

Abstract
Background
Human magneto/electrophysiology studies suggest that the phantom sound of tinnitus arises from spontaneous oscillatory neural activity in auditory cortex; however, in animal models, behavioral techniques suitable for testing this hypothesis in combination with electrophysiology recordings have yet to be evaluated. While electrophysiological studies of tinnitus have been reported in passive, awake animals, these studies fail to control for attentional mechanisms likely to play a role in the perception of tinnitus.
New Method
A novel appetitive operant conditioning, two-alternative identification task was developed for detecting acute tinnitus in rats. The procedure optimizes conditions for simultaneously recording oscillatory neural activity while controlling for the attentional state of the animal.
Results
Tinnitus was detected in six of seven rats following systemic injection with sodium salicylate (200 mg/kg IP), a known inducer of tinnitus. Analysis of ongoing local field potentials recorded from chronically implanted electrodes in auditory cortex of a rat reporting tinnitus revealed changes in the spectrum of ongoing neural activity.
Comparison with Existing Method(s)
Existing tinnitus-detection methods were not explicitly designed for the simultaneous recording of neural activity. The behavioral method reported here is the first to provide the conditions necessary for obtaining these recordings in chronically implanted rats.
Conclusions
The behavioral assay presented here will facilitate research into the neural mechanisms of tinnitus by allowing researchers to compare the electrophysiological data in animals with confirmed tinnitus.
Keywords: tinnitus, animal model, electrophysiology, auditory cortex, oscillatory activity, sodium salicylate
1. Introduction
Subjective tinnitus is the perception of a ringing or hissing sound with no acoustic source. In the United States, an estimated 50 million adults have experienced hearing tinnitus occasionally, and 16 million adults are estimated to experience frequent tinnitus perception (Shargorodsky et al., 2010). Despite decades of research, the neural circuits underlying tinnitus generation remain elusive. Recent human brain imaging studies have generated testable hypotheses regarding the relationships between abnormal patterns of neural activity in specific brain regions and tinnitus perception (see below). In order for these hypotheses to be more rigorously tested, behavioral models of tinnitus should be designed to allow for the simultaneous recording of neural activity during an auditory task.
Several magneto- and electroencephalographic studies in humans have reported abnormal patterns of ongoing neural activity in auditory and non-auditory brain regions of tinnitus subjects. Macroscopic fluctuations in brain activity are measured outside the cranium with magneto- or electroencephalography and reflect subthreshold currents generated within the brain which can be measured as voltage fluctuations called local field potentials (LFPs; Buzsaki et al., 2012). Oscillations of the LFP have been shown to modulate the probability and precise timing of neuron action potentials and have been demonstrated to play critical roles in various aspects of perceptual or cognitive function (for reviews on the topic, Basar et al., 2000; Buzsaki et al., 2012; Fries, 2009; Singer, 1999). Studies in human subjects suggest that specific changes in oscillatory activity are involved in tinnitus perception or an emotional response associated with the tinnitus percept (Adjamian et al., 2012; Ashton et al., 2007; Lorenz et al., 2009; Moazami-Goudarzi et al., 2010; Sedley et al., 2012; van der Loo et al., 2009; Weisz et al., 2005; Weisz et al., 2007b; Wienbruch et al., 2006). In particular, tinnitus subjects exhibit a reduction in alpha band activity (~ 10 Hz; Lorenz et al., 2009; Weisz et al., 2005) with an increase in delta band (1.5 to 4 Hz; Weisz et al., 2007a; Weisz et al., 2005) and gamma band (> 30 Hz; Ashton et al., 2007; Llinas et al., 1999; Lorenz et al., 2009; Weisz et al., 2007a) activity compared to control subjects.
While human studies on the neural mechanisms of tinnitus have provided important insights, animal models allow for a more precise and controlled manner of investigation, specifically the induction of tinnitus and its potential reversal. However, a major limitation of most animal studies on tinnitus is that the electrophysiological experiments were performed under anesthesia which significantly alters both spontaneous and sound-evoked neural activity in the auditory system (Kisley and Gerstein, 1999). Since anesthesia abolishes perception, electrophysiological recordings in conscious animals with behaviorally-confirmed tinnitus are needed to provide insight into the precise aberrant mechanisms responsible for generating the tinnitus percept. To date, only a handful of electrophysiological studies following manipulations intended to induce tinnitus perception have been reported in conscious, but passive animals (Norena et al., 2010; Yang et al., 2007; Zhang et al., 2011). Electrophysiological studies with animals actively reporting the presence of tinnitus, which would more appropriately model the human studies, have yet to be reported. Thus we endeavored to control for the effect of attention on neural activity by performing electrophysiological recordings when an animal was attending to its acoustic environment and actively reporting the presence of tinnitus via behavioral testing. To our knowledge, this study represents the first behavioral paradigm in animals that has been optimized for the simultaneous recording of neural activity during behaviorally-confirmed tinnitus perception.
Since Jastreboff and colleagues first established that tinnitus could be detected in rats (Jastreboff et al., 1988), a variety of behavioral paradigms have been developed for rats and other laboratory animals. Existing behavioral paradigms for tinnitus screening fall into roughly three main categories (for a review on tinnitus behavior paradigms see Turner, 2007): shock avoidance (Bauer et al., 1999; Guitton et al., 2003; Heffner, 2011; Heffner and Harrington, 2002; Jastreboff et al., 1988; Lobarinas et al., 2004), appetitive operant conditioning (Rüttiger et al., 2003), and gap pre-pulse inhibition of the acoustic startle reflex (Berger et al., 2013; Dehmel et al., 2012; Longenecker and Galazyuk, 2012; Turner et al., 2006; Turner and Parrish, 2008; Tzounopoulos, 2008; Yang et al., 2007). It is important to note that most of these behavioral paradigms include features that would make them incompatible for simultaneously measuring ongoing oscillatory neural activity while screening for tinnitus. Specifically, in order to properly assess oscillatory components of the LFP, neural activity needs to be sampled continuously over several seconds and therefore the behavioral paradigm requires relatively long and stable epochs. This requirement excludes most existing behavioral paradigms which detect tinnitus using discrete stimuli such as brief silent gaps in noise or tone-burst detection. Furthermore, behavioral paradigms that rely on sound-evoked responses would likely interfere with tinnitus-related neural activity that occurs spontaneously in the absence of sound. To overcome these limitations, we developed a novel two-alternative choice identification task for the detection of tinnitus in rats. This method optimized conditions for acquiring electrophysiological data during several seconds while performing a behavioral task in which the rat reports hearing a steady sound during quiet intervals; i.e. tinnitus.
