

Abstract
There is now considerable experimental data to suggest that inflammatory cells collaborate in the healing of skeletal fractures. In terms of mechanisms that contribute to the recruitment of inflammatory cells to the fracture site, chemokines and their receptors have received considerable attention. Our previous findings have shown that Duffy antigen receptor for chemokines (Darc), the non-classical chemokine receptor that does not signal, but rather acts as a scavenger of chemokines that regulate cell migration, is a negative regulator of peak bone density in mice. Furthermore, because Darc is expressed by inflammatory and endothelial cells, we hypothesized that disruption of Darc action will affect post-fracture inflammation and consequently will affect fracture healing. To test this hypothesis, we evaluated fracture healing in mice with targeted disruption of Darc and corresponding wild type (WT) control mice. We found that fracture callus cartilage formation was significantly greater (33%) at 7 days post-surgery in Darc-KO compared to WT mice. The increased cartilage was associated with greater Collagen (Col) II expression at 3 days post-fracture and Col-X at 7 days post-fracture compared to WT mice, suggesting that Darc deficiency led to early fracture cartilage formation and differentiation. We then compared the expression of cytokine and chemokine genes known to be induced during inflammation. Interleukin (Il)-1β, Il-6, and monocyte chemotactic protein 1 were all down regulated in the fractures derived from Darc-KO mice at one day post-fracture, consistent with an altered inflammatory response. Furthermore, the number of macrophages was significantly reduced around the fractures in Darc-KO compared to WT mice. Based on these data, we concluded that Darc plays a role in modulating the early inflammatory response to bone fracture and subsequent cartilage formation. However, the early cartilage formation was not translated with an early bone formation at the fracture site in Darc-KO compared to WT mice.
Introduction
Fracture healing is a complex process that involves the interaction of soluble mediators, extracellular matrix components, resident cells and infiltrating leukocyte subtypes, which participate differentially in the classically defined three phases of fracture healing: inflammation, bone formation and bone remodeling [1], [2]. In the inflammatory phase, a fibrin mesh develops and acts as a scaffold for the infiltrating neutrophils and macrophages to clear the tissue debris. A cytokine/chemokine cascade initiates the proliferation of fibrous cells in the fracture gap that differentiate to cartilage and cancellous bone, and thereafter remodel into lamellar bone through the coordinated action of bone forming osteoblast cells and bone resorbing osteoclast cells [3], [4], [5], [6]. The inflammatory phase that initiates this process is critical for successful bone repair, and its elucidation will identify new approaches to enhance bone repair in normal and impaired conditions.
The role of some of the chemokine receptors has been investigated in fracture healing [7]. However, the role of the duffy antigen receptor for chemokines (Darc), which is known to play an important role in chemokine-regulated leukocyte/neutrophil trafficking during inflammation has not been characterized in fracture healing. We have previously identified Darc as a negative regulator of bone mineral density [8]. Darc is known to bind chemokines that regulate cell trafficking [9]. It is highly expressed in erythrocytes as well as vascular endothelial cells [10], [11], the cell types that play key role in wound healing process [12], [13], [14]. Based on the established role of inflammation in fracture healing, and the predicted role of Darc in regulating function of inflammatory chemokines, we proposed that Darc expression plays an important role in post-fracture inflammation and fracture healing. To test this hypothesis, we have used Darc-KO mice and wild type mice to compare femur fracture healing and the post-fracture expression levels of inflammatory genes in the two lines of mice.
Materials and Methods
Femur Fracture Model
All procedures were performed with the approval of the institutional Animal Care and Use Committee (IACUC) in accordance with the Animal Welfare Act at the Jerry L. Pettis Memorial VA Medical Center, Loma Linda, CA, USA. Dr. A. Chaudhuri (New York Blood Center, NY, USA) provided the Darc-knockout (Darc-KO) mice, as well as the control wild type (WT) mice. Mice were generated as previously described by Luo et al. [15], and were bred and maintained as previously described [8].
Ten to twelve week old mice were placed under isoflurane anesthesia and femoral fractures were produced by the three-point bending technique, as previously described [16], [17]. Briefly, the femur was internally stabilized with a 0.5 mm diameter stainless steel pin surgically implanted prior to fracture, and a 4-0 PDSII suture was used to close the wound. Buprenorphine was administered subcutaneously after fracture for pain relief. Animals were allowed unrestricted movement during post-fracture healing. Tissues were harvested for analysis at different intervals of post-fracture healing.
Histomorphometry
Histomorphometry analyses were performed at 7, 11 and 21 days post-fracture. Bones were fixed in 10% formalin, demineralized in EDTA, paraffin-embedded and each sample was sectioned longitudinally at 5–6 µm thickness as described by Bancroft [18]. Sections were stained with Safranin-Orange and Fast Green to assess cartilage area and total callus area. Data are reported as the mean of callus area and cartilage area, and excluded the native cortical bone and the intramedullary space. Quantification was performed using Image Pro software 6.3 (Media Cybernetics, Silver Spring, MD, USA).
X-ray analysis
Fracture repair was analyzed by X-ray examination using a micro-computed tomography scanner (μ-CT; VivaCt 40 scanner, SCANCO Medical AG, Brüttisellen, Switzerland). Analyses were performed on fractured and unfractured bones at 21 days post-fracture, when healing of the fractured bone in the wild-type mouse normally approaches bony union of the fracture callus. Cortical bone and the surrounding mineralized tissue of the fracture callus were manually contoured. Total tissue volume was defined as the circumference of the bony callus. To normalize the length of bone examined, the analysis was performed on a standard length around the fracture, encompassing 591±5 slices of the scanned image and centered at the fracture site. We used two thresholds to differentiate between lower density callus cancellous bone (220–570 mg HA/cm3) and native cortical bone (570–1000 mg HA/cm3). To normalize for trabecular bone contributions to the fracture callus measurements, the unfractured bone trabecular values were subtracted from the fracture callus data.
