

Abstract
Plant organ development and polarity establishment is mediated by the action of several transcription factors. Among these, the KANADI (KAN) subclade of the GARP protein family plays important roles in polarity-associated processes during embryo, shoot and root patterning. In this study, we have identified a set of potential direct target genes of KAN1 through a combination of chromatin immunoprecipitation/DNA sequencing (ChIP-Seq) and genome-wide transcriptional profiling using tiling arrays. Target genes are over-represented for genes involved in the regulation of organ development as well as in the response to auxin. KAN1 affects directly the expression of several genes previously shown to be important in the establishment of polarity during lateral organ and vascular tissue development. We also show that KAN1 controls through its target genes auxin effects on organ development at different levels: transport and its regulation, and signaling. In addition, KAN1 regulates genes involved in the response to abscisic acid, jasmonic acid, brassinosteroids, ethylene, cytokinins and gibberellins. The role of KAN1 in organ polarity is antagonized by HD-ZIPIII transcription factors, including REVOLUTA (REV). A comparison of their target genes reveals that the REV/KAN1 module acts in organ patterning through opposite regulation of shared targets. Evidence of mutual repression between closely related family members is also shown.
Introduction
Plants achieve their final shoot architecture through the proper positioning of lateral organs such as leaves and flowers. In part this is mediated by the polar transport of the plant hormone auxin to specific locations, which then triggers organ initiation at these sites. The subsequent differentiation of organ progenitor cells into more specialized cell types results in highly organized tissues made up of many distinct cell types. The KAN subclade of the GARP family of transcription factors, as well as the set of class III homeodomain leucine zipper (HD-ZIPIII) transcription factors, play important roles in polarity-associated patterning processes. These transcription factors are key determinants in embryo, shoot and root patterning and during vegetative growth regulate several organ polarity processes [1-15]. In particular, during leaf development these two gene families have been shown to act antagonistically to maintain a stable abaxial/adaxial boundary (the boundary between the lower and upper side of the leaf) that is necessary for proper leaf blade growth. Here, the four members of the KAN group (KAN1-4) are required for abaxial cell fate, whereas the HD-ZIPIII genes, including PHABULOSA (PHB), PHAVOLUTA (PHV) and REV, promote adaxial cell identity in organ primordia [1,3,4,7,12-14].
Genetic studies have identified additional regulatory factors specifying the abaxial/adaxial sides of the leaf. The ASYMMETRIC LEAVES2 (AS2) gene, a LOB domain–containing plant-specific protein, and the ASYMMETRIC LEAVES1 (AS1) gene, a MYB domain transcription factor, are involved in the development of a symmetrical expanded lamina, and act to promote adaxial (upper leaf) fate in this regulatory network [16-18]. On the opposite side, members of the YABBY (YAB) gene family, such as FILAMENTOUS FLOWER (FIL), YAB3, YAB5 and YAB2, and two AUXIN RESPONSE FACTOR genes (ETTIN (ETT)/ARF3 and ARF4), specify abaxial (lower leaf) cell fate [19-23]. In addition to this set of transcription factors, small RNAs have also been found to play crucial roles in the establishment of organ polarity. HD-ZIPIII factors are targeted by microRNAs 165/166, which therefore act as abaxial determinants [24-27]. The ARF3 and ARF4 genes are controlled by the ta-siRNAs ta-siR2141 and ta-siR2142 (also referred as ta-siR-ARFs), thus implicating tasiR-ARFs as important adaxial regulators [27,28].
Genetic analysis indicates that some of these genes act antagonistically: loss-of-function mutations in genes promoting adaxial development typically produce an abaxialized phenotype that is accompanied by the expanded expression of abaxial genes, whereas loss-of-function mutations in abaxial genes produce an adaxialized phenotype that correlates with the expanded expression of adaxial genes. Transgenes or mutations that cause ectopic expression of these genes, usually lead to phenotypes opposite to that of the loss-of-function mutations. The antagonistic relationship between the adaxial and abaxial transcription factors could be mediated by direct cross regulation of each other's expression, or alternatively but not mutually exclusively, via opposite effects on common downstream targets of biochemical processes, both of which have been postulated [7,10,29-31]. One strategy to understand how transcription factors mediate their developmental functions is to identify the genes they directly regulate. In this study, we focus mainly on identifying KAN1 targets and, in addition, define potential shared targets between the abaxial factor KAN1 and the adaxial-fate promoting factor REV.
Up to now, only a small number of REV and KAN1 target genes have been reported. For instance, the LITTLE ZIPPER (ZPR) genes have been proposed as direct REV targets since they are transcriptionally up-regulated by REV and other HD-ZIPIII transcription factors. Furthermore, ZPR proteins interact with and repress HD-ZIPIII activity, forming a negative feedback loop [32,33]. Recently, we demonstrated that REV acts upstream of several class II HD-ZIP transcription factors (HAT2, HAT3, ATHB2/HAT4 and ATHB4) involved in shade signaling and leaf development [29,34], and the auxin biosynthetic enzymes TAA1 and YUCCA5 (YUC5). Expression of HAT2, TAA1 and YUC5 is reduced significantly by dexamethasone (DEX) in inducible KAN1 overexpression lines (35S::FLAG-GR-KAN1), indicating that at least one way to establish the leaf adaxial-abaxial pattern by the REV/KAN module is through the opposite regulation of shared target genes [29]. In addition, KAN activity has been proposed to negatively regulate PIN expression, and hence auxin movement, based on the ectopic expression of PIN1 in kan loss-of-function alleles, and the rapid down-regulation of PIN1 expression in response to induction of ectopic KAN1 activity [7,10]. It is not known whether KAN regulation is direct or indirect, but also suggests opposing actions of KAN and HD-ZIPIII on regulation of auxin biology. The adaxial factor AS2 is the best characterized target gene of KAN1, which represses the transcription of AS2 in abaxial tissue [17,18,31]. Mutation of a single nucleotide in a KAN1 binding site in the AS2 promoter causes ectopic AS2 expression in the abaxial domain, resulting in an adaxial phenotype. Furthermore, it has been shown that the abaxial expression of KAN1 is mediated directly by AS2 [31]. Based on these results, it has been proposed that KAN1 acts as a transcriptional repressor, and that mutual repression between KAN1 and AS2 contributes to the proper establishment of abaxial/adaxial polarity in plants.
Here, we provide a set of potential target genes of the KAN1 transcription factor identified through a combination of chromatin immunoprecipitation/deep sequencing (ChIP-Seq) and genome-wide transcriptional profiling using tiling arrays. Our dataset shows a strong over-representation of genes involved in the regulation of organ development as well as in the response to hormonal stimuli. In addition, the cis-element ‘VGAATAW’ has been identified to be enriched in the ChIP-seq dataset providing the first information about the KAN1-binding site. This cis-element is also present in the promoter of the KAN1 target gene AS2 and it has been shown to be recognized by KAN1 [31], validating our ChIP-seq analysis. Finally, the identification of genes potentially dually regulated by the REV/KAN1 module enables future elucidation of different genetic networks underlying the action of these antagonistic factors.
Materials and Methods
Plant material and treatments
For efficient chromatin immunoprecipitation, transgenic 35S::FLAG-GR-KAN1 plants were used [29]. The glucocorticoid receptor (GR) was cloned in frame with the FLAG epitope in the pJAN33 vector using the KpnI restriction site [35]. Therefore, these transgenic plants can be treated with dexamethasone (DEX), inducing the transition of the chimeric FLAG-GR-KAN1 protein from the cytoplasm to the nucleus, where it can bind to DNA to regulate its downstream targets. In order to achieve equal distribution and uptake of DEX, 35S:FLAG-GR-KAN1 plants were grown in liquid culture for 10 days and induced with 25µM DEX for 45 minutes prior to chromatin-immunoprecipitation. As a control, we used wild type Columbia (Col-0) plants.
ChIP-sequencing and ChIP analysis
Chromatin extraction and immunoprecipitation (ChIP) were carried out as described by Brandt et al. (2012) [29]. In total, we constructed one control library (Col-0) and two ChIP-Seq libraries for 35S:FLAG-GR-KAN1 using the Illumina® TruSeq® ChIP Sample Preparation Kit, according to the manufacturer’s protocol. For library preparation indexing adapters were ligated to the ends of the DNA fragments (AR003 for Col-0 library and AR011 and AR027 for 35S:FLAG-GR-KAN1 libraries). Indexed libraries were subsequently subjected to deep sequencing using the Illumina HiSeq instrument. The Illumina sequencing and data analysis were performed as described by Yant and colleagues (2010) [36], with the exception that the number of duplicate sequence reads was heuristically reduced prior to further analysis. This ChIP-Seq experiment resulted in the identification of 17402 positions in the Arabidopsis genome being enriched in 35S::FLAG-GR-KAN1 plants compared with Col-0 plants. ChIP-Seq raw data obtained in this study are available at the Gene Expression Omnibus database under series accession number GSE48081.
Tiling arrays
To examine genome-wide effects of high levels of KAN1 activity, we used ubiqutious expression of a steroid-dependent KAN1 variant, 35S:KAN1-GR [25]. KAN1 protein activity was induced by growing plants on 0.5x MS plates and submerging seedlings in 10 µM dexamethasone 21-acetate solution for 5 minutes. RNA was collected at three time points: 0 minutes (pre-induction) and 80, and 160 minutes post-induction. A total of 20-30 µg total RNA per sample using the RNeasy® Plant mini Kit (Qiagen, Valencia, CA, USA) was converted into a labeled probe for hybridization to Arabidopsis Tiling 1.0 Arrays (Affymetrix) at the Australian Genome Research Facility (The Walter and Eliza Hall Institute of Medical Research, Melbourne, Australia). The results were then calibrated and pooled per time point (2-3 biological replicates per time point) according to the tiling 1.0 array manual, and the resulting .chp files where loaded versus control into the Integrated Genome Browser (version 6.7) software for analyses [37]. The transcriptional changes from baseline were graphically assessed using selected threshold values and candidates with consistent up/down regulation along the full ORF/length of the predicted expressed sequence were identified.
Semi-quantitative PCRs (sqPCR) and quantitative real-time PCRs (qPCRs)
To test the 35S:KAN1-GR line used for the tiling array experiment, AS2 and cyclophilin were assessed as positive and negative controls, respectively, by sqPCR.
