studies have highlighted the importance of the arcuate fasciculus, superior longitudinal fasciculus, inferior fronto-occipital fasciculus, optic radiation, and callosal fibers in reading, and abnormalities of these tracts have been reported in RD (Frye et al., 2011; Wandell et al., 2012), as illustrated in Figure 7B. Of interest, with intervention both fMRI activation patterns and white matter pathways in the left anterior centrum semiovale have been shown to be reversible in RD (Keller & Just, 2009).
Figure 7.

Neuroanatomical substrates of RD. (A) Neuroanatomical studies have shown that children with RD have reduced grey matter (volume, density, and surface area) in left IFG, temporo-parietal region including IPL and superior temporal gyrus (STG), ventral occipitotemporal region (VOT) and cerebellum. Clusters in bilateral temporo-parietal region/ STG shown in orange have consistently reduced grey matter in RD compared to controls as reported in a recent meta-analysis (Richlan et al., in press). (B) White matter pathways that carry essential reading signals. The two images show several major white matter fascicles in the left hemisphere from different points of view. Three of these fascicles communicate information to and from the occipital lobe. The red ellipsoids are located in cortical regions activated in fMRI tasks of reading (Jobard, Crivello, & Tzourio-Mazoyer, 2003) (Reproduced with permission from (Wandell, Rauschecker, & Yeatman, 2012)). These pathways have been reported to be aberrant in RD in previous neuroimaging studies (except for the optic radiation) (Frye et al., 2011). IFGop: Inferior frontal gyrus, pars opercularis, IFGtr: IFG, pars triangularis, IPL: Inferior parietal lobule, MTG / STG: Middle temporal gyrus / superior temporal gyrus, SMG: Supramarginal gyrus, VWFA: Visual word form area.
Although still preliminary, as in the field of MD, these results point to macro- and micro-structural abnormalities in white matter pathways linking inferior frontoparietal, temporoparietal, and VOT regions, as well as corresponding gray matter regions, as key neuroanatomical correlates of RD. Important questions remain, however, and prompt future research into studies that examine brain regions or pathways that are more causal versus compensatory and those that reflect trait versus state markers.
Summary
Converging evidence from numerous functional and structural imaging studies underscores multiple pathways that underlie orthographic, phonological, and semantic processes that are impaired in RD. In sum it appears that the left hemisphere regions centered in the FG, temporoparietal cortex, and the IFG as well as their functional connectivity and associated white matter pathways are impaired in RD. The role of other regions such as the right IFG as well as subcortical and cerebellar regions is still controversial, perhaps because of variation in fMRI tasks, imaging modality, and samples used. Although it is clear that phonological deficits underlie RD, it is also apparent that there are many other deficits and factors involved in impaired reading, such as auditory-visual integration and executive function. Therefore, an important next step for RD research is to identify neural signatures for various subtypes of RD.
Neurobiology of MD+RD
As noted above, incidence rates of comorbid MD+RD can be as high as 50% (Lewis, Hitch, & Walker, 1994). Several behavioral studies have focused on whether MD and RD arise from independent factors. Converging evidence from these studies indicates that core deficits in RD arise primarily from phonological deficits whereas MD arises from deficits in nonsymbolic and symbolic representation of quantity (Landerl, Fussenegger, Moll, & Willburger, 2009; Rubinsten & Henik, 2006; van der Sluis, de Jong, & Leij, 2004; Willburger, Fussenegger, Moll, Wood, & Landerl, 2008). Most of these studies have focused on tasks that involve mapping between symbols and their internal magnitude representation or the ability to associate letters with phonemes. Beyond this, emerging behavioral research points to several potential loci of reading-related deficits that can affect math abilities. First, poor reading can impair basic phonological representation and mapping of numerical quantity (Zebian & Ansari, 2012). Second, arithmetic problem solving in number-word format is a particular source of difficulty for children with MD+RD (Compton, Fuchs, Fuchs, Lambert, & Hamlett, 2012; Fuchs et al., 2010; Fuchs & Fuchs, 2002; Jordan, Hanich, & Kaplan, 2003; Powell & Fuchs, 2010; Powell, Fuchs, Fuchs, Cirino, & Fletcher, 2009a, 2009b). Third, deficits in phonological awareness are also associated with difficulties in memorizing and retrieving basic arithmetic facts from long-term memory (De Smedt, Taylor, Archibald, & Ansari, 2010; Geary, 2004), especially when verbal retrieval is the optimal strategy (Boets & De Smedt, 2010).
To our knowledge, there has been no neuroscientific investigation of MD+RD. However, one case study using a 17-year-old male with a head injury resulting in a right parietal skull fracture and right temporal hemorrhage found evidence for both MD and RD despite normal intellectual function (Levin, 1996). Although such case studies have the potential to provide novel information about MD+RD, the nature of injury and specific compensatory mechanisms that accompany them precludes inferences regarding the larger MD+RD population.
Very little is currently known about differences in anatomical structure and connectivity in children with MD and MD+RD, when compared to TD children. Of importance, there have been no functional or structural neuroimaging studies of MD+RD reported in children or adults. Therefore, we focus here on the unique and shared processes underlying MD and RD and provide suggestions for future research on the neurobiology of MD+RD based on the known cognitive and neural processes involved in each disorder.
At the level of processing of symbolic and nonsymbolic magnitudes, the neurobiology of MD and RD presented above is largely consistent with the proposal that these disorders are associated with two independent cognitive deficits, namely a core phonological deficit in the case of RD and a deficit in number sense in the case of MD (Landerl et al., 2009). The distinct neurocognitive profiles are reflected in the differential role of the left ventral fronto-temporalparietal language circuits (IFG, STG/MTG, and IPL) in RD, and the predominantly right lateralized dorsal frontoparietal circuits (PMC/MFG and IPS/SPL) in MD. Yet this pattern of largely independent representations begins to overlap when we consider tasks involving verbally mediated fact retrieval. Indeed, at the level of arithmetic fact retrieval, new behavioral studies are beginning to suggest that adults with RD retrieve fewer arithmetic facts from memory and are less efficient in doing so (De Smedt & Boets, 2010). Phonological processing, particularly phonological awareness, is related to fact retrieval, and this association appears to be especially prominent in operations such as multiplication that rely more on fact retrieval, when compared to operations such as subtraction. Although there have been no neuroimaging studies of math fact retrieval in children or adults with RD, one recent study in TD adults suggested that multiplication elicits greater activity than subtraction in regions involved in verbal processing, including MTG and IFG areas that have been implicated in RD (Prado et al., 2011).