Gene Expression Analysis
Animals were sacrificed at 1, 3, 7 and 15 days post-surgery. Total RNA was isolated from fracture tissues as follows: Approximately 2 mm from each side of the fracture was quickly harvested and the fracture callus including bone marrow was stored in liquid nitrogen. The callus was then pulverized with Trizol under cryogenic conditions and following the protocol provided by Invitrogen. We have found this approach to be very successful in maintaining RNA integrity during purification. Reverse transcription was performed with MMLV Reverse Transcriptase (Promega, San Luis Obispo, CA, USA). Real-time PCR was performed using the SYBRgreen master mix (Applied Biosystems, Foster City, CA, USA) with gene-specific primers (Integrated DNA Technologies, Coralville, IA, USA). The information about the sequence of the primers used in this study is presented in Table S1. Changes in gene expression were determined by subtracting the Ct (threshold cycle) of target gene from the Ct value of the housekeeping gene; peptidylprolyl isomerase A (Ppia) (ΔCt = Ct of target gene – Ct of Ppia) as described in Table S2. Mean ΔCt was then used to calculate the difference in cycle thresholds between the wild-type (WT) unfractured bones and WT or Darc-KO fractures (ΔΔCt = mean ΔCt of WT unfractured bones - mean ΔCt of fractured bones from each genotype). The fold-activation was calculated as 2−ΔΔCt [19]. The genes examined were as follows: Receptor activator of nuclear factor-κB ligand (Rankl), osteoprotegerin (Opg), tumor necrosis factor (Tnf)-α, interleukin (Il)-1beta (β), Il-6, macrophage inflammatory protein-1 alpha (Mip-1α/Ccl3), monocyte chemotactic protein-1 (Mcp-1/Ccl2), as well as collagen II (Col-II) and collagen X (Col-X).
Detection and quantification of inflammatory cells in fracture tissues
Immunohistochemistry was performed using rat anti-mouse Ly-6B.2, F4/80 (ABD Serotec, A Divison of MorphoSys, Raleigh, NC, USA), and CD45R (BD Pharmingen, San Jose, CA, USA) to identify neutrophils, macrophages, and B lymphocytes, respectively. The longitudinal sections prepared as described above were incubated with primary antibodies for 60 min at 4°C. Conjugate and substrate were used according to manufacturer instructions and following the protocol provided by BIOCARE Medical (Biocare Medical, Concord, CA, USA). The number of positive cells in the fracture tissues was determined using OsteoMeasure software (Osteometrics Inc. GA, USA). Values were expressed as number of cells per mm2 tissue.
Statistical analysis
Statistical significance was evaluated using two-tailed Student's t-test. The difference is considered statistically significant, when p<0.05.
Results
Lack of Darc expression enhanced post-fracture cartilage formation
To determine if lack of Darc expression affects fracture healing process, we performed histomorphometric analysis of the fracture callus cartilage in Darc-KO and WT mice at 7, 11 and 21 days post-fracture ( Fig. 1 ). We found that fracture callus size ( Fig. 2A ) and fracture cartilage area ( Fig. 2B ) were significantly greater at 7 days post-surgery in Darc-KO mice compared to WT mice (45% and 33%, respectively, p<0.03). However, at 11 and 21 days post-fracture, the callus size was not significantly different between the two lines of mice. To evaluate fracture cartilage development, we compared the expression of Col-II and Col-X, markers of pre-hypertrophic and hypertrophic chondrocytes, respectively. We found the expression levels of both Col-II and Col-X were greater in both strains of mice at 7 days, compared to 3 and 15 days post-surgery, as would be expected in normal fracture repair ( Fig. 3 ). Darc-KO mice exhibited greater expression of Col-II at 3 days post-fracture ( Fig. 3A ) and Col-X at 7 days post-fracture ( Fig. 3B ) compared to WT mice. At 15 days post-surgery, the expression of both genes was significantly down-regulated in fracture calluses of both lines of mice compared to 7 days post-fractures, as is expected during the late stages of fracture cartilage development.
Figure 1. Callus and cartilage development during fracture healing in WT and Darc-KO mice.

Cartilage was stained with Safranin-Orange in fracture calluses derived from WT (A, C, E) and Darc-KO mice (B, D, F) at 7 (A, B), 11 (C, D) and 21 days post-fracture (E, F). Scale bar = 1 mm.
Figure 2. Quantification of callus and cartilage development during fracture healing.

Histomorphometric quantification of cartilage area (A) and fracture callus areas (B) derived from WT and Darc-KO mice at 7, 11 and 21 days post-fracture. We examined 5–9 animals/time point/strain of mice. *p<0.05 WT vs Darc-KO mice.
Figure 3. Expression of cartilage marker genes during fracture healing.
