

Abstract
Nitric oxide is a critical regulator of blood pressure (BP) and inflammation, and female spontaneously hypertensive rats (SHR) have higher renal nitric oxide bioavailability than males. We hypothesize that female SHR will have a greater rise in BP and renal T cell infiltration in response to nitric oxide synthase (NOS) inhibition than males. Both male and female SHR displayed a dose-dependent increase in BP to the nonspecific NOS inhibitor NG-nitro-l-arginine methyl ester (l-NAME: 2, 5, and 7 mg·kg−1·day−1 for 4 days each); however, females exhibited a greater increase in BP than males. Treatment of male and female SHR with 7 mg·kg−1·day−1 l-NAME for 2 wk significantly increased BP in both sexes; however, prior exposure to l-NAME only increased BP sensitivity to chronic NOS inhibition in females. l-NAME-induced hypertension increased renal T cell infiltration and indices of renal injury in both sexes, yet female SHR exhibited greater increases in Th17 cells and greater decreases in regulatory T cells than males. Chronic l-NAME was also associated with larger increases in renal cortical adhesion molecule expression in female SHR. The use of triple therapy to block l-NAME-mediated increases in BP attenuated l-NAME-induced increases in renal T cell counts and normalized adhesion molecule expression in SHR, suggesting that l-NAME-induced increases in renal T cells were dependent on both increases in BP and NOS inhibition. Our data suggest that NOS is critical in the ability of SHR, females in particular, to maintain BP and limit a pro-inflammatory renal T cell profile.
Keywords: nitric oxide synthase, kidney injury, hypertension, sex, inflammation
clinical and experimental studies have established that there is a sex difference in the incidence of hypertension, with males developing high blood pressure (BP) at younger ages than females (7, 33). The molecular mechanisms responsible for the sex difference remains unclear, although the nitric oxide (NO)/NO synthase (NOS) pathway has been implicated. NO is a key regulator of BP (4, 13), and the incidence and progression of hypertension has been linked with NO deficiencies in experimental animal models (3, 4). There is also a sex difference in the NO/NOS pathway; NO bioavailability is greater in females compared with males (11, 14). Therefore, a sex difference in the NO/NOS system may contribute to sexual dimorphisms in BP.
Spontaneously hypertensive rats (SHR) are a genetic model of hypertension where females have a slower elevation in BP with age compared with males (27, 33). We recently reported that female SHR have greater renal NO bioavailability than males (32). However, the implication of these findings on the role of NOS in BP control remains unknown. Chronic NOS inhibition induces hypertension in normotensive male rats and exacerbates the progression of hypertension in male SHR (4, 24, 35). There are conflicting reports in the literature regarding the impact of sex on the response to chronic NOS inhibition. Normotensive female rats have been reported to be more sensitive (36), less sensitive (19, 30, 35), or equally sensitive (38) to NOS inhibition-induced hypertension compared with age-matched males.
T cells play a critical role in the progression of hypertension and renal injury in male experimental models of hypertension (10, 16), and we recently published that female SHR have a less pro-inflammatory renal T cell profile than males (34). NO inhibits adhesion molecule expression in vitro and in vivo to decrease vascular monocyte adhesion (9, 15), and lymphocyte suppression with mycophenolate mofetil (MMF) attenuates NG-nitro-l-arginine methyl ester (l-NAME)-induced hypertension in male Sprague-Dawley rats (12, 26). However, it is unknown how chronic NOS inhibition impacts the renal T cell profile in either sex. Based on the importance of the NOS pathway in maintaining BP and the greater NO bioavailability in female SHR compared with males (32), the current study was designed to test the hypothesis that female SHR are more dependent on NOS to maintain BP compared with males. We also hypothesized that greater sensitivity of female SHR to l-NAME-induced hypertension will be associated with greater elevations in renal T cell infiltration.
METHODS
Animals
Male and female SHR (Harlan Laboratories, Indianapolis, IN) were studied in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved and monitored by the Georgia Regents University IACUC. At 10 wk of age male and female SHR were implanted with telemetry devices (Data Sciences, St. Paul, MN) to monitor BP as previously described (31, 33). Rats were allowed 1 wk recovery and 1 wk of baseline BP recording before being randomized to receive vehicle (tap water) or the NOS inhibitor l-NAME. Two experimental approaches were employed to assess the contribution of NO to BP control in male and female SHR: 1) BP dose response and 2) chronic l-NAME treatment. For all studies l-NAME was given in drinking water. Rats were individually housed to allow for daily measurements of water intake; all rats were weighed every 3 days. l-NAME was administered according to both weight and water intake to maintain appropriate dosage in each sex. At the end of all studies rats were anesthetized with ketamine-xylazine (50 and 6 mg/kg ip, respectively; Phoenix Pharmaceuticals, St. Joseph, MO), and a terminal blood sample was taken and kidneys were isolated.
