

Abstract
Formation of protein aggregates in the aging eye lens has been shown to correlate with progressive accumulation of specific low molecular weight (LMW) peptides derived from crystallins. Prominent among the LMW fragments is αA66-80, a peptide derived from αA-crystallin and present in increased concentrations in the water-insoluble (WIS) nuclear fractions of the aging lens. The αA66-80 peptide has amyloid-like properties and preferentially insolubilizes α-crystallin from soluble lens fractions. However, the specific interactions and mechanisms by which the peptide induces α-crystallin aggregation have not been delineated. To gain insights into the mechanisms of peptide-induced aggregation, we investigated the peptide interactions with α-crystallin by various biochemical approaches. The peptide diminishes α-crystallin chaperone ability and drastically promotes α-crystallin aggregation by formation of insoluble peptide-protein complexes through transient intermediates. Bis-ANS studies suggest that the peptide induces changes in hydrophobicity of α-crystallin that could trigger the formation and growth of aggregates. The peptide-α-crystallin aggregates were found to be resistant to dissociation by high ionic strength, whereas guanidium hydrochloride and urea were effective dissociating agents. We conclude that the αA66-80 peptide forms a hydrophobically driven, stable complex with α-crystallin and reduces its solubility. Using isotope-labeled chemical crosslinking and mass spectrometry, we show that the peptide binds to multiple sites, including the chaperone site, C-terminal extension and subunit interaction sites in αB-crystallin, which may explain the anti-chaperone property of the peptide and the consequential age-related accumulation of aggregated proteins. Thus, the α-crystallin-derived peptide could play a role in the pathogenesis of cataract formation in the aging lens.
Keywords: α-crystallin, aggregation, cataract, peptide interaction, cross-linking
Age-related cataract is common and causes a huge economic burden. Cataract is characterized by increasing cloudiness of the lens and progressive loss of vision due to the formation of protein aggregates.1 The formation of peptide and/or protein aggregates is associated with the pathogenesis of numerous debilitating human diseases, such as Alzheimer’s disease, systemic amyloidosis and Parkinson’s disease.2 The underlying processes behind the development of these protein aggregation disorders are unclear. A similar unexplained phenomenon operates behind the development of protein aggregation in lens. Multiple factors such as age-related modifications of lens proteins, oxidative stress and other conditions are believed to be involved, but how they influence the rate and extent of protein aggregation remain unknown.
Lens transparency relies on the solubility and stability of lens proteins, called crystallins, of which there are of three types: α-, β- and γ-crystallins. α-Crystallin, a small heat shock protein, is the most abundant of lens proteins and is composed of two subunits, αA and αB, each 20 kDa. α-Crystallin subunits oligomerize to a polydisperse protein with an average molecular mass of 800 kDa.1 α-Crystallin has chaperone activity3 and is responsible for suppressing the aggregation of other lens proteins. The chaperone activity of α-crystallin and the molecular interactions among the crystallins are responsible for maintaining lens transparency and the appropriate refractive index.3,4 Progressive age-related loss of lens transparency and cataract formation has been attributed to various post-translational modifications in lens proteins. These modifications include deamidation, oxidation, methylation, phosphorylation and truncation. Racemization of Ser, Thr, Glu/Gln and Phe in lens proteins has recently been reported in aged and cataract lenses.5 With these changes, aged lens proteins exhibit decreased chaperone activity, altered stability and decreased solubility, resulting in the formation of high molecular weight (HMW) aggregates.6,7 The most notable change observed in the aging lens is the progressive degradation of crystallins.8,9 In human lens, the quantity of degraded crystallins increases with the severity of cataract. Increased quantities of truncated crystallins and LMW peptide fragments are present in the water-insoluble aggregates of aging and cataract lenses.10–12 Decreased clearance of peptide fragments by peptidases or increased proteolysis has been implicated in the accumulation of peptide fragments in the aging lens. It is not known whether cleavage of crystallins to LMW peptides is an important step in the process of age-related cataract formation or is a normal event in the degradation of crystallins.
The release of peptide fragments from native proteins as a result of abnormal proteolysis of host proteins is a major contributing factor in the pathogenesis of various neurodegenerative diseases.13 In such diseases, specific peptide fragments that remain part of the three-dimensional conformation in native proteins do not have aggregation potential, but when peptide fragments become cleaved and released, they exhibit a propensity for aggregation and a tendency to influence the stability and aggregation behavior of proteins.14 Thus, the age-related appearance of peptide fragments and associated protein aggregation in the lens has led to the emerging hypothesis that peptides could be the initiating factor or a facilitating factor in protein aggregation and cataract formation. This hypothesis gains significance for the following reasons. First, aging and cataract lenses show increased proteolysis as compared to age-matched non-cataract lenses.15,16 Second, the crystallin fragments are largely found co-localized with aggregates in the opaque region of the lens as compared to the clear region of the same lens.8,11,12,15,16 Third, peptides have interaction sites in soluble crystallins and are found to modulate the normal physiology of the lens.17 For example, an oxidized peptide from βB3-crystallin has been shown to increase the aggregation of both β- and γ-crystallins.18 Moreover, peptides have been found to suppress the chaperone ability of α-crystallin, increase the oligomeric size of soluble crystallin,11,19 interact tightly with cytoskeletal and membrane components, alter the interaction of soluble α-crystallin with membrane20,21 and form amyloid-type aggregates in vitro.19,22 Such aberrant interactions with soluble proteins and cellular components are the characteristic features of pathological peptides such as p25, β-amyloid and polyglutamine fragments as well as the synthetic peptides with amyloidogenic potential.13,23,24
A degradation product of αA-crystallin, 66SDRDKFVIFLDVKHF80 (αA66-80), is prominent among the LMW crystallin fragments (<3.5 kDa) that accumulate in the aging lens (Figure 1). We have characterized the lens distribution, properties and aggregation potential of this peptide.19 The αA66-80 peptide is derived from the chaperone site in αA-crystallin25 and has regions homologous to Aβ(1–42) peptide, which is responsible for its polymerizing properties.26 The αA66-80 peptide preferentially insolubilizes α-crystallin from soluble lens crystallins composed of α-, β- and γ-crystallins. The peptide-induced α-crystallin aggregates serve as seeds for further aggregation with other soluble proteins.19 These attributes, combined with the observation that αA66-80 peptide is distributed primarily in the WIS fractions from nuclear region of the human lens,12,19 provide strong evidence, and an explanation, for the decrease in soluble α-crystallin with aging, especially in the lens nucleus and to a greater extent in the cataractous lens.27,28 Thus, it is probable that αA66-80 peptide plays a critical role in crystallin aggregation and cataract formation. However, the mechanisms underlying the generation of the peptide in the lens are yet to be elucidated, including the α-crystallin sequences involved in interactions with αA66-80 peptide and how the peptide induces aggregation of soluble α-crystallin. To gain insights into the effect of accumulating αA66-80 peptide on α-crystallin structure and function, we have investigated α-crystallin aggregation in the presence of the αA66-80 peptide. In the present study, we show a direct interaction between the peptide and αcrystallin subunits as well as the possible mechanism underlying peptide-induced aggregate formation. We identified the peptide interaction regions in αB-crystallin using a novel cross-linker. We found that the peptide suppresses α-crystallin chaperone activity, decreases the solubility of α-crystallin and increases the surface hydrophobicity in α-crystallin by forming stable non-covalent interactions with α-crystallin. Additionally, we describe the in vitro effects of the peptide on α-crystallin and discuss the potential impact of αA66-80 in initiating or facilitating aggregation in lens. Our findings enhance our understanding of the mechanisms of protein aggregation in cataract formation, which may influence therapeutic strategies to prevent or delay the onset of cataract.
Figure 1.

αA-crystallin sequence showing the αA66-80 peptide regions and chaperone site. Red – residues 66–80, Underlined sequence is the chaperone site in aα-crystallin.
MATERIALS AND METHODS
Materials
Synthetic peptides—66SDRDKFVIFLDVKHF80 (αA66-80), 43 TISPYYRQSLFRTV56 (αA43-56), and SDRDKFVIFLDVKHFK-Biotin (C-terminal biotinylated αA66-80), SDRDKFPIFLDVKHF (proline-substituted αA66-80) and EEKPAVTAAPK (C-terminal peptide of αB-crystallin)—were obtained from Genscript Corporation. The purity of the synthetic peptide exceeded 95%. The lyophilized peptide (2 mg) was dissolved in sterile water at 5 mg mL−1, sonicated for 10 sec, filtered through 0.2 μm filter and used immediately. The peptide incubations and assays were carried out in phosphate buffer (50 mM sodium phosphate buffer containing 150 mM NaCl, pH 7.2). Alcohol dehydrogenase (ADH) was obtained from Worthington Biochemical. The cross linking reagents BS2G d0/d4 (Bis (sulfo succindimydyl) glutarate) were purchased from Proteochem. Bis-ANS (4, 4′-dianilino -1,1′-binapthyl -5-5′-disulfonic acid) was obtained from Sigma-Aldrich. Bovine trypsin (sequencing grade) was obtained from G-Biosciences. Bio-Rad protein assay reagent was used to estimate the protein concentrations in various assays.