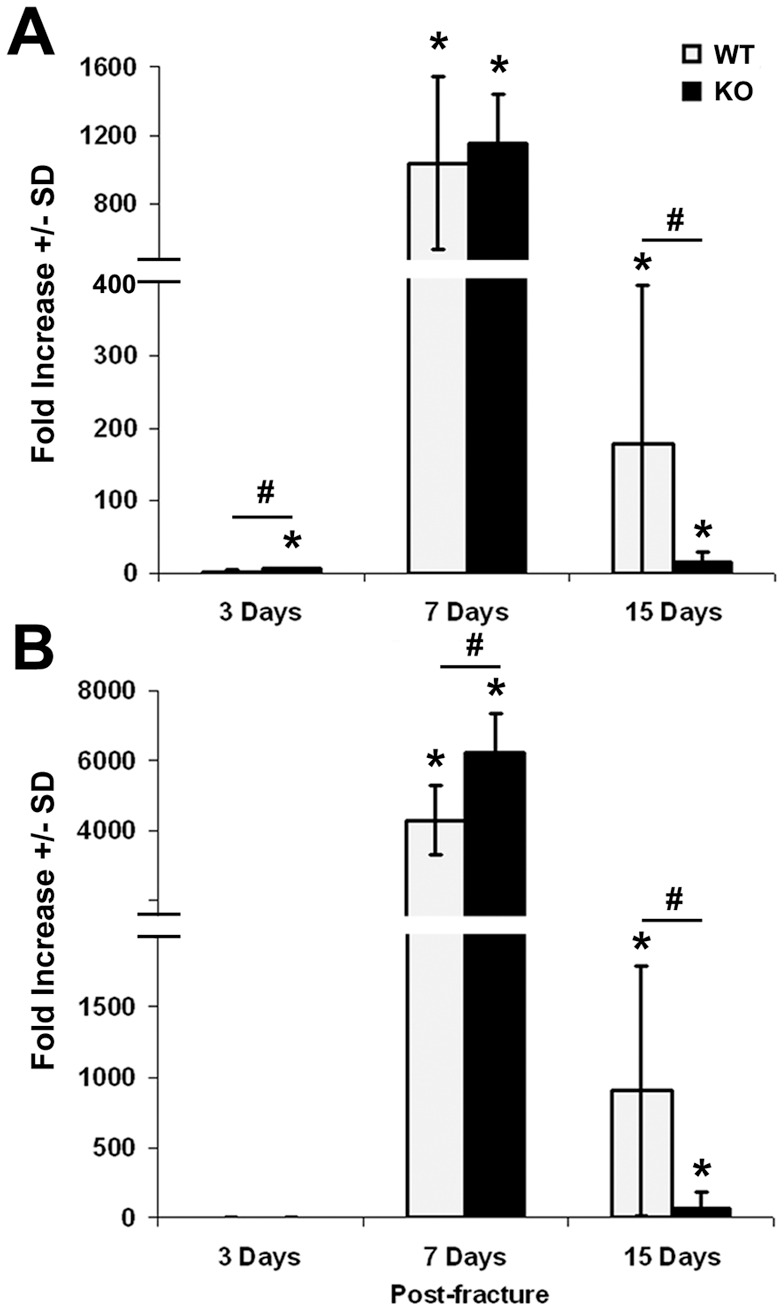
Collagen II (A) and Collagen X (B) expression in fracture calluses derived from WT and Darc-KO mice at 3, 7 and 15 days post-fracture. Data are expressed as fold change in the expression of the gene in the fractured bones compared to unfractured bone of WT mice. We analyzed 3–4 animals/mouse strain at 3 days post-surgery and 5–8 animals/mouse strain at 7 and 15 days post-fracture.*p<0.05 vs WT unfractured bones, #p<0.05 between fractured bones of the two lines of mice.
Darc deficiency did not improve fracture healing - Micro-CT data at 21 days post-fracture
To determine if the early cartilage formation in Darc-KO mice changed callus bone formation during fracture healing, we analyzed the mineralized tissues of the hard callus by micro-CT at 21 days post-fracture. Surprisingly, no significant difference in total volume or bone volume of the fracture calluses was observed between Darc-KO mice and WT mice at 21 days post-fracture ( Fig. 4 ). To determine if osteoclastogenesis was affected by lack of Darc expression; we compared the mRNA expression of Rankl and Opg, the decoy receptor at the fracture calluses between the two lines of mice ( Fig. 5 ). Both genes were up regulated at 7 days post-fracture when compared to WT unfractured bones, but no significant difference was observed between the two lines of mice. At 15 days post-fracture, mRNA expression of both genes was down-regulated in fracture calluses from both lines of mice compared to 7 days post-fracture, but no difference was observed between the two lines of mice at this time point.
Figure 4. Micro-CT analysis of the fracture calluses.

Data are presented as the percentage of WT. Five fractures were examined from each strain of mice. BV, bone volume; TV, total volume; BV/TV, bone volume fraction.
Figure 5. Expression of Rankl and Opg genes at 7 and 15 days post-fracture.
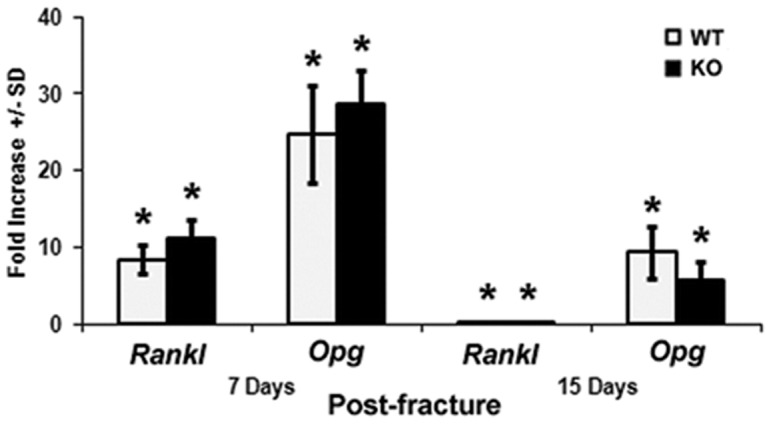
Data are expressed as fold-change in the expression of the gene in the fractured bones compared to unfractured bones of WT mice. We analyzed 6–8 animals/mouse strain. *p<0.05 vs WT unfractured bones.
Darc expression regulates post-fracture inflammation
The pro-inflammatory cytokines; TNF-α, IL-1β and IL-6 have been shown not only to coordinate the hematopoietic and immune systems, but also to contribute to bone repair by regulating osteoclastogenesis and the early recruitment and differentiation of osteoblastic lineage cells [20], [21], [22], [23], [24], [25], [26], [27], [28], [29]. Therefore, we have analyzed the effect of targeted disruption of Darc on the expression of these three inflammatory cytokines in bone fracture. As expected, the mRNA level of the three cytokines was enhanced after one day of bone fracture in both lines of mice ( Fig. 6 ), but the magnitude of increase in the expression of IL-1β and IL-6 was reduced by 52–54% in the fractures derived from Darc-KO mice compared to the fractures derived from WT mice at one day post-fracture. The expression levels of the three cytokines in unfractured bones were not different between the two lines of mice (data not shown).
Figure 6. Expression of cytokine genes at four time points post-fracture.