RNA was extracted from 15 day old seedlings grown on MS medium and after 80 minutes of dexamethasone treatment using the RNeasy® Plant mini Kit (Qiagen, Valencia, CA, USA). 1 µg of purified RNA was treated with DNAse RQ1 (Promega, Madison, WI, USA) and reverse transcribed using PrimeScriptTM Reverse Transcriptase (TaKaRa Biotech) for sqPCR. The sqPCR was performed with three biological replicates and visualized on 1.5% agarose gels using electrophoresis [10].
To analyze the gene expression of ATHB8, RNA was isolated from 10 day old Col-0 and transgenic 35S:FLAG-GR-KAN1 seedlings after 4 hours of DEX induction. Glyceraldehyde 3-phosphate dehydrogenase (GADPH) was used as a reference gene to evaluate the amounts of mRNA (Figure S2). Real-time PCR experiments were performed as described by Brandt et al. (2012) [29].
Results
Identification of direct KAN1 target genes using ChIP-Seq
To better understand processes downstream of KAN1 action, we constructed transgenic plants over-expressing KAN1 fused to the rat glucocorticoid receptor carrying an additional FLAG-epitope (35S:FLAG-GR-KAN1). After growing these plants on soil until the first true leaves were visible, the plants were sprayed once a day for one week with 25µM DEX solution. This treatment resulted in the development of partially abaxialized leaves with drastically reduced petioles (Figure 1A), whereas untreated control plants showed no mutant phenotype. In order to achieve equal distribution and uptake of DEX, 35S:FLAG-GR-KAN1 plants were grown in liquid culture for 10 days and induced with 25µM DEX for 45 minutes prior to chromatin-immunoprecipitation. As a control, we isolated chromatin from Col-0 wild-type plants. One Illumina control library and two ChIP-Seq libraries for 35S:FLAG-GR-KAN1 were sequenced. After filtering for read quality, sequencing reads were mapped to the Arabidopsis genome (TAIR10), resulting in the identification of 17402 peaks that were enriched in two independent ChIP-Seq experiments over the control sample. We subsequently limited our analysis to peaks showing at least three-fold enrichment. This dataset contains 4183 KAN1 bound regions. From a MEME-ChIP analysis (http://www.meme.sdsc.org) a VGAATAW motif was identified in 1802 of the 4183 regions (Figure 1B), corresponding to 3151 genes potentially regulated by KAN1 (see Dataset S1). These loci were equally distributed over the five Arabidopsis chromosomes, with a lack of enriched peaks in the centromeric regions (Figure 1C). A further analysis of the distribution of the peaks relative to the gene models revealed that the majority of binding sites were located within 1.0 kb upstream of the transcriptional start site (about 24%) or 1.0 kb downstream of the coding region (about 11%). Peaks were underrepresented in gene coding regions (Figure 1D).
Figure 1. Identification of KAN1 target genes.

A) Constructing an inducible KAN1 expression system. B) Sequence logos for the cis-element, forward and reverse orientation, enriched in the ChIP-Seq dataset C) Distribution of KAN1 binding sites across the five Arabidopsis chromosomes. D) Location of peaks identified by ChIP-Seq. About 25% of all peaks are located in the first 1000bp upstream of the transcriptional start site.
Next we examined whether our identified binding site is consistent with previous findings. The recently identified as2-5d mutation carries a G to A change in the promoter of AS2, causing ectopic AS2 expression due to uncoupling from KAN1 regulation [31]. Our analysis revealed enrichment at three positions in the AS2 promoter region previously identified to be recognized by KAN1. The sequence underlying the peak in the 5’ UTR of AS2 contains the VGAATAW motif, with the G being exchanged for A in as2-5d (Figure 2A). This finding supports the idea that the 1802 binding regions containing the VGAATAW motif are recognized by KAN1 and represent genuine binding regions. Regions for which we can detect enrichment in our ChIP-Seq dataset which do not contain the VGAATAW motif might represent regions where KAN1 is associated to, maybe in complex with other DNA-binding proteins.
Figure 2. Gene-ontology analysis of KAN1 targets.
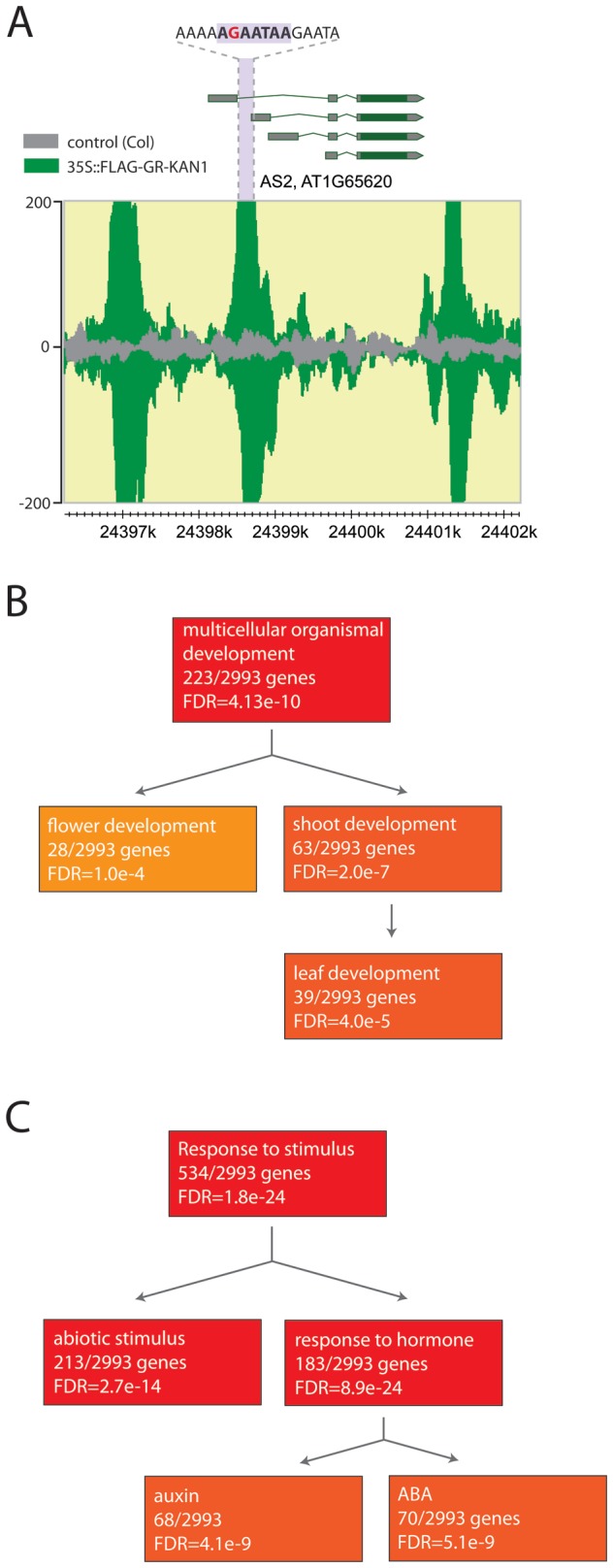
A) KAN1 binds to the ASYMMETRIC LEAVES2 (AS2) promoter. Three distinct binding regions were identified but only the second peak contains the VGAATAW motif. The guanine depicted in red is mutated to adenine in the as2-5d mutant. B) and C) Enrichment of GO terms identified in the set of genes located downstream of the KAN1-binding site. Over-representation of genes involved in multicellular organismal development and in the response to stimuli targeted by KAN1.
Taken together, we have developed an inducible system for KAN1 expression and used it to identify KAN1 binding sites across the Arabidopsis genome. Furthermore, we identified a cis-regulatory motif common to many of these targets that may represent a sequence directly recognized by KAN1.
Promoters bound by KAN1
Having identified 1802 binding regions, we were interested in investigating whether genes encoding proteins with specific functions are enriched in this dataset. We therefore performed gene ontology studies using the Agrigo tool (http:// bioinfo.cau.edu.cn/agriGO/). This analysis revealed that genes with a function in multicellular organismal development are strongly over-represented in our dataset with further enrichment in the sub-categories flower development and shoot/leaf patterning (Figure 2B). Since KAN1 is a major patterning factor and our target gene analysis revealed an enrichment of other genes involved in patterning, this dataset contains genes with a high probability to be regulated by KAN1 (Dataset S2). In addition to genes with a role in development, we also identified genes whose products have known roles in responding to stimuli (Figure 2C). Of the hormonal signaling pathways, enrichment is found for genes involved in auxin and abscisic acid signaling supporting previous findings [30].
Identification of genes transcriptionally regulated by KAN1
Having identified putative promoter regions bound by KAN1 using ChIP-seq, we next attempted to identify genes that respond transcriptionally to KAN1 activity. To this end, we utilized a line harboring a transgene resulting in widespread expression of a hormone inducible KAN1 protein, 35S:KAN1-GR [6]. When seeds homozygous for the 35S:KAN1-GR transgene were germinated in the presence of dexamethasone both shoot and root meristems were arrested, no leaf primordia were produced, and seedlings die a few weeks post germination, mimicking the phenotype of 35S:KAN1 plants [3]. As positive and negative controls we followed the expression of AS2 and cyclophilin, respectively. When assayed 80 minutes after dexamethasone treatment, expression of AS2 was reduced in hormone treated plants relative to controls, whereas cyclophilin expression was unchanged [10]. We next assayed genome-wide gene expression levels at two time points (80 minutes and 160 minutes) post-induction and identified 500 genes and 9 unannotated genomic regions in which gene expression was down-regulated at least at one of the time points (Dataset S3). In most instances down-regulation was observed at both time points, with 43 genes down-regulated only at 160 minutes and 4 genes down-regulated only at 80 minutes. Of the down-regulated genes, 42 are known to have a role in auxin biology (Dataset S4), including auxin transport or its regulation (PIN1, PIN3, PIN4, PIN7, AUX1, PGP4, PGP19, PID, BIG), auxin response (IAA2, IAA3, IAA13, IAA14, IAA16, ARF4, ARF19, HAT2), and auxin regulated genes (11 SAUR and 3 GH3 genes). Also down-regulated were 102 genes implicated in transcriptional regulation (Dataset S5), including some previously implicated in regulation of leaf polarity (e.g. PHB, YABBY5, ARF4). Some examples of each of these classes are shown in Figure S1. In contrast, up-regulation was detected at only 30 genes and 1 un-annotated region (Dataset S6). Since most potential target genes exhibited down-regulation, and KAN1 has been shown to interact with TOPLESS [38], a transcriptional co-repressor, we next identified genes that were both down-regulated and possessed local KAN1 binding sites.
