

Abstract
Objectives
Few model systems are amenable to developing multi-species biofilms in parallel under environmentally germane conditions. This is a problem when evaluating the potential real-world effectiveness of antimicrobials in the laboratory. One such antimicrobial is cetylpyridinium chloride (CPC), which is used in numerous over-the-counter oral healthcare products. The aim of this work was to develop a high-throughput microfluidic system that is combined with a confocal laser scanning microscope (CLSM) to quantitatively evaluate the effectiveness of CPC against oral multi-species biofilms grown in human saliva.
Methods
Twenty-four-channel BioFlux microfluidic plates were inoculated with pooled human saliva and fed filter-sterilized saliva for 20 h at 37°C. The bacterial diversity of the biofilms was evaluated by bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). The antimicrobial/anti-biofilm effect of CPC (0.5%–0.001% w/v) was examined using Live/Dead stain, CLSM and 3D imaging software.
Results
The analysis of biofilms by bTEFAP demonstrated that they contained genera typically found in human dental plaque. These included Aggregatibacter, Fusobacterium, Neisseria, Porphyromonas, Streptococcus and Veillonella. Using Live/Dead stain, clear gradations in killing were observed when the biofilms were treated with CPC between 0.5% and 0.001% w/v. At 0.5% (w/v) CPC, 90% of the total signal was from dead/damaged cells. Below this concentration range, less killing was observed. In the 0.5%–0.05% (w/v) range CPC penetration/killing was greatest and biofilm thickness was significantly reduced.
Conclusions
This work demonstrates the utility of a high-throughput microfluidic–CLSM system to grow multi-species oral biofilms, which are compositionally similar to naturally occurring biofilms, to assess the effectiveness of antimicrobials.
Keywords: multi-species biofilm, confocal scanning laser microscopy, Live/Dead staining, microfluidics, pyrosequencing
Introduction
Biofilms are surface-attached multi-species microbial communities.1,2 These communities represent the dominant mode of microbial life.2 Biofilm microorganisms individually and collectively possess properties that are distinct from their single-cell planktonic counterparts.3,4 In particular, biofilm bacteria are up to 1000-fold less susceptible to antimicrobials than planktonic cells.4–6 The reasons behind this reduced susceptibility are multifactorial and include retarded antimicrobial penetration of biofilm due to reaction diffusion limitation,7,8 altered growth rates,4 intraspecies and interspecies metabolite and/or cell–cell signalling interactions resulting in altered biofilm-specific phenotypes,9 and cross-species protection afforded by removal or inactivation of a given antimicrobial by a biofilm species.4,10 The environment in which biofilms form may also alter recalcitrance. For example, it has been shown that Acinetobacter baumannii possesses very different resistance profiles in laboratory media versus on ex vivo human ascites.11 Thus, in order to evaluate the possible real-world effectiveness of antimicrobials on bacterial populations, it is important to develop biofilms that contain species present in the environment of interest and under environmentally germane conditions.
Dental plaque is a multi-species biofilm that can contain >500 species of bacteria.12,13 Typical dominant species in both supragingival and subgingival plaque biofilms include those belonging to the genera Actinomyces, Aggregatibacter, Fusobacterium, Neisseria, Porphyromonas, Streptococcus and Veillonella.14 Following professional cleaning, there is a rapid successional integration of species; members of the streptococci are among the first to colonize surfaces, followed by numerous other Gram-positive and Gram-negative species.15 Depending upon the position of the community on tooth surfaces (e.g. within a subgingival pocket or on a supragingival surface exposed to rapidly flowing saliva), the biofilm community composition will be significantly different.16 These biofilm communities are not only heterogeneous with respect to the species they contain but also can be architecturally diverse; for instance, they can range from a few cells thick to visually conspicuous biofilms.17–19 The more diverse the community and the greater the biofilm biomass, the more likely it is that pathogenic species such as Porphyromonas gingivalis and Treponema denticola will integrate and promote periodontal disease.15,20,21 Approaches to controlling the species composition and overall density of dental plaque biofilm communities encompass abrasive regimens (e.g. tooth-brushing and flossing) and chemical treatments (e.g. mouthwash).22–24 The antimicrobial efficacy of mouthwash on multi-species biofilms has received attention, although few models exist that facilitate testing under in vivo or environmentally relevant in vitro conditions.25–31
Numerous model biofilm systems exist to examine biofilm development and/or the impact of antimicrobial compounds. These can be large-scale systems suitable for long-term studies, such as newly modified Robbins devices, Sorbarod-based biofilm systems and constant-depth film fermenters, or, for shorter durations, devices such as flow cells.26,30–32 A critical drawback to the operation of such model systems is their physical footprint (resulting in limited capabilities for performing parallel replicate studies) and the often limiting requirement for large amounts of media in which to develop biofilms. This latter point is of great importance if the medium is expensive or time-consuming to obtain, especially if it is from natural sources (e.g. saliva or wound exudate). For example, when conducting flow cell studies, an overnight experiment can require >500 mL.12,33 A microfluidic system, either custom-made or available commercially, removes such a limitation and also allows, by virtue of its small footprint, multiple biofilm experiments to be run in parallel.34 The potential for linking such a system to 3D imaging systems is only now just being realized and an opportunity to create high-throughput screens of antimicrobial or biofilm-structure-altering compounds can be envisaged.35,36
From the outset of this work, we wished to create a biofilm system that used saliva as the sole nutrient and inoculum source. Many biofilm systems use either medium or artificial saliva as the nutrient source.35–38 This is primarily due to the inherent difficulties in collecting large enough quantities of human saliva. However, these types of artificial media can conceivably have significant ramifications for biofilm composition (if a multi-species community is being grown) and for the responsiveness of the individual species (as well as the entire community) to environmental changes or chemical challenges. As a consequence, the use of pooled human saliva as an inoculum and as a medium source is gaining popularity in model oral biofilm systems,12,39–41 although the problem of obtaining sufficient quantities is still an issue. A key driver of the use of saliva relates to its complex physical and chemical nature.42,43 In particular, saliva contains complex mixtures of mucins, phosphate-containing compounds (including nucleic acids), proline-rich proteins, statherin, cystatins, histatins, enzymes (amylase, lysozyme, proteases, etc.) and trace elements, such as boron and zinc.42 In addition, the use of such a medium as an inoculum is beneficial as it contains many, if not all, of the organisms found in dental plaque biofilms.44 Saliva is therefore a unique mixture which will influence the colonization, growth and survival of microorganisms, and a model system that wishes to replicate the conditions of the oral cavity will need to use this as the nutrient source.
The aim of the work presented here was to develop a high-throughput dental plaque biofilm system that: (i) facilitates the growth of saliva-derived and -supported microbial communities under conditions representative of the human oral cavity and (ii) is amenable to testing with antimicrobials. This required the amalgamation of BioFlux (Fluxion, San Francisco, CA) microfluidic technologies with a cutting edge Leica confocal laser scanning microscope system (SPE, Leica, Buffalo Grove, IL). Using bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP), the developed multi-species biofilms were shown to contain obligate aerobic, facultative anaerobic and obligate anaerobic species common to those found in human subgingival and supragingival plaque biofilms. Relevant to the use of this system for high-throughput testing, we show that the susceptibility of these multi-species biofilms to cetylpyridinium chloride (CPC), a quaternary ammonium compound that is commonly used in mouthwash, was concentration-dependent and subject to reaction diffusion limitation.