Preparation of α-Crystallin
α-Crystallin from bovine lenses was isolated by gel filtration on Sephadex G-200 and ion-exchange chromatography on Q-Sepharose (GE Healthcare Biosciences) as described earlier.29
Expression and Purification of Recombinant αB-Crystallin
Human αB-crystallin cDNA (obtained from Dr. J. Mark Petrash, University of Colorado, Denver) was cloned in a pET23d vector (Novagen) at the NcoI/Hind III site and expressed in E. coli BL21 (DE3) pLysS cells (Invitrogen). The recombinant protein was purified by a combination of gel-filtration (Superdex G-200) and ion-exchange chromatography (Q-Sepharose), as described earlier.30 The purity of the protein was confirmed by SDS-PAGE. The recombinant αB-crystallin used for the cross linking experiments was of >95% purity.
Chaperone Assays
The ability of α-crystallin to suppress aggregation of ADH in the presence of αA66-80 peptide was assessed as described earlier.19 αA43-56 peptide was used as a control, as this peptide has no aggregation potential.31 α-Crystallin (50 μg) was incubated with ADH (150 μg) and αA66-80 or αA43-56 peptide (5 μg and 15 μg) in 1 ml of 50 mM phosphate buffer containing 50 mM EDTA (pH 7.2). Aggregation was monitored for 120 min by measuring the absorption at 360 nm at 37°C in a spectrophotometer equipped with a temperature-regulated multi-cell holder. After the chaperone assay, the samples were centrifuged at 8000 rpm for 15 min. The composition of the pellet was determined by SDS-PAGE.
Aggregation Assays
Susceptibility of the protein to peptide-induced aggregation was evaluated by different means. Samples containing α-crystallin (5 μM) and peptide (25 μM) in 1 ml of 50 mM phosphate buffer were incubated at 37°C. To assess protein aggregation, turbidity of 1 ml solution was determined spectrophotometrically by measuring the change in absorbance at 360 nm at different durations of incubation. α-Crystallin samples incubated without added peptide were used as a control. The concentration of the soluble α-crystallin remaining in the supernatant as a function of time during incubation was determined by Bio-Rad assay. In another experiment, α-crystallin (10 μM) was incubated with peptide (1.25–20 μM) in 1 ml of 50 mM phosphate buffer at 37°C for 24 h. The samples were then centrifuged at 8000 rpm for 15 min and pellets were run in SDS-PAGE.
Intrinsic Tryptophan Fluorescence Measurements
The fluorescence spectra of α-crystallin in the presence and absence of αA66-80 peptide were recorded using JASCO FP750 spectrofluorometer. α-Crystallin (5 μM) was mixed with increasing concentrations of peptide (molar ratio of α-crystallin: αA66-80 – 1:0, 1:0.25, 1:0.5, 1:1, 1:1.5, 1:2) in 1 ml of 50 mM phosphate buffer (pH 7.2). The samples were excited at 295 nm and tryptophan emission spectra were recorded between 300–400 nm (bandwidth 5 nm). All reaction mixtures were incubated for 10 min at 37°C to reach equilibrium prior to measurements.
Circular Dichroism Measurements
Far-UV circular dichroism (CD) experiments were carried out using a JASCO J-815 spectropolarimeter equipped with a thermostated cell holder. α-Crystallin (5 μM) was mixed with αA66-80 (molar ratio of α-crystallin : αA66-80 – 1:0, 1:0.25, 1:0.5, 1:1, 1:1.5, 1:2 and 1:4). Samples taken in 10 mM phosphate buffer (pH 7.2) were incubated at 37°C for 10 min before far-UV CD measurements. Measurements were acquired in the range of 200–240 nm using a 1-mm path quartz cuvette at a temperature of 25°C. All spectra were corrected by subtracting the buffer baseline and appropriate peptide controls. The spectrum derived represents an average of five scans.
Size Exclusion Chromatography (SEC)
α-Crystallin (5 μM) was incubated with αA66-80 (molar ratio of α-crystallin : αA66-80 – 1:0, 1:0.25, 1:0.5, 1:1, 1:2 and 1:4) in 250 μl of 50 mM phosphate buffer (pH 7.2) at 37°C for 10 min. Samples were then filtered through 0.45 μ filter (Millipore). Filtrate (150 μl) was injected onto a TSK G5000PWXL column (Tosoh Bioscience) gel-filtration column connected to high-performance liquid chromatography (HPLC) (Shimadzu) equipped with UV and refractive index detectors. Samples were eluted using 50 mM phosphate buffer (pH 7.2), and the elution was monitored at 280 nm.
Bis-ANS Binding Studies
Bis-ANS is an environment-sensitive hydrophobic fluorescent dye. The fluorescence quantum yield of bis-ANS increases upon binding to hydrophobic regions of proteins such as α-crystallin.32 Bis-ANS (20 μM) was added to α-crystallin (10 μM) incubated with αA66-80 peptide (molar ratio of α-crystallin : αA66-80 – 1:0, 1:0.25, 1:0.5, 1:1, 1:1.5 and 1:2) in 1 ml of 50 mM phosphate buffer, pH 7.2. The samples were thoroughly mixed and incubated for 10 min before measurements were obtained. Fluorescence of the bis-ANS bound to α-crystallins incubated in the presence and absence of peptides was measured using a JASCO FP750 spectrofluorometer. The interaction between the C-terminal region of αB-crystallin (EEKPAVTAAPK; αB-CTP) and αA66-80 peptide was confirmed in a separate experiment. Fluorescence intensity of bis-ANS bound to α-crystallin (100 μg) was measured in the presence of αB-CTP (10 μg) and αA66-80 (10 μg) or proline-substituted αA66-80 (10 μg). Fluorescence emission spectra were recorded at 400–600 nm using an excitation wavelength of 390 nm. The excitation and emission slits were set at 5 nm.
Dissociation Studies of α-Crystallin-αA66-80 Complexes
α-Crystallin (200 μg) was incubated with αA66-80 (100 μg) in 500 μl of 50 mM phosphate buffer for 24 h at 37°C. After centrifugation at 12,000 rpm for 30 min, aggregates were suspended in 100 μl of each of the dissociating agents for 1 h with gentle stirring at 37°C (50 mM phosphate buffer containing 1% SDS, 6 M GdmCl, 8 M urea, 1% Tween-20, 1% Triton X-100, 1 M NaCl and 1M MgCl2). The redissolved aggregates were then centrifuged at 12,000 rpm for 30 min. The soluble protein content of the supernate fractions of redissolved aggregates was determined by Bio-Rad protein assay and compared with the protein content of samples that were not incubated.
Cross-Linking Studies
The peptide interaction sites in αB-crystallin were evaluated using chemical cross-linking and mass spectrometry approaches. Isotope-labeled homobifunctional cross-linker N-hydroxysuccinimide esters BS2G-d0/d4 were used in 1:1 ratio mixtures of their nondeuterated (d0) and deuterated (d4) species. Reaction mixture containing αB-wild-type (WT) crystallin (200 μg) and biotin-labeled αA66-80 (100 μg) (2:1 W/W) in 1ml of 25 mM phosphate buffer (pH 7.4) was incubated at room temperature for 30 min prior to the addition of the cross-linker. A αB-WT crystallin sample without added peptide was used as control. One molar stock solution of BS2G- d0 and d4 cross linker in DMSO was prepared. A 20-fold molar excess of the cross-linker (1:1 ratio of d0 and d4) was added to the peptide-protein incubation mixture. Cross-linking reactions were allowed to proceed for 45 min in ice. A control sample containing the protein-peptide mixture without added cross-linker was maintained. The cross-linking reaction was terminated by adding 50 μl of Tris (final concentration 20 μM) solution to each sample. After incubation at room temperature for 10 min, the samples were passed through 10K centrifugal filter device (Centricon) to remove unreacted cross linker.
SDS-PAGE, Western Blotting and Digestion of Cross-Linked Products
The cross-linking reaction mixtures were separated by one-dimensional SDS-PAGE. The resolved samples were transferred to a PVDF membrane and, using Avidin-HRP (Pierce), assayed for the detection of cross-linked complex of αB-crystallin that is covalently linked with biotinylated αA66-80. Following this, the bands of interest containing the cross-linked product were excised and in-gel-digested using trypsin. The samples were dissolved in trypsin digestion buffer (0.2 M ammonium bicarbonate, pH 7.9), and 2 μg of bovine trypsin was added to 200 μg of total proteins. After incubation for 2 h, a second batch of trypsin (2 μg) was added to achieve complete digestion. The digestion took place in complete darkness at 37°C overnight.