A set of putative KAN1 target genes identified through ChIP-seq are also transcriptionally regulated by KAN1
Among the 3151 putative KAN1 target genes selected from the ChIP-seq data analysis, a set of 211 genes was also regulated by KAN1 at 80 and/or 160 minutes post-induction (Figure 3A and Dataset S7) in the tiling array experiments. In addition, gene ontology classification of these ChIP-seq/tiling array overlapping genes revealed again a strong over-representation of genes involved in multicellular organismal development and response to stimulus, with a significant enrichment of genes involved in shoot development and auxin response, respectively (Figure 3B). Interestingly, among 19 genes related to organ development and shoot patterning (Table 1), four genes were previously shown to be important factors in the genetic network controlling organ patterning: PHABULOSA (PHB) and ATHB8 (see also Figure S2), two class III HD-ZIP genes involved in the control of adaxial cell identity [1] and provascular patterning [1,39], respectively, MIR166F, which targets several HD-ZIPIII family members including PHV, PHB, REV, ATHB-8 and ATHB-15 [1,24,40], and PIN-FORMED 1 (PIN1), an auxin efflux carrier required for organ formation and positioning [41-43]. Moreover, several genes such as LONGIFOLIA 1 (LNG1) and LNG2 [44], the BEL1-like homeodomain protein SAW2 [45], associated with leaf shape establishment, the receptor-like kinase PXY/TDR (PHLOEM INTERCALATED WITH XYLEM/TDIF RECEPTOR), involved in the proliferation of procambial cells as well as in the maintenance of polarity during vascular tissue development [46,47] , NPY3, NPY5 (naked pins in yuc mutants) and the PINOID homolog WAG2, related to auxin-mediated organogenesis [48], were identified in both studies. Additional genes with a role in general aspects of shoot growth and development are listed in the Table 1.
Figure 3. Genome-wide comparison of genes bound and regulated by KAN1.

A) Venn-diagram showing numbers of genes bound by KAN1 and regulated by KAN1. The overlap contains 211 genes that are both bound and also regulated by KAN1. B) Gene ontology analysis of 211 potential direct KAN1 targets reveals a strong enrichment for genes involved in shoot patterning and the auxin response. Tables 1 and 2 contain these genes including the binding site information.
Table 1. Potential KAN1 target genes with a role in organ or shoot development.
|
ChIP-seq data
|
Tiling array data
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AGI | Gene Symbol | ORP-rank | Distance | Location | Enrichment replicate 1 | Enrichment replicate 2 | FDR replicate 1 | FDR replicate 2 | 80 min |
160 min
|
|
| AT1G13245 | RTFL17 | 4140 | 2414 | DOWN | 4,3 | 2,7 | 8,20E-04 | 3,93E-04 | yes | yes | |
| AT1G13260 | RAV1 | 1215 | 519 | DOWN | 5,5 | 2,7 | 2,66E-39 | 2,20E-23 | yes | yes | |
| AT1G13260 | RAV1 | 1003 | 9835 | UP | 7,8 | 3,4 | 6,93E-49 | 1,03E-23 | |||
| AT1G13260 | RAV1 | 882 | 6034 | UP | 5,1 | 2,8 | 1,54E-43 | 1,79E-33 | |||
| AT1G27320 | AHK3 | 5004 | 630 | DOWN | 4,3 | 2,1 | 8,20E-04 | 1,29E+02 | yes | yes | |
| AT1G56010 | NAC1 | 7940 | 1500 | UP | 4,1 | 2,0 | 2,30E+04 | 1,80E+07 | yes | yes | |
| AT1G73590 | PIN1 | 1344 | 1049 | DOWN | 6,9 | 3,6 | 3,37E-34 | 8,69E-23 | - | yes | |
| AT1G78240 | TSD2 | 3471 | 2653 | UP | 5,0 | 2,5 | 2,67E-11 | 4,39E-04 | yes | yes | |
| AT1G78240 | TSD2 | 6421 | 3459 | UP | 6,0 | 2,0 | 4,37E-03 | 1,80E+07 | |||
| AT1G78240 | TSD2 | 866 | 5964 | UP | 12,8 | 6,1 | 3,27E-49 | 1,04E-29 | |||
| AT1G78240 | TSD2 | 3094 | 7048 | UP | 6,7 | 2,6 | 1,98E-19 | 1,28E-02 | |||
| AT2G23760 | SAW2 | 1267 | 436 | UP | 7,2 | 3,8 | 9,78E-36 | 5,35E-25 | - | yes | |
| AT2G23760 | SAW2 | 1145 | 2496 | DOWN | 6,0 | 3,2 | 7,38E-38 | 5,57E-27 | |||
| AT2G31070 | TCP10 | 344 | 76 | UP | 8,0 | 4,3 | 6,40E-73 | 1,79E-54 | yes | yes | |
| AT2G34710 | PHB | 231 | 937 | UP | 10,2 | 4,2 | 1,91E-106 | 2,01E-52 | yes | yes | |
| AT3G14370 | WAG2 | 3554 | 1208 | UP | 7,9 | 3,5 | 1,08E-12 | 1,58E-02 | yes | yes | |
| AT5G60970 | TCP5 | 1164 | 2282 | DOWN | 7,3 | 4,4 | 1,15E-32 | 4,35E-30 | yes | yes | |
| AT5G60970 | TCP5 | 1019 | 3044 | DOWN | 7,8 | 4,9 | 2,90E-34 | 6,91E-34 | |||
| AT5G43603 | MIR166F | 2366 | 658 | UP | 3,8 | 2,7 | 2,25E-12 | 6,47E-18 | yes | yes | |
| AT5G61480 | PXY | 2404 | in CDS | 5,7 | 2,6 | 5,67E-22 | 7,40E-09 | - | yes | ||
| AT5G67440 | NPY3 | 434 | 413 | UP | 6,4 | 3,5 | 2,32E-65 | 1,24E-49 | yes | yes | |
| AT5G67440 | NPY3 | 1623 | 184 | DOWN | 5,5 | 2,5 | 1,19E-32 | 2,38E-16 | |||
| AT4G37590 | NPY5 | 979 | 1452 | UP | 9,9 | 5,0 | 2,37E-43 | 3,80E-29 | yes | yes | |
| AT3G02170 | LNG2 | 3204 | 3955 | UP | 5,3 | 2,9 | 3,08E-11 | 6,67E-07 | yes | yes | |
| AT5G15580 | LNG1 | 4944 | in CDS | 10,7 | 2,6 | 1,57E-12 | 3,27E+06 | yes | yes | ||
| AT5G61960 | AML1 | 2123 | 2369 | UP | 5,6 | 1,4 | 3,33E-34 | 4,59E-06 | yes | yes | |
| AT5G61960 | AML1 | 2265 | in CDS | 6,8 | 3,1 | 5,15E-23 | 3,05E-10 | ||||
| AT4G32880 | ATHB8 | 1402 | 561 | UP | 1,8 | 6,5 | 1,69E-30 | 3,33E-23 | yes | yes | |
| AT2G46685 | MIR166A | 4624 | 3218 | UP | 2,3 | 4,8 | 4,90E-07 | 1,49E+02 | - | - | |
| AT1G65620 | AS2 | 1750 | in CDS | 2,4 | 5,1 | 1,50E-34 | 6,53E-12 | - | - | ||
| AT1G65620 | AS2 | 2506 | 424 | DOWN | 2,4 | 5,9 | 1,72E-23 | 2,74E-06 | - | - | |
| AT5G16560 | KAN1 | 1996 | 173 | UP | 1,8 | 4,5 | 2,88E-21 | 5,12E-16 | - | - | |
| AT5G16560 | KAN1 | 2689 | 707 | UP | 2,6 | 9,3 | 8,92E-23 | 1,02E-04 | - | - | |
| AT5G16560 | KAN1 | 2930 | 5046 | UP | 2,9 | 7,8 | 8,53E-23 | 4,06E-03 | - | - | |
| AT1G32240 | KAN2 | 468 | 4442 | DOWN | 8,8 | 5 | 4,95E-61 | 4,17E-50 | - | - | |
Notes: By analyzing the ChIP-seq and the tiling array datasets and based on gene ontology (GO) analysis and literature contrast, we identified 23 genes involved in multicellular organismal development and shoot development. These genes are listed with the AGI (Arabidopsis Genome Initiative) gene code, the Gene Symbol, the ORP-rank, the distance from the binding site to the CDS, the location (UP=upstream of a gene, DOWN=downstream of a gene, in CDS), the enrichment of ChIP-seq replicates 1 and 2 (ratio of number of reads for a binding site in KAN1+DEX versus Col0+DEX), the false discovery rate (FDR) of ChIP-seq replicates 1 and 2, and the down-regulation at 80 and/or 160 min after KANADI1 activity induction (yes=the entire length of the predicted transcript was down-regulated; - no significant down-regulation).