Materials and methods
Test solutions for assessing biofilm viability
A range of CPC solutions (0.5%, 0.1%, 0.05%, 0.01%, 0.005% and 0.001% w/v) were prepared. PBS, pH 7.4, and 70% v/v ethanol were prepared for use as negative and positive controls, respectively.
Saliva collection and preparation: inoculum and nutrient source
A saliva collection protocol similar to that used by Rao et al.45 was used. Briefly, saliva was collected from six healthy adults who had not consumed food for 2 h prior to donation and had only imbibed water during that time. These individuals were non-smokers and had not taken antibiotics for at least 3 months prior to donation. The saliva was then pooled and prepared for one of two purposes: to be used as a cell-containing saliva (CCS) inoculum or to be used as a cell-free saliva (CFS) nutrient source for biofilm growth. CCS was prepared by mixing native, pooled saliva with glycerol in a 75%/25% ratio, respectively, and then split into 5 mL aliquots for storage at −80°C. CFS was prepared by adding 2.5 mM DTT to the saliva,46 allowing to stand for 10 min on ice and then centrifuging at 17 500 rpm for 30 min. The resulting supernatant was mixed with distilled water to a final concentration of 25% and filter sterilized through 0.22 μm pore-size surfactant-free cellulose acetate (SFCA) low-protein-binding filters (Nalge Nunc International, Rochester, NY). Aliquots of ∼30 mL were then stored at −20°C until required.
BioFlux microfluidic system
Forty-eight-well BioFlux plates (Fluxion, San Francisco, CA), in conjunction with the BioFlux 200 system (Fluxion, San Francisco, CA) and a Leica SPE confocal laser scanning microscope (CLSM), were used as the base platform for the microfluidic dental plaque biofilm system (Figure 1). Microfluidic plates were first pre-treated for cell attachment and biofilm development with CFS. One hundred microlitres of CFS was added to each outlet well then flowed towards the inlet well at 1.0 dyn/cm2 (flow rate of 93 μL/h, corresponding to a shear of 100 s−1 through the channel, which had a depth of 70 μm and a width of 370 μm) for 2 min at room temperature. Flow was then stopped and the plate was incubated at room temperature for 20 min. Once pre-treatment incubation was complete, the CFS remaining in the outlet wells was transferred to the corresponding inlet wells. One hundred microlitres of CCS was then added to each outlet well. To introduce cells into the BioFlux growth/viewing channel for biofilm growth, the CCS was flowed towards the inlet at 1.0 dyn/cm2 for precisely 6 s at 37°C. The plate was then set to incubate at 37°C for 45 min to allow cell adherence and initial growth prior to nutrient flow. Surface seeding was confirmed visually with a Nikon Eclipse TCS-100 inverted light microscope equipped with a 20 × 0.40 NA Ph1 ADL infinity-corrected objective. Following confirmation of seeding, CCS was aspirated from each outlet well and 750 μL of CFS was added to each inlet well. Plates were then incubated at 37°C for 20 h at 0.2 dyn/cm2 (flow rate of 19 μL/h, corresponding to a shear of 20 s−1) for overnight biofilm growth.
Figure 1.

The microfluidic confocal scanning laser microscope biofilm system. (a) Diagram depicting a vertical cross-section of the BioFlux microfluidic system mounted on a Leica SPE CLSM. (b) Annotated photograph showing the channels for two microfluidic channels and accompanying reservoir and waste wells. Black annotations highlight positions of flow channels. (c) An example of a Live/Dead-stained dental plaque biofilm rendered in 2D and 3D using Imaris. Figure 1(b) is used with permission from Fluxion (Fluxionbio.com). Figure 1(a) is modified with kind permission from the American Society for Microbiology (Benoit et al. Appl Environ Microbiol 2010; 76: 4136–42).
Following overnight incubation, the CFS that remained in each inlet and outlet well was aspirated. Biofilms were then treated in triplicate with 100 μL of a given treatment solution for 30 s at a flow rate of 2.0 dyn/cm2. All wells were subsequently aspirated and 100 μL of PBS was flowed at room temperature for 20 min at 0.2 dyn/cm2 to remove remaining treatment solution. Following the post-treatment wash, biofilms were stained with a BacLight LIVE/DEAD bacterial viability kit (Invitrogen, Grand Island, NY). The concentrations of component BacLight solutions were prepared according to the manufacturer's instructions and flowed at 0.2 dyn/cm2 for 45 min at room temperature to allow staining of the biofilms. Once staining was complete, stain remaining in the inlet wells was aspirated and a final 100 μL post-stain wash of PBS was added to each inlet. The post-stain PBS wash was flowed at 0.2 dyn/cm2 for 20 min at room temperature to remove any excess stain in the channel.
CLSM analysis of biofilms
Using a Leica SPE confocal laser scanning microscope, biofilms were imaged using a 40 × 1.25 NA HCX PL APO infinity-corrected oil objective. Image stacks were collected by exciting the biofilm sample with a 488 nm laser set to 15% power. This allowed the simultaneous excitation of Syto-9 and propidium iodide (both nucleic acid stains). Excitation capture ranges were kept constant for Syto-9 and propidium iodide (510–540 and 620–650 nm, respectively). Gain and offset for each channel (red and green) were calibrated using the Leica look-up table against the positive (70% ethanol) and negative (PBS) controls for the analysis of the CPC-treated biofilms. Following biofilm imaging, biofilms were rendered in the x–y–z planes with Imaris (Bitplane, Zurich, Switzerland) imaging software. Image stacks were treated equally and histograms generated in Imaris confirmed that the signal display of the rendered stack was correctly displayed in 3D. This allowed the 3D visualization of biofilm structure, penetration of antimicrobial into biofilms (inferred by the extent and degree of red signal) and the preparation of 3D files for the quantification of biofilm structure using the software program Comstat.47 Comstat software allowed the determination of biofilm biovolume (the amount of space/biomass occupied by a biofilm per field of view), average thickness (the average thickness, from the base at the biofilm–substratum interface to the top of the biofilm in the channel lumen, across the entire biofilm in the field of view) and roughness (a measure of variation in biofilm thickness across the field of view, an indicator of biofilm heterogeneity). The degree of kill, based on green (live) and red (dead) pixel intensity for every pixel in the x–y–z planes, was evaluated using ImageJ.48 The percentage red signal (dead/damaged cells) and green signal (viable cells) was determined by first multiplying the total number of pixels at a given intensity by the intensity value (0–255) then summing the total value for both the LIVE and DEAD signals from each image stack taken. This allowed the percentage viability (green signal) to be determined. All renderings and quantification analyses were performed on a dedicated computer equipped with an Intel i5 processor (Intel, Santa Clara, CA) supported with a Radeon 5850 1 Gb graphics card (AMD, Sunnyvale, CA). Captured renderings were assembled in CorelDRAW v. X4 (Corel, Mountain View, CA).
Culture-independent analysis
Biofilms were grown according to the protocol described above, washed in PBS and harvested. Biofilm cells were harvested by pulsing sterile distilled H2O through the channels at 8.0 dyn/cm2 (flow rate 745 μL/h, corresponding to a shear of 800 s−1) in both the forward and the reverse direction, in order to create shear stress to remove biofilm cells. This washing procedure was repeated until a visually large portion of the bound biofilm cells had been removed. Harvested cells were collected and stored at −80°C.