Even more prominent overlap with left STG/MTG and IFG systems involved in phonological and language processing is likely for more complex word problems that place stronger demands on verbal working memory. Consistent with this view, behavioral studies have shown that, when compared with children with MD, children with MD+RD show similar abilities on tasks involving basic numerical processing, but they show profound impairments in the solution of word problems (Jordan et al., 2003). Comparative neurobiological studies in MD, RD, and MD+RD groups can therefore help further clarify how phonological and other language deficits contribute to deficits in more complex problem-solving situations. Beyond this, studies in all three subtypes of LD also need to characterize how brain and cognitive processes in each domain interact, and the task conditions under which such interactions contribute synergistically to weak learning and problem-solving skills in children with comorbid MD+RD.
Although RD and MD are commonly represented as unique brain-based disorders, there is a high level of comorbidity for these disorders, and there is growing evidence that multiple, shared cognitive processes are required for reading and arithmetic (Houdé, Rossi, Lubin, & Joliot, 2010). These shared processes include executive function, attention, and working and semantic memory, in addition to the fundamental role of verbal processes inherent to arithmetic learning. Of importance, there have been no studies that have examined differences in neural function in participants with MD+RD in comparison to “pure” MD and RD conditions, which are necessary to describe the neural signature of the cognitive deficits in MD+RD. We propose three possible outcomes that could result from a study involving these three groups (i.e., MD, RD, and MD+RD; see Figure 1): First, the comorbid condition involves additive impairments for both reading and mathematics, both of which are included in the neural markers for “pure” RD and MD (Figure 1A). Second, the neural basis of the comorbid condition involves brain-based impairments that underlie verbal processes that preclude both normal reading and arithmetic development (Figure 1B). This hypothesis is based on the known association between verbal and calculation abilities (Prado et al., 2011). Third, the neural basis of MD+RD is a result of impairments of shared neural resources necessary for both reading and mathematics, and may include executive function and working memory. Deficits in prefrontal brain regions, including both VLPFC and DLPFC, may be related to these impaired, shared neural resources (Figure 1C).
Conclusions
It is not surprising that the literatures describing the cognitive and neural bases of reading and mathematics have evolved largely independent of one another; there are vast differences between the fundamental cognitive processes involved in these two skills, and, given this divide, making meaningful connections between these two domains is not straightforward. From a neuroscientific perspective, there are a number of gross differences between reading and mathematics: (a) reading and mathematics appear to rely on neural activity in different hemispheres, with reading primarily activating left-hemisphere structures and mathematics relying on predominantly right-hemisphere structures; (b) different regions of the parietal lobe appear to be important for domain-specific aspects of reading and mathematics, with ventral IPL regions (supramarginal and angular gyri) being closely associated with reading-related processes whereas the IPS appears to be critical for performing arithmetic calculations; and (c) a distributed network involving left-hemisphere IPL and IFG appears to play a fundamental role in many reading-related processes, whereas mathematics does not appear to engage this network. Therefore, from a theoretical perspective, it might be argued that simultaneously considering the neuroscientific bases of math and reading is not a useful endeavor.
From a practical perspective, however, understanding the cognitive and neuroscientific relationships between reading and mathematical skills represents an essential and unexplored question in the extant literature. In support of this statement, the comorbidity rates of MD and RD are remarkably high, suggesting that overlapping neural systems may affect performance in both reading and arithmetic domains in individuals with MD+RD. Therefore, despite the significant neurocognitive differences between reading and mathematics, we argue that the intersection of MD and RD provides a unique opportunity to clarify the unique and shared brain systems that adversely affect learning and skill acquisition in these populations. Furthermore, a more comprehensive understanding of the cognitive and neurobiological factors that contribute to high levels of comorbidity will inform methods for remediating deficits in some of the most academically challenged students in our educational systems.
In this review, we have described both the cognitive theories and neuroscientific evidence for MD, RD, and MD+RD, and it is important to consider how the brain-based data have informed our understanding of the behavioral impairments in these populations. With regard to both MD and RD, it is widely acknowledged that a phonological or quantity processing deficits represents core cognitive impairments in MD and RD, respectively; however, there are many other theories that link basic domain-specific and domain-general functions in each disorder. Brain-based data have not yet unambiguously clarified the cognitive basis of MD or RD since several different cognitive theories described here have been supported by neuroimaging results. This raises the possibility that MD and RD both represent highly heterogeneous and/or systemic impairment, which affects multiple domain-specific and domain-general brain systems, notions that have not always been embraced in either the MD or RD research communities. Nevertheless, a strong case can be made for heterogeneity: Skilled math and reading both rely on the confluence of a host of basic sensory and cognitive functions, and it is plausible that impairments in any of these functions could result in impaired skills in both domains. It is hoped that future study designs will target multiple, competing hypotheses using large cohorts of subjects as a means of addressing known heterogeneities in these populations and bringing clarity to the many cognitive theories of MD and RD. Such an endeavor takes on added significance when we consider the high levels of comorbidity between MD and RD and the critical need to address reading and problem-solving skills in a largely neglected group of individuals.
Our review has underscored that mathematical and reading tasks involve both domain-general and domain-specific brain networks, and there is evidence that many of these circuits have shown impairments in MD and RD. Furthermore, we have learned that the parietal cortex appears to be a critical processing station for both reading and mathematical domains, further supporting an essential role of this brain region for high-level integration and computation. Finally, we have learned that the PFC plays a significant role in attention and working memory in both domains, and impairments in PFC function may underlie deficits associated with MD-RD comorbidity. Building on these findings, we hope that future research will explore the neuroscientific bases of MD+RD; neuroimaging studies provide important objective information about the etiology and subsequent course of MD and RD independently, as well as the comorbidity of the two disorders. Specifically, if there is a neural signature for RD and for MD (and MD+RD), then brain-based identification, which is invariant to environment and diagnostic criteria (such as cutoff scores), could provide an important clinical and diagnostic tool.