Out of 211 genes identified as putative KAN1 targets by both the ChIP-seq and the tiling array approaches, 21 are involved in auxin response. Table 2 shows a set of KAN1 target genes encoding proteins involved in auxin signaling as well as in auxin transport. This set of genes includes several early auxin-regulated genes with a role in auxin signaling pathways such as two Aux/IAA genes (IAA2 and IAA13), which encode short-lived transcription factors that function as repressors of auxin response genes [49], three GH3 genes (DFL1, DFL2 and WES1), encoding acyl adenylate–forming isozymes that covalently modify indole-3-acetic acid (IAA) [50], and three SAUR-like genes (AT1G19840, AT1G75590 and AT2G21210), which encode short-lived nuclear proteins involved in auxin signaling by interacting with calmodulin [51,52]. Furthermore, an AUXIN RESPONSE FACTOR gene, ARF4, was identified in both experiments while ARF3/ETT was identified only in our ChIP-seq data. ARF4, together with the redundant gene ARF3/ETT (ETTIN), act to promote abaxial identity in association with KAN or its downstream targets [20]. In addition, it has been shown that the negative transcriptional, post-transcriptional and epigenetic regulation of these ARFs by AS1 and AS2 is important for the establishment of early leaf adaxial/abaxial polarity [53]. Among this set of genes, we also found the class II HD-ZIP gene HAT2, which is an early auxin-inducible gene with opposite functions in regulating auxin-mediated morphogenesis in the shoot and root tissues [54]. In a previous study, we also showed that HAT2 acts downstream of REV in the shade avoidance response [29]. Regarding those genes involved in auxin transport, two PIN genes, PIN3 and PIN4, which are important for tropic growth of the root [55] and root patterning [56], respectively, as well as for creating local auxin gradients required for the establishment of primordia and organ development [41], were found in both studies. Furthermore, a phospholipase required for PIN protein trafficking to the plasma membrane in the root (phospholipase A2; PLA2A) [57], and the PINOID protein kinase (PID), which controls PIN polarity and mediates changes in auxin flow to create local gradients for patterning processes [58], were identified. Additionally, the auxin influx transporter AUX1 and the ATP-binding cassette transporter AtMDR1 found in both studies regulate root gravitropism, and photomorphogenesis and root development, respectively, by mediating auxin polar transport [59,60].
Table 2. Potential KAN1 target genes involved in auxin response.
|
ChIP-seq data
|
Tiling array data
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AGI | Gene Symbol | ORP-rank | Distance | Location | Enrichment replicate 1 | Enrichment replicate 2 | FDR replicate 1 | FDR replicate 2 | 80 min | 160 min |
| AT1G19840 | SAUR-like | 2341 | 6906 | DOWN | 5,1 | 2,8 | 6,76E-18 | 1,05E-12 | yes | yes |
| AT1G75590 | SAUR-like | 3037 | 1915 | UP | 5,8 | 2,5 | 2,21E-17 | 1,86E-04 | yes | yes |
| AT2G21210 | SAUR-like | 157 | 149 | UP | 8,3 | 3,4 | 4,47E-119 | 9,21E-58 | yes | yes |
| AT2G33310 | IAA13 | 1242 | in CDS | 7,4 | 4,1 | 1,00E-33 | 1,71E-27 | yes | yes | |
| AT2G34650 | PID | 2130 | 3923 | UP | 5,7 | 2,8 | 6,51E-23 | 1,53E-12 | yes | yes |
| AT2G38120 | AUX1 | 158 | 10052 | UP | 7,3 | 4,2 | 3,04E-94 | 1,24E-79 | yes | yes |
| AT3G23030 | IAA2 | 165 | 171 | UP | 9,0 | 4,4 | 4,11E-103 | 3,88E-67 | yes | yes |
| AT3G28860 | ATMDR1 | 3763 | 3456 | UP | 5,0 | 2,2 | 1,49E-11 | 1,45E-01 | yes | yes |
| AT3G28860 | ATMDR1 | 3075 | 8677 | DOWN | 4,4 | 2,0 | 3,47E-16 | 8,40E-05 | ||
| AT4G03400 | DFL2 | 2385 | 2241 | DOWN | 7,4 | 3,1 | 9,71E-24 | 9,10E-08 | yes | yes |
| AT4G03400 | DFL2 | 1583 | 2654 | DOWN | 5,6 | 2,6 | 2,76E-33 | 1,08E-16 | ||
| AT4G27260 | GH3.5, WES1 | 2996 | 1074 | UP | 4,0 | 2,2 | 6,37E-13 | 1,14E-07 | yes | yes |
| AT4G27260 | GH3.5, WES1 | 5906 | 971 | DOWN | 5,0 | 1,3 | 4,55E-09 | 1,63E+07 | ||
| AT5G47370 | HAT2 | 3245 | 86 | UP | 4,6 | 2,3 | 1,64E-12 | 1,76E-05 | yes | yes |
| AT5G54510 | GH3.6, DFL1 | 2785 | 2222 | DOWN | 4,8 | 1,9 | 1,29E-22 | 1,16E-04 | yes | yes |
| AT5G59780 | MYB59 | 1240 | 5918 | UP | 5,6 | 3,3 | 2,16E-32 | 7,10E-28 | yes | yes |
| AT5G61420 | MYB28 | 1759 | 769 | UP | 6,2 | 2,7 | 5,59E-32 | 2,11E-13 | yes | yes |
| AT5G61420 | MYB28 | 1753 | 2117 | UP | 4,4 | 2,7 | 4,69E-22 | 6,78E-21 | ||
| AT5G63160 | BT1 | 16 | 1273 | UP | 9,1 | 4,7 | 2,98E-185 | 8,71E-136 | - | yes |
| AT5G67300 | ATMYB44 | 2070 | 381 | UP | 4,5 | 2,4 | 1,11E-21 | 1,87E-14 | yes | yes |
| AT5G67300 | ATMYB44 | 53 | 2438 | UP | 11,3 | 5,4 | 9,41E-140 | 1,06E-92 | ||
| AT1G70940 | PIN3 | 5237 | 20 | UP | 6,4 | 3,2 | 2,54E-03 | 2,51E+02 | yes | yes |
| AT2G01420 | PIN4 | 1622 | 367 | UP | 6,3 | 2,8 | 3,70E-35 | 3,35E-15 | yes | yes |
| AT2G01420 | PIN4 | 1630 | 973 | DOWN | 5,5 | 3,0 | 1,65E-27 | 7,06E-20 | ||
| AT2G26560 | PLA2A | 2249 | 1401 | UP | 6,1 | 3,1 | 1,59E-20 | 5,88E-12 | yes | yes |
| AT5G60450 | ARF4 | 294 | 2112 | UP | 6,7 | 4,0 | 6,92E-70 | 1,80E-64 | yes | yes |
| AT2G33860 | ARF3/ETT | 4041 | 1151 | UP | 3,9 | 3,1 | 6,30E-01 | 2,50E-07 | - | - |
| AT1G15690 | AVP1 | 3612 | 2238 | UP | 8,1 | 3,4 | 4,21E-13 | 8,87E-02 | yes | yes |
Notes: By analyzing the ChIP-seq and the tiling array datasets and based on gene ontology (GO) analysis and literature contrast, we identified 22 genes involved in response to auxin. These genes are listed with the AGI (Arabidopsis Genome Initiative) gene code, the Gene Symbol, the ORP-rank, the distance from the binding site to the CDS, the location (UP=upstream of a gene, DOWN=downstream of a gene, in CDS), the enrichment of ChIP-seq replicates 1 and 2 (ratio of number of reads for a binding site in KAN1+DEX versus Col0+DEX), the false discovery rate (FDR) of ChIP-seq replicates 1 and 2, and the down-regulation at 80 and/or 160 min after KANADI1 activity induction (yes=the entire length of the predicted transcript was down-regulated; - no significant down-regulation).
Finally, several genes previously described as being involved in adaxial/abaxial patterning of the leaf and the vascular tissues such as MIR166A and AS2 [1,17,18,24,40] were identified as KAN1 targets exclusively through the ChIP-seq approach. Moreover, KAN1 itself and KAN2 were isolated as putative targets suggesting that KAN1 may control its own expression as well as the expression of other KAN gene family members.
Genes oppositely regulated by the REV/KAN1 module
REV and KAN1 have opposite functions in early leaf patterning. In order to determine whether the antagonistic roles can be attributed to an opposite regulation of common downstream target genes, we compared potential downstream REV target genes identified by ChIP-Seq [29] with the list of genes bound and regulated by KAN1 (Dataset S7). This analysis resulted in the identification of 26 genes, which are candidates for dual regulation (Table 3). Interestingly, five genes are bound by REV and KAN1 in a region less than 100bp apart, suggesting that, besides dual regulation, REV and KAN1 might also compete for chromatin accessibility. All five genes (TEM, ZFP4, SUC1, a receptor protein kinase and a NPH3-like protein) seem to be involved in the control of development corroborating the idea that they act downstream of developmental regulators.
Table 3. Genes potentially cross-regulated by the REV/KAN1 module.