In order to evaluate the efficacy of our paradigm, temporary tinnitus was induced in trained rats by systemic injection of sodium salicylate which reliably induces temporary tinnitus in normal hearing humans (Mongan et al., 1973) and rats (Jastreboff et al., 1988; Lobarinas et al., 2004). To illustrate its utility, we include a subset of electrophysiological recordings from a chronically implanted electrode in auditory cortex (AC) of a rat concurrently reporting tinnitus.
2. Materials and Methods
2.1. Objective
To obtain estimates of ongoing oscillatory neural activity from the AC during tinnitus, we developed a novel two-alternative choice identification task for the detection of tinnitus in rats. The use of rats in this project was approved by the Institutional Animal Care and Use Committee at the University of Buffalo and was carried out in accordance with National Institutes of Health guidelines.
2.2. Behavior Apparatus
The behavioral apparatus was constructed of transparent acrylic walls and equipped with 3 infrared (IR) detectors; one located at the center nose poke and one at the right feeder trough and one at the left feeder trough (Vulintus, Dallas, TX, USA or MedAssociates Inc., St. Albans, VT, USA). A bright light (green or white light-emitting diode) was located directly above the center nose poke and served as the GO cue (Figure 1). The behavior apparatus was illuminated by an ambient house light and housed in a double-walled, electrically shielded sound-attenuating cubicle constructed of 2 layers of dense fiber board.
Figure 1.
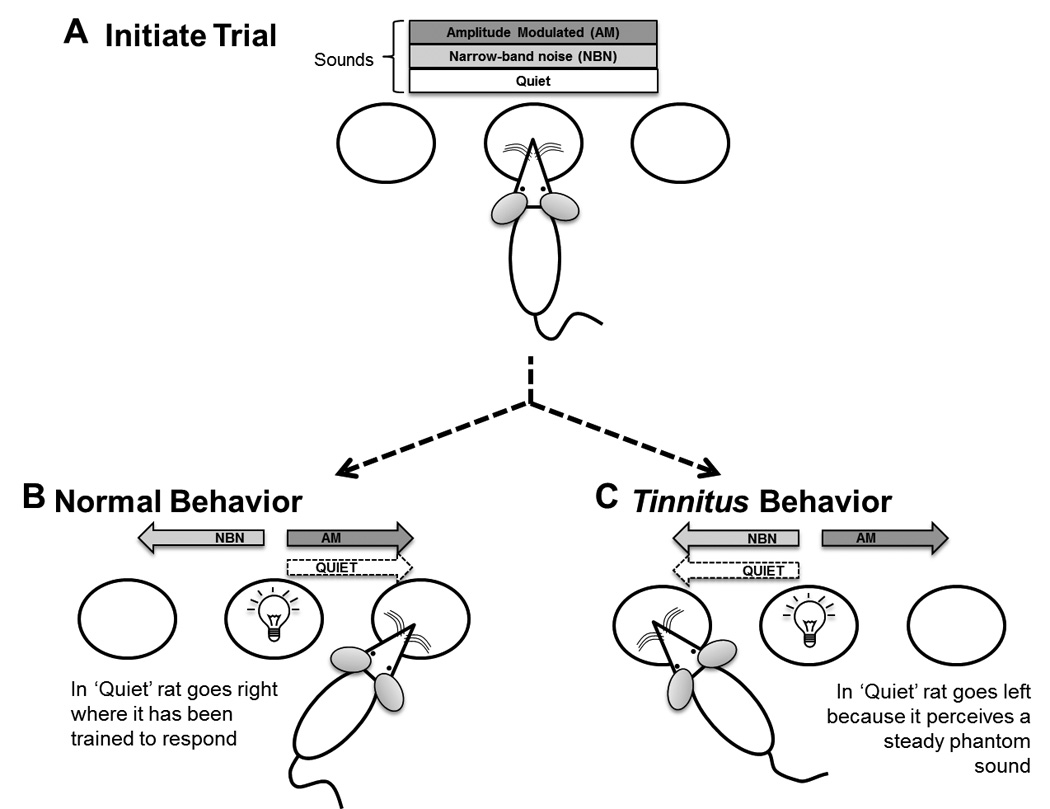
Overview of the behavioral paradigm. A rat initiates a trial by poking and holding its nose in the center port (A). One of three acoustic conditions, AM, NBN, or Quiet, are present. A light emitting diode illuminates (serving as the GO cue) after a random interval (4 – 8 s) directing the rat to respond at the left feeder trough for NBN trials or the right feeder for AM or Quiet trials (B). After salicylate, if an animal misidentifies the Quiet trials by responding to the left feeder trough (C) it is presumed to be experiencing a steady NBN-like phantom sound. See Methods for details.
The behavior apparatus was computer-controlled with custom software written in Matlab (MathWorks, Nattick, MA, USA) interfaced with real-time processing hardware (RX6-5, Tucker-Davis Technologies, Alachua, FL, USA). The speaker (FT28D, Fostex, Tokyo, Japan) was located on the roof of the cage. The intensity of the acoustic stimuli used was calibrated with a microphone (1/4” mic model 2520, Larson-Davis, Depew, NY, USA) at the approximate location of where the rat’s head would be when it poked its nose into the center nose poke. Sound levels were ~60 dB SPL for narrow band noise (NBN) and sinusoidally amplitude modulated (AM) broad-band noise stimuli.
2.3. Training
Male Sprague-Dawley albino rats (n = 7; tested between 6 and 7 months of age) were food restricted to 85 – 90% of free feeding weight in order to promote exploratory behavior when placed in the behavioral boxes. Initially, incidental spontaneous nose pokes into a center port (detected by the breaking of an infrared beam) immediately illuminated a bright light emitting diode located above the head of the center port. When the rat removed its nose from the center port it was immediately reinforced with a food pellet (45 mg dustless grain pellets, Bio-Serv, Frenchtown, NJ, USA) delivered to the feeder trough associated with a sound generated from the overhead speaker; i.e. left feeder for NBN and right feeder for AM and Quiet (see below for details on acoustic stimuli). Once rats learned to nose-poke for food-pellets, the rat’s home cage food intake was adjusted so that their weights steadily increased with age while maintaining motivation to perform at least 100 trials in a session. Rats were run in 1 h training sessions once per day, 6 days per week.
Once rats exhibited frequent nose poking behavior into the center port (typically within 2 days), the behavioral response was transferred from the center port to the left feeder, associated with NBN, or to the right feeder which was associated with AM noise or quiet trials. Rats were required to nose poke to the center port and then respond to the correct feeder trough (detected by breaking an infrared beam at either trough) based on the sound presented; a correct response was immediately reinforced with a food pellet.