Advanced neuroimaging techniques may also aid in disentangling the multiple theories purported to underlie the disorders, especially in dissociating the domain-general theories (such as working memory and attention) from the domain-specific theories underlying MD and RD. It is now widely accepted that cognitive functions, such as arithmetic processing, are served by networks of brain regions distributed across the cortex (Supekar & Menon, 2012). Therefore, understanding the integrity of the underlying anatomy of these regions, and the strength of connections between nodes of these brain networks, is critical for understanding the neural basis of normal and impaired arithmetic function. Multivariate approaches to understanding aberrant stimulus representations underlying different subtypes of LD are likely to contribute new information about developmental LD, with important practical implications for educational neuroscience (Ashkenazi et al., 2012). Finally, an important advancement in the field of LD research and practice is the use of the response to intervention model. Understanding which groups of individuals respond better to specific targeted interventions has important clinical and educational significance and may further elucidate diagnostic criteria. In sum, advancements in experimental design and neuroimaging methods have the potential to substantially improve the way we define and diagnose MD, RD, and MD+RD. These advancements may also provide more effective pathways to prediction and intervention as a means of improving the likelihood that all children will be provided enhanced opportunities to succeed scholastically.
Acknowledgments
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by grants from the National Institutes of Health (HD047520, HD059205, K23HD054720, and F32DC010322).
Footnotes
Authors’ Note
Figures 2–7 appear in color in the online version of this article, available at http://journaloflearningdisabilities.sagepub.com.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
- Abrams DA, Nicol T, Zecker SG, Kraus N. Auditory brainstem timing predicts cerebral asymmetry for speech. Journal of Neuroscience. 2006;26(43):11131–11137. doi: 10.1523/JNEUROSCI.2744-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abrams DA, Nicol T, Zecker S, Kraus N. Abnormal cortical processing of the syllable rate of speech in poor readers. Journal of Neuroscience. 2009;29(24):7686–7693. doi: 10.1523/JNEUROSCI.5242-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahissar M. Dyslexia and the anchoring-deficit hypothesis. Trend in Cognitive Science. 2007;11(11):458–465. doi: 10.1016/j.tics.2007.08.015. [DOI] [PubMed] [Google Scholar]
- Ansari D. Effects of development and enculturation on number representation in the brain. Nature Review Neuroscience. 2008;9(4):278–291. doi: 10.1038/nrn2334. [DOI] [PubMed] [Google Scholar]
- Ansari D, Dhital B. Age-related changes in the activation of the intraparietal sulcus during nonsymbolic magnitude processing: An event-related functional magnetic resonance imaging study. Journal of Cognitive Neuroscience. 2006;18(11):1820–1828. doi: 10.1162/jocn.2006.18.11.1820. [DOI] [PubMed] [Google Scholar]
- Arsalidou M, Taylor MJ. Is 2+2=4? Meta-analyses of brain areas needed for numbers and calculations. Neuroimage. 2011;54(3):2382–2393. doi: 10.1016/j.neuroimage.2010.10.009. [DOI] [PubMed] [Google Scholar]
- Ashcraft MH, Fierman BA. Mental addition in third, fourth, and sixth graders. Journal of Experimental Child Psychology. 1982;33:216–234. [Google Scholar]
- Ashkenazi S, Henik A. Attentional networks in developmental dyscalculia. Behavioral and Brain Functions. 2010a;6(1):1–12. doi: 10.1186/1744-9081-6-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashkenazi S, Henik A. A disassociation between physical and mental number bisection in developmental dyscalculia. Neuropsychologia. 2010b;48(10):2861–2868. doi: 10.1016/j.neuropsychologia.2010.05.028. [DOI] [PubMed] [Google Scholar]
- Ashkenazi S, Mark-Zigdon N, Henik A. Numerical distance effect in developmental dyscalculia. Cognitive Development. 2009;24(4):387–400. [Google Scholar]
- Ashkenazi S, Rosenberg-Lee M, Tenison C, Menon V. Weak task-related modulation and stimulus representations during arithmetic problem solving in children with developmental dyscalculia. Developmental Cognitive Neuroscience. 2012;2(Suppl. 1[0]):S152–S166. doi: 10.1016/j.dcn.2011.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berninger VW, Raskind W, Richards T, Abbott R, Stock P. A multidisciplinary approach to understanding developmental dyslexia within working-memory architecture: Genotypes, phenotypes, brain, and instruction. Developmental Neuropsychology. 2008;33(6):707–744. doi: 10.1080/87565640802418662. [DOI] [PubMed] [Google Scholar]
- Binder JR, Medler DA, Westbury CF, Liebenthal E, Buchanan L. Tuning of the human left fusiform gyrus to sublexical orthographic structure. Neuroimage. 2006;33(2):739–748. doi: 10.1016/j.neuroimage.2006.06.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Black JM, Tanaka H, Stanley L, Nagamine M, Zakerani N, Thurston A, Hoeft F. Maternal history of reading difficulty is associated with reduced language-related gray matter in beginning readers. NeuroImage. 2012;59(3):3021–3032. doi: 10.1016/j.neuroimage.2011.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blair KP, Rosenberg-Lee M, Tsang JM, Schwartz DL, Menon V. Beyond natural numbers: Representation of negative numbers in the parietal cortex. Frontiers in Human Neuroscience. 2011;6:7. doi: 10.3389/fnhum.2012.00007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boets B, De Smedt B. Single-digit arithmetic in children with dyslexia. Dyslexia. 2010;16(2):183–191. doi: 10.1002/dys.403. [DOI] [PubMed] [Google Scholar]
- Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TB, Mesulam MM. Functional anatomy of intra- and cross-modal lexical tasks. Neuroimage. 2002;16(1):7–22. doi: 10.1006/nimg.2002.1081. [DOI] [PubMed] [Google Scholar]
- Bowers PG, Steffy R, Tate E. Comparison of the effects of IQ control methods on memory and naming speed predictors of reading disability. Reading Research Quarterly. 1988;23(3):304–319. [Google Scholar]
- Butterworth B, Varma S, Laurillard D. Dyscalculia: From brain to education. Science. 2011;332(6033):1049–1053. doi: 10.1126/science.1201536. [DOI] [PubMed] [Google Scholar]
- Cantlon JF, Brannon EM. Shared system for ordering small and large numbers in monkeys and humans. Psychological Science. 2006;17(5):401–406. doi: 10.1111/j.1467-9280.2006.01719.x. [DOI] [PubMed] [Google Scholar]
- Cantlon JF, Brannon EM, Carter EJ, Pelphrey KA. Functional imaging of numerical processing in adults and 4-y-old children. PLoS Biology. 2006;4(5):e125. doi: 10.1371/journal.pbio.0040125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantlon JF, Libertus ME, Pinel P, Dehaene S, Brannon EM, Pelphrey KA. The neural development of an abstract concept of number. Journal of Cognitive Neuroscience. 2009;21(11):2217–2229. doi: 10.1162/jocn.2008.21159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Bitan T, Booth JR. Effective brain connectivity in children with reading difficulties during phonological processing. Brain and Language. 2008;107(2):91–101. doi: 10.1016/j.bandl.2007.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao F, Bitan T, Chou TL, Burman DD, Booth JR. Deficient orthographic and phonological representations in children with dyslexia revealed by brain activation patterns. Journal of Child Psychology and Psychiatry. 2006;47(10):1041–1050. doi: 10.1111/j.1469-7610.2006.01684.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carpenter TP, Moser JM. The acquisition of addition and subtraction concepts in grades one through three. Journal for Research in Mathematics Education. 1984;15(3):179–202. [Google Scholar]
- Casanova MF, El-Baz AS, Giedd J, Rumsey JM, Switala AE. Increased white matter gyral depth in dyslexia: Implications for corticocortical connectivity. Journal of Autism and Developmental Disorders. 2010;40(1):21–29. doi: 10.1007/s10803-009-0817-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen Kadosh R, Cohen Kadosh K, Schuhmann T, Kaas A, Goebel R, Henik A, Sack AT. Virtual dyscalculia induced by parietal-lobe TMS impairs automatic magnitude processing. Current Biology. 2007;17(8):689–693. doi: 10.1016/j.cub.2007.02.056. [DOI] [PubMed] [Google Scholar]
- Compton DL, Fuchs LS, Fuchs D, Lambert W, Hamlett C. The cognitive and academic profiles of reading and mathematics learning disabilities. Journal of Learning Disabilities. 2012;45(1):79–95. doi: 10.1177/0022219410393012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis N, Cannistraci CJ, Rogers BP, Gatenby JC, Fuchs LS, Anderson AW, Gore JC. Aberrant functional activation in school age children at-risk for mathematical disability: A functional imaging study of simple arithmetic skill. Neuropsychologia. 2009;47(12):2470–2479. doi: 10.1016/j.neuropsychologia.2009.04.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Smedt B, Boets B. Phonological processing and arithmetic fact retrieval: Evidence from developmental dyslexia. Neuropsychologia. 2010;48(14):3973–3981. doi: 10.1016/j.neuropsychologia.2010.10.018. [DOI] [PubMed] [Google Scholar]
- De Smedt B, Taylor J, Archibald L, Ansari D. How is phonological processing related to individual differences in children’s arithmetic skills? Developmental Science. 2010;13(3):508–520. doi: 10.1111/j.1467-7687.2009.00897.x. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Cohen L. The unique role of the visual word form area in reading. Trends in Cognitive Sciences. 2011;15(6):254–262. doi: 10.1016/j.tics.2011.04.003. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Piazza M, Pinel P, Cohen L. Three parietal circuits for number processing. Cognitive Neuropsychology. 2003;20(3):487–506. doi: 10.1080/02643290244000239. [DOI] [PubMed] [Google Scholar]
- Delazer M, Domahs F, Bartha L, Brenneis C, Lochy A, Trieb T, Benke T. Learning complex arithmetic—An fMRI study. Cognitive Brain Research. 2003;18(1):76–88. doi: 10.1016/j.cogbrainres.2003.09.005. [DOI] [PubMed] [Google Scholar]
- Dhar M, Been PH, Minderaa RB, Althaus M. Reduced interhemispheric coherence in dyslexic adults. Cortex. 2010;46(6):794–798. doi: 10.1016/j.cortex.2009.09.006. [DOI] [PubMed] [Google Scholar]
- Eckert MA, Leonard CM, Richards TL, Aylward EH, Thomson J, Berninger VW. Anatomical correlates of dyslexia: Frontal and cerebellar findings. Brain. 2003;126(Pt. 2):482–494. doi: 10.1093/brain/awg026. [DOI] [PubMed] [Google Scholar]
- Farris EA, Odegard TN, Miller HL, Ring J, Allen G, Black J. Functional connectivity between the left and right inferior frontal lobes in a small sample of children with and without reading difficulties. Neurocase. 2011;17(5):425–439. doi: 10.1080/13554794.2010.532141. [DOI] [PubMed] [Google Scholar]
- Feigenson L, Dehaene S, Spelke E. Core systems of number. Trends in Cognitive Sciences. 2004;8(7):307–314. doi: 10.1016/j.tics.2004.05.002. [DOI] [PubMed] [Google Scholar]
- Fiez JA, Petersen SE. Neuroimaging studies of word reading. Proceedings of the National Academy of Sciences. 1998;95(3):914–921. doi: 10.1073/pnas.95.3.914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frye RE, Liederman J, Hasan KM, Lincoln A, Malmberg B, McLean J, III, Papanicolaou A. Diffusion tensor quantification of the relations between microstructural and macrostructural indices of white matter and reading. Human Brain Mapping. 2011;32(8):1220–1235. doi: 10.1002/hbm.21103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuchs LS, Fuchs D. Mathematical problem-solving profiles of students with mathematics disabilities with and without comorbid reading disabilities. Journal of Learning Disabilities. 2002;35(6):563–573. doi: 10.1177/00222194020350060701. [DOI] [PubMed] [Google Scholar]