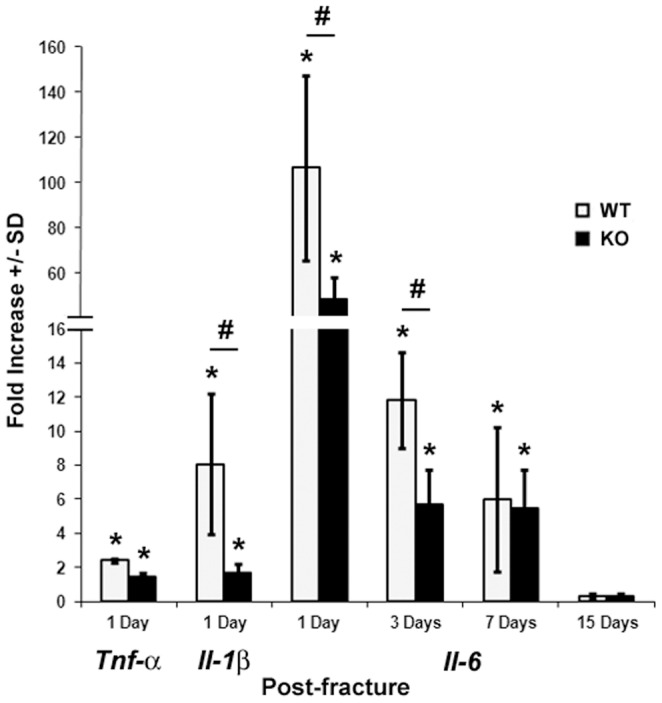
Data are expressed as fold-change in the expression of the gene in the fractured bones compared to unfractured bones of WT mice. We analyzed 3–4 animals/mouse strain at 1 and 3 days and 5–8 mice at 7 and 15 days post-fracture.*p<0.05 vs WT unfractured bones, #p<0.05 between fractured bones of the two lines of mice.
The post-fracture inflammatory response is marked by the infiltration of cells that release inflammatory mediators such as cytokines and chemokines. Thus, to determine if the migration of inflammatory cells was affected by the lack of Darc expression in KO mice, we evaluated the expression of two CC chemokines, monocyte chemotactic protein 1 (Mcp-1), also called Ccl2 and macrophage inflammatory protein 1 (Mip-1α), Ccl3. The expression level of both genes was increased at one day post-fracture, but the magnitude of increase in the expression was reduced by three days post-fracture ( Fig. 7 ), when inflammation starts to subside. The expression of Ccl2 but not Ccl3 was reduced in Darc-KO compared to WT mice at one day post-fracture ( Fig. 7A, B ).
Figure 7. Expression of chemokine genes at four time points post-fracture.
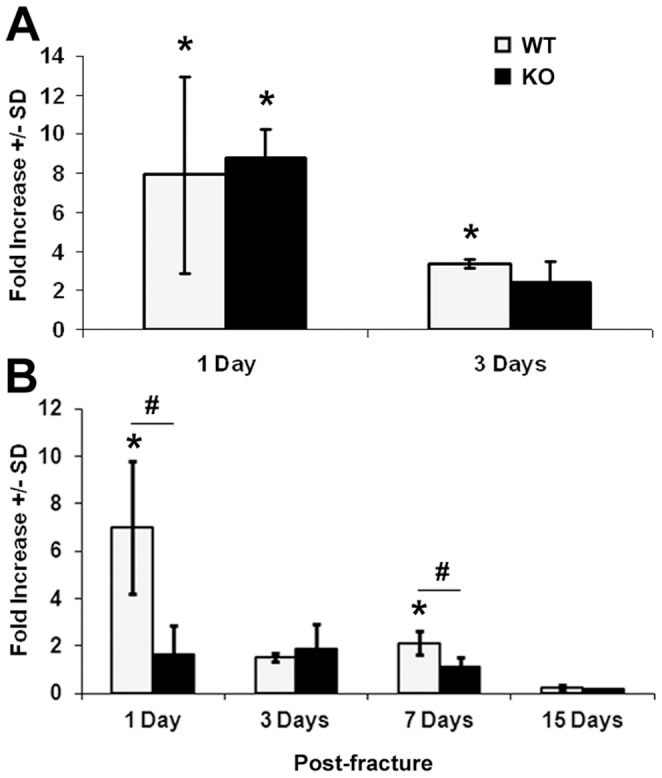
A. Ccl3, B. Ccl2. Data are expressed as fold-change in the expression of the gene in the fractured bones compared to WT unfractured bones. We examined 3–4 animals/time point/mouse strain at 1 and 3 days and 5–8 mice at 7 and 15 days post-fracture. *p<0.05 vs WT unfractured bone, #p<0.05 between fractured bones of the two lines of mice.
Since Il-6 and Ccl2 are the genes that showed the biggest difference in mRNA expression both between fractured and unfractured bones and between the two lines of mice after fracture, we have evaluated the expression of Il-6 ( Fig. 6 ) and Ccl2 ( Fig. 7B ) at additional post-fracture time points. While the increase in the expression of Il-6 in response to fracture was greater in WT compared to KO mice at 1 and 3 days post-fracture, no difference was observed at 7 days post-fracture between the two lines of mice when inflammation normally has subsided. Though mRNA expression of Ccl2 in the fracture calluses decreased at 7 days, it remained significantly greater in fractured bones compared to unfractured bones in WT mice. Furthermore, Ccl2 expression in fracture calluses derived from WT mice was greater at 1 and 7 days post-fracture compared to fractures derived from KO mice.
To determine if the expression of cytokines and chemokines is associated with infiltration of inflammatory cells to fractures, we quantified the inflammatory cell population in the bone marrow and soft tissues around the fracture sites ( Fig. 8 ). At one day post-fracture, neutrophils were the most abundant and B lymphocytes were the least abundant at the fracture site (data not shown). While the expression levels of markers of neutrophils (Ly-6B.2), B-lymphocytes (CD45R) and macrophages (F4/80) were reduced at the fracture site of Darc-KO mice compared to WT mice, the reduction was statistically significant only for the macrophage marker, F4/80 (p = 0.04, Fig. 8 ).
Figure 8. Evaluation of the inflammatory cells around the fractures in Darc-KO and WT mice at one day post-fracture.