The purpose of this behavioral paradigm was to record and analyze oscillatory LFP activity during the behavior. In order to resolve low frequency oscillatory activity, a relatively long continuous trace of neural activity was required. Therefore rats were required to maintain relatively still for an interval (at least 4 seconds) before responding to the GO cue. The duration rats were required to hold their nose in the center port was dynamically adjusted using a computer controlled adaptive algorithm which progressively increased the delay to the GO cue in small steps (+250 ms for 5 consecutive hits) until they could wait for more than 4 seconds. Once rats were highly proficient at the task (hit-rate ≥ 80% for all sound conditions for at least 2 consecutive days) the rate of reinforcement was progressively reduced from 100% to 70%. We observed that reducing the rate of reinforcement seemed to improve performance on the paradigm.
One of the three acoustic conditions was always present while the rat was in the behavior box, regardless of whether the rat had initiated a trial or not. Acoustic conditions only changed after the rat had responded to one of the feeder troughs which completed the trial. A correct response to a narrow-band noise (NBN: 1/8th octave band, center frequencies randomized: 4, 8, 16, 22.6, or 32 kHz) was paired with the left feeder trough. The center frequency of the NBN stimulus was varied across trials in order to avoid making an assumption about the pitch of the tinnitus percept. A correct response to either an amplitude modulated (AM: broad-band noise, 100% modulation depth, 5 Hz) or Quiet (speaker off) trial was assigned to the right feeder trough. Correct responses were immediately reinforced with a food pellet delivered to the corresponding trough. Incorrect responses were indicated by turning off the house light and punished with a 20 second time-out period during which a new trial could not be initiated. NBN trials (left feeder trough) were presented for 50% of trials, whereas AM and Quiet (right feeder trough) trials were presented for 30% and 20% of trials, respectively. In order to prevent habits of alternation where a subject can perform above chance (> 50% hit-rate) using guessing strategies, trial sequences were selected according to specific criteria outlined by (Gellermann (1933); Heffner, 2011). We observed that implementation of these trial selection criteria reduced guessing behavior and improved performance on the task. Once proficient at the task, a rat was permitted up to 1 h to perform up to 125 trials. Naïve rats required between two to three months of training to become proficient on this task (data not shown).
2.4. Testing for Acute Tinnitus
Subjective tinnitus following a moderate dose of salicylate is characterized by the perception of a steady sound (McFadden et al. 1984). Therefore, if a rat was experiencing a steady tinnitus on Quiet trials after salicylate treatment, then responses on Quiet trials would be expected to shift from the right feeder (Figure 1B, previously a correct response) to the left feeder (Figure 1C, previously an incorrect response). In other words, tinnitus would be more similar to a steady NBN than AM noise.
Baseline behavior was established using an animal’s performance for each acoustic condition (NBN, AM, and Quiet) on the 4 most recent training days prior to administering saline control or sodium salicylate. One day of each week was considered a ‘testing day.’ On testing day, the reinforcement schedule described in the previous section was modified such that responses to either feeder trough (left or right) during Quiet trials were never reinforced or punished, but a response to either feeder trough was still required to initiate a new trial. Once trained, none of the rats were observed attempting to re-initiate a new trial without first responding to a feeder trough. Since Quiet trials accounted for 20% of all trials, the reinforcement rate of both NBN and AM trials was increased from 70% during baseline to 90% during testing to compensate for the removal of food pellets on quiet trials. Consequently, the overall rate of reinforcement in a testing session would be approximately equivalent to that received during baseline days.
Tinnitus-like behavior was defined as a significant drop in responses to the correct feeder during Quiet trials on testing day compared to the response rate on Quiet trials during the preceding 4 baseline days. The criterion used to define the presence of tinnitus in individual rats was evaluated by calculating z-scores based on the preceding 4 baseline days for each acoustic condition separately. Performance on testing day was tested against the baseline sample distribution of performance on each acoustic condition separately. The null hypothesis that no tinnitus was perceived was rejected if the computed z-score for the Quiet condition was less than the one-tailed criterion of p < 0.01 (z < mean - standard deviation * 2.33; similar to Heffner, 2011).
The raw hit rates of all animals as a group were compared across testing conditions using a repeated measures two-way analysis of variance. The significance of changes between saline vehicle control and sodium salicylate within each sound condition was determined using the Bonferroni post-test.
2.5. Inducing Tinnitus
On testing day, either a single injection of sodium salicylate (200 mg/kg, 50 mg/ml IP, referred to also as salicylate), dissolved in normal saline, or an equivalent volume of normal saline, was administered 2 h before beginning testing. This dose of salicylate has been demonstrated to reliably induce temporary tinnitus-like behavior in rats (Lobarinas et al., 2004). One rat was excluded for not performing a sufficient number of trials following salicylate injection (< 100 trials), possibly due to discomfort resulting from the high dose of salicylate.
2.6. Electrode Implantation Procedure
A subset of the fully trained rats were chronically implanted with a 16-channel polyimide insulated, tungsten microwire electrode array (wire diameter 33 µm, wire length ~1 mm, wires spaced 250 µm × 375 µm, arranged 2 × 8; Tucker-Davis Technologies). The microwires were laser cut at a 60° angle to minimize dimpling during insertion into cortex. Microwire electrodes were fixed to a flexible cable so the zero-insertion force headstage connector could be mounted to the top of the skull. In order to reduce the immune response associated with electrode implantation and to increase the long-term viability of electrode recordings, rats were treated with minocycline in their home cage drinking water (100 mg/L) for 2 days pre- and 5 days post-implantation (Rennaker et al., 2007).
Surgical plane of anesthesia was induced with a systemic injection (intraperitoneal) of a ketamine (70 mg/kg) and xylazine (7 mg/kg) mixture and maintained with smaller intramuscular doses as needed to maintain an areflexive surgical plane for the duration of the surgery. The animal was placed on a homeothermic heating pad (maintained core temperature at ~37° C), fixed in a standard stereotaxic frame with blunted ear bars, and the scalp swabbed with alcohol and then triiodine. An incision was made in the scalp midline, connective tissue was removed by blunt-dissection around the skull, and the top of the skull scraped with the scalpel blade to remove any remaining tissue. Six bone screws were fixed in the skull; 1 in the left and right frontal bones and 2 in the left and 2 in the right parietal bones. The muscle overlying the left temporal bone was resected, clamped with a hemostat, and mostly removed. A small craniotomy, centered −4.5 mm caudal to bregma and approximately 3 – 5 mm ventral from the temporal bone ridge, was made using a motorized drill with a carbide drill bit, flushing with saline every few seconds to avoid overheating of underlying cortex. A small aluminum flathead rivet was fixed on bregma with a drop of superglue and dental cement so that the animal could be removed from the stereotaxic frame to simplify insertion of the microwire array into cortex.