l-NAME Dose Response
Rats were randomized to receive vehicle (tap water; n = 5) or increasing doses of l-NAME in drinking water: 2, 5, and 7 mg·kg−1·day−1 (n = 5–6; Sigma-Aldrich, St. Louis, MO). Each dose was given for 4 days. Pilot studies using 0.1, 0.5, and 2 mg·kg−1·day−1 l-NAME revealed 2 mg·kg−1·day−1 l-NAME as the lowest dose to result in a change in BP from baseline in either sex (data not shown). l-NAME treatment was then suspended for 1 wk to assess the ability of BP to return to baseline values in both sexes before initiating chronic BP studies.
Chronic l-NAME
The dose of 7 mg·kg−1·day−1 (equivalent to ∼80 mg/l) l-NAME is comparable to commonly used doses in the literature to induce l-NAME hypertension (5, 24). Two groups of male and female SHR were treated with l-NAME at 7 mg·kg−1·day−1 for 14 days (n = 7–9). Group 1 were rats from the dose-response study; group 2 were age-matched rats that had not previously been exposed to l-NAME (l-NAME naïve). l-NAME naïve rats were included to assess not only the impact of prior exposure to l-NAME on the chronic BP responses, but also because BP did not return to baseline values in female SHR following the l-NAME dose-response studies in group 1.
Triple Therapy
An additional set of male and female SHR were treated with l-NAME at 7 mg·kg−1·day−1 for 14 days in the presence of triple therapy [TTx: reserpine, hydralazine, and hydrochlorothiazide (20); n = 5] to block l-NAME-induced increases in BP. TTx was not used to abolish hypertension in SHR; BP in these rats was maintained at levels comparable to vehicle control SHR.
Renal Health and Histological Analysis
A subset of rats were placed in metabolic cages for 24-h urine collection before beginning and at the end of the 7 mg·kg−1·day−1 l-NAME treatment to measure total protein (Bradford assay; Bio-Rad, Hercules, CA), albumin, and nephrin excretion (Exocell, Philadelphia, PA). Kidneys were prepared for histological analysis as previously described to assess renal injury and slides were examined in a blinded manner (33). Histological examination of kidneys was performed following staining using periodic acid-Schiff (PASH), Masson's trichrome, and Picro-Sirus Red staining.
Analytical Flow Cytometry
Whole blood and cell suspensions of kidneys were prepared and phenotypic and intracellular analyses performed as previously described to determine numbers of CD3+ and CD4+ T cells, regulatory T cells (Tregs; CD3+/CD4+/Foxp3+) or Th17 cells (CD3+/CD4+/ROR-γ+; all antibodies from BD Biosciences, San Diego, CA) (1, 34). Antibody specificity was confirmed using isotype controls.
Real-Time Polymerase Chain Reaction
RNA was isolated from 30 to 50 mg of renal cortex and medulla from control, l-NAME, and l-NAME/TTx-treated SHR (n = 5–6) using RNeasy Plus Mini kit (all reagents and primers from Qiagen, Valencia, CA). A blend of oligonucleotide and random hexanucleotide primers were used for reverse transcription of 1 μg of RNA using Quantifect Reverse Transcription kit and real-time polymerase chain reaction (RT-PCR) was performed with QuantiTect SYBR Green RT-PCR kit. GAPDH was the internal standard, and mRNA levels were expressed relative to control in each sex.
Statistical Analysis
All data are expressed as means ± SE. Telemetry data within each sex were analyzed using repeated measures ANOVA with Greenhouse-Geisser correction. Telemetry data between sexes and between control and l-NAME-treated rats were compared using T-test. Flow cytometry, adhesion molecule, and urinary excretion data were compared using one-way ANOVA followed by a Newman-Keul's post hoc test. For all comparisons, differences were statistically significant with P < 0.05. Analyses were performed using GraphPad Prism Version 5.0 (GraphPad Software, La Jolla, CA).
RESULTS
Animal Characteristics
Dose response.
At baseline, male SHR were larger than females (274 ± 6 vs. 177 ± 2 g; P < 0.05) and consumed more water (30 ± 1 vs. 26 ± 2 ml; P < 0.05). During the study, both sexes gained weight (males: 42 ± 4 g; females: 16 ± 2 g). There was not a difference in weight gain in vehicle versus l-NAME-treated rats. Water consumption did not change with l-NAME.