Data are expressed as number of cells/mm2 of fracture callus. We examined 5–7 animals/mouse strain. The inflammatory cells were identified by antibodies specific for CD45 (B-lymphocytes), F4/80 (macrophages) and Ly6.b.2 (neutrophils). *p<0.05 WT vs Darc-KO mice.
Discussion
Unlike typical chemokine receptors, Darc is present on both red blood and endothelial cells but not on leukocytes [30], [31], [32]. Previous studies have shown that Darc is crucial for chemokine-mediated leukocyte migration in vivo, for the changes in chemokine and chemokine receptor homeostasis associated with Darc gene deficiency, and it exerts strong anti-inflammatory effects [33]. Fracture repair is a local event regulated by locally expressed inflammatory mediators. The onset of acute inflammation initiates the early stages of fracture repair and its resolution promotes cartilage formation immediately thereafter, so it would stand to reason that Darc regulates this process. However, the involvement of Darc in post-fracture inflammation and fracture repair has never been investigated. Therefore, in this study we tested the effect of targeted disruption of Darc expression on femoral bone fracture repair and post-fracture inflammation.
We have evaluated local inflammation at one day post-fracture, when the predominant cells in the hematoma are neutrophils [34]. Recruited neutrophils are thought to mediate this switch by producing several macrophage chemoattractants, such as CCL2 and IL-6 [35], [36]. Thus, since lack of Darc expression was previously reported to reduce the infiltration of neutrophils to local inflammation site [37], we expected fractures derived from Darc-KO mice to show less chemokine and cytokine expression/secretion in fracture tissues, and consequently less macrophage infiltration to the fracture calluses.
Darc-KO mice showed a significant increase in cartilage abundance in the fracture calluses at 7 days post-fracture, which was associated with an earlier induction of Collagen-II gene expression and Collagen-X expression in fracture repair in the Darc-KO mouse compared to WT strain ( Fig. 3 ). These data suggest that fracture callus cartilage has developed earlier in Darc-KO than in WT mice, though there is no evidence that Darc directly regulates collagen gene expression. It has been reported that inflammation alters chondrocyte proliferation [38] as is observed with persistent inflammation in conditions, such as infection where cartilage development can be impaired. Inflammatory mediators such as IL-1β have a pivotal role in sustaining both inflammation and cartilage erosion, at least in the pathology associated with arthritis [39], [40], [41], [42]. It is known that chemokines can induce cartilage degradation [43], [44], [45], so it is possible that lack of Darc expression in the KO mice affected fracture cartilage development through reduced inflammatory cell recruitment and chemokine expression that favored cartilage formation in Darc-KO fracture calluses.
The increased cartilage development that occurred very early in fracture repair and well before maximum bone remodeling in Darc-KO mice compared to WT mice possibly was associated with reduced post-fracture inflammation in Darc-KO fractures. However, the early cartilage formation in Darc-KO mice did not translate into accelerated conversion to bone and early bony union, since micro-CT measurements of the hard callus did not reveal any significant differences in the volume of the newly formed bone between the two lines of mice at 21 days post-surgery ( Fig. 4 ). Our previous studies demonstrated reduced resorbing surfaces in unfractured bones derived from Darc-KO mice compared to WT mice [8], which would be expected to reduce osteoclast recruitment and subsequently delay cartilage resorption in Darc-KO fracture calluses compared to WT mice. If cartilage resorption was delayed in Darc-KO fractures, its effects on the development of the bony fracture callus were not significant, and we conclude from the cartilage abundance and collagen gene expression differences that the increased fracture cartilage was the result of differences in chondrocyte development. These results suggest that Darc deficiency reduces the inflammation that would normally delay cartilage development and Collagen-II gene expression until inflammation is resolved, normally after the first three days post-fracture.
To investigate inflammatory cell infiltration to the fracture tissues, we compared the numbers of inflammatory cells around the fractures in the two lines of mice using antibodies specific for markers of neutrophils, macrophages and B lymphocytes. We quantified the cell numbers at one day post-fracture, because we had observed a tremendous decrease in the expression of the inflammatory cytokines and chemokines in the Darc-KO fractures at one day post-fracture ( Fig. 6 and 7 ). An evaluation of these three cell types revealed a trend toward reduced numbers of these cells in the fracture calluses from Darc-KO, although only macrophage numbers were significantly reduced in Darc-KO fractures compared to WT fractures ( Fig. 8 ). We believe that the reason why we did not reach significance between the two lines of mice for neutrophils was that the size of the groups was insufficient to detect significance with the modest differences in cartilage formation and chondrocyte-specific collagen gene expression between the two lines of mice (Figures 2 and 3, respectively). As expected, neutrophils were the most abundant cells in the fracture tissues at one day post-fracture, as these cells are known to migrate to the injured areas immediately after injury, and as the first inflammatory cells to reach sites of tissue injury, they secrete cytokines and chemokines that play crucial roles in attracting other cells to the injured tissue. They are followed by macrophages that act as phagocytes to clear the wound of matrix and cell debris [46]. Each of these inflammatory cells secrete growth factors and cytokines that are presumed to function as tissue repair signals and direct resident cell functions in fracture callus development during healing. It follows that if the number of neutrophils is reduced in Darc-KO fracture calluses; the recruitment of the other inflammatory cells to the fracture will be reduced and will affect healing. Therefore, we speculate that down-regulation of cytokine expression level due to delayed recruitment of inflammatory cells to the fracture callus in Darc-KO mice could be responsible for the early cartilage formation in KO mice fractures.
Though the expression of the Opg and Rankl markers of osteoclast development was significantly greater in fracture calluses compared to unfractured bones in both lines of mice, no difference was observed between the two lines of mice at 7 days post-fracture ( Fig. 5 ), suggesting that local osteoclastogenesis was not affected in Darc-KO fracture calluses. Since our previous studies have established that bone resorption is reduced in the unfractured bones of the Darc-KO mice [8], we propose that cartilage resorption at the fracture site of Darc-KO mice could be delayed due to reduced recruitment of osteoclast precursors to the fracture calluses in KO mice compared to WT mice. However, without further studies on osteoclast development and recruitment during fracture healing we cannot conclude that fracture callus remodeling was reduced in Darc-KO mice.