Identification of Cross-Linked Products by LC-MS/MS
The extracted peptides were lyophilized and resuspended in 8 μl of 5% acetonitrile and 1% formic acid. Separation of peptides was achieved in two steps. A portion of the digest was loaded onto a C8 trap column (Michrom Bioresources). Eluates were then passed through C18 reversed-phase resin. Peptides were eluted on 5% to 40% acetonitrile (in 0.1% formic acid) over 70 min. Eluted peptides were reversed-phase separated using integrated Proxeon nano-HPLC coupled on-line to an LTQ-Orbitrap XL spectrometer (Thermo-Scientific). Mass acquisition was done using a high-resolution FTMS scan of the eluting peptides. Post-acquisition, the LC-MS experimental peptide mass lists obtained were matched with theoretical mass lists of cross-linked peptides generated using GPMAW (General Protein Mass Analysis for Windows, version 9.2) software with mass tolerance set at 5 ppm. The GPMAW software program allows the user to define the protein and the peptide sequences as well as the homobifunctional amine-reactive cross-linker –BS2G used for the study. Amine group in the protein and peptide were chosen as reactive sites for the cross-linker. With this approach theoretical peak lists corresponding to peptides cross-linked with light (d0) and heavy (d4) precursors were generated. Subsequently by comparison with the GPMAW generated possible cross-linked peptides, the m/z values obtained from LC-MS were assigned to cross-linked products. The identified cross-linked peptides (parent ion) from LC-MS were validated by MS/MS analysis. In the second MS, the parent ion selected is subjected to peptide fragmentation (CID in ion trap and HCD in C-trap) producing the MS/MS spectrum. The b and the y ion series resulting from fragmentation along the peptide backbone were interpreted from the low mass end through to the highest mass ion. Peptide sequences were confirmed using MASCOT distiller software.
RESULTS
αA66-80 Peptide Suppresses the Chaperone Activity of α-Crystallin
The ability of α-crystallin to suppress aggregation of ADH in the presence of peptides was assessed. When native ADH was incubated at 37°C in the presence of EDTA containing phosphate buffer (50 mM pH 7.5), ADH unfolded and aggregated into particles sufficiently large enough to scatter light at 360 nm (Figure 2, curve 1). In contrast, incubation with α-crystallin suppressed aggregation of the ADH in its denatured state due to α-crystallin chaperone activity (Figure 2, curve 2). The control peptide αA43-56 had no effect on α-crystallin chaperone activity (Figure 2, curves 3 & 4) and did not cause aggregation of samples. The addition of αA66-80 diminished the chaperone activity of α-crystallin and increased in a concentration-dependent manner the aggregation and precipitation of α-crystallin and ADH substrate (Figure 2, curves 5 & 6). Additionally, the peptides by themselves did not precipitate and scatter light under the experimental conditions (data not shown). SDS-PAGE analysis of the samples after the chaperone assay (Figure 2 inset) revealed that the samples containing control peptide αA43-56 (Figure 2 inset Lanes 3 & 4) had less ADH substrate and α-crystallin in the pellets than the samples containing αA66-80 peptide (Figure 2 inset Lanes 5 & 6). Samples that contained αA66-80 peptide had significantly more precipitation of α-crystallin (20 kDa bands), ADH substrate and αA66-80 peptide in the pellets. The data suggest that αA66-80 peptide facilitates the aggregation of α-crystallin and suppresses its chaperone activity. The interaction of the peptide with α-crystallin or with the α-crystallin-ADH complex might be responsible for the enhanced aggregation and increased light scattering. These data suggest that α-crystallin is incorporated into larger aggregates composed of α-crystallin, ADH and peptide.
Figure 2.

Effects of αA66-80 peptide on chaperone activity of α-crystallin. The kinetics of aggregation were monitored by measuring the light scattering of the samples at 360 nm for 120 min. α-crystallin was preincubated with peptides separately for 1 h in 37°C before the chaperone activity assay. 1 ADH (150 μg), 2 ADH (150 μg) + α-crystallin (50 μg), 3 ADH (150 μg) + α-crystallin (50 μg) + αA43-56 (5 μg), 4 ADH (150 μg) + α-crystallin (50 μg) + αA43-56 (15 μg), 5 ADH (150 μg) +α-crystallin (50 μg) + αA66-80 (5 μg), 6 ADH (150 μg) + α-crystallin (50 μg)+ αA66-80 (15 μg). InsetT SDS-PAGE analysis: Lanes 1–6 are precipitates of chaperone assay samples 1–6. Samples containing αA66-80 peptide (Lanes 5 & 6) have more α-crystallin, ADH substrate and peptide precipitated than samples containing αA43–56 peptide (Lanes 3 & 4).
αA66-80 Peptide Facilitates α-Crystallin Aggregation
To investigate whether α-crystallins incubated with αA66-80 peptide are more prone to aggregate, the turbidity of the samples was monitored spectrophotometrically by measuring the change in absorbance at 360 nm. When incubated with αA66-80, α-crystallin showed a progressive increase in turbidity during 0 to 300 min of incubation. Under the same conditions, the control α-crystallin and the αA66-80 peptide alone did not show an increase in turbidity, indicating that the peptide is causing the turbidity in α-crystallin solution (Figure 3A). The solubility of α-crystallin decreased as the duration of incubation with peptide increased (Figure 3B), thus confirming the influence of αA66-80 on turbidity. The peptide-mediated aggregation is also evidenced by the precipitation of α-crystallins, as confirmed by SDS-PAGE analysis (Figure 3C). The α-crystallin used in the study contained 12 and 17 kDa polypeptides as minor contaminants. However, these impurities by themselves did not contribute to the aggregation of α-crystallin, since the incubation of crystallin fraction by itself did not result in appreciable precipitation (Figure 3C, lane 1) whereas α-crystallin fraction containing increasing concentration of αA66-80 peptide showed significant precipitation of proteins (Figure 3C, lanes 5, 6 and 7). This indicates that α66–80 was primarily responsible for the aggregation and precipitation of α-crystallins. Precipitation seems to be more prominent when the crystallin-peptide ratio is 1:1, 1:1.5 and 1:2 μM (lanes 5, 6 and 7). Thus, an α-crystallin–αA66-80 peptide ratio of at least 1:1 may be the critical concentration required for precipitation. Thus, the presence of the αA66-80 peptide and α-crystallin together in the aggregates correlates with the in vivo accumulation of the peptide in water-insoluble aggregates19 and points to α-crystallin as a major component in cataractous inclusions.8 Intrinsic tryptophan fluorescence was measured to investigate the effect of the peptide on α-crystallin conformation. α-Crystallin subunits have three tryptophan residues (9th position in αA-crystallin and 9th and 60th positions in αB-crystallin), whereas the peptide has no tryptophan residues. Therefore, the fluorescence emission of α-crystallin was monitored in the presence and absence of αA66-80 peptide to evaluate the change in tryptophan fluorescence following the binding of the peptide to α-crystallin. Native α-crystallin showed emission maxima at 339 nm when excited at 295 nm. Emission spectra of α-crystallin mixed with αA66-80 peptide showed a decrease in the fluorescence intensity with increasing peptide concentration (Figure 4A). However, there was no shift in the emission maximum. These results suggest the possible association of the peptide with the protein and quenching of the tryptophan emission due to peptide-induced aggregation. Since there are no available binding models under such aggregating conditions, no meaningful Kd was deducible from our intrinsic fluorescence measurements. Aggregation of α-crystallin in the presence of αA66-80 peptide was also evident from our CD (Figure 4B) and SEC (Figure 4C) studies. Far-UV spectrum of the control αcrystallin sample showed a minimum at 218 nm, consistent with previously published results.33 In α-crystallin samples incubated with αA66-80 peptide, the spectra did not change, but the CD signal intensity decreased with increased αA66-80 peptide concentration. The CD signal decrease is probably a consequence of aggregate formation, which eventually causes precipitation and loss of the signal (Figure 4B). SEC elution profiles demonstrated no additional soluble aggregate peaks of α-crystallin at any of the peptide concentrations. However, the α-crystallin oligomer showed reduced peak intensity in association with increased αA66-80 concentration, indicating the formation of insoluble aggregates that are held up by pre-column filters in SEC (Figure 4C).
Figure 3.