The animal was carefully removed from the stereotaxic frame, transferred to a platform and stabilized to the aluminum head-post using a custom made holding device. The arrangement provided an open field to work. In some animals, a single tungsten electrode (impedance ≈ 2 MΩ, FHC, Bowdin, ME, USA) was used to map out the excitatory frequency receptive field of multiunit activity in the AC prior to implanting the microwire electrode. The caudal-rostral, low-high frequency tonotopic gradient of primary AC (Polley et al., 2007) was used to identify the optimal site for electrode array implantation.
The dura overlying the AC was carefully resected and a silver ground wire was tightly wrapped around a screw fixed in the parietal bone ipsilateral to the craniotomy. The landing bridge which secured the tungsten microwires in the array to the flexible cable was fixed to a stiff copper wire with a small drop of nail acrylic. The copper rod was used to support the array during insertion into cortex using a micromanipulator. The microwire array was rapidly inserted into exposed cortex with the long axis of the array oriented along the caudal-rostral axis and with rostral electrodes slightly dorsal to the caudal electrodes to approximate the tonotopic gradient of primary AC. The array was implanted at a depth of ~500 µm from the cortical surface during insertion. The microwire array was subsequently covered with a non-toxic compound (Kwik-Sil, WPI Inc., Sarasota, FL, USA) to protect the brain and to secure the array to the skull. The array was then fixed to the temporal bone and bone screws on the top of the skull with dental cement. The headstage connector attached to the microwire array was routed to the top of the skull and fixed with dental cement. Rats were allowed 5 days to recover from surgery before returning to training on the behavioral tinnitus task. In order to accustom the rats to being tethered while performing the behavior, implanted rats were routinely tethered to the headstage and commutator during training sessions.
2.7. Electrophysiology Data Analysis
Electrophysiological data was recorded only on testing days. Multichannel extracellular signals were digitized and streamed to disk at ~25 kHz sampling rate (RZ5-2, TDT) for offline filtering and analysis. The streamed data was down-sampled to ~1.2 kHz and low-pass filtered (cutoff at 250 Hz) for the LFP signal. The continuous LFP was segmented into 4 s epochs preceding the onset of the GO cue (see Figure 2). Individual epochs were subjected to multitaper spectral analysis (Chronux v2.10 toolbox for Matlab, Bokil et al., 2010; Thomson, 1982). The multi-taper approach applies optimal windowing functions (Slepian sequences) to a time-domain signal, reducing frequency bias of the spectral estimation compared to arbitrarily chosen windowing functions (such as with Hanning or Hamming functions) (Bokil et al., 2010; Thomson, 1982). The frequency bandwidth parameter, W, of the multi-taper sequence is an explicitly controlled parameter which maximally concentrates energy in the frequency range of [f − W, f + W]. An estimate of the averaged multi-taper spectrum for multiple trials can be realized as:
Figure 2.
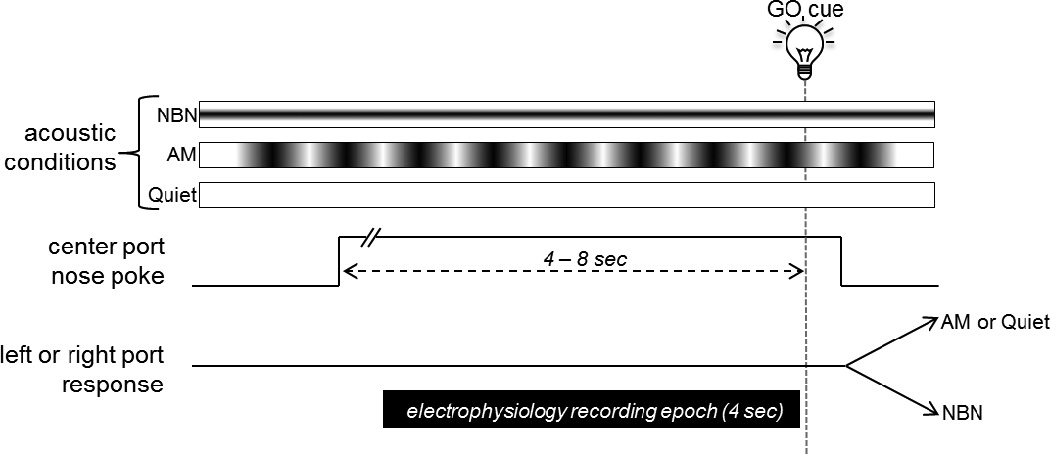
Schematic representation of the paradigm. One of three possible acoustic conditions (NBN, AM, Quiet) are presented from the overhead speaker. The rat initiates a trial by poking its nose in the center port and waiting a variable interval (4 – 8 s) for a bright light serving as the GO cue. After illumination of the GO cue, the rat must then identify the acoustic condition by responding at the left or right feeder trough. Correctly pairing the NBN with the left feeder trough or the AM or Quiet with the right feeder trough was reinforced with a food pellet at a variable rate of reinforcement. For electrophysiological recordings, a 4 s epoch was extracted from the streamed data set, filtered, and subjected to spectral analysis.
The multi-taper spectral estimate Ŝ(f) of N trials is computed by applying K orthogonal tapers u, (K = 2WT − 1, and T is the duration of the time signal in seconds) to the time-domain data, Xn, for each trial. The discrete Fourier transform is applied to each tapered trial signal and the resulting spectra are averaged. This procedure results in a principled trade-off between spectral variance and a reduction of precision in frequency resolution in the power spectrum (Bokil et al. 2006). Because of this trade-off, and since oscillatory neural activity occurring at higher frequencies tends to span broader bandwidth than at lower frequencies, we chose to analyze low frequencies (< 20 Hz) with narrower bandwidth tapers (2W = 2 Hz) than the high frequencies (20 – 150 Hz; 2W = 8 Hz).
The multi-taper approach to spectral estimation has also been extended to provide a robust statistical comparison between conditions and within a subject. Bokil et al. (2007) demonstrated that by combining Jackknife estimates of variance of two conditions, one can generate confidence intervals to evaluate the null hypothesis that the two spectra are equal. The multitaper approach effectively smoothes data across neighboring frequency bins within the range of [−W, +W] and therefore significant deviations smaller than 2W may be spurious. Only continuous bands of significant deviations larger than the full taper bandwidth 2W are considered when interpreting the statistic (Bokil et al., 2007).
3. Results
3.1. Salicylate-induced tinnitus behavior
Behavioral results from a typical rat tested 2 h following treatment with saline or salicylate (200 mg/kg, IP) are presented in figure 3. Saline treatment did not alter the rat’s ability to correctly identify Quiet trials. Moreover, the rat still correctly identified NBN (left feeder) and AM (right feeder) trials after saline injection. This result indicates that the behavioral paradigm is robust regardless of altering reinforcement rates during testing day (see section 2.4, Testing for Acute Tinnitus). Following salicylate treatment, however, the rat shifted its response on Quiet trials from the right feeder (associated with AM and Quiet) to the left feeder (associated with NBN); behavior consistent with the rat hearing the phantom sound of tinnitus on Quiet trials. The rat still correctly identified NBN (left feeder) and AM (right feeder) trials after salicylate. This result indicates that the rat is still able to discriminate between the acoustic stimuli and therefore any hearing loss attributable to salicylate (Cazals, 2000) is not a confounding factor in the test.