Most of the previous studies on Darc regulation of inflammation were based upon LPS challenges, chemokine injections or chronic diseases [37], [47], [48], [49]. Dawson et al., [47] reported that there was no difference between the WT and Darc-KO mice in LPS-induced leukocyte recruitment to the peritoneal cavity which was the injection site at two hours post-LPS injection, but they noticed more leukocytes accumulation in the lung and liver in the KO mice compared to WT mice. Lee et al., [37] have found that mice that lack Darc in erythrocytes showed reduced neutrophils in lung airspaces compared to WT control mice 4 hours after intratracheal instillation of LPS. However, the MIP-2 concentration was found to be higher in the lung airspaces of mice that lack Darc expression in both erythrocytes and endothelial cells and from mice that lack Darc expression only in endothelial cells but not in erythrocytes at 4 hours (h) but not 2 h post LPS injection compared to control mice. In lung tissue vascular compartment, the concentration of MIP-2 was greater only in mice lacking Darc in erythrocytes at 2 h but not 4 h post-LPS injection compared to WT control mice. However, in the plasma, MIP-2 concentration was reduced in Darc-KO mice lacking Darc in endothelial (4 h) and in erythrocyte cells (2 h) compared to WT control mice. In addition, Fukuma et al., [48] have shown that plasma concentrations of eotaxin (Ccl11) and MCP-1 (Ccl2), the two chemokines that bind to Darc, were significantly lower in Darc-KO compared to WT mice in physiological conditions. The clearance from the blood circulation after intravenous injection of these two chemokines was faster in Darc-KO compared to WT mice. These data suggest that the lack of Darc expression enhances the clearance of chemokines from blood circulation, and that chemokine-Darc regulated secretion of soluble chemokines is time- and tissue- dependent, two variables that were different in the healing fracture calluses of our study.
Other studies on fracture repair have established that the acute inflammation that normally initiates fracture repair is a local phenomenon [50] and that a systemic inflammatory response to traumatic injuries impairs fracture healing [51]. We therefore expect the inflammatory response to bone fracture to be local, and that systemic measurements of cytokines in bone repair would be misleading. Our evaluation of post-fracture inflammation was performed at the site of the fracture not in any other organ; we did not evaluate the circulating chemokines or leukocytes after fracture. Because we limited our evaluation to the local fracture tissues and did not examine systemic inflammation, we expect that Darc regulation of fracture inflammation would differ from the studies on the systemic effect of Darc expression.
Results from human studies suggest that Darc expression might function in human fracture repair as it did in the mouse fracture model. It has been demonstrated that rs2814778, a Darc-deficient haplotype known to protect the African-American population from malaria infection, was related to low white blood cell and neutrophil counts [52]. However, to our knowledge, only one study compared fracture healing between different ethnicities [53], and they did not find any differences in fracture healing, although they noticed small differences in pain scores during fracture healing. Further investigation with larger patient populations is required to characterize the effect of Darc deficiency on fracture healing in human populations.
Based on the previous studies and the present study, we hypothesize that Darc delays the clearance of chemokines from the blood circulation and induces the inflammatory cell migration to the injured areas. In the absence of Darc, chemokines are rapidly cleared from the blood circulation and are probably retained in the lung and liver which delays the recruitment of inflammatory cells to the injured areas. Taken together, our findings suggest that Darc expression promotes the resolution of inflammation in fracture repair. Furthermore, our previous studies have established that bone resorption is reduced in the unfractured bones of the Darc-KO mice [8].These data suggest that the delayed cartilage resorption at the fracture site of Darc-KO mice could also be due to reduced recruitment of osteoclast precursors to the fracture calluses in KO mice compared to WT mice.
Conclusions
We have shown for the first time, using mice with targeted disruption of the mouse Darc gene that Darc plays a role in modulating the regulation of inflammatory response to bone injury and that lack of Darc expression promotes cartilage formation in fracture calluses but does not affect bony union or fracture healing at 21 days post-fracture.
Supporting Information
Information on the primers used for quantitative PCR.
(DOC)
Real Time PCR. Raw Data. Real Time PCR was done using cDNA prepared from RNA isolated from fracture calluses derived from Darc-KO and wild type mice at three time points post-fracture.
(XLS)
Acknowledgments
All work was performed at the facilities provided by the Veterans Administration in Loma Linda, CA, USA. The authors thank Jomana Bashir and Yan Hu for their technical assistance.