αA66-80-induced aggregation of α-crystallin. (A) α-crystallin (5 μM) incubated with αA66-80 (25 μM) in 1 ml of 50 mM phosphate buffer monitored at 360 nm at different durations of incubation (data presented as mean ± SD of 3 independent measurements). (B) The concentration of soluble proteins at different time points during incubation (data represented as mean ± SD of 3 independent measurements). (C) SDS-PAGE analysis: α-crystallin (10 μM) was incubated with different concentrations of αA66-80 (1.25 μM- 20 μM) in 1ml of 50 mM phosphate buffer (pH 7.2) at 37°C for 24 h. After incubation, samples were centrifuged at 8000 rpm for 15 min and pellets were dissolved in SDS-PAGE sample buffer and run in 4–20% gel. α: αA66-80 ratio - Lane 1 - 1: 0 μM, Lane 2 - 1: 0.125 μM, Lane 3 - 1: 0.25 μM, Lane 4 - 1: 0.5 μM, Lane 5 - 1: 1 μM, Lane 6 - 1: 1.5 μM, Lane 7 - 1: 2 μM.
Figure 4.

The interaction between α-crystallin and the αA66-80 peptide analyzed by (A) tryptophan fluorescence measurements. The arrow points to the decrease in intrinsic tryptophan fluorescence of α-crystallin in the presence of increasing concentrations of peptide. (B) Far-UV CD spectra. The signal intensity decreases, in concomitance with the formation of insoluble precipitate. (C) Size exclusion chromatography. Chromatograms represent the decrease in the soluble α-crystallin fraction in the presence of αA66-80 peptide.
αA66-80 Peptide Interaction Increases the Surface Hydrophobicity of α-Crystallin
The hallmark of aggregating proteins is their exposed hydrophobic groups.13 Therefore, to determine if a concomitant change in hydrophobicity occurs during the peptide binding, bis-ANS binding experiments were performed. Figure 5 illustrates the emission spectra of α-crystallin in the presence and absence of the peptide. The fluorescence intensity of bis-ANS in α-crystallin samples gradually increased in the presence of increasing concentrations of αA66-80 peptide, indicating the increased availability of hydrophobic sites on α-crystallin upon interaction with the peptide.
Figure 5.

Changes in surface hydrophobicity of α-crystallin in the presence of peptide. Emission spectrum of bis-ANS (20 μM) samples bound to α-crystallin in the presence and absence of αA66-80 peptide. Arrow indicates a gradual increase in Bis-ANS fluorescence intensity bound to α-crystallin with increase in αA66-80 peptide concentration.
Non-covalent Interactions Stabilize α-Crystallin-αA66-80 Complexes
We have shown that the αA66-80 peptide is able to promote aggregation of α-crystallin. The characteristics of the α-crystallin-αA66-80 aggregate were examined by studying the effects of dissociating agents, including SDS, NaCl, GdmCl and urea, on the peptide-induced α-crystallin aggregates. The α-crystallin-αA66-80 aggregates were resistant to dissociation (solubilization) by high ionic strength buffers that contained 1 M NaCl and 2 M MgCl2 and were only partially affected by PBS-TritonX-100. Maximum solubility (about 62% to 68%) of the α-crystallin and peptide from the aggregates was observed in the presence of strong dissociating agents, urea and GdmCl (Table 1). This suggests that the non-covalent interactions stabilize the peptide-crystallin associations and contribute to the aggregation. These data, combined with the bis-ANS binding studies, indicate that intermolecular hydrophobic interactions play a crucial role in the peptide-mediated α-crystallin aggregation.
Table 1.
Stability of the peptide-protein complexes in dissociating agentsa
| Dissociating Agents | Protein recovered in soluble fraction ( % of total)b |
|---|---|
| Phosphate buffer (PBS) | 0.01 |
| PBS – 1% Tween-20 | 0.012 |
| PBS-1% Triton X-100 | 12 |
| 1 M NaCl | 0.001 |
| 2M MgCl2 | 0.001 |
| 8 M Urea | 62 |
| 1% SDS | 12 |
| 6 M Guanidium Hydrochloride | 68 |
α-crystallin and αA66-80 peptide (2:1) ratio (W/W) were incubated at 37°C for 18 h. After incubation, the samples were centrifuged (10,000 g, 10 min) and the aggregates were resuspended in different dissociating agents and placed in roller shaker at 37°C for 1 h. The samples were again centrifuged at 8000 rpm for 10 min. Protein content of the supernate was estimated by Bio-Rad protein assay. Protein content of the sample at 0 min before incubation was taken as 100%.
Bold-faced percentages indicate the maximum solubility of the aggregate in 8 M urea and 6 M guanidium hydrochloride
Site-specific Interaction Between αA66-80 and αB-crystallin
Our previous studies showed that the αA66-80 peptide brings about preferential insolubilization of α-crystallin from soluble lens extracts.19 The preferential insolubilization could be mediated by a specific interaction between the peptide and α-crystallin. To examine the interaction, we used a chemical cross-linking and mass spectrometry approach. In our experiments with α-crystallin, αA66-80 peptide induced aggregation and precipitation of both αA- and αB-crystallin. To minimize the complexity of analyzing the large number of the peptides in mass spectra arising from both αA- and αB-crystallin subunits, we focused on studying the interaction between αA66-80 and αB-crystallin by using a novel cross-linking reagent. Figure 6 depicts a schematic overview of the analytical strategy we formulated. A biotinylated αA66-80 peptide was used in our experiments to enable detection of the αB-crystallin-peptide complex in gels. The presence of biotin did not affect the interaction between the peptide and αB-crystallin. We used isotope-labeled homobifunctional NHS ester (N-hydroxy succinimide)-BS2G-d0/d4 (bis(sulfosuccinmidyl) glutarate) cross-linker to examine the peptide interaction site in αB-crystallin (Figure 7A). Homobifunctional cross-linking reagents contain two identical functional groups on either side of the molecule that are separated by a spacer bridging of a defined distance. NHS esters are highly reactive towards primary amines (i.e., ε-amino groups of lysines and the free N-terminus of a protein), but less reactive with hydroxyl groups of serine and threonine side chains.34 Upon cross linking, NHS esters created amide bonds in peptides that resulted in a mass increase of 96.021 u and 100.045 u with BS2G-d0 and BS2G-d4, respectively. BS2G was used in 1:1 mixtures of their deuterated and non-deuterated species to facilitate identification of cross-linked products by means of their distinct doublet isotope patterns with mass differences of 4.025 u (d0/d4 ) for mono protonated forms in the deconvoluted mass spectra.35 BS2G was employed in 20-fold molar excess and allowed to react for 45 min in ice, an adequate condition for the formation of a cross-linked complex between αB-crystallin and the peptide without aggregation. The presence of the αB-crystallin-αA66-80 peptide cross-linked complex was detected by western blot analysis of the SDS-PAGE gel using Avidin-HRP that binds to the biotin tag in the peptide. We observed avidin reactivity only in the sample where peptide was cross-linked to αB-crystallin (Figure 7B, lane 4). The cross-linked species migrated as two distinct bands, a trace band at 22 kDa and a strong dimer band at 44 kDa in SDS-PAGE, indicating the formation of a well-defined cross-linked complex. The cross-linked species from 22 kDa band was excised and subsequently used in enzymatic in-gel digestion with trypsin.
Figure 6.

General analytical strategy for the identification of αA66-80 peptide interaction site in αB-crystallin using isotope-labeled cross linker (BS2G d0/d4).
Figure 7.

(A) Chemical structure of amine- reactive, homobifunctional, and isotope-labeled (do and d4) cross-linking reagent BS2G - Bis (sulfosuccinimidyl) glutarate. The corresponding mass shifts associated with d0 and d4 cross-linked peptides are shown. (B) SDS-PAGE of αB-crystallin. Biotinylated αA66-80 peptide cross-linking reaction mixture with BS2G d0/d4 and western blot of similarly run gel. The reaction was carried out as described under methods with 20-fold molar excess for 45 min. SDS-PAGE and Western blot: Lane 1- Marker, Lane 2- αBWT control with cross-linker (without peptide), Lane 3- αBWT + Biotin-αA66-80 without cross-linker, Lane 4- αBWT + Biotin-αA66-80 with cross-linker. Arrows in the SDS-PAGE gel point to cross-linked complexes containing biotin-labeled peptide confirmed by western blot of the gel.