In order to conduct culture-independent analyses, bTEFAP, using the primers 939F (TTGACGGGGGCCCGCAC) and 1492R (TACCTTGTTACGACTT), was performed according to the method of Vornhagen and co-workers.49 Briefly, DNA was extracted and prepared using a Qiagen (Valencia, CA) DNA preparation system according to the manufacturer's instructions. Single-step PCR was conducted to generate barcoded amplicons with linkers and the size and concentration of DNA fragments were determined by using DNA chips within an Experion Automated Electrophoresis Station (Bio-Rad Laboratories, Hercules, CA) and a TBS-380 Fluorometer (Promega Corporation, Madison, WI). A sample of 9.6 × 106 double-stranded DNA molecules/μL with an average size of 625 bp was mixed with 9.6 million DNA capture beads and subsequently amplified by emulsion PCR. Subsequently, bead-attached DNA samples were denatured with NaOH and sequencing primers were annealed. A two-region 454 sequencing run was performed on a 70 × 75 GS PicoTiterPlate using a Genome Sequencer FLX System (Roche, Nutley, NJ). All failed sequence reads, low-quality sequence ends (average phred quality score for sequence ends was required to be >Q25), short reads <150 bp (final mean length 412 bp) and tags and primers were removed. Sequence collections were then depleted of any non-bacterial sequences, sequences with ambiguous base calls, sequences with homopolymers >5 bp in length and chimeras.50–52 To determine the predicted identity of bacteria, remaining sequences were de-noised and de-replicated and OTU clustering was performed (uClust, http://www.drive5.com) and then queried using BLASTn against a highly curated custom database of 16S bacterial sequences curated from the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/). BLASTn outputs were compiled and data reduction analysis was performed as described previously.53–55 Bacteria were classified at the closest well-characterized genus.
Statistical analyses
The significance of differences between untreated and treated biofilms was examined using two-tailed Student's t-test modified for unequal variance. Values of P < 0.05 were considered significant and values of P < 0.01 were considered highly significant.
Results
Microscopic characterization of microfluidic dental plaque biofilm
Microscopic examination was performed using an epifluorescence microscope following the staining of the biofilm cells with Syto9 stain (a component of the Live/Dead stain). This enabled clear visualization and differentiation of a variety of cell types and cellular arrangements on outer layers of the biofilm and within small cell clusters/microcolonies on the glass surface of the microfluidic channel. Cell types included cocci that were typically arranged in chains (Figure 2a) and rod-shaped cells that were often autoaggregated (self-aggregated), and the rods were occasionally observed to have formed rosette-like structures. Both were coaggregated with other cell types (Figure 2b–d). Rod-shaped cells were seldom observed on their own. Fusiform cell types, suspected to be Fusobacterium nucleatum or a closely related species, were also observed both as single cells and in multi-cell-type corn-cob-like coaggregates (Figure 2c and d). Single cells were also detected. In addition, occasional yeast-like cells were observed, although these were often surrounded or encased in bacterial cells and difficult to image. No flagellated/motile cells or spirochaetes were observed to be present, although these may have been exclusively located in the dense biofilm masses and therefore difficult to visualize.
Figure 2.
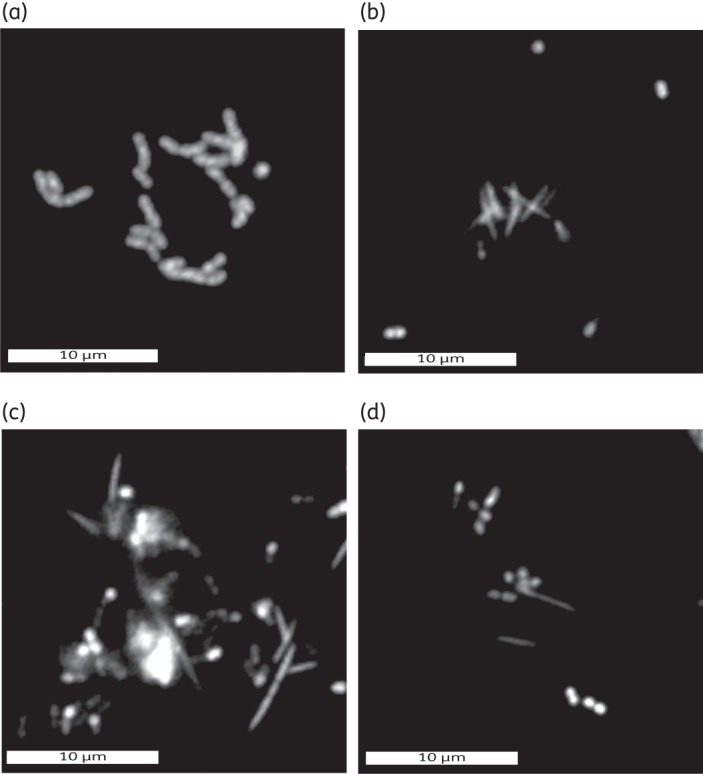
Epifluorescence images of observed cell shapes and arrangements demonstrating the multi-species composition of the biofilm in the microfluidic system. (a) Chains of cocci. (b) Aggregated rod-shaped cells with outlying diplococci. (c) Fusiforms co-localized with mixtures of other cell types (including diplococci and streptococci). (d) Simple multi-species coaggregated clusters of cells. Bars represent 10 μm.
Culture-independent characterization of microfluidic dental plaque biofilm
After data curation, an average of 2848 sequences were analysed for each sample. Rarefaction analysis was conducted using QIIME.56 Rarefaction analysis was performed on the three biofilm channels from which the biofilm cells were harvested to determine how much of the microbiome was evaluated. It should be noted that visual inspection of the channels after biofilm harvesting did reveal that sparsely attached cells/microcolonies remained, but repeated attempts to harvest these cells were not successful. Biofilm community richness for each of the three channels was estimated using the Chao1 estimator57 averaged over 10 iterations and normalized at 2790 sequences for each of the three channels. This predicted 230, 224 and 220 OTUs for each. Rarefaction analysis of the sequencing data identified 158, 145 and 141 OTUs (Figure S1, available as Supplementary data at JAC Online). Thus, the percentages of each population evaluated were predicted to be 68.7%, 64.7% and 64.1%, respectively.
The phyla identified by bTEFAP in the three channels were similar to those documented to be present within oral biofilms. Variation was observed in the relative amounts of each phylum within each channel, but the relative order of the dominant phyla was constant between each channel. The two dominant phyla were always Proteobacteria and Firmicutes (average percentage abundance of 59% and 37.7%, respectively) (Figure 3a). Less dominant phyla included Bacteroidetes (2.4%), Actinobacteria (0.8%), Fusobacteria (0.06%) and TM7 (0.03%) and were always in that order of abundance in each channel.
Figure 3.

Average percentage abundance of each phylum (a) and genus (b) based on bacterial tag-encoded FLX amplicon pyrosequencing of dental plaque biofilms grown in three randomly harvested BioFlux microfluidic channels.