|
REV ChIP-seq
|
KAN1 ChIP-seq |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AGI | Gene symbol | Distance | Location | FDR replicate 1 | FDR replicate 2 | Enrichment replicate 1 | Enrichment replicate 2 | Distance | Location | Distance REV/KAN1 binding |
| AT1G22570 | Major facilitator protein | 1129 | DOWN | 1,19E-112 | 3,31E-68 | 13,5 | 6,2 | 920 | UP | 4085 |
| AT1G25560 | TEM1 | 2926 | UP | 2,15E-64 | 2,70E-31 | 11,7 | 4,9 | 3024 | UP | 61 |
| AT1G51940 | LysM-domain protein | 1525 | UP | 1,22E-10 | 3,24E+02 | 4,5 | 1,8 | 5999 | UP | 4504 |
| AT1G61660 | bHLH transcription factor | in CDS | 2,26E-37 | 1,47E-22 | 6,8 | 3,4 | 1365 | UP | 3405 | |
| AT1G66140 | ZFP4 | in CDS | 3,75E-31 | 4,10E-17 | 6,1 | 2,9 | in CDS | 26 | ||
| AT1G66140 | ZFP4 | 1,49E-01 | 1,35E+02 | 4,0 | 2,2 | in CDS | 434 | |||
| AT1G67710 | ARR11 | 1466 | UP | 2,20E-54 | 6,92E-40 | 6,3 | 3,4 | 1375 | UP | 113 |
| AT1G68130 | IDD14 | 3051 | UP | 1,08E-12 | 9,72E-11 | 3,8 | 2,2 | 3357 | DOWN | 8739 |
| AT1G68520 | B-BOX zinc finger protein | 741 | DOWN | 4,34E-26 | 3,09E-09 | 3,8 | 2,3 | 248 | UP | 2757 |
| AT1G68520 | B-BOX zinc finger protein | 5,97E-14 | 2,76E-06 | 5,9 | 2,8 | 829 | UP | 3408 | ||
| AT1G71880 | SUC1 | 5858 | UP | 2,80E-74 | 1,99E-51 | 8,1 | 4,1 | 5878 | UP | 41 |
| AT1G72300 | Leucine-rich receptor protein | 3145 | UP | 4,42E-22 | 4,03E-12 | 8,3 | 4,1 | 691 | UP | 2424 |
| AT1G72300 | Leucine-rich receptor protein | 6,72E-09 | 1,69E-09 | 4,0 | 2,4 | 1465 | UP | 1630 | ||
| AT3G02140 | TMAC2 | 893 | UP | 2,04E-69 | 5,94E-57 | 10,5 | 5,9 | 201 | UP | 653 |
| AT3G02140 | TMAC2 | 1,01E-48 | 5,34E-34 | 7,4 | 3,7 | 2737 | UP | 1863 | ||
| AT3G02140 | TMAC2 | 4,50E-08 | 5,09E-03 | 4,8 | 2,5 | 3049 | UP | 2231 | ||
| AT3G12920 | BRG3 | 1579 | DOWN | 1,73E-04 | 8,92E-01 | 4,3 | 2,3 | 1782 | UP | 5019 |
| AT3G12920 | BRG3 | 1,22E-181 | 7,71E-141 | 13,2 | 7,0 | 4565 | UP | 7652 | ||
| AT3G12920 | BRG3 | 6,75E-90 | 8,25E-62 | 11,6 | 5,8 | 983 | DOWN | 568 | ||
| AT3G12920 | BRG3 | 1,13E-63 | 9,51E-49 | 9,9 | 5,4 | 2338 | DOWN | 797 | ||
| AT3G15570 | NPH3 family protein | 1009 | UP | 6,12E-93 | 1,02E-63 | 8,9 | 4,4 | 1036 | UP | 14 |
| AT3G54400 | Aspartyl protease protein | 602 | UP | 4,51E+02 | 8,26E+04 | 4,6 | 2,4 | 120 | UP | 402 |
| AT3G56050 | Protein kinase family protein | 208 | UP | 4,29E-87 | 5,13E-48 | 8,8 | 3,9 | 274 | UP | 42 |
| AT3G61460 | BRH1 | 2196 | UP | 1,85E-17 | 4,61E-12 | 5,9 | 3,2 | 314 | UP | 1826 |
| AT4G18700 | CIPK12 | 282 | DOWN | 3,22E+00 | 3,57E+03 | 7,5 | 3,9 | 26 | UP | 2468 |
| AT4G18700 | CIPK12 | 4,36E-101 | 1,53E-92 | 8,4 | 5,0 | 133 | DOWN | 146 | ||
| AT4G22190 | unknown protein | 2827 | UP | 1,45E-38 | 9,85E-30 | 8,1 | 4,5 | 1709 | UP | 1166 |
| AT4G26540 | Leucine rich repeat receptor | 2234 | UP | 3,06E-86 | 1,40E-45 | 8,5 | 3,7 | 2160 | UP | 129 |
| AT4G26540 | Leucine rich repeat receptor | 8,31E-78 | 1,31E-53 | 7,1 | 3,6 | 896 | DOWN | 6893 | ||
| AT4G27260 | GH3.5, WES1 | 2494 | DOWN | 6,37E-13 | 1,14E-07 | 4,0 | 2,2 | 1074 | UP | 6295 |
| AT4G27260 | GH3.5, WES1 | 4,55E-09 | 1,63E+07 | 5,0 | 1,3 | 971 | DOWN | 1591 | ||
| AT5G05690 | CPD | 4847 | UP | 1,81E-06 | 1,67E-02 | 5,1 | 2,7 | 5642 | UP | 894 |
| AT5G47370 | HAT2 | 1548 | UP | 1,64E-12 | 1,76E-05 | 4,6 | 2,3 | 86 | UP | 1403 |
| AT5G51550 | EXL3 | 2573 | UP | 2,22E-174 | 1,08E-139 | 14,1 | 7,7 | 687 | UP | 1892 |
| AT5G51550 | EXL3 | 9,95E-19 | 1,35E-11 | 4,0 | 2,1 | 2133 | UP | 480 | ||
| AT5G52060 | ATBAG1, BAG1 | 739 | UP | 1,28E-20 | 2,48E-03 | 5,4 | 2,1 | 8 | UP | 745 |
| AT5G64570 | XYL4 | 2389 | UP | 1,84E-39 | 5,08E-19 | 4,9 | 2,2 | in CDS | 5597 | |
| AT5G64570 | XYL4 | 5,70E-50 | 5,28E-46 | 7,7 | 4,6 | 331 | UP | 2090 | ||
| AT5G67190 | DEAR2 | 2710 | UP | 1,46E-33 | 1,01E-22 | 4,9 | 2,6 | 276 | UP | 2557 |
| AT5G67190 | DEAR2 | 5,02E-178 | 3,45E-151 | 14,2 | 8,1 | 1541 | UP | 1248 | ||
| AT5G67190 | DEAR2 | 1,28E-61 | 2,35E-45 | 6,9 | 3,6 | 1319 | UP | 6884 | ||
Notes: By comparing the REV target genes identified by ChIP-Seq [29] with the list of genes bound and regulated by KAN1 (Dataset S7), we identified 26 genes which are candidates for dual regulation. These genes are listed with the AGI (Arabidopsis Genome Initiative) gene code, the Gene Symbol, the false discovery rate (FDR) of ChIP-seq replicates 1 and 2, the enrichment of ChIP-seq replicates 1 and 2 (ratio of number of reads for a binding site in KAN1+DEX versus Col0+DEX), the distance from the binding site to the CDS and the location (UP=upstream of a gene, DOWN=downstream of a gene, in CDS), and the distance between REV and KAN1 binding sites.
Discussion
In this study, we utilize inducible overexpression of KAN1 to identify KAN1 responsive genes and direct targets. Although such an approach may lead to artifacts because of the ectopic and artificially high expression levels used, the set of genes we have identified shows enrichment for genes involved in development and auxin biology, suggesting our experiments have identified genes that are biologically relevant.
Our results show that the VGAATAW motif may be a common cis-regulatory element recognized by KAN1, which includes the motif affected by the as2-5d point mutation that causes ectopic AS2 expression due to its regulation being uncoupled from KAN1 [31]. We have focused our attention on the 1802 binding regions containing this motif (corresponding to 3151 genes potentially regulated by KAN1) and, especially, on those genes that exhibit gene expression changes in response to induction of KAN1 activity. Several of the identified downstream targets have a role in organ development, shoot patterning or auxin response and transport. In addition, we present a set of genes that are potentially controlled by both KAN1 and REV. The potential regulation of the selected genes by KAN1 and its link with patterning processes and auxin-related events as well as the gene regulation by the module KAN1/REV are discussed below.
KAN1 regulates many genes related to organ patterning
In our study, we find that KAN1 binds to the promoter of two HD-ZIPIII genes, PHABULOSA and ATHB8 (Table 1) and represses their expression, suggesting that both HD-ZIPIII genes are direct targets of KAN1 during organ polarity establishment. In previous studies, it was proposed that the antagonistic role between KAN and HD-ZIPIII activities in vascular tissue formation is mediated by affecting the canalization of auxin flow rather than through a direct interaction between both families of transcription factors [10]. However, our results suggest that there may be contexts in which KAN1 acts directly on PHB and ATHB8.
We also find that KAN1 binds directly to the promoters of MIR166A and MIR166F and down-regulates the expression of MIR166F (Table 1), suggesting that at least KAN1 may directly regulate MIR166F. In addition, our results indicate that KAN1 binds to its own promoter and KAN2 via the VGAATAW motif (Table 1) but also potentially KAN3 (Dataset S1), although no VGAATAW motif was found for this binding event. Taken together, these results suggest that in some contexts KAN1 may direct a negative feedback loop that limits the levels of several abaxial factors including KAN1 itself.
KAN1 binds to the proximal promoters and represses the expression of genes involved in different aspects of organ development such as PXY/TDR, LNG1/2 and SAW2 (Table 1). Like KAN1, PXY is a key gene in vasculature polarity establishment. In particular, PXY is required for the proper orientation of cell divisions in the vascular meristem, which gives rise to specialized and spatially separated xylem and phloem cells [46,47]. The homologous genes LNG1 and LNG2 regulate leaf morphology by positively promoting longitudinal polar cell elongation [44]. The adaxial epidermal cells of the midveins and the leaf blade are longitudinally elongated in the lng1-1D mutant plants compared with wild type. SAW2 controls leaf shape and exhibits adaxial expression in developing lateral organs [45]. Therefore, our results suggest that KAN1 may directly regulate genes involved in the development of lateral organs and vascular tissue, known sites of KAN1 activity.
KAN1 regulates auxin-related genes
Organ patterning is in part modulated by the polar transport of auxin to specific locations, generating auxin maxima that promote organ initiation and growth. PIN proteins play an important role in the regulation of auxin distribution. Loss of proper PIN polarity establishment, as in PIN multiple mutants and gn mutants, leads to embryo patterning defects [61-65]. Previous studies have shown a negative effect of KAN1 on PIN1 activity. Thus, ectopic expression of PIN1 is observed in kan1 kan2 kan4 embryos, suggesting that KAN genes may act to restrict auxin flow during embryogenesis by regulating PIN1 gene expression [7]. PIN1 gene expression alterations have also been observed at the ectopic abaxial leaf outgrowths of kan1 kan2 plants. In particular, PIN1 expression was higher in the outgrowths than in the surrounding leaf tissue, suggesting that the outgrowths may be due to ectopic auxin maxima forming in the lamina [3,4]. In addition, it has been shown that polar auxin flow is essential to form procambium cells in vascular tissues, and KAN genes play a role in the distribution of this auxin flow by restricting PIN1 activity [10]. In agreement with these findings, we have identified a binding site for KAN1 downstream from PIN1 that likely mediates direct repression of PIN1 by KAN1 (Table 1). Motifs adjacent to other PIN genes such as PIN3 and PIN4 were also bound by KAN1, and their expression was repressed by KAN1 as well (Table 2). Therefore, KAN1 may directly regulate several PIN family members supporting previous findings that showed that, at least in some contexts, KAN proteins may act in patterning processes through auxin transport modulation. Additionally, and reinforcing this hypothesis, KAN1 bound and repressed several genes involved in the regulation of PIN activity and trafficking such as PINOID and PLA2A [57,58,66], respectively. NPY3, NPY5 and WAG2, which are thought to act together to determine what side of the cell PIN accumulates at [48,67-69], and genes involved in auxin polar transport such as the auxin influx transporter AUX1 and the ATP-binding cassette transporter AtMDR1 [59,60] were also bound and repressed by KAN1 (Table 2).