Nano-MS analysis of tryptic peptides of cross-linked products from gel bands generated numerous peptide signals. The deconvoluted peak lists from LC-MS data, when compared with theoretical mass lists generated by GPMAW (as described in Methods), revealed five inter-cross-linked species between αB-WT and αA66-80 peptide from the tryptic peptides of 22 kDa band (Table 2). Of the five intercross-linked pairs identified in LC-MS, two (identified by boldface in Table 2) were confirmed by MS/MS (Figure 8 and Figure S1). Figure 8A gives the LC-MS spectra of the intermolecular cross-linked peptide pair 1612.82/1616.84 [MH+], which appears as doubly protonated 806.91/808.93 [M2H+] ion pairs with a mass difference of 2.02Da. The identified parent ion was subjected to MS/MS analysis. Figure 8B shows the CID and HCD spectrum with a series of y-type and b-type ions observed. Fragmentation pattern analysis in MS/MS led to the identification of a cross-link between Lys-166 in αB-WT and the amino group in the N-terminal serine of αA66-80 peptide. The cross-link product has the C-terminal region of αB-WT (amino acids 164–174) attached to the αA66-80 peptide (amino acids 66–68) (Figure 8C). The fragmentation pattern of a triply charged ion pair having a mass difference of 1.34 (m/z of 622.34/623.68) is shown in Figure S1B. The pair corresponds to the cross-linked site between αB-WT (amino acids 91–103) and the αA66-80 peptide (amino acids 66–68) (Figure S1C). The other three presumptive crosslinks identified (Figure S2) were not abundant enough to perform MS/MS analysis. Figure 9 illustrates the potential αA66-80 interacting sites in αB-crystallin identified by GPMAW from the LC-MS data of the cross-linked band.
Table 2.
Identification of potential αB-crystallin and αA66-80 crosslink pairs from the LC-MS data using GPMAWa,b
| Observed mass pairs[MH+] | Intensity | Actual mass[MH+] | αB- crystallin sequence | αA66-80 sequence | type | Delta | ppm |
|---|---|---|---|---|---|---|---|
|
1612.820 1616.843 |
161427 189204 |
1612.818 | 164–174 | 1–3 | X-link | −0.002 | −1 |
| 1979.082 1983.106 |
484245 703659 |
1979.085 | 70–74 | 4–13 | X-link | 0.003 | 1 |
| 2361.165 2365.192 |
1337170 1336468 |
2361.172 | 75–90 | 1–3 | X-link | 0.007 | 3 |
| 2189.084 2193.114 |
206855 229689 |
2189.095 | 93–107 | 1–3 | X-link | 0.011 | 5 |
|
1865.009 1869.034 |
342231 596374 |
1865.013 | 91–103 | 1–3 | X-link | 0.004 | 2 |
The deconvoluted peak lists from MS-spectra were compared with theoretically generated cross-linked peptide masses. The theoretical peak lists were generated using GPMAW. The table gives the observed mass in the LC-MS, actual mass (theoretical) generated by GPMAW, the amino acid segments in the interacting partners. Delta corresponds to mass deviation between the observed and actual mass.
Mass pairs and the corresponding cross-linked sites in bold were confirmed by MS/MS.
Figure 8.

Nano-LC-LTQ Orbitrap identification of the αA66-80 peptide interaction with C-terminal extension of αB-crystallin. (A) Extracted ion chromatogram of tryptic digest of cross-linked complex. Signal of doubly charged peptides with m/z 806.91 and 808.93, cross-linked with light (d0) and heavy (d4) precursor ions presenting a mass difference of 2.02 Da is indicated by arrow. (B) Fragmentation mass spectrum (both CID and HCD) of the identified precursor ion (m/z 806.9) obtained using MS/MS. The identified y and b ions are indicated. (C) The interacting region of the N-terminal amino group of 66th residue of αA66-80 with Lys166 of αB-crystallin is shown in the identified cross-linked sequences.
Figure 9.

Amino acid sequences of αB-crystallin and biotinyl αA66-80 (SDRDKFVIFLDVKHFK-Biotin). Sequences in blue denote C-terminal extension (151–175) region in αB-crystallin. Sequences in red denote the chaperone site in αB-crystallin. The underlined sequences in αB-crystallin are presumptive peptide interaction region identified by the GPMAW from the LC-MS data. Sequences underlined in green are confirmed by MS/MS.
To confirm that the αA66-80 peptide interaction at the C-terminal extension of αB-crystallin is a specific interaction and not a random interaction, we used αB-crystallin C-terminal peptide 164–174 (αB-CTP) to titrate αA66-80 and measured the changes in surface hydrophobicity of α-crystallin with Bis-ANS. As control, we used a Proline-substituted αA66-80 peptide that does not interact with α-crystallin. The results are shown in Figure S3. The addition of αA66-80 peptide or αB-CTP increased the surface hydrophobicity of α-crystallin, whereas the Proline–substituted αA66-80 did not. The presence of both αB-CTP and αA66-80 peptides in the sample resulted in no additional increase in the surface hydrophobicity of α-crystallin. These observations suggest that interaction between αB-CTP and αA66-80 peptide prevents the αA66-80 peptide from binding to α-crystallin (Figure S3). Thus, the cross linking and dissociation studies together suggest that the αA66-80 peptide binds at multiple sites and forms a tight complex with αB-crystallin.
DISCUSSION
High-molecular weight (HMW) aggregates composed of native, modified and truncated proteins and peptide fragments are the hallmark of aging and cataract lenses.1,7,11,12,15 Recent evidence points to the involvement of crystallin fragments in the aggregation of lens proteins.19 The amyloidogenic peptide αA66-80 is known to progressively accumulate in the water-insoluble aggregates of the aging lens,12,19 and the peptide preferentially insolubilizes α-crystallin from the soluble lens fractions.19 However, it is not clear how the peptide influences the structure and function of soluble α-crystallins. Since α-crystallins constitute the major protein in the water-insoluble cataract aggregates, it is important to determine the nature of the interactions between the peptide and α-crystallins. The aims of this study were to identify the sites at which αA66-80 peptide binds to α-crystallin and the mechanism by which it induces aggregation of soluble α-crystallins. We present here the first evidence of a striking potential of the αA66-80 peptide to form a stable complex with α-crystallin and induce α-crystallin aggregation.
Impact of the αA66-80 Peptide on α-Crystallin
The αA66-80 peptide is localized and concentrated in the nuclear region of the lens.12,19 In aged human lenses (>70 year-old), the concentration of αA66-80 peptide is 2.33 ± 0.6 nmol per gram lens tissue and almost all of this is in the nuclear region.19 In addition, the lens also contains additional peptides that are capable of inducing aggregation of α-crystallin.11,12,19 In our experiments we use peptide concentrations in the μM range. Such use of high levels of peptide in in vitro experiments to demonstrate the in vivo effect of the peptide has been reasoned out earlier.19,36 The in vivo process of cataract pathogenesis and the associated accumulation of peptides takes place in a span of years to decades, whereas the time frame of the in-vitro experiments are limited. Further, the effective concentration of the αA66-80 in the lens could be increased by its interaction with other lens components and due to factors like crowding and nuclear localization of most of the peptides. In the presence of αA66-80, α-crystallin has diminished chaperone activity and aggregates into insoluble form (Figures 2 and 3). Thus, the age-related insolubility of α-crystallin specifically observed in the nuclear region of the lens and the predominant presence of α-crystallin in the water-insoluble aggregates of lens, which has been known for years, can be attributed to the interactions between peptide and soluble α-crystallins.
Rapid aggregation of soluble proteins can be due to self-association of proteins (which may or may not be accompanied by subtle conformational changes),37 leading to the formation of small oligomers or HMW soluble or insoluble aggregates38–40 as a result of interactions with peptides and small molecules.41–44 Our data provide the evidence in support of this phenomenon. First, the quenching of tryptophan fluorescence of α-crystallin by the peptide suggests that the microenvironment of intrinsic tryptophan residues in α-crystallin is altered as a consequence of peptide binding. A steady decrease in the tryptophan emission intensity, with no change in the emission maximum, in samples containing αA66-80 peptide could also indicate a compactness of the tertiary structure or the formation of aggregates.45–47
Second, α-crystallin samples incubated with αA66-80 peptide exhibited reduced peak/signal intensity in our SEC and CD experiments (Figure 4). The decrease in fluorescence intensity and CD signal, coupled with the reduction in peak intensity in SEC measurements, suggests that the peptide-induced α-crystallin aggregates are insoluble and hence are lost as precipitates. Such aggregation behavior is due to rapid HMW aggregate formation and precipitation, resulting in particle formation with no detectable intermediates.45,48
Third, the bis-ANS signal intensity of α-crystallin was enhanced in the presence of the αA66-80 peptide, demonstrating a conformational change and exposure of hydrophobic residues. This finding correlates well with earlier studies which demonstrated that HMW aggregates in aged and cataract lenses result from exposure of buried β-pleated sheets and increased hydrophobic interactions.49,50 In both intrinsic tryptophan fluorescence measurement and Bis-ANS binding experiments, we did not see any shift in the emission maximum. tryptophan fluorescence quenching with no shift in emission maximum under the conditions of aggregation has been reported earlier.46,51 Increase in Bis-ANS intensity with no shift in emission maximum has been observed for mutant crystallin associated with congenital cataract.52 The apolarity implied in our Bis-ANS studies is also relatively small when compared to large increase in Bis-ANS intensity seen in “molten globule” forms where the tertiary structure is substantially disturbed.53 We therefore conclude that the marked aggregation of α-crystallin by the peptide occurs through minor structural alterations.