Figure 3.

Performance of an individual rat on the behavioral task following systemic treatment with saline or salicylate. Top panel: For each of the acoustic conditions (AM, Quiet and NBN), the proportion of trials identified as NBN is plotted over 16 days of testing. Bottom panel: When the rat’s behavioral performance was plotted according to z-score, saline treatment (Day 5) did not significantly affect the rat’s ability to identify Quiet or the other two conditions (NBN or AM), whereas following treatment with salicylate (Day 10), the rat incorrectly identified the Quiet condition as NBN significantly more than baseline indicating the presence of a steady NBN-like phantom sound (i.e., tinnitus). The horizontal black line in the bottom panel indicates the one-tailed criterion for significance.
None of the 7 rats exhibited tinnitus-like behavior after saline treatment (Figures 4A and 5) indicating the behavior is robust; i.e., the rats selected the correct feeder on Quiet trials despite not being reinforced or punished for these trials on testing day. Following salicylate (200 mg/kg IP), 6 of 7 rats shifted from responding to the right feeder (AM, Quiet) to the left feeder (NBN) on Quiet trials; behavior suggesting that 6 of the 7 rats were perceiving a phantom sound following systemic treatment with salicylate (Figure 4B).
Figure 4.

The effect of saline and salicylate on behavioral performance of trained rats. Panels A and B: Performance of 7 rats on Quiet trials on the days before (Days −4 to −1), during (Day 0) and after (Days 1 to 4) systemic treatment with saline or salicylate. 2 h after saline injection (Day 0; panel A), all of the rats correctly identified Quiet trials as such, whereas 2 h following injection with salicylate (200 mg/kg IP), 6 of 7 rats misidentified Quiet trials as NBN trials indicating the presence of a steady phantom sound (Day 0; panel B). The horizontal black line indicates the one-tailed criterion for significance.
Figure 5.

Performance of trained rats (N = 7) to identify the three acoustic conditions following systemic treatment with saline or salicylate. Panels A and B present the mean (±SD) proportion of trials identified as NBN over days relative to treatment (day 0). Panel C compares performance of rats (each circle-square pair represents one rat) between saline and salicylate treatments. (A, C) AM and Quiet conditions were infrequently misidentified as NBN, whereas NBN was identified correctly following saline treatment. Following salicylate (B, C), Quiet conditions were significantly more likely to be misidentified as NBN. This indicates that, as a group, salicylate induces phantom sound perception in rats. ns = not significant; *** p < 0.001.
Group performance on this behavioral task was assessed by comparing hit rates for each condition following saline or salicylate injection (Figure 5). Results presented in figure 5 are shown in reference to the proportion of responses to the left feeder which was associated with a steady NBN feeder trough. The proportion of responses to the left NBN feeder on AM, Quiet and NBN trials were determined separately for two experimental conditions, saline and salicylate. Following saline, rats infrequently misidentified Quiet trials (i.e., no evidence of tinnitus). However, following salicylate treatment rats identified Quiet trials as NBN significantly more than following saline (Figure 5C); behavior consistent with the perception of tinnitus during Quiet intervals. AM trials were still correctly identified as AM and rarely misidentified as NBN trials following salicylate (two-tailed paired t tests on saline vs. salicylate: Quiet t(6) = 6.346, p < 0.001; AM t(6) = 0.6297, p = 0.5521, NBN t(6) = 2.086, p = 0.0821).
3.2. Recordings from auditory cortex
To test the feasibility of acquiring electrophysiological data while rats were reporting experiencing tinnitus, we recorded from electrodes chronically implanted in AC while rats were discriminating Quiet from AM and NBN trials. Behavioral and electrophysiological measurements were obtained concurrently following treatment with saline and salicylate. Multichannel recordings of LFPs were obtained from a chronically implanted rat (performance of this rat is presented in Figure 3) while performing the behavioral task following systemic saline or salicylate injection. Time-domain signals were extracted from a 4 s period preceding the onset of the GO cue (see Figure 2) and subjected to multi-taper spectral analysis. Taper bandwidths (2W) of 2 Hz and 8 Hz were used for spectral analysis of low and high frequencies, respectively (see section 2.7, Electrophysiology Data Analysis). The characteristic frequency of multiunit frequency receptive field of the electrode, recorded under ketamine anesthesia following implantation, was ~6 kHz (data not shown). Mean data are shown for low (Figure 6, left column) and high frequency (Figure 6, right column) components of the LFP obtained during NBN, Quiet, and AM trials. Each panel shows data obtained after saline and salicylate treatments. Salicylate treatment caused a decrease in power in the theta (~5 Hz) and alpha bands (~10 Hz), an increase in the low gamma band (centered at ~50 Hz), and a broadband decrease in the high gamma band (> 100 Hz). Similar changes in power spectra were observed for NBN and Quiet trials, whereas power spectra for AM trials exhibited a relatively smaller increase in low gamma than the other two conditions. The observation that power spectra were altered during all trial types indicates that tinnitus may have been present during most trials regardless of the presence of a real sound; however, the AM stimulus was sufficiently salient for the rat to still identify correctly.
Figure 6.