In-depth analysis of the sequences of the identified phyla demonstrated that the biofilms contained genera common to the oral cavity and that the relative abundance of each was similar to that found in supragingival plaque. Across three experiments (channels) there was an average of 14 genera. Neisseria and streptococci were the most abundant (average percentage abundance 55.8% and 20.2% respectively), while other more metabolically fastidious oral species, such as Porphyromonas and Fusobacterium (average percentage abundance 2.4% and 0.06%, respectively), were also always present in the dental plaque biofilms (Figure 3b and Figure S2, available as Supplementary data at JAC Online). Of note, none of the sequences belonging to the genus Porphyromonas could be speciated because identities were to database sequences for which species designations have not been assigned. As our database is up to date with the NCBI and RDP taxonomy database, these sequences are likely from novel species of Porphyromonas. Members of the candidate division TM7 were also detected, although at low levels (<0.05%). Further analysis of the dominant speciated members belonging to the genera Neisseria and Streptococcus were Neisseria subflava (28.2%), Neisseria mucosa (1.7%), Streptococcus infantis (4.1%) and Streptococcus oralis (3.0%) (Table S1, available as Supplementary data at JAC Online).
Relationship between CPC concentration and changes in cell damage/inactivation
Biofilms that developed within the channels of 48-well (24 channels) BioFlux microfluidic plates displayed increased cell damage/death that was proportional, in a sigmoidal fashion, to increased CPC test solution concentration (Figure 4). Inferred from pixel colour (red versus green) and intensity (0–255 shades), the amount of dead or damaged cells within the untreated biofilms washed with PBS was 21%. Conversely, washing with 70% ethanol resulted in near maximal and highly significant cell damage/inactivation (P < 0.01%, 84.4%) (Figure 4). Over the entire CPC test solution concentration range (0.5%–0.001% w/v), a concentration-dependent sigmoidal-like kill curve was observed, with standard deviations greatest over the linear range (0.05%–0.005%). There was a highly significant level of cell damage/inactivation (P < 0.01%) when the PBS control was compared with treatments with 0.5%–0.01% CPC (Figure 4). Furthermore, there was no significant difference (P < 0.05%) in total biofilm cell damage/inactivation between those treated with 70% ethanol and those treated, within this range, between 0.5% and 0.05% CPC.
Figure 4.

Average percentage signal from biofilms accounted for by ‘live’/viable signal (light grey bars) and ‘dead’/damaged signal (dark grey bars) in relation to total signal captured for both. A sigmoidal relationship was observed between CPC concentration and cell viability, with greatest replicate variability for CPC concentrations falling within the linear range. Significant differences from the PBS control are highlighted (*P < 0.05, **P < 0.01).
Computational 3D reconstruction and rendering of image Z-stacks of the treated and untreated biofilms (Figure 5) allowed inspection of the penetrance of CPC within the biofilms. As inferred by cell damage/death and consequent labelling with the red fluorescent probe of the Baclight LIVE/DEAD kit, penetration was also found to be proportional to CPC concentration. Almost complete penetration and cell damage/death was observed for the 0.5% CPC-treated biofilm, almost no penetration/killing was observed for the 0.001% CPC-treated biofilm, and killing/penetrance increasing from the 0.005% to 0.01% CPC-treated biofilms (Figure 4). Close inspection of the 0.01% and 0.005% 3D-rendered Z-stacks showed that cell death was often restricted to the perimeter of the biofilms. This suggests that reaction–diffusion limitation was a factor in cell damage/inactivation, as is removal of CPC from the bulk fluid to a concentration below the threshold required for cell damage.
Figure 5.

Representative biofilm renderings following each treatment. Green signal indicates viable live cells (Syto 9), red signal indicates damaged/dead cells (propidium iodide). Upper images (A1–H1) are of the x–y plane. Middle images (A2–H2) are of the x–z plane. Lower images (A3–H3) are an angled view of each plane (x–y–z). PBS is the negative control, 70% EtOH (ethanol) is the positive control, and all CPC treatments shown in decreasing concentration from left to right. Both controls indicate that the assay functioned correctly and decreasing biofilm damage corresponds to decreasing CPC concentration. Bars represent 30 μm.
Relationship between CPC concentration and changes in biofilm architecture
Inspection of 3D-rendered images indicated that the treatment of biofilms with different concentrations of CPC altered biofilm structure (Figure 5). Specifically, biofilms treated with 0.5%, 0.1% and 0.05% CPC possessed a more densely packed, flatter-looking biofilm that lacked pronounced undulating masses. In order to assess and quantify any such changes, Comstat was used to determine the biomass, thickness distribution and roughness coefficient of the biofilms (Table 1). Regardless of treatment concentration, no significant change in biomass was observed, even though clear differences in biofilm architecture were observed (Figure 5a versus Figure 5c–e). The average thickness of the biofilms, however, reflected the visual observations, and significant differences (P < 0.05) were observed for biofilms treated with 0.5%, 0.1% and 0.05% CPC. In this range, biofilms were reduced in thickness by 34.5%–43.0%. In addition, it was in this concentration range that the vast majority of the cells were dead/damaged. Biofilm roughness was seemingly little changed by CPC treatments, with the exception of that following exposure to 0.01% CPC, which yielded a P value of 0.038. Collectively, the quantitative data support the qualitative 3D imaging data and show that the microfluidic dental plaque biofilms exposed to 0.5%–0.05% CPC undergo architectural change, but no significant loss of biofilm biomass was evident.
Table 1.
Effect of treatments with different CPC concentrations on average biofilm biomass, average thickness and biofilm roughness
| Biofilm parameter | PBS | 70% EtOH | 0.5% CPC | 0.1% CPC | 0.05% CPC | 0.01% CPC | 0.005% CPC | 0.001% CPC |
|---|---|---|---|---|---|---|---|---|
| Biomass (μm3/μm2) | 7.52 (4.44) | 8.07 (2.47) | 7.00 (4.60) | 9.84 (4.67) | 6.84 (3.86) | 8.82 (7.35) | 13.28 (2.32) | 10.41 (8.56) |
| Thickness (μm) | 25.96 (4.86) | 20.30 (5.47)* | 14.81 (8.41)* | 17.01 (5.44)* | 14.89 (5.94)** | 19.05 (11.15) | 26.59 (10.58) | 24.78 (10.93) |
| Roughness | 0.41 (0.33) | 0.37 (0.16) | 0.69 (0.48) | 0.58 (0.22) | 0.64 (0.20) | 0.73 (0.30)* | 0.55 (0.35) | 0.55 (0.30) |
Data [mean (SD)] were derived from imaging of at least three separate biofilm channels (experiments).
*P < 0.05 and **P < 0.01: significant differences from the PBS control.
EtOH, ethanol.
Discussion
The work presented here demonstrates the potential of using a high-throughput approach to reproducibly grow oral multi-species biofilms that contain species that are indigenous to dental plaque. In addition, the utility of such a device for evaluating antimicrobial/anti-biofilm compounds was demonstrated by exposing the biofilm communities to different concentrations of CPC and evaluating cell damage and death through a combination of 3D imaging and image analysis software packages. Of significance to this work, few in vitro, high-throughput oral model biofilm systems have been developed that contain multi-species biofilm communities grown under conditions representative of the oral cavity.12,25,27,58–60 The development of such representative in vitro model biofilms is important to accurately predict the in vivo efficacy of current or candidate antimicrobials that may be used in oral hygiene products.