Funding Statement
The project described was supported by USAMRMC ORTHOPAEDIC RESEARCH PROGRAM (http://mrmc-rad6.army.mil/). Grant/Contract/MIPR No.: W81XWH-10-1-0952. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Cruess RL, Dumont J (1975) Fracture healing. Can J Surg 18: 403–413. [PubMed] [Google Scholar]
- 2.Simmons DJ (1985) Fracture healing perspectives. Clin Orthop Relat Res: 100–113. [PubMed]
- 3. Cho TJ, Gerstenfeld LC, Einhorn TA (2002) Differential temporal expression of members of the transforming growth factor beta superfamily during murine fracture healing. J Bone Miner Res 17: 513–520. [DOI] [PubMed] [Google Scholar]
- 4. Gerstenfeld LC, Cullinane DM, Barnes GL, Graves DT, Einhorn TA (2003) Fracture healing as a post-natal developmental process: molecular, spatial, and temporal aspects of its regulation. J Cell Biochem 88: 873–884. [DOI] [PubMed] [Google Scholar]
- 5. Lange J, Sapozhnikova A, Lu C, Hu D, Li X, et al. (2010) Action of IL-1beta during fracture healing. J Orthop Res 28: 778–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Mumme M, Scotti C, Papadimitropoulos A, Todorov A, Hoffmann W, et al. (2012) Interleukin-1beta modulates endochondral ossification by human adult bone marrow stromal cells. Eur Cell Mater 24: 224–236. [DOI] [PubMed] [Google Scholar]
- 7. Xing Z, Lu C, Hu D, Yu YY, Wang X, et al. (2010) Multiple roles for CCR2 during fracture healing. Dis Model Mech 3: 451–458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Edderkaoui B, Baylink DJ, Beamer WG, Wergedal JE, Porte R, et al. (2007) Identification of mouse Duffy antigen receptor for chemokines (Darc) as a BMD QTL gene. Genome Res 17: 577–585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Peiper SC, Wang ZX, Neote K, Martin AW, Showell HJ, et al. (1995) The Duffy antigen/receptor for chemokines (DARC) is expressed in endothelial cells of Duffy negative individuals who lack the erythrocyte receptor. J Exp Med 181: 1311–1317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Chaudhuri A, Rodriguez M, Zbrzezna V, Luo H, Pogo AO, et al. (2003) Induction of Duffy gene (FY) in human endothelial cells and in mouse. Cytokine 21: 137–148. [DOI] [PubMed] [Google Scholar]
- 11. Chaudhuri A, Zbrzezna V, Johnson C, Nichols M, Rubinstein P, et al. (1989) Purification and characterization of an erythrocyte membrane protein complex carrying Duffy blood group antigenicity. Possible receptor for Plasmodium vivax and Plasmodium knowlesi malaria parasite. J Biol Chem 264: 13770–13774. [PubMed] [Google Scholar]
- 12. Bao P, Kodra A, Tomic-Canic M, Golinko MS, Ehrlich HP, et al. (2009) The role of vascular endothelial growth factor in wound healing. J Surg Res 153: 347–358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Harrison SL, Vavken P, Murray MM (2011) Erythrocytes inhibit ligament fibroblast proliferation in a collagen scaffold. J Orthop Res 29: 1361–1366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. McGrath MH, Emery JM 3rd (1985) The effect of inhibition of angiogenesis in granulation tissue on wound healing and the fibroblast. Ann Plast Surg 15: 105–122. [DOI] [PubMed] [Google Scholar]
- 15. Luo H, Chaudhuri A, Zbrzezna V, He Y, Pogo AO (2000) Deletion of the murine Duffy gene (Dfy) reveals that the Duffy receptor is functionally redundant. Mol Cell Biol 20: 3097–3101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Bonnarens F, Einhorn TA (1984) Production of a standard closed fracture in laboratory animal bone. J Orthop Res 2: 97–101. [DOI] [PubMed] [Google Scholar]
- 17. Rundle CH, Wang X, Wergedal JE, Mohan S, Lau KH (2008) Fracture healing in mice deficient in plasminogen activator inhibitor-1. Calcif Tissue Int 83: 276–284. [DOI] [PubMed] [Google Scholar]
- 18.Bancroft JD (2002) Enzyme histochemistry and its diagnostic applications; Bancroft JD GMe, editor. Harcourt: Churchill Livingstone Elsevier.
- 19. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402–408. [DOI] [PubMed] [Google Scholar]
- 20. Einhorn TA, Majeska RJ, Rush EB, Levine PM, Horowitz MC (1995) The expression of cytokine activity by fracture callus. J Bone Miner Res 10: 1272–1281. [DOI] [PubMed] [Google Scholar]
- 21. Frost A, Jonsson KB, Nilsson O, Ljunggren O (1997) Inflammatory cytokines regulate proliferation of cultured human osteoblasts. Acta Orthop Scand 68: 91–96. [DOI] [PubMed] [Google Scholar]
- 22. Hierl T, Borcsok I, Sommer U, Ziegler R, Kasperk C (1998) Regulation of interleukin-6 expression in human osteoblastic cells in vitro. Exp Clin Endocrinol Diabetes 106: 324–333. [DOI] [PubMed] [Google Scholar]
- 23. Johnson CS, Jerome CP, Brommage R (2000) Unbiased determination of cytokine localization in bone: colocalization of interleukin-6 with osteoblasts in serial sections from monkey vertebrae. Bone 26: 461–467. [DOI] [PubMed] [Google Scholar]
- 24. Kon T, Cho TJ, Aizawa T, Yamazaki M, Nooh N, et al. (2001) Expression of osteoprotegerin, receptor activator of NF-kappaB ligand (osteoprotegerin ligand) and related proinflammatory cytokines during fracture healing. J Bone Miner Res 16: 1004–1014. [DOI] [PubMed] [Google Scholar]
- 25. Linkhart TA, Linkhart SG, MacCharles DC, Long DL, Strong DD (1991) Interleukin-6 messenger RNA expression and interleukin-6 protein secretion in cells isolated from normal human bone: regulation by interleukin-1. J Bone Miner Res 6: 1285–1294. [DOI] [PubMed] [Google Scholar]
- 26. Morioka M, Shimodaira K, Kuwano Y, Fujikawa H, Saito H, et al. (2000) Effect of interleukin-1beta on aromatase activity and cell proliferation in human osteoblast-like cells (HOS). Biochem Biophys Res Commun 268: 60–64. [DOI] [PubMed] [Google Scholar]
- 27. Olmedo ML, Landry PS, Sadasivan KK, Albright JA, Marino AA (2000) Programmed cell death in post-traumatic bone callus. Cell Mol Biol (Noisy-le-grand) 46: 89–97. [PubMed] [Google Scholar]
- 28. Olmedo ML, Landry PS, Sadasivan KK, Albright JA, Meek WD, et al. (1999) Regulation of osteoblast levels during bone healing. J Orthop Trauma 13: 356–362. [DOI] [PubMed] [Google Scholar]