Our data also point to a direct effect of KAN1 on auxin signaling pathways. For instance KAN1 bound near and repressed early auxin response genes including three GH3 genes (DFL1, DFL2 and WES1), three SAUR-like genes (AT1G19840, AT1G75590 and AT2G21210) as well as two Aux/IAA genes (IAA2 and IAA13) [49,70] (Table 2).
A connection of auxin signaling comes in addition from direct repression of ARF4 and binding of ARF3 by KAN1 (Table 2). The phenotype of ett arf4 leaves resembles the phenotype of kan1 kan2 leaves, leading to the proposal that ARF proteins act together with KAN proteins or its downstream targets to regulate transcription [20]. While this previous study suggested a positive interaction between these transcription factors, our findings suggest that there may also be negative feedback between KAN1 and ARF4 and ARF3/ETT, potentially again (see above), as a mechanism to maintain homeostasis among factors controlling abaxial identity.
We have identified several additional genes involved in auxin transport and its regulation or auxin signaling as being repressed after KAN1 induction and, in some cases, also bound by KAN1 (Dataset S1 and Figure S1). This set of genes includes PIN7, which is involved in apical–basal axis formation of the embryo [62], YABBY5, a transcription factor involved in abaxial cell fate specification and auxin distribution [21,23], different early auxin-responsive genes such as GH3.3, the SMALL AUXIN UP RNAs (SAUR) SAUR19, SAUR20 and SAUR63, which regulate auxin polar transport and promote auxin-mediated organ elongation [71,72], three SAUR-like genes (AT1G19840, AT4G38840 and AT5G18030), ARF19, IAA3, IAA16, IAA14, and an auxin receptor belonging to the TIR1 subfamily (AFB1) that interacts with Aux/IAA proteins [73,74]. In addition, in a previous study [29], we demonstrated that the expression of HAT2, which was also bound and repressed by KAN1 in the current study, and two genes that encode auxin biosynthetic enzymes, TAA1 and YUC5, is reduced significantly after KAN1 induction. These results together with our findings reflect that, certainly, KAN1 may control the influence of auxin on organ development through complex interactions and at different levels: biosynthesis, transport and its regulation, and signaling.
Finally, our results suggest that KAN1 may act on other hormone pathways through the regulation of genes involved in the response to abscisic acid, jasmonic acid, brassinosteroids, ethylene, cytokinins and gibberellins (Dataset S2 and Figure S1).
Regulation by KAN1 and REV of common downstream target genes
Genetic analysis has indicated that the HD-ZIPIII and KAN factors act oppositely in organ patterning [1,3,7]. However, it remains unclear whether this interaction occurs by direct mutually antagonistic regulation, through opposing regulation of a set of common direct targets or through opposing regulation of indirect targets. With respect to direct antagonistic regulation, in the current study, we did not find evidence of direct regulation of REV by KAN1, although KAN1 appears to bind other HD-ZIPIII genes such as PHB and ATHB8 and to repress their expression. On the other hand, published work identifying HAT2, TAA1 and YUC5 as genes oppositely regulated by REV and KAN [29] supports the shared common targets hypothesis. To further investigate whether REV and KAN1 act on additional common target genes, we compared the ChIP-Seq data for KAN1 with those recently obtained for REV [29] and found an additional set of overlapping putative target genes that bring the total to 26 genes (Table 3). Among these, we found genes encoding transcription factors and proteins involved in hormone-associated processes. Finally, several genes involved in auxin transcriptional response and auxin transport are repressed by KAN1, whereas auxin biosynthesis and transport are positively regulated by HD-ZIPIII activity. Thus, another mechanism by which KAN1 and HD-ZIPIII activities have opposing effects is via antagonistic regulation of auxin biology, which does not necessarily occur at the level of the same transcriptional targets but will create steep auxin gradients that could function as positional signals.
The vast majority of KAN1 targets identified were down-regulated. Together with the observations that KAN1 directly represses the expression of the adaxial factor AS2 [31] and that TOPLESS, a co-repressor protein, directly interacts with KAN1 [38], our data suggests that KAN1 primarily acts as a repressor. According to the opposite regulation of common targets hypothesis, if KAN1 acts as a repressor, the HD-ZIPIII proteins should act as activators of those genes that are common targets. Consistent with HD-ZIPIII proteins acting as activators, expression of REV translationally fused with a repressor domain (REV-SRDX) phenocopies phb phv rev plants (Dyani Lewis and J. L. Bowman, unpublished data). Thus, our findings together with published work [1,3,7,29,31,38] indicate that HD-ZIPIII and KAN genes function antagonistically both through mutual regulation as well as through the opposite regulation of common direct targets and indirect targets. Mutual regulation may ensure the proper partitioning of adaxial and abaxial tissues while the opposite regulation of common targets may help set up contrasting transcriptional activities that distinguish adaxial and abaxial cell types.
Supporting Information
All ChIP-Seq identified regions containing the VGAATAW element.
(XLS)
Putative KAN1 targets with roles in system and organ development and hormone signaling.
(XLS)
All genes down-regulated by KAN1.
(XLS)
Genes involved in auxin biology down-regulated by KAN1.
(XLS)
Genes involved in transcriptional regulation down-regulated by KAN1.
(XLS)
All genes up-regulated by KAN1.
(XLS)
Overlap ChIP-Seq/tiling array.
(XLS)
Examples of raw tiling array data. The lower two lines in each figure represent the 80 minute time point and the upper two lines represent the 160 minute time point. The upper of the lines in each time point are from a single biological experiment, whereas the lower are the average from two biological replicates. Genes are identified by their AtNg and common names, and those genes that were detected as also bound by KAN1 are denoted by an *.
(TIF)
Genes bound by KAN1 are also regulated by KAN1 at the transcriptional level. A) ChIP-Seq graphs show enrichment for KAN1 binding in the 3’ region of the ATHB8 gene. The enriched region contains the VGAATAW motif. B) ATHB8 expression is strongly repressed in DEX-treated 35S::FLAG-GR-KAN1 transgenic plants. Plotted are relative qRT-PCR expression values of two independent biological replicates. Each biological experiment was carried out with four technical replicates and average values with standard deviation were calculated. *p≤0.01; **p≤1.0E-06.
(TIF)
Acknowledgments
Excellent technical support by Christa Lanz, Ronny Brandt, Gesine Seibold and Ingrid Blumberg is gratefully acknowledged.
Funding Statement
MH acknowledges the Australian Research Council (http://www.arc.gov.au/) and European Research Council (GA Nº261081)(http://erc.europa.eu/) for current funding. The SW laboratory is funded by grants of the Deutsche Forschungsgemeinschaft (WE4281/6-1, WE4281/7-1) (http://www.dfg.de/) and the European Union (FP7 No. 256502) (http://cordis.europa.eu/fp7/home_en.html). LB was supported by the Swiss national Science Foundation (PBZHA-1188060)(http://www.snf.ch/E/Pages/default.aspx) and JLB acknowledges support from the Australian Research Council (DP0771232, DP110100070)(http://www.arc.gov.au/). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Emery JF, Floyd SK, Alvarez J, Eshed Y, Hawker NP et al. (2003) Radial Patterning of Arabidopsis Shoots by Class III HD-ZIP and KANADI Genes. Curr Biol 13: 1768-1774. doi: 10.1016/j.cub.2003.09.035. PubMed: 14561401. [DOI] [PubMed] [Google Scholar]
- 2. Eshed Y, Baum SF, Bowman JL (1999) Distinct mechanisms promote polarity establishment in carpels of Arabidopsis. Cell 99: 199-209. doi: 10.1016/S0092-8674(00)81651-7. PubMed: 10535738. [DOI] [PubMed] [Google Scholar]
- 3. Eshed Y, Baum SF, Perea JV, Bowman JL (2001) Establishment of polarity in lateral organs of plants. Curr Biol 11: 1251-1260. doi: 10.1016/S0960-9822(01)00392-X. PubMed: 11525739. [DOI] [PubMed] [Google Scholar]
- 4. Eshed Y, Izhaki A, Baum SF, Floyd SK, Bowman JL (2004) Asymmetric leaf development and blade expansion in Arabidopsis are mediated by KANADI and YABBY activities. Development 131: 2997-3006. doi: 10.1242/dev.01186. PubMed: 15169760. [DOI] [PubMed] [Google Scholar]
- 5. Bowman JL, Floyd SK (2008) Patterning and Polarity in Seed Plant Shoots. Annu Rev Plant Biol 59: 67-88. doi: 10.1146/annurev.arplant.57.032905.105356. PubMed: 18031217. [DOI] [PubMed] [Google Scholar]
- 6. Hawker NP, Bowman JL (2004) Roles for Class III HD-Zip and KANADI genes in Arabidopsis root development. Plant Physiol 135: 2261-2270. doi: 10.1104/pp.104.040196. PubMed: 15286295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Izhaki A, Bowman JL (2007) KANADI and class III HD-Zip gene families regulate embryo patterning and modulate auxin flow during embryogenesis in Arabidopsis. Plant Cell 19: 495-508. doi: 10.1105/tpc.106.047472. PubMed: 17307928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Smith ZR, Long JA (2010) Control of Arabidopsis apical-basal embryo polarity by antagonistic transcription factors. Nature 464: 423-426. doi: 10.1038/nature08843. PubMed: 20190735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Heisler MG, Ohno C, Das P, Sieber P, Reddy GV et al. (2005) Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr Biol 15: 1899-1911. doi: 10.1016/j.cub.2005.09.052. PubMed: 16271866. [DOI] [PubMed] [Google Scholar]