In order to characterize the species composition of the microfluidic dental plaque biofilms and validate use of the model system for future biofilm studies, microscopic examination of biofilms and bTEFAP were performed. Both microscopic and bTEFAP analyses demonstrated that there were numerous cell types/species in microfluidic-grown biofilms that are common to dental plaque. These not only formed biofilms but were often juxtaposed to form the cellular arrangements commonly observed in dental plaque. For example, multi-generic aggregates (possibly coaggregates) containing streptococci, and different cell types that are commonly observed in dental plaque were often observed (Figure 2).15,61 The ability of bacteria to aggregate via autoaggregative interactions (self-aggregation) and coaggregation (the specific recognition and adhesion of different species of bacteria to one another) is proposed to be integral to biofilm development.62,63 While the different cell types are an indication of the multi-species nature of the biofilms, a more descriptive study of the bacterial diversity (and reproducibility between channels) was desirable. Thus, we performed shallow (an average of 2848 reads) bTEFAP on three randomly chosen biofilms grown in three channels to identify the dominant phyla and constituent genera present in the biofilms. Even though a relatively shallow approach was taken, and we estimate that we captured ∼65.8% of the total diversity, the predominant populations were covered and the well represented types of species found in the biofilms were consistent with those found in human dental plaque biofilms. An unexpected outcome of bTEFAP was that Neisseria was shown to be a reproducibly dominant member of biofilms, as well as Streptococcus in second place. Neisseria constitutes a large genus (>25 species) belonging to the β-Proteobacteria, which are facultative anaerobes.64 Evidence suggests that that Neisseria is present in young dental plaque biofilms65,66 and can be one of a number of dominant members in supragingival plaque44 and on dental plaque implants.67 It should be noted that the amount of Neisseria in the microfluidic system was at least 2-fold greater than often observed in normal dental plaque biofilm communities. However, work by Bik et al.68 has shown that Neisseria can strongly dominate in healthy dental plaque harvested from healthy volunteers. Streptococci are often considered ‘primary’ or ‘pioneer’ colonizers15 because of their ability to adhere to saliva-coated surfaces,69 adhere to one another,70 use salivary components such as mucins as a nutrient source71,72 and survive under a range of environmental conditions (pH, oxygen tension, etc).73–75 Considering our approach to removing biofilm from within the microfluidic system (which did leave some cell clusters still attached), it is possible that the amount of streptococci, as determined by bTEFAP, is an underestimate because they are pioneer colonizers and many would be at the base of the biofilms. It should also be noted that obligate anaerobic species were also present, including Porphyromonas and Veillonella spp. (Figure 3 and Table S1, available as Supplementary data at JAC Online). These two genera are often detected in young dental plaque biofilms, although, as observed in the microfluidic system, at an abundance that is approximately an order of magnitude lower than streptococci.76 Another interesting observation was the absence of detectable amounts of Actinomyces, which may be due to selection pressures, such as fluid shear, exerted by the microfluidic system. For example, it has been shown that Actinomyces are less capable of residing in biofilms under high shear, as opposed to streptococci, especially during early stages of biofilm development.77 Our bTEFAP protocol is predicted to be able to detect Actinomyces spp. and, using Actinomyces-selective media,78 we were able to culture bacteria that were Gram-positive and possessed Actinomyces-like filamentous rod-shaped morphologies from the saliva inoculum. Thus, taking these findings together, the developed biofilms grown in the microfluidic system represent a community that is similar to supragingival plaque biofilms, albeit with seemingly elevated numbers of Neisseria spp. and below detectable amounts of Actinomyces spp.—possibly a result of environmental conditions (oxygen tension, shear, etc.) in the microfluidic system and/or the approach of harvesting biofilm cells. An interesting follow-up study would be to grow biofilms for longer periods or to grow them under anaerobic conditions.
A required utility for the microfluidic biofilm system was that it would be amenable to testing antimicrobials. In order to test our system we used CPC, a cationic quaternary ammonium compound used as the key antimicrobial ingredient in many over-the-counter mouthwashes. As positive and negative controls we used 70% ethanol and PBS for all experimental runs. It should be noted that, in the absence of antimicrobial treatment, the average amount of damaged/inactivated cells was 21%, suggesting that the biofilm contains some dead and damaged cells prior to antimicrobial treatment. Such an observation has been reported previously for in vitro single-species biofilms26 and in vivo multi-species biofilms.79 Not only is CPC an antimicrobial (due to its ability to integrate into bacterial membranes) but it is also a bio-surfactant and may have de-adhesive effects on biofilm bacteria.80–82 With respect to cell damage/inactivation, a clear concentration-dependent response effect was observed (Figures 4 and 5). CPC treatment caused significant cell inactivation/damage, as compared with the PBS-treated biofilms, between the concentrations of 0.5% and 0.01% (P < 0.05, Figure 4). In addition, the efficacy of CPC was not significantly different (P < 0.05) from that of 70% ethanol at a concentration range between 0.5% and 0.05. At the lower concentration ranges, discrete regions of cell damage/inactivation (as indicated by Live/Dead stain) of biofilms were observed, where outer cells were more damaged than their deeper-positioned counterparts (Figure 4, 0.001%–0.01% CPC). While the degree of CPC-induced damage/inactivation to biofilm cells varies among studies in the literature,25,26,60 at least one supports our observation that cells in the outer extremities of biofilms tend to have a greater propensity to be damaged by CPC than deeper-lying cells.26 This phenomenon likely relates to the cationic bacterial membrane-reactive nature of CPC83 and the retardation of access of CPC to the biofilm depths. This retardation process is often described as reaction–diffusion limitation.4,84 The subject of de-adhesion in response to CPC (or other quaternary ammonium compounds), has also previously received attention,45,59,80,81 although no tests have been performed on saliva-grown multi-species biofilms. Here we qualitatively (3D-rendered CLSM images; Figure 1) and quantitatively (Comstat data, Table 1) show that CPC, at a concentration between 0.5 and 0.05%, significantly alters biofilm architecture, presumably due to de-adhesion events (Figure 5; Table 1). This was displayed as a general compacting of the biofilm and loss of biofilm projections. There was no significant decrease in biomass, however, suggesting that either de-adhesion may only be transitory and damaged cells re-integrate into the biofilm or that CPC weakens biofilm architecture and causes biofilm collapse. This latter possibility is particularly intriguing as work by Hope and Wilson85 has shown that a similar phenomenon occurs in biofilms exposed to chlorhexidine. Chlorhexidine is similar to CPC in its mechanism of action against bacterial membranes and it is also a cationic antimicrobial and thus likely interacts with biofilm EPS and may cause contraction or collapse of the biofilm structure.83
In conclusion, the work presented here demonstrates the utility of a microfluidic system in the development of oral biofilms under environmentally germane conditions to yield communities with cell morphotypes, cellular arrangements and species abundances that are similar to those found in dental plaque. Furthermore, the extent of bacterial damage/inactivation in biofilms exposed to different concentrations of CPC was demonstrated to be qualitatively and quantitatively measurable. From a real-world perspective, the effective concentrations of CPC required to damage dental plaque biofilm bacteria are in line with those currently used in oral hygiene products (0.01%–0.1%). While the focus of this work has been on saliva and dental plaque biofilms, arguably the greatest implication of this work is in the translational potential for the combined BioFlux microfluidic/CLSM system to be used in other biofilm studies using nutrient and/or inoculum sources that are difficult to obtain, such as urine, wound exudate, tears and sweat.
Funding
This work was supported by the National Institutes of Health (NIH: R21DE018820 to A. H. R.) and University of Michigan start-up funds to A. H. R.