- 29. Rickard DJ, Gowen M, MacDonald BR (1993) Proliferative responses to estradiol, IL-1 alpha and TGF beta by cells expressing alkaline phosphatase in human osteoblast-like cell cultures. Calcif Tissue Int 52: 227–233. [DOI] [PubMed] [Google Scholar]
- 30. Horne K, Woolley IJ (2009) Shedding light on DARC: the role of the Duffy antigen/receptor for chemokines in inflammation, infection and malignancy. Inflamm Res 58: 431–435. [DOI] [PubMed] [Google Scholar]
- 31. Iwamoto S, Li J, Omi T, Ikemoto S, Kajii E (1996) Identification of a novel exon and spliced form of Duffy mRNA that is the predominant transcript in both erythroid and postcapillary venule endothelium. Blood 87: 378–385. [PubMed] [Google Scholar]
- 32. Reutershan J, Harry B, Chang D, Bagby GJ, Ley K (2009) DARC on RBC limits lung injury by balancing compartmental distribution of CXC chemokines. Eur J Immunol 39: 1597–1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Zarbock A, Bishop J, Muller H, Schmolke M, Buschmann K, et al. (2010) Chemokine homeostasis vs. chemokine presentation during severe acute lung injury: the other side of the Duffy antigen receptor for chemokines. Am J Physiol Lung Cell Mol Physiol 298: L462–471. [DOI] [PubMed] [Google Scholar]
- 34. Chung R, Cool JC, Scherer MA, Foster BK, Xian CJ (2006) Roles of neutrophil-mediated inflammatory response in the bony repair of injured growth plate cartilage in young rats. J Leukoc Biol 80: 1272–1280. [DOI] [PubMed] [Google Scholar]
- 35. Hurst SM, Wilkinson TS, McLoughlin RM, Jones S, Horiuchi S, et al. (2001) Il-6 and its soluble receptor orchestrate a temporal switch in the pattern of leukocyte recruitment seen during acute inflammation. Immunity 14: 705–714. [DOI] [PubMed] [Google Scholar]
- 36. Jung YS, Lee HY, Kim SD, Park JS, Kim JK, et al. (2013) Wnt5a stimulates chemotactic migration and chemokine production in human neutrophils. Experimental & Molecular Medicine 45: e27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Lee JS, Wurfel MM, Matute-Bello G, Frevert CW, Rosengart MR, et al. (2006) The Duffy antigen modifies systemic and local tissue chemokine responses following lipopolysaccharide stimulation. J Immunol 177: 8086–8094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Studer RK, Bergman R, Stubbs T, Decker K (2004) Chondrocyte response to growth factors is modulated by p38 mitogen-activated protein kinase inhibition. Arthritis Research & Therapy 6: : R56–R64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Auron PE (1998) The interleukin 1 receptor: ligand interactions and signal transduction. Cytokine Growth Factor Rev 9: 221–237. [DOI] [PubMed] [Google Scholar]
- 40.Goldring MB (1992) Degradation of articular cartilage in culture: regulatory factors; Woessner JJaH, DS, Marcel Dekker, NY, editor. New York: Marcel Dekker, Inc. 556 p. [Google Scholar]
- 41. van den Berg WB, Joosten LA, Kollias G, van De Loo FA (1999) Role of tumour necrosis factor alpha in experimental arthritis: separate activity of interleukin 1beta in chronicity and cartilage destruction. Ann Rheum Dis 58 Suppl 1I40–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Westacott CI, Sharif M (1996) Cytokines in osteoarthritis: mediators or markers of joint destruction? Semin Arthritis Rheum 25: 254–272. [DOI] [PubMed] [Google Scholar]
- 43. Borzi RM, Mazzetti I, Cattini L, Uguccioni M, Baggiolini M, et al. (2000) Human chondrocytes express functional chemokine receptors and release matrix-degrading enzymes in response to C-X-C and C-C chemokines. Arthritis Rheum 43: 1734–1741. [DOI] [PubMed] [Google Scholar]
- 44. Yuan GH, Masuko-Hongo K, Nishioka K (2001) Role of chemokines/chemokine receptor systems in cartilage degradation. Drug News Perspect 14: 591–600. [PubMed] [Google Scholar]
- 45. Yuan GH, Masuko-Hongo K, Sakata M, Tsuruha J, Onuma H, et al. (2001) The role of C-C chemokines and their receptors in osteoarthritis. Arthritis Rheum 44: 1056–1070. [DOI] [PubMed] [Google Scholar]
- 46. Martin P, Leibovich SJ (2005) Inflammatory cells during wound repair: the good, the bad and the ugly. Trends Cell Biol 15: 599–607. [DOI] [PubMed] [Google Scholar]
- 47. Dawson TC, Lentsch AB, Wang Z, Cowhig JE, Rot A, et al. (2000) Exaggerated response to endotoxin in mice lacking the Duffy antigen/receptor for chemokines (DARC). Blood 96: 1681–1684. [PubMed] [Google Scholar]
- 48. Fukuma N, Akimitsu N, Hamamoto H, Kusuhara H, Sugiyama Y, et al. (2003) A role of the Duffy antigen for the maintenance of plasma chemokine concentrations. Biochem Biophys Res Commun 303: 137–139. [DOI] [PubMed] [Google Scholar]
- 49. Vielhauer V, Allam R, Lindenmeyer MT, Cohen CD, Draganovici D, et al. (2009) Efficient renal recruitment of macrophages and T cells in mice lacking the duffy antigen/receptor for chemokines. Am J Pathol 175: 119–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Kolar P, Gaber T, Perka C, Duda GN, Buttgereit F (2011) Human early fracture hematoma is characterized by inflammation and hypoxia. Clin Orthop Relat Res 469: 3118–3126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Recknagel S, Bindl R, Brochhausen C, Gockelmann M, Wehner T, et al. (2013) Systemic inflammation induced by a thoracic trauma alters the cellular composition of the early fracture callus. J Trauma Acute Care Surg 74: 531–537. [DOI] [PubMed] [Google Scholar]
- 52. Reich D, Nalls MA, Kao WH, Akylbekova EL, Tandon A, et al. (2009) Reduced neutrophil count in people of African descent is due to a regulatory variant in the Duffy antigen receptor for chemokines gene. PLoS Genet 5: e1000360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Walsh M, Davidovitch RI, Egol KA (2010) Ethnic disparities in recovery following distal radial fracture. J Bone Joint Surg Am 92: 1082–1087. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Information on the primers used for quantitative PCR.
(DOC)
Real Time PCR. Raw Data. Real Time PCR was done using cDNA prepared from RNA isolated from fracture calluses derived from Darc-KO and wild type mice at three time points post-fracture.
(XLS)