- Fuchs LS, Geary DC, Compton DL, Fuchs D, Hamlett CL, Seethaler PM, Schatschneider C. Do different types of school mathematics development depend on different constellations of numerical versus general cognitive abilities? Developmental Psychology. 2010;46(6):1731–1746. doi: 10.1037/a0020662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geary DC. Mathematical disabilities: Cognitive, neuropsychological, and genetic components. Psychological Bulletin. 1993;114(2):345–362. doi: 10.1037/0033-2909.114.2.345. [DOI] [PubMed] [Google Scholar]
- Geary DC. Children’s mathematical development: Research and practical applications. Washington, DC: American Psychological Association; 1994. [Google Scholar]
- Geary DC. Mathematics and learning disabilities. Journal of Learning Disabilities. 2004;37(1):4–15. doi: 10.1177/00222194040370010201. [DOI] [PubMed] [Google Scholar]
- Geschwind N. Disconnexion syndromes in animals and man. II. Brain. 1965;88(3):585–644. doi: 10.1093/brain/88.3.585. [DOI] [PubMed] [Google Scholar]
- Golestani N, Molko N, Dehaene S, LeBihan D, Pallier C. Brain structure predicts the learning of foreign speech sounds. Cerebral Cortex. 2007;17(3):575–582. doi: 10.1093/cercor/bhk001. [DOI] [PubMed] [Google Scholar]
- Goswami U. A temporal sampling framework for developmental dyslexia. Trends in Cognitive Sciences. 2011;15(1):3–10. doi: 10.1016/j.tics.2010.10.001. [DOI] [PubMed] [Google Scholar]
- Hecht SA, Torgesen JK, Wagner RK, Rashotte CA. The relations between phonological processing abilities and emerging individual differences in mathematical computation skills: A longitudinal study from second to fifth grades. Journal of Experimental Child Psychology. 2001;79(2):192–227. doi: 10.1006/jecp.2000.2586. [DOI] [PubMed] [Google Scholar]
- Hoeft F, McCandliss BD, Black JM, Gantman A, Zakerani N, Hulme C, Gabrieli JD. Neural systems predicting long-term outcome in dyslexia. Proceedings of the National Academy of Sciences. 2011;108(1):361–366. doi: 10.1073/pnas.1008950108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoeft F, Meyler A, Hernandez A, Juel C, Taylor-Hill H, Martindale JL, Gabrieli JD. Functional and morphometric brain dissociation between dyslexia and reading ability. Proceedings of the National Academy of Sciences. 2007;104(10):4234–4239. doi: 10.1073/pnas.0609399104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horwitz B, Rumsey JM, Donohue BC. Functional connectivity of the angular gyrus in normal reading and dyslexia. Proceedings of the National Academy of Sciences. 1998;95(15):8939–8944. doi: 10.1073/pnas.95.15.8939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Houdé O, Rossi S, Lubin A, Joliot M. Mapping numerical processing, reading, and executive functions in the developing brain: An fMRI meta-analysis of 52 studies including 842 children. Developmental Science. 2010;13(6):876–885. doi: 10.1111/j.1467-7687.2009.00938.x. [DOI] [PubMed] [Google Scholar]
- Jobard G, Crivello F, Tzourio-Mazoyer N. Evaluation of the dual route theory of reading: A metaanalysis of 35 neuroimaging studies. NeuroImage. 2003;20(2):693–712. doi: 10.1016/S1053-8119(03)00343-4. [DOI] [PubMed] [Google Scholar]
- Johansen-Berg H, Behrens TE. Just pretty pictures? What diffusion tractography can add in clinical neuroscience. Current Opinion in Neurology. 2006;19(4):379–385. doi: 10.1097/01.wco.0000236618.82086.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jordan NC, Hanich LB, Kaplan D. A longitudinal study of mathematical competencies in children with specific mathematics difficulties versus children with comorbid mathematics and reading difficulties. Child Development. 2003;74(3):834–850. doi: 10.1111/1467-8624.00571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufmann L, Vogel SE, Starke M, Kremser C, Schocke M. Numerical and non-numerical ordinality processing in children with and without developmental dyscalculia: Evidence from fMRI. Cognitive Development. 2009;24(4):486–494. [Google Scholar]
- Kaufmann L, Vogel SE, Starke M, Kremser C, Schocke M, Wood G. Developmental dyscalculia: Compensatory mechanisms in left intraparietal regions in response to nonsymbolic magnitudes. Behavioral and Brain Functions. 2009;5:35. doi: 10.1186/1744-9081-5-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller TA, Just MA. Altering cortical connectivity: Remediation-induced changes in the white matter of poor readers. Neuron. 2009;64(5):624–631. doi: 10.1016/j.neuron.2009.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerns JG, Cohen JD, MacDonald AW, Cho RY, Stenger VA, Carter CS. Anterior cingulate conflict monitoring and adjustments in control. Science. 2004;303(5660):1023–1026. doi: 10.1126/science.1089910. [DOI] [PubMed] [Google Scholar]
- Koyama MS, Di Martino A, Zuo XN, Kelly C, Mennes M, Jutagir DR, Milham MP. Resting-state functional connectivity indexes reading competence in children and adults. Journal of Neuroscience. 2011;31(23):8617–8624. doi: 10.1523/JNEUROSCI.4865-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucian K, Grond U, Rotzer S, Henzi B, Schonmann C, Plangger F, von Aster M. Mental number line training in children with developmental dyscalculia. Neuroimage. 2011;57(3):782–795. doi: 10.1016/j.neuroimage.2011.01.070. [DOI] [PubMed] [Google Scholar]