Transparency declarations
A. H. R has received research awards from the National Institutes of Health (NIDCR), Colgate-Palmolive (Piscataway, NJ) and the Society for Applied Microbiology to fund research studies in his lab over the past five years. S. E. D. is CEO and the General Partner of Molecular Research LP (MR DNA) where pyrosequencing and associated analyses were conducted. S. E. D. has no other conflicts or declarations associated with this work. J. D. is a Regional Application Specialist for W. Nuhsbaum Inc. J. B. is the Global Director, BioFlux Sales & Support at Fluxion Biosciences (although, he does not hold a seat on board of directors of the corporation and holds no equity position in the company). All other authors: none to declare.
Supplementary data
Acknowledgements
The authors are grateful for the support provided by the Microbial Ecology: Relationships to Human and Environmental Health group at the University of Michigan, USA.
References
- 1.Costerton JW, Cheng KJ, Geesey GG, et al. Bacterial biofilms in nature and disease. Annu Rev Microbiol. 1987;41:435–64. doi: 10.1146/annurev.mi.41.100187.002251. [DOI] [PubMed] [Google Scholar]
- 2.Stoodley P, Sauer K, Davies DG, et al. Biofilms as complex differentiated communities. Annu Rev Microbiol. 2002;56:187–209. doi: 10.1146/annurev.micro.56.012302.160705. [DOI] [PubMed] [Google Scholar]
- 3.Armitage GC. Basic features of biofilms–why are they difficult therapeutic targets? Ann R Australas Coll Dent Surg. 2004;17:30–4. [PubMed] [Google Scholar]
- 4.Gilbert P, Maira-Litran T, McBain AJ, et al. The physiology and collective recalcitrance of microbial biofilm communities. Adv Microb Physiol. 2002;46:202–56. [PubMed] [Google Scholar]
- 5.Mah TF, O'Toole GA. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001;9:34–9. doi: 10.1016/s0966-842x(00)01913-2. [DOI] [PubMed] [Google Scholar]
- 6.Roberts AP, Mullany P. Oral biofilms: a reservoir of transferable, bacterial, antimicrobial resistance. Expert Rev Anti Infect Ther. 2010;8:1441–50. doi: 10.1586/eri.10.106. [DOI] [PubMed] [Google Scholar]
- 7.Stewart PS, Rayner J, Roe F, et al. Biofilm penetration and disinfection efficacy of alkaline hypochlorite and chlorosulfamates. J Appl Microbiol. 2001;91:525–32. doi: 10.1046/j.1365-2672.2001.01413.x. [DOI] [PubMed] [Google Scholar]
- 8.Stewart PS. Theoretical aspects of antibiotic diffusion into microbial biofilms. Antimicrob Agents Chemother. 1996;40:2517–22. doi: 10.1128/aac.40.11.2517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Peters BM, Jabra-Rizk MA, O'May GA, et al. Polymicrobial interactions: impact on pathogenesis and human disease. Clin Microbiol Rev. 2012;25:193–213. doi: 10.1128/CMR.00013-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Leriche V, Briandet R, Carpentier B. Ecology of mixed biofilms subjected daily to a chlorinated alkaline solution: spatial distribution of bacterial species suggests a protective effect of one species to another. Environ Microbiol. 2003;5:64–71. doi: 10.1046/j.1462-2920.2003.00394.x. [DOI] [PubMed] [Google Scholar]
- 11.Umland TC, Schultz LW, MacDonald U, et al. In vivo-validated essential genes identified in Acinetobacter baumannii by using human ascites overlap poorly with essential genes detected on laboratory media. MBio. 2012;3:e00113-12. doi: 10.1128/mBio.00113-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Foster JS, Kolenbrander PE. Development of a multispecies oral bacterial community in a saliva-conditioned flow cell. Appl Environ Microbiol. 2004;70:4340–8. doi: 10.1128/AEM.70.7.4340-4348.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Whittaker CJ, Klier CM, Kolenbrander PE. Mechanisms of adhesion by oral bacteria. Annu Rev Microbiol. 1996;50:513–52. doi: 10.1146/annurev.micro.50.1.513. [DOI] [PubMed] [Google Scholar]
- 14.Kolenbrander PE, Andersen RN, Blehert DS, et al. Communication among oral bacteria. Microbiol Mol Biol Rev. 2002;66:486–505. doi: 10.1128/MMBR.66.3.486-505.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kolenbrander PE, Palmer RJ, Jr, Rickard AH, et al. Bacterial interactions and successions during plaque development. Periodontol 2000. 2006;42:47–79. doi: 10.1111/j.1600-0757.2006.00187.x. [DOI] [PubMed] [Google Scholar]
- 16.Aas JA, Paster BJ, Stokes LN, et al. Defining the normal bacterial flora of the oral cavity. J Clin Microbiol. 2005;43:5721–32. doi: 10.1128/JCM.43.11.5721-5732.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Marsh PD. Dental plaque as a biofilm and a microbial community - implications for health and disease. BMC Oral Health. 2006;6(Suppl 1):S14. doi: 10.1186/1472-6831-6-S1-S14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bernimoulin JP. Recent concepts in plaque formation. J Clin Periodontol. 2003;30(Suppl 5):7–9. doi: 10.1034/j.1600-051x.30.s5.3.x. [DOI] [PubMed] [Google Scholar]
- 19.White DJ. Dental calculus: recent insights into occurrence, formation, prevention, removal and oral health effects of supragingival and subgingival deposits. Eur J Oral Sci. 1997;105:508–22. doi: 10.1111/j.1600-0722.1997.tb00238.x. [DOI] [PubMed] [Google Scholar]
- 20.Whitmore SE, Lamont RJ. The pathogenic persona of community-associated oral streptococci. Mol Microbiol. 2011;81:305–14. doi: 10.1111/j.1365-2958.2011.07707.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lamont RJ, Jenkinson HF. Subgingival colonization by Porphyromonas gingivalis. Oral Microbiol Immunol. 2000;15:341–9. doi: 10.1034/j.1399-302x.2000.150601.x. [DOI] [PubMed] [Google Scholar]
- 22.Marsh PD. Controlling the oral biofilm with antimicrobials. J Dent. 2010;38(Suppl 1):S11–5. doi: 10.1016/S0300-5712(10)70005-1. [DOI] [PubMed] [Google Scholar]
- 23.Davies RM. Toothpaste in the control of plaque/gingivitis and periodontitis. Periodontol 2000. 2008;48:23–30. doi: 10.1111/j.1600-0757.2008.00261.x. [DOI] [PubMed] [Google Scholar]
- 24.Teles RP, Teles FR. Antimicrobial agents used in the control of periodontal biofilms: effective adjuncts to mechanical plaque control. Braz Oral Res. 2009;23(Suppl 1):39–48. doi: 10.1590/s1806-83242009000500007. [DOI] [PubMed] [Google Scholar]
- 25.Pan PC, Harper S, Ricci-Nittel D, et al. In-vitro evidence for efficacy of antimicrobial mouthrinses. J Dent. 2010;38(Suppl 1):S16–20. doi: 10.1016/S0300-5712(10)70006-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Foster JS, Pan PC, Kolenbrander PE. Effects of antimicrobial agents on oral biofilms in a saliva-conditioned flowcell. Biofilms. 2004;1:5–12. [Google Scholar]