The peptide–α-crystallin complexes are stabilized by strong non-covalent interactions because they are partially disrupted and dissociated only by chaotropic agents (6 M GdmCl and 8 M urea) (Table 1), agents known to solubilize amyloid cores in the Alzheimer’s brain.54 We were unable to achieve 100% solubilization of aggregates in the presence of urea and GdmCl. At this time we do not know the reasons for this. A myriad of such non-covalent interactions govern the stability of amyloidal and other protein aggregates in neurodegenerative diseases, suggesting a mechanistic similarity among the disease-causing peptides. Such interactions have been shown to enhance the rate of aggregation and have been identified in the process of both physiological and pathological aggregation of proteins.38
Peptides have specific interaction sites on crystallins.18,55 Using isotope-labeled cross linking and mass spectrometry, we identified residues 70–74, 75–90, 91–103, 93–107 and 164–174 of αB-crystallin as specific αA66-80 peptide interaction sites (Figure 9). These regions in αB-crystallin are important for their oligomerization, chaperone function and solubility. In earlier studies we found that residues 73–92 in αB-crystallin are involved in chaperone function.56,57 The flexible C-terminal extension of αB-crystallin (residues 164–175) is shown to be important in the solubility, chaperone activity and the oligomeric assembly of the α-crystallin molecule.58–60 Other interaction sites identified are important for the chaperone ability and sub-unit interaction.59 Thus, the peptide interaction at the C-terminal extension, chaperone site and subunit interaction site might have led to decreased flexibility and diminished chaperone ability, affecting oligomerization and resulting in decreased solubilization potential of α-crystallin.
Mechanism of αA66-80 Peptide-Induced Aggregation
Based on our results, we hypothesize that peptide-induced crystallin aggregation occurs through the following mechanisms: Peptide binding to α-crystallin results in decreased solubility and increased surface hydrophobicity. This acts as the key trigger in the self-association/aggregation of protein-peptide complexes into HMW aggregates stabilized by non-covalent interactions. Many such hydrophobic site-mediated nuclei then coalesce together and the aggregates increase in size (Figure 10). This view is supported by our time lapse microscopic recordings that show the formation and growth of HMW aggregates from the nucleus of the α-crystallin–αA66-80 complex.19 The events may prevent the association of α-crystallin with a denatured substrate, and thus suppress α-crystallin chaperone activity (Figure 2) and allow accumulation of unchaperoned, misfolded proteins and their aggregation. In support of this mechanism are our chaperone assays that demonstrated the appearance of peptide, substrate and α-crystallin chaperone as insoluble complexes in the aggregate pellet (Figure 2 inset). Thus the peptide-mediated interactions alter the conformational characteristics and facilitate the aggregation of α-crystallin. Such aggregation and insolubilization of soluble proteins by peptides with amyloidogenic potential has been reported earlier.23,24,46
Figure 10.
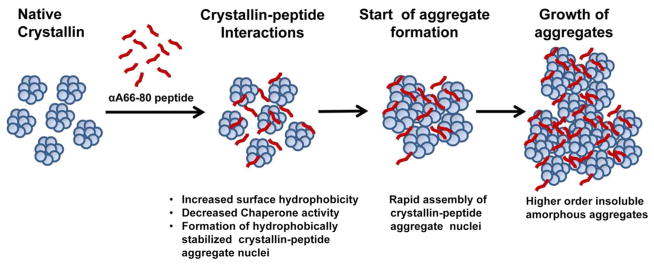
Proposed mechanism of peptide-induced aggregation of soluble crystallins. Stable non-covalent interactions between the αA66-80 peptide and crystallins increases surface hydrophobicity and facilitate formation of aggregation nuclei. The rapid assembly of nuclei with soluble crystallins favors formation of large aggregates.
Biological Relevance of Peptide-Induced Aggregation
The data presented here, combined with our previous work, provide strong support of the hypothesis that peptide interactions with soluble crystallins may be an important contributor in inducing or facilitating protein aggregation and cataract formation. We have developed a model that places the interaction of peptide and α-crystallin in the context of protein aggregation and cataract formation. The aggregation behavior of proteins is influenced by their solution environment and their stability in the native state.13 In aging lenses, the solution environment is significantly altered. Slow and progressive degradation of crystallins results in the generation of peptides. The imbalance between the production and the clearance of peptides could result in peptide accumulation in aging lenses.7 The accumulating peptides could alter the aggregation behavior of soluble lens proteins as a result of specific interactions with crystallins. Additionally, the interactions may limit the access of these peptides to peptidases, resulting in further accumulation of peptides.
Structural studies on a large number of peptide-protein complexes have shown that the association of peptide to protein causes non-local long-range effects. For example, antigenic peptide binding to MHC induces conformational changes in the MHC molecule, which in turn facilitates or decreases the binding of other proteins to the MHC molecule.61 The peptide binding could thus alter the intra- and intermolecular interactions among crystallins. Altered interactions disturb the spatial arrangement of the proteins necessary for lens transparency, which in turn leads to fluctuations of protein density and refractive index, culminating in increased light scattering.4 In addition to altering the crystallin-crystallin interactions, the insoluble form of peptide-protein complex could act as a template for the conversion of soluble components of the lens into the aggregates. The peptides or the peptide-protein complexes may initiate aberrant interactions with other cellular components, resulting in impairment of cellular function. In vitro evidence suggests that these peptides are capable of interacting with membranes as well as modulating crystallin-membrane interactions.21,62 These protein-peptide aggregates may recruit other proteins and peptides and thus become resistant to degradation or clearance. As with other polymerization reactions, such as those at play in Alzheimer and other tauopathies, peptide fragments in the lens, either alone or in complex with crystallins, could act as a seed for the growth of larger aggregates, consistent with the observation that the peptide fragments are associated with the insoluble aggregates. Thus, a steady accumulation of the LMW peptides initiates a cascade that results in aggregate formation and aggregate growth in the lens, causing an increase of light scattering and leading to loss of vision and cataract. Therefore, mechanisms and strategies to inhibit peptide generation and peptide interactions could delay or prevent the onset of age-related cataract.
There are several critical questions that remain regarding peptide-induced crystallin aggregation and its relationship to cataract formation. What is the in vivo significance of the observed effects that peptides exhibit in in-vitro? In addition to αA66-80, do other peptides with amyloidogenic potential exist in the lens? Do specific peptides have particular contributions to the aggregation of lens proteins? If so, what exactly are the components that form the nucleating complex? What proteases or mechanisms are responsible for the generation of peptides and why and when do the peptides get cleaved from the crystallins during the aging process? Recently we have identified endogenous proteases in bovine and human lenses with the potential to generate αA66-80 peptide63 and studies on endogenous proteases will likely shed some light on in vivo generation of the peptides. To determine whether peptide-crystallin interactions have physiological significance, the challenge now is to explore in more detail how responses of crystallins to accumulating peptides are coupled to those manifested via other environmental insults. It is of clinical interest that aggregation can be avoided by inhibiting the proteases that generate the peptide fragments. Developing a way to inhibit protease activity may open a novel window to suppress or delay age-related cataract formation.
Supplementary Material
Acknowledgments
Funding Source Statement. Supported by National Institutes of Health grant EY 019878 and an unrestricted grant-in-aid from Research to Prevent Blindness to the Department of Ophthalmology.
We thank Sharon Morey for help in the preparation of the manuscript.
Abbreviations
- ADH
Alcohol dehydrogenase
- HMW
high molecular weight
- LMW
low molecular weight
- WIS
water-insoluble
- WS
water-soluble
- Bis-ANS
4,4′-bis( 1-anilinonapthalene 8- sulfonate)
- GdmCl
guanidium hydrochloride
- CD
circular dichroism
- SEC
size exclusion chromatography
- Aβ
beta-amyloid
- NHS
N-hydroxy succinimide esters
- BS2G d0/d4
Bis (sulfo succindimydyl)glutarate)- non-deuterated/deuterated
- DMSO
dimethyl sulfoxide
- PVDF
polyvinylidene difluoride
- HRP
horse radish peroxidase
- GPMAW
general protein mass analysis for Windows
- CID
collision-induced dissociation
- HCD
high energy C-trap collision-induced dissociation
Footnotes
SUPPORTING INFORMATION AVAILABLE
LC-MS and MS/MS spectra of additional cross-linked peptides identified and the data supporting the specific interaction between αA66-80 peptide and the C-terminal extension of αB-crystallin are given under supporting info.
The above mentioned supporting information can be accessed free of charge at http://pubs.acs.org.