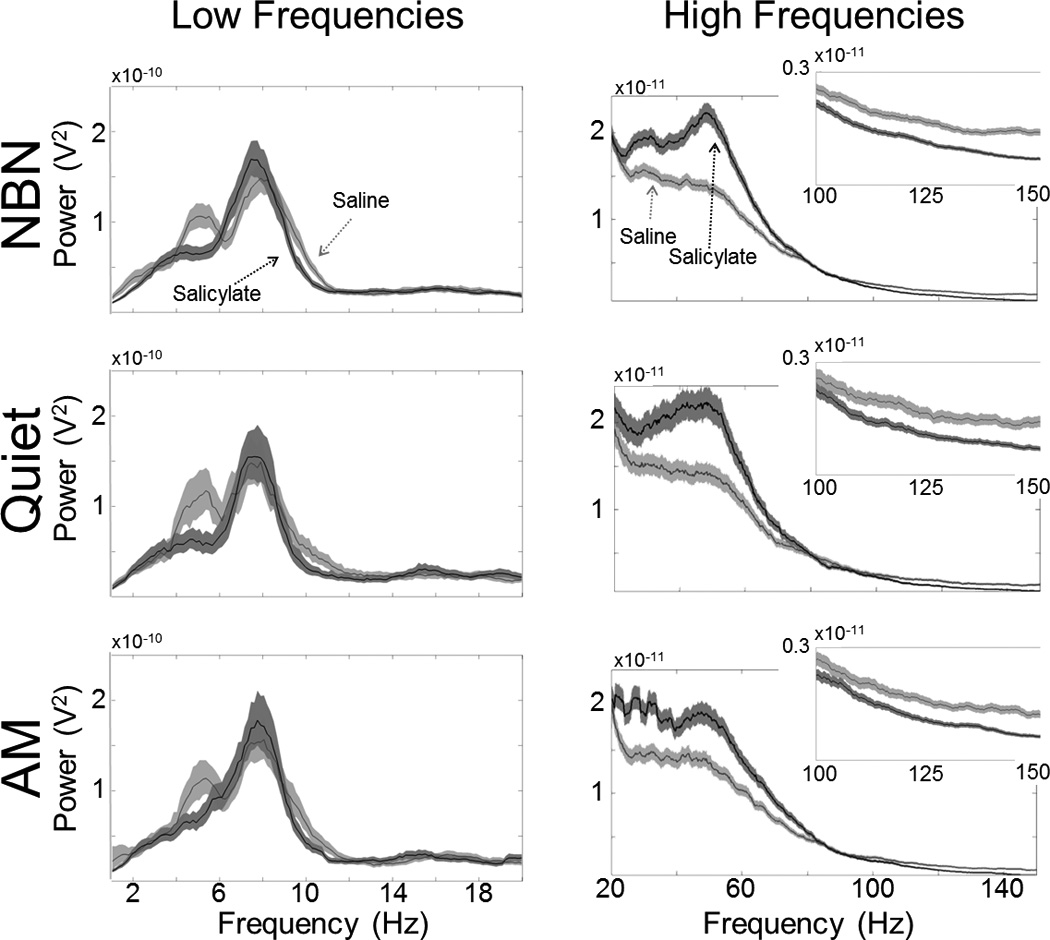
Salicylate-induced tinnitus corresponds to changes in ongoing oscillatory activity in auditory cortex. AC LFP activity was recorded from chronically implanted microwire electrode array of the rat performing the behavioral task (same animal as in figure 2). Power spectra were computed from a 4 s epoch preceding the GO cue (see figure 1). Low (< 20 Hz; W = 1 Hz, K = 3) and high (20 – 150 Hz; W = 4 Hz, K = 15) frequency components of the LFP were separately subjected to multitaper spectral analysis (see Methods). Light and dark gray bands represent jackknife estimate of 95% confidence intervals following saline or salicylate, respectively. High frequency plots are magnified 10 times compared with low frequencies. Insets plots rescale very high frequencies (100 – 150 Hz) for clarity.
4. Discussion
This report demonstrates the efficacy of a novel two-alternative choice identification task for detecting acute tinnitus perception in rats; the relatively long hold period and reduced motor activity during Quiet, AM and NBN conditions was optimized for the simultaneous recording of neural activity while the rat was attending to one of three acoustic conditions. In agreement with previous studies (Jastreboff et al., 1988; Lobarinas et al., 2004), a single moderately high dose of salicylate (200 mg/kg, IP) reliably induced acute tinnitus in rats. A unique feature of this behavioral paradigm is that it provides relatively long epochs (4 s was used in this study) during which neural activity on slow and fast scales can be sampled and analyzed. Furthermore, this paradigm has particular utility in the recording of ongoing neural activity because the rats are relatively quiescent during the extended holding period thereby reducing motion artifacts from the recordings. Finally, since tinnitus is a novel auditory sensation to which the animal is likely attending in order to make an informed response, sampling electrophysiological activity while the animal is attending and responding to different sounds of similar quality (NBN) serves as an appropriate control for neural activity related to changes in arousal while the animal experiences drug-induced tinnitus.
In addition to our efforts to control for attention and to record neural activity during the moments when tinnitus was reported behaviorally, we also designed this behavioral paradigm to be largely resistant to the hearing loss that often accompanies drug- and noise-induced tinnitus. For example, a moderate hearing loss would not be expected to impair the animal’s performance during this task; provided the animal can still hear the ~60 dB NBN and AM stimuli. In this particular case, performance during our study was not significantly altered by salicylate-induced hearing loss (figures 4 and 5).
In this report we have demonstrated that this novel behavioral paradigm can detect acute salicylate-induced tinnitus in rats. This paradigm will likely be of use in detecting and recording neural activity following acute noise trauma which may be more similar to how humans typically acquire tinnitus. It should be noted, however, that the metric used in this paradigm to assess the presence of tinnitus in individual subjects is based on comparison with a recent history of performance (see figures 3, 4, and 5), and as such, the paradigm may need to be modified to effectively evaluate chronic tinnitus.
The analysis of LFP activity from AC recorded while an animal reported tinnitus perception demonstrates proof of concept that this behavioral paradigm provides the appropriate conditions for measuring neural activity, including oscillatory activity, while an animal is actively informing the investigator that it is experiencing tinnitus during Quiet intervals. In this study, a spectral analysis of the neural activity revealed a significant decrease in alpha and an increase in low gamma band activity during a time when the animal was reporting tinnitus-like behavior (Figure 6). The preliminary results reported here are similar to neuromagnetic recordings for humans with acute tinnitus (Lorenz et al., 2009). However, some features of the power spectrum during salicylate-induced tinnitus differ from reports of humans with tinnitus. In particular, a significant decrease in theta band activity (4 – 6 Hz) diverges from the results reported in a study in humans by Llinas et al. (1999) which noted an increase in this frequency band in subjects with tinnitus. We also did not observe any significant changes in slow-wave delta band (< 4 Hz) activity which has been proposed to be critical in tinnitus perception (Kahlbrock and Weisz, 2008; Weisz et al., 2007a; Weisz et al., 2007b). That we did not observe these slow-wave changes, however, may be due to a high-pass filter at 2 Hz in our pre-amplifying hardware or that acute tinnitus was pharmacologically induced compared with human subjects with chronic tinnitus. We also observed a broadband decrease in the high gamma band (> 100 Hz) which may reflect a decrease in multiunit spiking activity (Ray and Maunsell, 2011) which has been reported previously in the passive rat AC following treatment with salicylate (Yang et al., 2007).
While the electrophysiological results reported here are preliminary and included as proof of principle, a larger sample size and additional manipulations are required to specifically evaluate the hypothesis that oscillatory activity plays a critical role in tinnitus production. Future studies may utilize this behavioral paradigm with chronically implanted, moveable electrodes to investigate single neuron activity during acute tinnitus in various brain regions. This protocol will also facilitate the comparison of neural activity related to the perception of a real acoustic stimulus with that of a phantom sound perception. Furthermore, this behavioral paradigm was designed specifically for rats which are easily trained on appetitive conditioning paradigms; however, we believe it may be possible to utilize this paradigm to judge the presence of tinnitus in other animal species such as guinea pigs and primates, whereas mouse behavior may be too frenetic to maintain still for the duration of the recording period. Understanding the neural correlates of tinnitus perception in awake animals actively reporting phantom sound perception will provide better strategies for pharmacological or other interventions, such as transcranial magnetic stimulation, in the treatment of tinnitus.