- 10. Ilegems M, Douet V, Meylan-Bettex M, Uyttewaal M, Brand L et al. (2010) Interplay of auxin, KANADI and Class III HD-ZIP transcription factors in vascular tissue formation. Development 137: 975-984. doi: 10.1242/dev.047662. PubMed: 20179097. [DOI] [PubMed] [Google Scholar]
- 11. Juarez MT, Kui JS, Thomas J, Heller BA, Timmermans MCP (2004) microRNA-mediated repression of rolled leaf1 specifies maize leaf polarity. Nature 428: 84-88. doi: 10.1038/nature02363. PubMed: 14999285. [DOI] [PubMed] [Google Scholar]
- 12. Kerstetter RA, Bollman K, Taylor RA, Bomblies K, Poethig RS (2001) KANADI regulates organ polarity in Arabidopsis. Nature 411: 706-709. doi: 10.1038/35079629. PubMed: 11395775. [DOI] [PubMed] [Google Scholar]
- 13. McConnell JR, Barton MK (1998) Leaf polarity and meristem formation in Arabidopsis. Development 125: 2935-2942. PubMed: 9655815. [DOI] [PubMed] [Google Scholar]
- 14. McConnell JR, Emery J, Eshed Y, Bao N, Bowman J et al. (2001) Role of PHABULOSA and PHAVOLUTA in determining radial patterning in shoots. Nature 411: 709-713. doi: 10.1038/35079635. PubMed: 11395776. [DOI] [PubMed] [Google Scholar]
- 15. Otsuga D, DeGuzman B, Prigge MJ, Drews GN, Clark SE (2001) REVOLUTA regulates meristem initiation at lateral positions. Plant J 25: 223-236. doi: 10.1046/j.1365-313x.2001.00959.x. PubMed: 11169198. [DOI] [PubMed] [Google Scholar]
- 16. Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M et al. (2000) Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature 408: 967-971. doi: 10.1038/35050091. PubMed: 11140682. [DOI] [PubMed] [Google Scholar]
- 17. Iwakawa H, Ueno Y, Semiarti E, Onouchi H, Kojima S et al. (2002) The ASYMMETRIC LEAVES2 gene of Arabidopsis thaliana, required for formation of a symmetric flat leaf lamina, encodes a member of a novel family of proteins characterized by cysteine repeats and a leucine zipper. Plant Cell Physiol 43: 467-478. doi: 10.1093/pcp/pcf077. PubMed: 12040093. [DOI] [PubMed] [Google Scholar]
- 18. Iwakawa H, Iwasaki M, Kojima S, Ueno Y, Soma T et al. (2007) Expression of the ASYMMETRIC LEAVES2 gene in the adaxial domain of Arabidopsis leaves represses cell proliferation in this domain and is critical for the development of properly expanded leaves. Plant J 51: 173-184. doi: 10.1111/j.1365-313X.2007.03132.x. PubMed: 17559509. [DOI] [PubMed] [Google Scholar]
- 19. Chitwood DH, Nogueira FTS, Howell MD, Montgomery TA, Carrington JC et al. (2009) Pattern formation via small RNA mobility. Genes Dev 23: 549-554. doi: 10.1101/gad.1770009. PubMed: 19270155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Pekker I, Alvarez JP, Eshed Y (2005) Auxin response factors mediate Arabidopsis organ asymmetry via modulation of KANADI activity. Plant Cell 17: 2899-2910. doi: 10.1105/tpc.105.034876. PubMed: 16199616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Sarojam R, Sappl PG, Goldshmidt A, Efroni I, Floyd SK et al. (2010) Differentiating Arabidopsis shoots from leaves by combined YABBY activities. Plant Cell 22: 2113-2130. doi: 10.1105/tpc.110.075853. PubMed: 20628155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Sawa S, Watanabe K, Goto K, Liu YG, Shibata D et al. (1999) FILAMENTOUS FLOWER, a meristem and organ identity gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domains. Genes Dev 13: 1079-1088. doi: 10.1101/gad.13.9.1079. PubMed: 10323860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Siegfried KR, Eshed Y, Baum SF, Otsuga D, Drews GN et al. (1999) Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 126: 4117-4128. PubMed: 10457020. [DOI] [PubMed] [Google Scholar]
- 24. Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP (2002) MicroRNAs in plants. Genes Dev 16: 1616-1626. doi: 10.1101/gad.1004402. PubMed: 12101121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Mallory AC, Reinhart BJ, Jones-Rhoades MW, Tang G, Zamore PD et al. (2004) MicroRNA control of PHABULOSA in leaf development: importance of pairing to the microRNA. p. 5 [prime] region. EMBO J 23: 3356-3364 [DOI] [PMC free article] [PubMed]
- 26. Kim J, Jung JH, Reyes JL, Kim YS, Kim SY et al. (2005) microRNA-directed cleavage of ATHB15 mRNA regulates vascular development in Arabidopsis inflorescence stems. Plant J 42: 84-94. doi: 10.1111/j.1365-313X.2005.02354.x. PubMed: 15773855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Williams L, Carles CC, Osmont KS, Fletcher JC (2005) A database analysis method identifies an endogenous trans-acting short-interfering RNA that targets the Arabidopsis ARF2, ARF3, and ARF4 genes. Proc Natl Acad Sci U S A 102: 9703-9708. doi: 10.1073/pnas.0504029102. PubMed: 15980147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Allen E, Xie Z, Gustafson AM, Carrington JC (2005) microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207-221. doi: 10.1016/j.cell.2005.04.004. PubMed: 15851028. [DOI] [PubMed] [Google Scholar]
- 29. Brandt R, Salla-Martret M, Bou-Torrent J, Musielak T, Stahl M et al. (2012) Genome-wide binding-site analysis of REVOLUTA reveals a link between leaf patterning and light-mediated growth responses. Plant J 72: 31-42. doi: 10.1111/j.1365-313X.2012.05049.x. PubMed: 22578006. [DOI] [PubMed] [Google Scholar]
- 30. Liu T, Reinhart BJ, Magnani E, Huang T, Kerstetter R et al. (2013) Of Blades and Branches: Understanding and Expanding the Arabidopsis Ad/Abaxial Regulatory Network through Target Gene Identification. Cold Spring Harb Symp Quant Biol [DOI] [PubMed] [Google Scholar]
- 31. Wu G, Lin WC, Huang T, Poethig RS, Springer PS et al. (2008) KANADI1 regulates adaxial-abaxial polarity in Arabidopsis by directly repressing the transcription of ASYMMETRIC LEAVES2. Proc Natl Acad Sci U S A 105: 16392-16397. doi: 10.1073/pnas.0803997105. PubMed: 18849474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Wenkel S, Emery J, Hou BH, Evans MM, Barton MK (2007) A feedback regulatory module formed by LITTLE ZIPPER and HD-ZIPIII genes. Plant Cell 19: 3379-3390. doi: 10.1105/tpc.107.055772. PubMed: 18055602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Kim Y-S, Kim S-G, Lee M, Lee I, Park H-Y et al. (2008) HD-ZIP III Activity Is Modulated by Competitive Inhibitors via a Feedback Loop in Arabidopsis Shoot Apical Meristem Development. Plant Cell 20: 920-933. doi: 10.1105/tpc.107.057448. PubMed: 18408069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Bou-Torrent J, Salla-Martret M, Brandt R, Musielak T, Palauqui J-C et al. (2012) ATHB4 and HAT3, two class II HD-ZIP transcription factors, control leaf development in Arabidopsis. Plant Signal Behav 7: 1382-1387. doi: 10.4161/psb.21824. PubMed: 22918502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Weigel M, Varotto C, Pesaresi P, Finazzi G, Rappaport F et al. (2003) Plastocyanin Is Indispensable for Photosynthetic Electron Flow in Arabidopsis thaliana. J Biol Chem 278: 31286-31289. doi: 10.1074/jbc.M302876200. PubMed: 12773541. [DOI] [PubMed] [Google Scholar]
- 36. Yant L, Mathieu J, Dinh TT, Ott F, Lanz C et al. (2010) Orchestration of the Floral Transition and Floral Development in Arabidopsis by the Bifunctional Transcription Factor APETALA2. Plant Cell Available: 22: 2156-2170 PubMed: 20675573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Nicol JW, Helt GA, Blanchard SG, Raja A, Loraine AE (2009) The Integrated Genome Browser: free software for distribution and exploration of genome-scale datasets. Bioinformatics 25: 2730-2731. doi: 10.1093/bioinformatics/btp472. PubMed: 19654113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Causier B, Ashworth M, Guo W, Davies B (2012) The TOPLESS Interactome: A Framework for Gene Repression in Arabidopsis. Plant Physiol 158: 423-438. doi: 10.1104/pp.111.186999. PubMed: 22065421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Baima S, Nobili F, Sessa G, Lucchetti S, Ruberti I et al. (1995) The expression of the Athb-8 homeobox gene is restricted to provascular cells in Arabidopsis thaliana. Development 121: 4171-4182. PubMed: 8575317. [DOI] [PubMed] [Google Scholar]
- 40. Rhoades MW, Reinhart BJ, Lim LP, Burge CB, Bartel B et al. (2002) Prediction of plant microRNA targets. Cell 110: 513-520. doi: 10.1016/S0092-8674(02)00863-2. PubMed: 12202040. [DOI] [PubMed] [Google Scholar]
- 41. Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D et al. (2003) Local, Efflux-Dependent Auxin Gradients as a Common Module for Plant Organ Formation. Cell 115: 591-602. doi: 10.1016/S0092-8674(03)00924-3. PubMed: 14651850. [DOI] [PubMed] [Google Scholar]
- 42. Reinhardt D, Pesce E-R, Stieger P, Mandel T, Baltensperger K et al. (2003) Regulation of phyllotaxis by polar auxin transport. Nature 426: 255-260. doi: 10.1038/nature02081. PubMed: 14628043. [DOI] [PubMed] [Google Scholar]
- 43. Vernoux T, Kronenberger J, Grandjean O, Laufs P, Traas J (2000) PIN-FORMED 1 regulates cell fate at the periphery of the shoot apical meristem. Development 127: 5157-5165. PubMed: 11060241. [DOI] [PubMed] [Google Scholar]
- 44. Lee YK, Kim GT, Kim IJ, Park J, Kwak SS et al. (2006) LONGIFOLIA1 and LONGIFOLIA2, two homologous genes, regulate longitudinal cell elongation in Arabidopsis. Development 133: 4305-4314. doi: 10.1242/dev.02604. PubMed: 17038516. [DOI] [PubMed] [Google Scholar]