References
- 1.Bloemendal H, de Jong W, Jaenicke R, Lubsen NH, Slingsby C, Tardieu A. Ageing and vision: structure, stability and function of lens crystallins. Prog Biophys Mol Biol. 2004;86:407–485. doi: 10.1016/j.pbiomolbio.2003.11.012. [DOI] [PubMed] [Google Scholar]
- 2.Chiti F, Dobson CM. Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem. 2006;75:333–366. doi: 10.1146/annurev.biochem.75.101304.123901. [DOI] [PubMed] [Google Scholar]
- 3.Horwitz J. Alpha-crystallin can function as a molecular chaperone. Proc Natl Acad Sci U S A. 1992;89:10449–10453. doi: 10.1073/pnas.89.21.10449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Takemoto L, Sorensen CM. Protein-protein interactions and lens transparency. Exp Eye Res. 2008;87:496–501. doi: 10.1016/j.exer.2008.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hooi MY, Truscott RJ. Racemisation and human cataract. D-Ser, D-Asp/Asn and D-Thr are higher in the lifelong proteins of cataract lenses than in age-matched normal lenses. Age (Dordr) 2011;33:131–141. doi: 10.1007/s11357-010-9171-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hains PG, Truscott RJ. Post-translational modifications in the nuclear region of young, aged, and cataract human lenses. J Proteome Res. 2007;6:3935–3943. doi: 10.1021/pr070138h. [DOI] [PubMed] [Google Scholar]
- 7.Sharma KK, Santhoshkumar P. Lens aging: effects of crystallins. Biochim Biophys Acta. 2009;1790:1095–1108. doi: 10.1016/j.bbagen.2009.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Harrington V, Srivastava OP, Kirk M. Proteomic analysis of water insoluble proteins from normal and cataractous human lenses. Mol Vis. 2007;13:1680–1694. [PubMed] [Google Scholar]
- 9.Grey AC, Schey KL. Age-related changes in the spatial distribution of human lens alpha-crystallin products by MALDI imaging mass spectrometry. Invest Ophthalmol Vis Sci. 2009;50:4319–4329. doi: 10.1167/iovs.09-3522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Roy D, Spector A. Human insoluble lens protein. II Isolation and characterization of a 9600 dalton polypeptide. Exp Eye Res. 1978;26:445–459. doi: 10.1016/0014-4835(78)90131-8. [DOI] [PubMed] [Google Scholar]
- 11.Santhoshkumar P, Udupa P, Murugesan R, Sharma KK. Significance of interactions of low molecular weight crystallin fragments in lens aging and cataract formation. J Biol Chem. 2008;283:8477–8485. doi: 10.1074/jbc.M705876200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Su SP, McArthur JD, Andrew Aquilina J. Localization of low molecular weight crystallin peptides in the aging human lens using a MALDI mass spectrometry imaging approach. Exp Eye Res. 2010;91:97–103. doi: 10.1016/j.exer.2010.04.010. [DOI] [PubMed] [Google Scholar]
- 13.Stefani M, Dobson CM. Protein aggregation and aggregate toxicity: new insights into protein folding, misfolding diseases and biological evolution. J Mol Med (Berl) 2003;81:678–699. doi: 10.1007/s00109-003-0464-5. [DOI] [PubMed] [Google Scholar]
- 14.Stefani M. Structural features and cytotoxicity of amyloid oligomers: implications in Alzheimer’s disease and other diseases with amyloid deposits. Prog Neurobiol. 2012;99:226–245. doi: 10.1016/j.pneurobio.2012.03.002. [DOI] [PubMed] [Google Scholar]
- 15.Srivastava OP. Age-related increase in concentration and aggregation of degraded polypeptides in human lenses. Exp Eye Res. 1988;47:525–543. doi: 10.1016/0014-4835(88)90092-9. [DOI] [PubMed] [Google Scholar]
- 16.Harrington V, McCall S, Huynh S, Srivastava K, Srivastava OP. Crystallins in water soluble-high molecular weight protein fractions and water insoluble protein fractions in aging and cataractous human lenses. Mol Vis. 2004;10:476–489. [PubMed] [Google Scholar]
- 17.Surolia I, Sinha S, Sarkar DP, Reddy PY, Reddy GB, Surolia A. Concurrence of Danish dementia and cataract: insights from the interactions of dementia associated peptides with eye lens alpha-crystallin. PLoS One. 2008;3:e2927. doi: 10.1371/journal.pone.0002927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Udupa EG, Sharma KK. Effect of oxidized betaB3-crystallin peptide on lens betaL-crystallin: interaction with betaB2-crystallin. Invest Ophthalmol Vis Sci. 2005;46:2514–2521. doi: 10.1167/iovs.05-0031. [DOI] [PubMed] [Google Scholar]
- 19.Santhoshkumar P, Raju M, Sharma KK. alphaA-crystallin peptide SDRDKFVIFLDVKHF accumulating in aging lens impairs the function of alpha-crystallin and induces lens protein aggregation. PLoS One. 2011;6:e19291. doi: 10.1371/journal.pone.0019291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Su SP, McArthur JD, Truscott RJ, Aquilina JA. Truncation, cross-linking and interaction of crystallins and intermediate filament proteins in the aging human lens. Biochim Biophys Acta. 2011;1814:647–656. doi: 10.1016/j.bbapap.2011.03.014. [DOI] [PubMed] [Google Scholar]
- 21.Su SP, McArthur JD, Friedrich MG, Truscott RJ, Aquilina JA. Understanding the alpha-crystallin cell membrane conjunction. Mol Vis. 2011;17:2798–2807. [PMC free article] [PubMed] [Google Scholar]
- 22.Srivastava OP, Srivastava K. In-vivo existing human lens crystallin fragments form amyloid-type aggregates in vitro. ARVO. 2012;53:E-Abstract 2278. [Google Scholar]
- 23.Konno T. Amyloid-induced aggregation and precipitation of soluble proteins: an electrostatic contribution of the Alzheimer’s beta(25–35) amyloid fibril. Biochemistry. 2001;40:2148–2154. doi: 10.1021/bi002156h. [DOI] [PubMed] [Google Scholar]
- 24.Olzscha H, Schermann SM, Woerner AC, Pinkert S, Hecht MH, Tartaglia GG, Vendruscolo M, Hayer-Hartl M, Hartl FU, Vabulas RM. Amyloid-like aggregates sequester numerous metastable proteins with essential cellular functions. Cell. 2011;144:67–78. doi: 10.1016/j.cell.2010.11.050. [DOI] [PubMed] [Google Scholar]
- 25.Sharma KK, Kumar RS, Kumar GS, Quinn PT. Synthesis and characterization of a peptide identified as a functional element in alphaA-crystallin. J Biol Chem. 2000;275:3767–3771. doi: 10.1074/jbc.275.6.3767. [DOI] [PubMed] [Google Scholar]
- 26.Santhoshkumar P, Sharma KK. Inhibition of amyloid fibrillogenesis and toxicity by a peptide chaperone. Mol Cell Biochem. 2004;267:147–155. doi: 10.1023/b:mcbi.0000049373.15558.b8. [DOI] [PubMed] [Google Scholar]
- 27.Roy D, Spector A. Absence of low-molecular-weight alpha crystallin in nuclear region of old human lenses. Proc Natl Acad Sci U S A. 1976;73:3484–3487. doi: 10.1073/pnas.73.10.3484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Truscott RJ, Augusteyn RC. Changes in human lens proteins during nuclear cataract formation. Exp Eye Res. 1977;24:159–170. doi: 10.1016/0014-4835(77)90256-1. [DOI] [PubMed] [Google Scholar]
- 29.Sharma KK, Kaur H, Kester K. Functional elements in molecular chaperone alpha-crystallin: identification of binding sites in alpha B-crystallin. Biochem Biophys Res Commun. 1997;239:217–222. doi: 10.1006/bbrc.1997.7460. [DOI] [PubMed] [Google Scholar]
- 30.Santhoshkumar P, Murugesan R, Sharma KK. Deletion of (54)FLRAPSWF(61) residues decreases the oligomeric size and enhances the chaperone function of alphaB-crystallin. Biochemistry. 2009;48:5066–5073. doi: 10.1021/bi900085v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sreelakshmi Y, Santhoshkumar P, Bhattacharyya J, Sharma KK. AlphaA-crystallin interacting regions in the small heat shock protein, alphaB-crystallin. Biochemistry. 2004;43:15785–15795. doi: 10.1021/bi048151s. [DOI] [PubMed] [Google Scholar]
- 32.Sharma KK, Kumar GS, Murphy AS, Kester K. Identification of 1,1′-bi(4-anilino)naphthalene-5,5′-disulfonic acid binding sequences in alpha-crystallin. J Biol Chem. 1998;273:15474–15478. doi: 10.1074/jbc.273.25.15474. [DOI] [PubMed] [Google Scholar]