- 27.Shapiro S, Giertsen E, Guggenheim B. An in vitro oral biofilm model for comparing the efficacy of antimicrobial mouthrinses. Caries Res. 2002;36:93–100. doi: 10.1159/000057866. [DOI] [PubMed] [Google Scholar]
- 28.Wilson M. Susceptibility of oral bacterial biofilms to antimicrobial agents. J Med Microbiol. 1996;44:79–87. doi: 10.1099/00222615-44-2-79. [DOI] [PubMed] [Google Scholar]
- 29.Wilson M, Patel H, Fletcher J. Susceptibility of biofilms of Streptococcus sanguis to chlorhexidine gluconate and cetylpyridinium chloride. Oral Microbiol Immunol. 1996;11:188–92. doi: 10.1111/j.1399-302x.1996.tb00356.x. [DOI] [PubMed] [Google Scholar]
- 30.McBain AJ. Chapter 4: In vitro biofilm models: an overview. Adv Appl Microbiol. 2009;69:99–132. doi: 10.1016/S0065-2164(09)69004-3. [DOI] [PubMed] [Google Scholar]
- 31.Bercy P, Lasserre J. Susceptibility to various oral antiseptics of Porphyromonas gingivalis W83 within a biofilm. Adv Ther. 2007;24:1181–91. doi: 10.1007/BF02877764. [DOI] [PubMed] [Google Scholar]
- 32.Wilson M, Patel H, Noar JH. Effect of chlorhexidine on multi-species biofilms. Curr Microbiol. 1998;36:13–8. doi: 10.1007/s002849900272. [DOI] [PubMed] [Google Scholar]
- 33.Cuadra-Saenz G, Rao DL, Underwood AJ, et al. Autoinducer-2 influences interactions amongst pioneer colonizing streptococci in oral biofilms. Microbiology. 2012;158:1783–95. doi: 10.1099/mic.0.057182-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Benoit MR, Conant CG, Ionescu-Zanetti C, et al. New device for high-throughput viability screening of flow biofilms. Appl Environ Microbiol. 2010;76:4136–42. doi: 10.1128/AEM.03065-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wong L, Sissons C. A comparison of human dental plaque microcosm biofilms grown in an undefined medium and a chemically defined artificial saliva. Arch Oral Biol. 2001;46:477–86. doi: 10.1016/s0003-9969(01)00016-4. [DOI] [PubMed] [Google Scholar]
- 36.Sissons CH. Artificial dental plaque biofilm model systems. Adv Dent Res. 1997;11:110–26. doi: 10.1177/08959374970110010201. [DOI] [PubMed] [Google Scholar]
- 37.Pratten J, Wills K, Barnett P, et al. In vitro studies of the effect of antiseptic-containing mouthwashes on the formation and viability of Streptococcus sanguis biofilms. J Appl Microbiol. 1998;84:1149–55. doi: 10.1046/j.1365-2672.1998.00462.x. [DOI] [PubMed] [Google Scholar]
- 38.Bjorklund M, Ouwehand AC, Forssten SD. Improved artificial saliva for studying the cariogenic effect of carbohydrates. Curr Microbiol. 2011;63:46–9. doi: 10.1007/s00284-011-9937-x. [DOI] [PubMed] [Google Scholar]
- 39.Ledder RG, Gilbert P, Pluen A, et al. Individual microflora beget unique oral microcosms. J Appl Microbiol. 2006;100:1123–31. doi: 10.1111/j.1365-2672.2006.02847.x. [DOI] [PubMed] [Google Scholar]
- 40.McBain AJ, Sissons C, Ledder RG, et al. Development and characterization of a simple perfused oral microcosm. J Appl Microbiol. 2005;98:624–34. doi: 10.1111/j.1365-2672.2004.02483.x. [DOI] [PubMed] [Google Scholar]
- 41.Pratten J, Wilson M. Antimicrobial susceptibility and composition of microcosm dental plaques supplemented with sucrose. Antimicrob Agents Chemother. 1999;43:1595–9. doi: 10.1128/aac.43.7.1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Amerongen AV, Veerman EC. Saliva–the defender of the oral cavity. Oral Dis. 2002;8:12–22. doi: 10.1034/j.1601-0825.2002.1o816.x. [DOI] [PubMed] [Google Scholar]
- 43.Orstavik D. Bacterial adherence related to agglutination and surface film formation. Adv Exp Med Biol. 1978;107:869–75. [PubMed] [Google Scholar]
- 44.Yamanaka W, Takeshita T, Shibata Y, et al. Compositional stability of a salivary bacterial population against supragingival microbiota shift following periodontal therapy. PLoS One. 2012;7:e42806. doi: 10.1371/journal.pone.0042806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rao D, Arvanitidou E, Du-Thumm L, et al. Efficacy of an alcohol-free CPC-containing mouthwash against oral multispecies biofilms. J Clin Dent. 2011;22:187–94. [PubMed] [Google Scholar]
- 46.De Jong MH, Van der Hoeven JS, Van Os JH. Growth of micro-organisms from supragingival dental plaque on saliva agar. J Dent Res. 1986;65:85–8. doi: 10.1177/00220345860650021601. [DOI] [PubMed] [Google Scholar]
- 47.Heydorn A, Nielsen AT, Hentzer M, et al. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology. 2000;146:2395–407. doi: 10.1099/00221287-146-10-2395. [DOI] [PubMed] [Google Scholar]
- 48.Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012;9:671–5. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Vornhagen J, Stevens M, McCormick DW, et al. Coaggregation occurs amongst bacteria within and between biofilms in domestic showerheads. Biofouling. 2013;29:53–68. doi: 10.1080/08927014.2012.744395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ishak HD, Plowes R, Sen R, et al. Bacterial diversity in Solenopsis invicta and Solenopsis geminata ant colonies characterized by 16S amplicon 454 pyrosequencing. Microb Ecol. 2011;61:821–31. doi: 10.1007/s00248-010-9793-4. [DOI] [PubMed] [Google Scholar]
- 51.Capone KA, Dowd SE, Stamatas GN, et al. Diversity of the human skin microbiome early in life. J Invest Dermatol. 2011;131:2026–32. doi: 10.1038/jid.2011.168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bailey MT, Dowd SE, Parry NM, et al. Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium. Infect Immun. 2010;78:1509–19. doi: 10.1128/IAI.00862-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Handl S, Dowd SE, Garcia-Mazcorro JF, et al. Massive parallel 16S rRNA gene pyrosequencing reveals highly diverse fecal bacterial and fungal communities in healthy dogs and cats. FEMS Microbiol Ecol. 2011;76:301–10. doi: 10.1111/j.1574-6941.2011.01058.x. [DOI] [PubMed] [Google Scholar]
- 54.Callaway TR, Dowd SE, Edrington TS, et al. Evaluation of bacterial diversity in the rumen and feces of cattle fed different levels of dried distiller's grains plus solubles using bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP) J Anim Sci. 2010;88:3977–83. doi: 10.2527/jas.2010-2900. [DOI] [PubMed] [Google Scholar]
- 55.Pitta DW, Pinchak E, Dowd SE, et al. Rumen bacterial diversity dynamics associated with changing from bermudagrass hay to grazed winter wheat diets. Microb Ecol. 2010;59:511–22. doi: 10.1007/s00248-009-9609-6. [DOI] [PubMed] [Google Scholar]