- Kucian K, Loenneker T, Dietrich T, Dosch M, Martin E, von Aster M. Impaired neural networks for approximate calculation in dyscalculic children: A functional MRI study. Behavioral and Brain Functions. 2006;2:31. doi: 10.1186/1744-9081-2-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucian K, Loenneker T, Martin E, von Aster M. Non-symbolic numerical distance effect in children with and without developmental dyscalculia: A parametric fMRI study. Developmental Neuropsychology. 2011;36(6):741–762. doi: 10.1080/87565641.2010.549867. [DOI] [PubMed] [Google Scholar]
- Landerl K, Fussenegger B, Moll K, Willburger E. Dyslexia and dyscalculia: Two learning disorders with different cognitive profiles. Journal of Experimental Child Psychology. 2009;103(3):309–324. doi: 10.1016/j.jecp.2009.03.006. [DOI] [PubMed] [Google Scholar]
- Laycock R, Crewther SG. Towards an understanding of the role of the “magnocellular advantage” in fluent reading. Neuroscience & Biobehavioral Reviews. 2008;32(8):1494–1506. doi: 10.1016/j.neubiorev.2008.06.002. [DOI] [PubMed] [Google Scholar]
- Laycock SK, Wilkinson ID, Wallis LI, Darwent G, Wonders SH, Fawcett AJ, Nicolson RI. Cerebellar volume and cerebellar metabolic characteristics in adults with dyslexia. Annals of the New York Academy of Sciences. 2008;1145:222–236. doi: 10.1196/annals.1416.002. [DOI] [PubMed] [Google Scholar]
- Levin HS. Dyscalculia and dyslexia after right hemisphere injury in infancy. Archives of Neurology. 1996;53(1):88–96. doi: 10.1001/archneur.1996.00550010108024. [DOI] [PubMed] [Google Scholar]
- Lewis C, Hitch GJ, Walker P. The prevalence of specific arithmetic difficulties and specific reading difficulties in 9- to 10-year-old boys and girls. Journal of Child Psychology and Psychiatry. 1994;35(2):283–292. doi: 10.1111/j.1469-7610.1994.tb01162.x. [DOI] [PubMed] [Google Scholar]
- Ligges C, Ungureanu M, Ligges M, Blanz B, Witte H. Understanding the time variant connectivity of the language network in developmental dyslexia: New insights using Granger causality. Journal of Neural Transmission. 2010;117(4):529–543. doi: 10.1007/s00702-010-0367-x. [DOI] [PubMed] [Google Scholar]
- Lyon GR, Shaywitz SE, Shaywitz BA. A definition of dyslexia. Annals of Dyslexia. 2003;53(1):1–14. [Google Scholar]
- Maisog JM, Einbinder ER, Flowers DL, Turkeltaub PE, Eden GF. A meta-analysis of functional neuroimaging studies of dyslexia. Annals of the New York Academy of Sciences. 2008;1145:237–259. doi: 10.1196/annals.1416.024. [DOI] [PubMed] [Google Scholar]
- Mazzocco MM, Feigenson L, Halberda J. Impaired acuity of the approximate number system underlies mathematical learning disability (dyscalculia) Child Development. 2011;82(4):1224–1237. doi: 10.1111/j.1467-8624.2011.01608.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menon V, Mackenzie K, Rivera SM, Reiss AL. Prefrontal cortex involvement in processing incorrect arithmetic equations: Evidence from event-related fMRI. Human Brain Mapping. 2002;16(2):119–130. doi: 10.1002/hbm.10035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menon V, Rivera SM, White CD, Glover GH, Reiss AL. Dissociating prefrontal and parietal cortex activation during arithmetic processing. Neuroimage. 2000;12(4):357–365. doi: 10.1006/nimg.2000.0613. [DOI] [PubMed] [Google Scholar]
- Molko N, Cachia A, Riviere D, Mangin JF, Bruandet M, Le Bihan D, Dehaene S. Functional and structural alterations of the intraparietal sulcus in a developmental dyscalculia of genetic origin. Neuron. 2003;40(4):847–858. doi: 10.1016/s0896-6273(03)00670-6. [DOI] [PubMed] [Google Scholar]
- Moyer RS, Landauer TK. Time required for judgements of numerical inequality. Nature. 1967;215(5109):1519–1520. doi: 10.1038/2151519a0. [DOI] [PubMed] [Google Scholar]
- Mussolin C, De Volder A, Grandin C, Schlogel X, Nassogne MC, Noël MP. Neural correlates of symbolic number comparison in developmental dyscalculia. Journal of Cognitive Neuroscience. 2010;22(5):860–874. doi: 10.1162/jocn.2009.21237. [DOI] [PubMed] [Google Scholar]
- Mussolin C, Mejias S, Noël MP. Symbolic and nonsymbolic number comparison in children with and without dyscalculia. Cognition. 2010;115(1):10–25. doi: 10.1016/j.cognition.2009.10.006. [DOI] [PubMed] [Google Scholar]
- Newman RL, Joanisse MF. Modulation of brain regions involved in word recognition by homophonous stimuli: An fMRI study. Brain Research. 2011;1367:250–264. doi: 10.1016/j.brainres.2010.09.089. [DOI] [PubMed] [Google Scholar]
- Nicolson RI, Fawcett AJ. Dyslexia, dysgraphia, procedural learning and the cerebellum. Cortex. 2009 doi: 10.1016/j.cortex.2009.08.016. [DOI] [PubMed] [Google Scholar]
- Norman KA, Polyn SM, Detre GJ, Haxby JV. Beyond mind-reading: Multi-voxel pattern analysis of fMRI data. Trends in Cognitive Sciences. 2006;10(9):424–430. doi: 10.1016/j.tics.2006.07.005. [DOI] [PubMed] [Google Scholar]
- Piazza M, Facoetti A, Trussardi AN, Berteletti I, Conte S, Lucangeli D, Zorzi M. Developmental trajectory of number acuity reveals a severe impairment in developmental dyscalculia. Cognition. 2010;116(1):33–41. doi: 10.1016/j.cognition.2010.03.012. [DOI] [PubMed] [Google Scholar]
- Poeppel D, Idsardi WJ, van Wassenhove V. Speech perception at the interface of neurobiology and linguistics. Philosophical Transactions of the Royal Society B: Biological Sciences. 2008;363(1493):1071–1086. doi: 10.1098/rstb.2007.2160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell SR, Fuchs LS. Contribution of equal-sign instruction beyond word-problem tutoring for thirdgrade students with mathematics difficulty. Journal of Educational Psychology. 2010;102(2):381–394. doi: 10.1037/a0018447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell SR, Fuchs LS, Fuchs D, Cirino PT, Fletcher JM. Do word-problem features differentially affect problem difficulty as a function of students’ mathematics difficulty with and without reading difficulty? Journal of Learning Disabilities. 2009a;42(2):99–110. doi: 10.1177/0022219408326211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powell SR, Fuchs LS, Fuchs D, Cirino PT, Fletcher JM. Effects of fact retrieval tutoring on third-grade students with math difficulties with and without reading difficulties. Learning Disabilities Research & Practice. 2009b;24(1):1–11. doi: 10.1111/j.1540-5826.2008.01272.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prado J, Mutreja R, Zhang H, Mehta R, Desroches AS, Minas JE, Booth JR. Distinct representations of subtraction and multiplication in the neural systems for numerosity and language. Human Brain Mapping. 2011;32(11):1932–1947. doi: 10.1002/hbm.21159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price GR, Holloway I, Rasanen P, Vesterinen M, Ansari D. Impaired parietal magnitude processing in developmental dyscalculia. Current Biology. 2007;17(24):R1042–R1043. doi: 10.1016/j.cub.2007.10.013. [DOI] [PubMed] [Google Scholar]