Highlights.
-
□
A novel behavioral task was developed for detecting tinnitus perception in rats.
-
□
The task was designed for simultaneous recording of neural activity.
-
□
Tinnitus was identified following a single systemic dose of sodium salicylate.
-
□
Significant changes in oscillatory neural activity were detected and discussed.
Acknowledgements
We would like to acknowledge the contribution of Erickson Nielans and Michael Dent, Ph.D. for their helpful consultation on the behavioral methods and analysis.
Research supported in part by NIH National Institute on Deafness and Other Communication Disorders NRSA fellowship (5F31DC010931, DS), NIH grants (R01DC009091 and R01DC009219, RS; and R03DC011374, BLA) and Office of Naval Research (N0001412107, RS)
Abbreviations
- AC
auditory cortex
- LFP
local field potential
- NBN
Narrow-band noise
- AM
Amplitude modulated noise
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
Sarah H. Hayes, Email: shhayes@buffalo.edu.
Nina Kashanian, Email: ninakash@buffalo.edu.
Kelly Radziwon, Email: radziwon@buffalo.edu.
Richard J. Salvi, Email: salvi@buffalo.edu.
Brian L. Allman, Email: brian.allman@schulich.uwo.ca.
References
- Adjamian P, Sereda M, Zobay O, Hall DA, Palmer AR. Neuromagnetic Indicators of Tinnitus and Tinnitus Masking in Patients with and without Hearing Loss. J Assoc Res Otolaryngol. 2012;13:715–831. doi: 10.1007/s10162-012-0340-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashton H, Reid K, Marsh R, Johnson I, Alter K, Griffiths T. High frequency localised "hot spots" in temporal lobes of patients with intractable tinnitus: a quantitative electroencephalographic (QEEG) study. Neurosci Lett. 2007;426:23–48. doi: 10.1016/j.neulet.2007.08.034. [DOI] [PubMed] [Google Scholar]
- Basar E, Basar-Eroglu C, Karakas S, Schurmann M. Brain oscillations in perception and memory. Int J Psychophysiol. 2000;35:95–124. doi: 10.1016/s0167-8760(99)00047-1. [DOI] [PubMed] [Google Scholar]
- Bauer CA, Brozoski TJ, Rojas R, Boley J, Wyder M. Behavioral model of chronic tinnitus in rats. Otolaryngol Head Neck Surg. 1999;121:457–462. doi: 10.1016/S0194-5998(99)70237-8. [DOI] [PubMed] [Google Scholar]
- Berger JI, Coomber B, Shackleton TM, Palmer AR, Wallace MN. A novel behavioural approach to detecting tinnitus in the guinea pig. J Neurosci Methods. 2013;213:188–195. doi: 10.1016/j.jneumeth.2012.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bokil H, Andrews P, Kulkarni JE, Mehta S, Mitra PP. Chronux: a platform for analyzing neural signals. J Neurosci Methods. 2010;192:146–151. doi: 10.1016/j.jneumeth.2010.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bokil H, Purpura K, Schoffelen JM, Thomson D, Mitra P. Comparing spectra and coherences for groups of unequal size. J Neurosci Methods. 2007;159:337–345. doi: 10.1016/j.jneumeth.2006.07.011. [DOI] [PubMed] [Google Scholar]
- Buzsaki G, Anastassiou CA, Koch C. The origin of extracellular fields and currents--EEG, ECoG, LFP and spikes. Nat Rev Neurosci. 2012;13:407–420. doi: 10.1038/nrn3241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cazals Y. Auditory sensori-neural alterations induced by salicylate. Prog Neurobiol. 2000;62:583–631. doi: 10.1016/s0301-0082(00)00027-7. [DOI] [PubMed] [Google Scholar]
- Dehmel S, Pradhan S, Koehler S, Bledsoe S, Shore S. Noise overexposure alters long-term somatosensory-auditory processing in the dorsal cochlear nucleus--possible basis for tinnitus-related hyperactivity? J Neurosci. 2012;32:1660–1671. doi: 10.1523/JNEUROSCI.4608-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fries P. Neuronal gamma-band synchronization as a fundamental process in cortical computation. Annu Rev Neurosci. 2009;32:209–224. doi: 10.1146/annurev.neuro.051508.135603. [DOI] [PubMed] [Google Scholar]
- Gellermann LW. Chance orders of alternating stimuli in visual discrimination experiments. The Pedagogical Seminary and Journal of Genetic Psychology. 1933;42:206–228. [Google Scholar]
- Guitton MJ, Caston J, Ruel J, Johnson RM, Pujol R, Puel JL. Salicylate induces tinnitus through activation of cochlear NMDA receptors. J Neurosci. 2003;23:3944–3952. doi: 10.1523/JNEUROSCI.23-09-03944.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heffner HE. A two-choice sound localization procedure for detecting lateralized tinnitus in animals. Behav Res Methods. 2011;43:577–589. doi: 10.3758/s13428-011-0061-4. [DOI] [PubMed] [Google Scholar]
- Heffner HE, Harrington IA. Tinnitus in hamsters following exposure to intense sound. Hear Res. 2002;170:83–95. doi: 10.1016/s0378-5955(02)00343-x. [DOI] [PubMed] [Google Scholar]
- Jastreboff PJ, Brennan JF, Coleman JK, Sasaki CT. Phantom auditory sensation in rats: an animal model for tinnitus. Behav Neurosci. 1988;102:811–822. doi: 10.1037//0735-7044.102.6.811. [DOI] [PubMed] [Google Scholar]
- Kahlbrock N, Weisz N. Transient reduction of tinnitus intensity is marked by concomitant reductions of delta band power. BMC Biol. 2008;6:4. doi: 10.1186/1741-7007-6-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kisley MA, Gerstein GL. Trial-to-trial variability and state-dependent modulation of auditory-evoked responses in cortex. J Neurosci. 1999;19:10451–10560. doi: 10.1523/JNEUROSCI.19-23-10451.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas RR, Ribary U, Jeanmonod D, Kronberg E, Mitra PP. Thalamocortical dysrhythmia: A neurological and neuropsychiatric syndrome characterized by magnetoencephalography. Proc Natl Acad Sci U S A. 1999;96:15222–15237. doi: 10.1073/pnas.96.26.15222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lobarinas E, Sun W, Cushing R, Salvi R. A novel behavioral paradigm for assessing tinnitus using schedule-induced polydipsia avoidance conditioning (SIP-AC) Hear Res. 2004;190:109–114. doi: 10.1016/S0378-5955(04)00019-X. [DOI] [PubMed] [Google Scholar]