- 45. Kumar R, Kushalappa K, Godt D, Pidkowich MS, Pastorelli S et al. (2007) The Arabidopsis BEL1-LIKE HOMEODOMAIN proteins SAW1 and SAW2 act redundantly to regulate KNOX expression spatially in leaf margins. Plant Cell 19: 2719-2735. doi: 10.1105/tpc.106.048769. PubMed: 17873098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Hirakawa Y, Shinohara H, Kondo Y, Inoue A, Nakanomyo I et al. (2008) Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proc Natl Acad Sci USA 105: 15208-15213. doi: 10.1073/pnas.0808444105. PubMed: 18812507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Fisher K, Turner S (2007) PXY, a receptor-like kinase essential for maintaining polarity during plant vascular-tissue development. Curr Biol 17: 1061-1066. doi: 10.1016/j.cub.2007.05.049. PubMed: 17570668. [DOI] [PubMed] [Google Scholar]
- 48. Cheng Y, Qin G, Dai X, Zhao Y (2008) NPY genes and AGC kinases define two key steps in auxin-mediated organogenesis in Arabidopsis. Proc Natl Acad Sci U S A 105: 21017-21022. doi: 10.1073/pnas.0809761106. PubMed: 19075219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Tiwari SB, Hagen G, Guilfoyle TJ (2004) Aux/IAA Proteins Contain a Potent Transcriptional Repression Domain. Plant Cell Available: 16: 533-543 PubMed: 14742873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Staswick PE, Tiryaki I, Rowe ML (2002) Jasmonate Response Locus JAR1 and Several Related Arabidopsis Genes Encode Enzymes of the Firefly Luciferase Superfamily That Show Activity on Jasmonic, Salicylic, and Indole-3-Acetic Acids in an Assay for Adenylation. Plant Cell Available: 14: 1405-1415 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Yang T, Poovaiah BW (2000) Molecular and Biochemical Evidence for the Involvement of Calcium/Calmodulin in Auxin Action. J Biol Chem 275: 3137-3143. doi: 10.1074/jbc.275.5.3137. PubMed: 10652297. [DOI] [PubMed] [Google Scholar]
- 52. Knauss S, Rohrmeier T, Lehle L (2003) The Auxin-induced Maize Gene ZmSAUR2 Encodes a Short-lived Nuclear Protein Expressed in Elongating Tissues. J Biol Chem 278: 23936-23943. doi: 10.1074/jbc.M212585200. PubMed: 12695517. [DOI] [PubMed] [Google Scholar]
- 53. Iwasaki M, Takahashi H, Iwakawa H, Nakagawa A, Ishikawa T et al. (2013) Dual regulation of ETTIN (ARF3) gene expression by AS1-AS2, which maintains the DNA methylation level, is involved in stabilization of leaf adaxial-abaxial partitioning in Arabidopsis. Development 140: 1958-1969. doi: 10.1242/dev.085365. PubMed: 23571218. [DOI] [PubMed] [Google Scholar]
- 54. Sawa S, Ohgishi M, Goda H, Higuchi K, Shimada Y et al. (2002) The HAT2 gene, a member of the HD-Zip gene family, isolated as an auxin inducible gene by DNA microarray screening, affects auxin response in Arabidopsis. Plant J 32: 1011-1022. doi: 10.1046/j.1365-313X.2002.01488.x. PubMed: 12492842. [DOI] [PubMed] [Google Scholar]
- 55. Friml J, Wiśniewska J, Benková E, Mendgen K, Palme K (2002) Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 415: 806-809. doi: 10.1038/415806a. PubMed: 11845211. [DOI] [PubMed] [Google Scholar]
- 56. Friml J, Benková E, Blilou I, Wisniewska J, Hamann T et al. (2002) AtPIN4 Mediates Sink-Driven Auxin Gradients and Root Patterning in Arabidopsis. Cell 108: 661-673. doi: 10.1016/S0092-8674(02)00656-6. PubMed: 11893337. [DOI] [PubMed] [Google Scholar]
- 57. Lee OR, Kim SJ, Kim HJ, Hong JK, Ryu SB et al. (2010) Phospholipase A(2) is required for PIN-FORMED protein trafficking to the plasma membrane in the Arabidopsis root. Plant Cell 22: 1812-1825. doi: 10.1105/tpc.110.074211. PubMed: 20525850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Benjamins R, Quint A, Weijers D, Hooykaas P, Offringa R (2001) The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 128: 4057-4067. PubMed: 11641228. [DOI] [PubMed] [Google Scholar]
- 59. Carrier DJ, Bakar NT, Swarup R, Callaghan R, Napier RM et al. (2008) The binding of auxin to the Arabidopsis auxin influx transporter AUX1. Plant Physiol 148: 529-535. doi: 10.1104/pp.108.122044. PubMed: 18614710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Lin R, Wang H (2005) Two homologous ATP-binding cassette transporter proteins, AtMDR1 and AtPGP1, regulate Arabidopsis photomorphogenesis and root development by mediating polar auxin transport. Plant Physiol 138: 949-964. doi: 10.1104/pp.105.061572. PubMed: 15908594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Steinmann T, Geldner N, Grebe M, Mangold S, Jackson CL et al. (1999) Coordinated Polar Localization of Auxin Efflux Carrier PIN1 by GNOM ARF GEF. Science 286: 316-318. doi: 10.1126/science.286.5438.316. PubMed: 10514379. [DOI] [PubMed] [Google Scholar]
- 62. Friml J, Vieten A, Sauer M, Weijers D, Schwarz H et al. (2003) Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 426: 147-153. doi: 10.1038/nature02085. PubMed: 14614497. [DOI] [PubMed] [Google Scholar]
- 63. Geldner N, Anders N, Wolters H, Keicher J, Kornberger W et al. (2003) The Arabidopsis GNOM ARF-GEF Mediates Endosomal Recycling, Auxin Transport, and Auxin-Dependent Plant Growth. Cell 112: 219-230. doi: 10.1016/S0092-8674(03)00003-5. PubMed: 12553910. [DOI] [PubMed] [Google Scholar]
- 64. Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I et al. (2005) The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433: 39-44. doi: 10.1038/nature03184. PubMed: 15635403. [DOI] [PubMed] [Google Scholar]
- 65. Vieten A, Vanneste S, Wisniewska J, Benková E, Benjamins R et al. (2005) Functional redundancy of PIN proteins is accompanied by auxin-dependent cross-regulation of PIN expression. Development 132: 4521-4531. doi: 10.1242/dev.02027. PubMed: 16192309. [DOI] [PubMed] [Google Scholar]
- 66. Friml J, Yang X, Michniewicz M, Weijers D, Quint A et al. (2004) A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux. Science 306: 862-865. doi: 10.1126/science.1100618. PubMed: 15514156. [DOI] [PubMed] [Google Scholar]
- 67. Li Y, Dai X, Cheng Y, Zhao Y (2011) NPY Genes Play an Essential Role in Root Gravitropic Responses in Arabidopsis. Molecular Plants 4: 171-179. doi: 10.1093/mp/ssq052. PubMed: 20833732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Furutani M, Kajiwara T, Kato T, Treml BS, Stockum C et al. (2007) The gene MACCHI-BOU 4/ENHANCER OF PINOID encodes a NPH3-like protein and reveals similarities between organogenesis and phototropism at the molecular level. Development 134: 3849-3859. doi: 10.1242/dev.009654. PubMed: 17913786. [DOI] [PubMed] [Google Scholar]
- 69. Spalding EP (2013) Diverting the downhill flow of auxin to steer growth during tropisms. Am J Bot 100: 203-214. doi: 10.3732/ajb.1200420. PubMed: 23284058. [DOI] [PubMed] [Google Scholar]
- 70. Tiwari SB, Hagen G, Guilfoyle T (2003) The Roles of Auxin Response Factor Domains in Auxin-Responsive Transcription. Plant Cell Available: 15: 533-543 PubMed: 12566590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Chae K, Isaacs CG, Reeves PH, Maloney GS, Muday GK et al. (2012) Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant J 71: 684-697. doi: 10.1111/j.1365-313X.2012.05024.x. PubMed: 22507274. [DOI] [PubMed] [Google Scholar]
- 72. Spartz AK, Lee SH, Wenger JP, Gonzalez N, Itoh H et al. (2012) The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J 70: 978-990. doi: 10.1111/j.1365-313X.2012.04946.x. PubMed: 22348445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Cecchetti V, Altamura MM, Falasca G, Costantino P, Cardarelli M (2008) Auxin Regulates Arabidopsis Anther Dehiscence, Pollen Maturation, and Filament Elongation. Plant Cell Available: 20: 1760-1774 PubMed: 18628351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Parry G, Calderon-Villalobos LI, Prigge M, Peret B, Dharmasiri S et al. (2009) Complex regulation of the TIR1/AFB family of auxin receptors. Proc Natl Acad Sci USA 106: 22540-22545. doi: 10.1073/pnas.0911967106. PubMed: 20018756. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
All ChIP-Seq identified regions containing the VGAATAW element.
(XLS)
Putative KAN1 targets with roles in system and organ development and hormone signaling.
(XLS)
All genes down-regulated by KAN1.
(XLS)
Genes involved in auxin biology down-regulated by KAN1.
(XLS)
Genes involved in transcriptional regulation down-regulated by KAN1.
(XLS)
All genes up-regulated by KAN1.
(XLS)
Overlap ChIP-Seq/tiling array.
(XLS)
Examples of raw tiling array data. The lower two lines in each figure represent the 80 minute time point and the upper two lines represent the 160 minute time point. The upper of the lines in each time point are from a single biological experiment, whereas the lower are the average from two biological replicates. Genes are identified by their AtNg and common names, and those genes that were detected as also bound by KAN1 are denoted by an *.
(TIF)
Genes bound by KAN1 are also regulated by KAN1 at the transcriptional level. A) ChIP-Seq graphs show enrichment for KAN1 binding in the 3’ region of the ATHB8 gene. The enriched region contains the VGAATAW motif. B) ATHB8 expression is strongly repressed in DEX-treated 35S::FLAG-GR-KAN1 transgenic plants. Plotted are relative qRT-PCR expression values of two independent biological replicates. Each biological experiment was carried out with four technical replicates and average values with standard deviation were calculated. *p≤0.01; **p≤1.0E-06.
(TIF)