- 56.Caporaso JG, Kuczynski J, Stombaugh J, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7:335–6. doi: 10.1038/nmeth.f.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Chao A. Nonparametric estimation of the number of classes in a population. Scand J Stat. 1984;11:265–70. [Google Scholar]
- 58.Rudney JD, Chen R, Lenton P, et al. A reproducible oral microcosm biofilm model for testing dental materials. J Appl Microbiol. 2012;113:1540–53. doi: 10.1111/j.1365-2672.2012.05439.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Busscher HJ, White DJ, Atema-Smit J, et al. Surfactive and antibacterial activity of cetylpyridinium chloride formulations in vitro and in vivo. J Clin Periodontol. 2008;35:547–54. doi: 10.1111/j.1600-051X.2008.01230.x. [DOI] [PubMed] [Google Scholar]
- 60.Corbin A, Pitts B, Parker A, et al. Antimicrobial penetration and efficacy in an in-vitro oral biofilm model. Antimicrob Agents Chemother. 2011;55:338–44. doi: 10.1128/AAC.00206-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Listgarten MA. The structure of dental plaque. Periodontol 2000. 1994;5:52–65. doi: 10.1111/j.1600-0757.1994.tb00018.x. [DOI] [PubMed] [Google Scholar]
- 62.Rickard AH, Gilbert P, High NJ, et al. Bacterial coaggregation: an integral process in the development of multi-species biofilms. Trends Microbiol. 2003;11:94–100. doi: 10.1016/s0966-842x(02)00034-3. [DOI] [PubMed] [Google Scholar]
- 63.Kolenbrander PE. Oral microbial communities: biofilms, interactions, and genetic systems. Annu Rev Microbiol. 2000;54:413–37. doi: 10.1146/annurev.micro.54.1.413. [DOI] [PubMed] [Google Scholar]
- 64.Short HB, Clark VL, Kellogg DS, Jr, et al. Anaerobic survival of clinical isolates and laboratory strains of Neisseria gonorrhoea: use in transfer and storage. J Clin Microbiol. 1982;15:915–9. doi: 10.1128/jcm.15.5.915-919.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ritz HL. Fluorescent antibody staining of Neisseria, Streptococcus and Veillonella in frozen sections of human dental plaque. Arch Oral Biol. 1969;14:1073–83. doi: 10.1016/0003-9969(69)90077-6. [DOI] [PubMed] [Google Scholar]
- 66.Ritz HL. Microbial population shifts in developing human dental plaque. Arch Oral Biol. 1967;12:1561–8. doi: 10.1016/0003-9969(67)90190-2. [DOI] [PubMed] [Google Scholar]
- 67.Komoria R, Sato T, Takano-Yamamotoa T, et al. Microbial composition of dental plaque microflora on first molars with orthodontic bands and brackets, and the acidogenic potential of these bacteria. J Oral Biosci. 2012;54:107–12. [Google Scholar]
- 68.Bik EM, Long CD, Armitage GC, et al. Bacterial diversity in the oral cavity of 10 healthy individuals. ISME J. 2010;4:962–74. doi: 10.1038/ismej.2010.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gibbons RJ, Houte JV. Bacterial adherence in oral microbial ecology. Annu Rev Microbiol. 1975;29:19–44. doi: 10.1146/annurev.mi.29.100175.000315. [DOI] [PubMed] [Google Scholar]
- 70.Kolenbrander PE, London J. Adhere today, here tomorrow: oral bacterial adherence. J Bacteriol. 1993;175:3247–52. doi: 10.1128/jb.175.11.3247-3252.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Van der Hoeven JS, Camp PJ. Synergistic degradation of mucin by Streptococcus oralis and Streptococcus sanguis in mixed chemostat cultures. J Dent Res. 1991;70:1041–4. doi: 10.1177/00220345910700070401. [DOI] [PubMed] [Google Scholar]
- 72.van der Hoeven JS, van den Kieboom CW, Camp PJ. Utilization of mucin by oral Streptococcus species. Antonie Van Leeuwenhoek. 1990;57:165–72. doi: 10.1007/BF00403951. [DOI] [PubMed] [Google Scholar]
- 73.Burne RA. Oral streptococci … products of their environment. J Dent Res. 1998;77:445–52. doi: 10.1177/00220345980770030301. [DOI] [PubMed] [Google Scholar]
- 74.Burne RA, Quivey RG, Jr, Marquis RE. Physiologic homeostasis and stress responses in oral biofilms. Methods Enzymol. 1999;310:441–60. doi: 10.1016/s0076-6879(99)10035-1. [DOI] [PubMed] [Google Scholar]
- 75.Marquis RE. Oxygen metabolism, oxidative stress and acid-base physiology of dental plaque biofilms. J Ind Microbiol. 1995;15:198–207. doi: 10.1007/BF01569826. [DOI] [PubMed] [Google Scholar]
- 76.Diaz PI, Chalmers NI, Rickard AH, et al. Molecular characterization of subject-specific oral microflora during initial colonization of enamel. Appl Environ Microbiol. 2006;72:2837–48. doi: 10.1128/AEM.72.4.2837-2848.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Sharma PK, Gibcus MJ, van der Mei HC, et al. Influence of fluid shear and microbubbles on bacterial detachment from a surface. Appl Environ Microbiol. 2005;71:3668–73. doi: 10.1128/AEM.71.7.3668-3673.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Lewis R, McKenzie D, Bagg J, et al. Experience with a novel selective medium for isolation of Actinomyces spp. from medical and dental specimens. J Clin Microbiol. 1995;33:1613–6. doi: 10.1128/jcm.33.6.1613-1616.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Auschill TM, Arweiler NB, Netuschil L, et al. Spatial distribution of vital and dead microorganisms in dental biofilms. Arch Oral Biol. 2001;46:471–6. doi: 10.1016/s0003-9969(00)00136-9. [DOI] [PubMed] [Google Scholar]
- 80.Neu TR. Significance of bacterial surface-active compounds in interaction of bacteria with interfaces. Microbiol Rev. 1996;60:151–66. doi: 10.1128/mr.60.1.151-166.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Smith RN, Andersen RN, Kolenbrander PE. Inhibition of intergeneric coaggregation among oral bacteria by cetylpyridinium chloride, chlorhexidine digluconate and octenidine dihydrochloride. J Periodontal Res. 1991;26:422–8. doi: 10.1111/j.1600-0765.1991.tb01732.x. [DOI] [PubMed] [Google Scholar]
- 82.Marsh PD. Microbiological aspects of the chemical control of plaque and gingivitis. J Dent Res. 1992;71:1431–8. doi: 10.1177/00220345920710071501. [DOI] [PubMed] [Google Scholar]
- 83.Gilbert P, Moore LE. Cationic antiseptics: diversity of action under a common epithet. J Appl Microbiol. 2005;99:703–15. doi: 10.1111/j.1365-2672.2005.02664.x. [DOI] [PubMed] [Google Scholar]
- 84.Stewart PS, Grab L, Diemer JA. Analysis of biocide transport limitation in an artificial biofilm system. J Appl Microbiol. 1998;85:495–500. doi: 10.1046/j.1365-2672.1998.853529.x. [DOI] [PubMed] [Google Scholar]
- 85.Hope CK, Wilson M. Analysis of the effects of chlorhexidine on oral biofilm vitality and structure based on viability profiling and an indicator of membrane integrity. Antimicrob Agents Chemother. 2004;48:1461–8. doi: 10.1128/AAC.48.5.1461-1468.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.