- Longenecker RJ, Galazyuk AV. Methodological optimization of tinnitus assessment using prepulse inhibition of the acoustic startle reflex. Brain Res. 2012;1485:54–62. doi: 10.1016/j.brainres.2012.02.067. [DOI] [PubMed] [Google Scholar]
- Lorenz I, Muller N, Schlee W, Hartmann T, Weisz N. Loss of alpha power is related to increased gamma synchronization-A marker of reduced inhibition in tinnitus? Neurosci Lett. 2009;453:225–238. doi: 10.1016/j.neulet.2009.02.028. [DOI] [PubMed] [Google Scholar]
- Moazami-Goudarzi M, Michels L, Weisz N, Jeanmonod D. Temporo-insular enhancement of EEG low and high frequencies in patients with chronic tinnitus. QEEG study of chronic tinnitus patients. BMC Neurosci. 2010;11:40. doi: 10.1186/1471-2202-11-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mongan E, Kelly P, Nies K, Porter WW, Paulus HE. Tinnitus as an indication of therapeutic serum salicylate levels. JAMA. 1973;226:142–145. [PubMed] [Google Scholar]
- Norena AJ, Moffat G, Blanc JL, Pezard L, Cazals Y. Neural changes in the auditory cortex of awake guinea pigs after two tinnitus inducers: salicylate and acoustic trauma. Neuroscience. 2010;166:1194–1209. doi: 10.1016/j.neuroscience.2009.12.063. [DOI] [PubMed] [Google Scholar]
- Polley DB, Read HL, Storace DA, Merzenich MM. Multiparametric auditory receptive field organization across five cortical fields in the albino rat. J Neurophysiol. 2007;97:3621–3638. doi: 10.1152/jn.01298.2006. [DOI] [PubMed] [Google Scholar]
- Ray S, Maunsell JH. Different origins of gamma rhythm and high-gamma activity in macaque visual cortex. PLoS Biol. 2011;9:e1000610. doi: 10.1371/journal.pbio.1000610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rennaker RL, Miller J, Tang H, Wilson DA. Minocycline increases quality and longevity of chronic neural recordings. J Neural Eng. 2007;4:L1–L5. doi: 10.1088/1741-2560/4/2/L01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rüttiger L, Ciuffani J, Zenner H-P, Knipper M. A behavioral paradigm to judge acute sodium salicylate-induced sound experience in rats: a new approach for an animal model on tinnitus. Hearing Research. 2003;180:39–50. doi: 10.1016/s0378-5955(03)00075-3. [DOI] [PubMed] [Google Scholar]
- Sedley W, Teki S, Kumar S, Barnes GR, Bamiou DE, Griffiths TD. Single-subject oscillatory gamma responses in tinnitus. Brain. 2012;135:3089–3100. doi: 10.1093/brain/aws220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shargorodsky J, Curhan GC, Farwell WR. Prevalence and Characteristics of Tinnitus among US Adults. Am J Med. 2010;123:711–728. doi: 10.1016/j.amjmed.2010.02.015. [DOI] [PubMed] [Google Scholar]
- Singer W. Neuronal synchrony: a versatile code for the definition of relations? Neuron. 1999;24:49–65. 111–125. doi: 10.1016/s0896-6273(00)80821-1. [DOI] [PubMed] [Google Scholar]
- Thomson DJ. Spectrum Estimation and Harmonic-Analysis. Proceedings of the Ieee. 1982;70:1055–1096. [Google Scholar]
- Turner JG. Behavioral measures of tinnitus in laboratory animals. Prog Brain Res. 2007;166:147–156. doi: 10.1016/S0079-6123(07)66013-0. [DOI] [PubMed] [Google Scholar]
- Turner JG, Brozoski TJ, Bauer CA, Parrish JL, Myers K, Hughes LF, Caspary DM. Gap detection deficits in rats with tinnitus: a potential novel screening tool. Behav Neurosci. 2006;120:188–195. doi: 10.1037/0735-7044.120.1.188. [DOI] [PubMed] [Google Scholar]
- Turner JG, Parrish J. Gap detection methods for assessing salicylate-induced tinnitus and hyperacusis in rats. Am J Audiol. 2008;17:S185–S192. doi: 10.1044/1059-0889(2008/08-0006). [DOI] [PubMed] [Google Scholar]
- Tzounopoulos T. Mechanisms of synaptic plasticity in the dorsal cochlear nucleus: plasticity-induced changes that could underlie tinnitus. Am J Audiol. 2008;17:S170–S185. doi: 10.1044/1059-0889(2008/07-0030). [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Loo E, Gais S, Congedo M, Vanneste S, Plazier M, Menovsky T, Van deHeyning P, De Ridder D. Tinnitus intensity dependent gamma oscillations of the contralateral auditory cortex. PLoS One. 2009;4:e7396. doi: 10.1371/journal.pone.0007396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weisz N, Dohrmann K, Elbert T. The relevance of spontaneous activity for the coding of the tinnitus sensation. Prog Brain Res. 2007a;166:61–70. doi: 10.1016/S0079-6123(07)66006-3. [DOI] [PubMed] [Google Scholar]
- Weisz N, Moratti S, Meinzer M, Dohrmann K, Elbert T. Tinnitus perception and distress is related to abnormal spontaneous brain activity as measured by magnetoencephalography. PLoS Med. 2005;2:e153. doi: 10.1371/journal.pmed.0020153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weisz N, Muller S, Schlee W, Dohrmann K, Hartmann T, Elbert T. The neural code of auditory phantom perception. J Neurosci. 2007b;27:1479–1484. doi: 10.1523/JNEUROSCI.3711-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wienbruch C, Paul I, Weisz N, Elbert T, Roberts LE. Frequency organization of the 40-Hz auditory steady-state response in normal hearing and in tinnitus. Neuroimage. 2006;33:180–194. doi: 10.1016/j.neuroimage.2006.06.023. [DOI] [PubMed] [Google Scholar]
- Yang G, Lobarinas E, Zhang L, Turner J, Stolzberg D, Salvi R, Sun W. Salicylate induced tinnitus: behavioral measures and neural activity in auditory cortex of awake rats. Hear Res. 2007;226:244–253. doi: 10.1016/j.heares.2006.06.013. [DOI] [PubMed] [Google Scholar]
- Zhang X, Yang P, Cao Y, Qin L, Sato Y. Salicylate induced neural changes in the primary auditory cortex of awake cats. Neuroscience. 2011;172:232–245. doi: 10.1016/j.neuroscience.2010.10.073. [DOI] [PubMed] [Google Scholar]