- Quaglino V, Bourdin B, Czternasty G, Vrignaud P, Fall S, Meyer ME, de Marco G. Differences in effective connectivity between dyslexic children and normal readers during a pseudoword reading task: An fMRI study. Neurophysiologie Clinique/Clinical Neurophysiology. 2008;38(2):73–82. doi: 10.1016/j.neucli.2007.12.007. [DOI] [PubMed] [Google Scholar]
- Raschle NM, Chang M, Gaab N. Structural brain alterations associated with dyslexia predate reading onset. NeuroImage. 2011;57(3):742–749. doi: 10.1016/j.neuroimage.2010.09.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards TL, Berninger VW. Abnormal fMRI connectivity in children with dyslexia during a phoneme task: Before but not after treatment. Journal of Neurolinguistics. 2008;21(4):294–304. doi: 10.1016/j.jneuroling.2007.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richlan F, Kronbichler M, Wimmer H. Functional abnormalities in the dyslexic brain: A quantitative meta-analysis of neuroimaging studies. Human Brain Mapping. 2009;30(10):3299–3308. doi: 10.1002/hbm.20752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richlan F, Kronbichler M, Wimmer H. Metaanalyzing brain dysfunctions in dyslexic children and adults. Neuroimage. 2011;56(3):1735–1742. doi: 10.1016/j.neuroimage.2011.02.040. [DOI] [PubMed] [Google Scholar]
- Richlan F, Kronbichler M, Wimmer H. Structural abnormalities in the dyslexia brain: A meta-analysis of voxelbased morphometry studies. Human Brain Mapping. doi: 10.1002/hbm.22127. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivera SM, Reiss AL, Eckert MA, Menon V. Developmental changes in mental arithmetic: Evidence for increased functional specialization in the left inferior parietal cortex. Cerebral Cortex. 2005;15(11):1779–1790. doi: 10.1093/cercor/bhi055. [DOI] [PubMed] [Google Scholar]
- Rosenberg-Lee M, Barth M, Menon V. What difference does a year of schooling make? Maturation of brain response and connectivity between 2nd and 3rd grades during arithmetic problem solving. Neuroimage. 2011;57(3):796–808. doi: 10.1016/j.neuroimage.2011.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenberg-Lee M, Chang TT, Young CB, Wu S, Menon V. Functional dissociations between four basic arithmetic operations in the human posterior parietal cortex: A cytoarchitectonic mapping study. Neuropsychologia. 2011;49(9):2592–2608. doi: 10.1016/j.neuropsychologia.2011.04.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rotzer S, Kucian K, Martin E, von Aster M, Klaver P, Loenneker T. Optimized voxel-based morphometry in children with developmental dyscalculia. Neuroimage. 2008;39(1):417–422. doi: 10.1016/j.neuroimage.2007.08.045. [DOI] [PubMed] [Google Scholar]
- Rotzer S, Loenneker T, Kucian K, Martin E, Klaver P, von Aster M. Dysfunctional neural network of spatial working memory contributes to developmental dyscalculia. Neuropsychologia. 2009;47(13):2859–2865. doi: 10.1016/j.neuropsychologia.2009.06.009. [DOI] [PubMed] [Google Scholar]
- Rousselle L, Noël MP. Basic numerical skills in children with mathematics learning disabilities: A comparison of symbolic vs non-symbolic number magnitude processing. Cognition. 2007;102(3):361–395. doi: 10.1016/j.cognition.2006.01.005. [DOI] [PubMed] [Google Scholar]
- Rubinsten O, Henik A. Automatic activation of internal magnitudes: A study of developmental dyscalculia. Neuropsychology. 2005;19(5):641–648. doi: 10.1037/0894-4105.19.5.641. [DOI] [PubMed] [Google Scholar]
- Rubinsten O, Henik A. Double dissociation of functions in developmental dyslexia and dyscalculia. Journal of Educational Psychology. 2006;98(4):854–867. [Google Scholar]
- Rykhlevskaia E, Uddin LQ, Kondos L, Menon V. Neuroanatomical correlates of developmental dyscalculia: Combined evidence from morphometry and tractography. Frontiers in Human Neuroscience. 2009;3:51. doi: 10.3389/neuro.09.051.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlaggar BL, McCandliss BD. Development of neural systems for reading. Annual Review of Neuroscience. 2007;30:475–503. doi: 10.1146/annurev.neuro.28.061604.135645. [DOI] [PubMed] [Google Scholar]
- Shalev R, Auerbach J, Manor O, Gross-Tsur V. Developmental dyscalculia: Prevalence and prognosis. European Child & Adolescent Psychiatry. 2000;9(0):S58–S64. doi: 10.1007/s007870070009. [DOI] [PubMed] [Google Scholar]
- Shaywitz BA, Shaywitz SE, Pugh KR, Mencl WE, Fulbright RK, Skudlarski P, Gore JC. Disruption of posterior brain systems for reading in children with developmental dyslexia. Biological Psychiatry. 2002;52(2):101–110. doi: 10.1016/s0006-3223(02)01365-3. [DOI] [PubMed] [Google Scholar]
- Shaywitz SE, Shaywitz BA. Dyslexia. Biological Psychiatry. 2005;57(11):1301–1309. doi: 10.1016/j.biopsych.2005.01.043. [DOI] [PubMed] [Google Scholar]
- Shaywitz SE, Shaywitz BA. Paying attention to reading: The neurobiology of reading and dyslexia. Development and Psychopathology. 2008;20(4):1329–1349. doi: 10.1017/S0954579408000631. [DOI] [PubMed] [Google Scholar]