- 33.Raman B, Ramakrishna T, Rao CM. Rapid refolding studies on the chaperone-like alpha-crystallin. Effect of alpha-crystallin on refolding of beta- and gamma-crystallins. J Biol Chem. 1995;270:19888–19892. doi: 10.1074/jbc.270.34.19888. [DOI] [PubMed] [Google Scholar]
- 34.Madler S, Bich C, Touboul D, Zenobi R. Chemical cross-linking with NHS esters: a systematic study on amino acid reactivities. J Mass Spectrom. 2009;44:694–706. doi: 10.1002/jms.1544. [DOI] [PubMed] [Google Scholar]
- 35.Ihling C, Schmidt A, Kalkhof S, Schulz DM, Stingl C, Mechtler K, Haack M, Beck-Sickinger AG, Cooper DM, Sinz A. Isotope-labeled cross-linkers and Fourier transform ion cyclotron resonance mass spectrometry for structural analysis of a protein/peptide complex. J Am Soc Mass Spectrom. 2006;17:1100–1113. doi: 10.1016/j.jasms.2006.04.020. [DOI] [PubMed] [Google Scholar]
- 36.Hu X, Crick SL, Bu G, Frieden C, Pappu RV, Lee JM. Amyloid seeds formed by cellular uptake, concentration, and aggregation of the amyloid-beta peptide. Proc Natl Acad Sci U S A. 2009;106:20324–20329. doi: 10.1073/pnas.0911281106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Rousseau F, Schymkowitz J, Serrano L. Protein aggregation and amyloidosis: confusion of the kinds? Curr Opin Struct Biol. 2006;16:118–126. doi: 10.1016/j.sbi.2006.01.011. [DOI] [PubMed] [Google Scholar]
- 38.Wang W, Nema S, Teagarden D. Protein aggregation--pathways and influencing factors. Int J Pharm. 2010;390:89–99. doi: 10.1016/j.ijpharm.2010.02.025. [DOI] [PubMed] [Google Scholar]
- 39.Weiss WFt, Young TM, Roberts CJ. Principles, approaches, and challenges for predicting protein aggregation rates and shelf life. J Pharm Sci. 2009;98:1246–1277. doi: 10.1002/jps.21521. [DOI] [PubMed] [Google Scholar]
- 40.Roberts CJ. Non-native protein aggregation kinetics. Biotechnol Bioeng. 2007;98:927–938. doi: 10.1002/bit.21627. [DOI] [PubMed] [Google Scholar]
- 41.Uversky VN, Li J, Fink AL. Metal-triggered structural transformations, aggregation, and fibrillation of human alpha-synuclein. A possible molecular NK between Parkinson’s disease and heavy metal exposure. J Biol Chem. 2001;276:44284–44296. doi: 10.1074/jbc.M105343200. [DOI] [PubMed] [Google Scholar]
- 42.Shalova IN, Asryants RA, Sholukh MV, Saso L, Kurganov BI, Muronetz VI, Izumrudov VA. Interaction of polyanions with basic proteins, 2(a) : influence of complexing polyanions on the thermo-aggregation of oligomeric enzymes, Macromol. Biosci. 2005;5:1184–1192. doi: 10.1002/mabi.200500142. [DOI] [PubMed] [Google Scholar]
- 43.Ahn HJ, Zamolodchikov D, Cortes-Canteli M, Norris EH, Glickman JF, Strickland S. Alzheimer’s disease peptide beta-amyloid interacts with fibrinogen and induces its oligomerization. Proc Natl Acad Sci U S A. 2010;107:21812–21817. doi: 10.1073/pnas.1010373107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Artemova N, Stein-Margolina V, Smirnova E, Gurvits B. Formation of supramolecular structures of a native-like protein in the presence of amphiphilic peptides: Variations in aggregate morphology. FEBS Lett. 2012;586:186–190. doi: 10.1016/j.febslet.2011.12.017. [DOI] [PubMed] [Google Scholar]
- 45.Malik R, Roy I. Probing the mechanism of insulin aggregation during agitation. Int J Pharm. 2011;413:73–80. doi: 10.1016/j.ijpharm.2011.04.024. [DOI] [PubMed] [Google Scholar]
- 46.Artemova NV, Stein-Margolina VA, Bumagina ZM, Gurvits BY. Acceleration of protein aggregation by amphiphilic peptides: transformation of supramolecular structure of the aggregates. Biotechnol Prog. 2011;27:846–854. doi: 10.1002/btpr.574. [DOI] [PubMed] [Google Scholar]
- 47.Duy C, Fitter J. How aggregation and conformational scrambling of unfolded states govern fluorescence emission spectra. Biophys J. 2006;90:3704–3711. doi: 10.1529/biophysj.105.078980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Golub N, Meremyanin A, Markossian K, Eronina T, Chebotareva N, Asryants R, Muronets V, Kurganov B. Evidence for the formation of start aggregates as an initial stage of protein aggregation. FEBS Lett. 2007;581:4223–4227. doi: 10.1016/j.febslet.2007.07.066. [DOI] [PubMed] [Google Scholar]
- 49.Fujii N, Awakura M, Takemoto L, Inomata M, Takata T, Fujii N, Saito T. Characterization of alphaA-crystallin from high molecular weight aggregates in the normal human lens. Mol Vis. 2003;9:315–322. [PubMed] [Google Scholar]
- 50.Liang JJ, Akhtar NJ. Human lens high-molecular-weight alpha-crystallin aggregates. Biochem Biophys Res Commun. 2000;275:354–359. doi: 10.1006/bbrc.2000.3306. [DOI] [PubMed] [Google Scholar]
- 51.Jiang W, Han Y, Zhou R, Zhang L, Liu C. DNA is a template for accelerating the aggregation of copper, zinc superoxide dismutase. Biochemistry. 2007;46:5911–5923. doi: 10.1021/bi062234m. [DOI] [PubMed] [Google Scholar]
- 52.Singh D, Raman B, Ramakrishna T, Rao Ch M. The cataract-causing mutation G98R in human alphaA-crystallin leads to folding defects and loss of chaperone activity. Mol Vis. 2006;12:1372–1379. [PubMed] [Google Scholar]
- 53.Matulis D, Lovrien R. 1-Anilino-8-naphthalene sulfonate anion-protein binding depends primarily on ion pair formation. Biophys J. 1998;74:422–429. doi: 10.1016/S0006-3495(98)77799-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc Natl Acad Sci U S A. 1985;82:4245–4249. doi: 10.1073/pnas.82.12.4245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Rao G, Santhoshkumar P, Sharma KK. Anti-chaperone betaA3/A1(102–117) peptide interacting sites in human alphaB-crystallin. Mol Vis. 2008;14:666–674. [PMC free article] [PubMed] [Google Scholar]
- 56.Bhattacharyya J, Padmanabha Udupa EG, Wang J, Sharma KK. Mini-alphaB-crystallin: a functional element of alphaB-crystallin with chaperone-like activity. Biochemistry. 2006;45:3069–3076. doi: 10.1021/bi0518141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Santhoshkumar P, Sharma KK. Conserved F84 and P86 residues in alphaB-crystallin are essential to effectively prevent the aggregation of substrate proteins. Protein Sci. 2006;15:2488–2498. doi: 10.1110/ps.062338206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Smulders R, Carver JA, Lindner RA, van Boekel MA, Bloemendal H, de Jong WW. Immobilization of the C-terminal extension of bovine alpha-crystallin reduces chaperone-like activity. J Biol Chem. 1996;271:29060–29066. doi: 10.1074/jbc.271.46.29060. [DOI] [PubMed] [Google Scholar]
- 59.Pasta SY, Raman B, Ramakrishna T, Rao Ch M. Role of the C-terminal extensions of alpha-crystallins. Swapping the C-terminal extension of alpha-crystallin to alphaB-crystallin results in enhanced chaperone activity. J Biol Chem. 2002;277:45821–45828. doi: 10.1074/jbc.M206499200. [DOI] [PubMed] [Google Scholar]
- 60.Kallur LS, Aziz A, Abraham EC. C-Terminal truncation affects subunit exchange of human alphaA-crystallin with alphaB-crystallin. Mol Cell Biochem. 2008;308:85–91. doi: 10.1007/s11010-007-9615-2. [DOI] [PubMed] [Google Scholar]
- 61.London N, Movshovitz-Attias D, Schueler-Furman O. The structural basis of peptide-protein binding strategies. Structure. 2010;18:188–199. doi: 10.1016/j.str.2009.11.012. [DOI] [PubMed] [Google Scholar]
- 62.Friedrich MG, Lam J, Truscott RJ. Degradation of an old human protein: age-dependent cleavage of gammaS-crystallin generates a peptide that binds to cell membranes. J Biol Chem. 2012;287:39012–39020. doi: 10.1074/jbc.M112.391565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hariharapura R, Santhoshkumar P, Krishna Sharma K. Profiling of lens protease involved in generation of alphaA-66–80 crystallin peptide using an internally quenched protease substrate. Exp Eye Res. 2013;109:51–59. doi: 10.1016/j.exer.2013.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.