- Siegler RS. Emerging minds: The process of change in children’s thinking. New York, NY: Oxford University Press; 1996. [Google Scholar]
- Silani G, Frith U, Demonet JF, Fazio F, Perani D, Price C, Paulesu E. Brain abnormalities underlying altered activation in dyslexia: A voxel based morphometry study. Brain. 2005;128(Pt. 10):2453–2461. doi: 10.1093/brain/awh579. [DOI] [PubMed] [Google Scholar]
- Stanovich KE. Phenotypic performance profile of children with reading disabilities: A regression-based tests of the phonological-core variable-difference model. Journal of Educational Psychology. 1994;86(1):24–53. [Google Scholar]
- Stein J. The magnocellular theory of developmental dyslexia. Dyslexia. 2001;7(1):12–36. doi: 10.1002/dys.186. [DOI] [PubMed] [Google Scholar]
- Supekar K, Menon V. Developmental maturation of dynamic causal control signals in higher-order cognition: A neurocognitive network model. PLoS Computational Biology. 2012;8(2):e1002374. doi: 10.1371/journal.pcbi.1002374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swanson HL, Howard CB, Saez L. Do different components of working memory underlie different subgroups of reading disabilities? Journal of Learning Disabilities. 2006;39(3):252–269. doi: 10.1177/00222194060390030501. [DOI] [PubMed] [Google Scholar]
- Tallal P. Improving language and literacy is a matter of time. Nature Reviews Neuroscience. 2004;5(9):721–728. doi: 10.1038/nrn1499. [DOI] [PubMed] [Google Scholar]
- Toll SWM, Van der Ven SHG, Kroesbergen EH, Van Luit JEH. Executive functions as predictors of math learning disabilities. Journal of Learning Disabilities. 2011;44(6):521–532. doi: 10.1177/0022219410387302. [DOI] [PubMed] [Google Scholar]
- Torgesen JK, Wagner RK, Rashotte CA, Burgess S, Hecht S. Contributions of phonological awareness and rapid automatic naming ability to the growth of word-reading skills in second- to fifth-grade children. Scientific Studies of Reading. 1997;1(2):161–185. [Google Scholar]
- Uddin LQ, Supekar K, Amin H, Rykhlevskaia E, Nguyen DA, Greicius MD, Menon V. Dissociable connectivity within human angular gyrus and intraparietal sulcus: Evidence from functional and structural connectivity. Cerebral Cortex. 2010;20(11):2636–2646. doi: 10.1093/cercor/bhq011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Mark S, Klaver P, Bucher K, Maurer U, Schulz E, Brem S, Brandeis D. The left occipitotemporal system in reading: Disruption of focal fMRI connectivity to left inferior frontal and inferior parietal language areas in children with dyslexia. NeuroImage. 2011;54(3):2426–2436. doi: 10.1016/j.neuroimage.2010.10.002. [DOI] [PubMed] [Google Scholar]
- van der Sluis S, de Jong PF, Leij A. Inhibition and shifting in children with learning deficits in arithmetic and reading. Journal of Experimental Child Psychology. 2004;87(3):239–266. doi: 10.1016/j.jecp.2003.12.002. [DOI] [PubMed] [Google Scholar]
- Vidyasagar TR, Pammer K. Dyslexia: A deficit in visuo-spatial attention, not in phonological processing. Trends in Cognitive Sciences. 2010;14(2):57–63. doi: 10.1016/j.tics.2009.12.003. [DOI] [PubMed] [Google Scholar]
- Vigneau M, Beaucousin V, Herve PY, Duffau H, Crivello F, Houde O, Tzourio-Mazoyer N. Metaanalyzing left hemisphere language areas: Phonology, semantics, and sentence processing. NeuroImage. 2006;30(4):1414–1432. doi: 10.1016/j.neuroimage.2005.11.002. [DOI] [PubMed] [Google Scholar]
- von Aster M, Shalev R. Number development and developmental dyscalculia. Developmental Medicine & Child Neurology. 2007;49(11):868–873. doi: 10.1111/j.1469-8749.2007.00868.x. [DOI] [PubMed] [Google Scholar]
- Vourkas M, Micheloyannis S, Simos PG, Rezaie R, Fletcher JM, Cirino PT, Papanicolaou AC. Dynamic task-specific brain network connectivity in children with severe reading difficulties. Neuroscience Letters. 2011;488(2):123–128. doi: 10.1016/j.neulet.2010.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner RK, Torgesen JK. The nature of phonological processing and its role in the acquisition of reading skills. Psychological Bulletin. 1987;101:192–212. [Google Scholar]
- Wandell BA, Rauschecker AM, Yeatman JD. Learning to see words. Annual Review of Psychology. 2012;63:31–53. doi: 10.1146/annurev-psych-120710-100434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willburger E, Fussenegger B, Moll K, Wood G, Landerl K. Naming speed in dyslexia and dyscalculia. Learning and Individual Differences. 2008;18(2):224–236. [Google Scholar]
- Wilson AJ, Dehaene S. Number sense and developmental dyscalculia. In: Coch D, Dawson G, Fischer KW, editors. Human behavior, learning, and the developing brain: Atypical development. New York, NY: Guilford; 2007. pp. 212–238. [Google Scholar]
- Wolf M. Naming speed and reading: The contribution of the cognitive neurosciences. Reading Research Quarterly. 1991;26(2):123–141. [Google Scholar]
- Wolf M, Bowers PG. The double-deficit hypothesis for the developmental dyslexias. Journal of Educational Psychology. 1999;91(3):415–438. [Google Scholar]
- Xu F, Carey S. Infants’ metaphysics: The case of numerical identity. Cognitive Psychology. 1996;30(2):111–153. doi: 10.1006/cogp.1996.0005. [DOI] [PubMed] [Google Scholar]
- Xu F, Spelke ES. Large number discrimination in 6-month-old infants. Cognition. 2000;74(1):B1–B11. doi: 10.1016/s0010-0277(99)00066-9. [DOI] [PubMed] [Google Scholar]
- Zago L, Petit L, Turbelin MR, Andersson F, Vigneau M, Tzourio-Mazoyer N. How verbal and spatial manipulation networks contribute to calculation: An fMRI study. Neuropsychologia. 2008;46(9):2403–2414. doi: 10.1016/j.neuropsychologia.2008.03.001. [DOI] [PubMed] [Google Scholar]
- Zebian S, Ansari D. Differences between literates and illiterates on symbolic but not nonsymbolic numerical magnitude processing. Psychonomic Bulletin & Review. 2012;19(1):93–100. doi: 10.3758/s13423-011-0175-9. [DOI] [PubMed] [Google Scholar]