Chronic l-NAME.
Male and female SHR in group 1, previously exposed to l-NAME, gained weight during l-NAME treatment (male: 318 ± 5 to 340 ± 3 g; female: 205 ± 2 to 212 ± 7; P < 0.05). Male SHR in group 2, naïve to previous l-NAME, were larger than females at baseline (315 ± 6 vs. 185 ± 2 g, P < 0.05); neither sex gained weight during l-NAME treatment (313 ± 7 vs. 185 ± 3 g). TTx did not alter weight gain in males (319 ± 4 to 343 ± 2 g; P < 0.05), although female SHR receiving TTx did not gain weight (197 ± 3 to 198 ± 5 g).
l-NAME Increases BP in Male and Female SHR
Low-dose l-NAME results in a greater increase in BP in female SHR.
Female and male SHR were treated with increasing doses of l-NAME and BP was measured via telemetry (Fig. 1A). BP was greater in male SHR than female SHR at baseline (P < 0.05), and the sex difference in BP was abolished upon treatment with l-NAME. Male SHR exhibited 7%, 12%, and 14% increases in BP in response to 2, 5, and 7 mg·kg−1·day−1 l-NAME, respectively, compared with age-matched vehicle controls. BP returned to vehicle control levels with termination of l-NAME treatment in males (Fig. 1B). l-NAME increased BP in female SHR 11%, 12%, and 20% in response to 2, 5, and 7 mg·kg−1·day−1 l-NAME, respectively, compared with vehicle controls (Fig. 1C). BP remained elevated (by 7%) 4 days after l-NAME treatment was terminated in female SHR (Fig. 1C, P = 0.042). BP in vehicle control rats did not change during the study.
Fig. 1.

Blood pressure (BP) measured by telemetry in response to increasing doses of NG-nitro-l-arginine methyl ester (l-NAME) or vehicle in age-matched male and female spontaneously hypertensive rats (SHR). Female SHR have greater BP sensitivity to l-NAME compared with male SHR (A), although both male (B) and female SHR (C) exhibit a dose-dependent increase in BP compared with same-sex vehicle controls. *Significant difference from male SHR; #significant difference from same sex baseline BP and vehicle control.
Prior l-NAME exposure alters the BP response to chronic LNAME in female SHR.
BP in female SHR remained elevated 7 days after the l-NAME dose-response study was stopped; as a result there was not a sex difference in BP when starting 2 wk of 7 mg·kg−1·day−1 l-NAME in group 1 (Fig. 2A). BP increased over the treatment period to the same extent in both male and female SHR such that BP was comparable in both sexes throughout l-NAME treatment. Male SHR in group 2 (l-NAME naïve) had a higher baseline BP compared with female SHR (Fig. 2B). BP significantly increased within 24 h of initiating l-NAME and continued to increase throughout the treatment period; however, BP increased to a greater degree in female SHR during the final 2 days of l-NAME treatment abolishing the sex difference in BP. The BP response to chronic l-NAME was comparable in male SHR regardless of prior l-NAME exposure (Fig. 2C). In contrast, female SHR from group 1 exhibited a greater increase in BP in the first week of l-NAME treatment compared with l-NAME naïve females in group 2 (24 ± 3% vs. 14 ± 2%, P < 0.05; Fig. 2D). BP continued to increase in both groups during the second week of l-NAME treatment; however, the increase was greater in l-NAME naïve female SHR, particularly in the last 2 days of treatment (43 ± 1% vs. 35 ± 5%, P < 0.05) such that BPs were comparable at the end of the 2 wk.
Fig. 2.

BP in response to chronic l-NAME treatment (7 mg·kg−1·day−1) in male and female SHR. Chronic l-NAME significantly increased BP in both males and females that have (A) and have not (B) been previously exposed to l-NAME. Prior exposure to l-NAME did not impact BP responses to l-NAME in male SHR (C). In contrast, previous l-NAME treatment sensitized BP responses in female SHR (D). #Significant difference from the baseline BP in both groups; *significant difference from male SHR; &significant difference from age-matched rat.
Chronic l-NAME Induces Renal Injury in Male and Female SHR
Histological examination of kidneys revealed that l-NAME treatment was associated with thickening, necrosis, and thrombosis of the glomerular and interstitial arteries, tubular cast formation and fibrosis, and glomerular necrosis and ischemia in both sexes. Kidneys of vehicle control male and female SHR were structurally unremarkable. For all histological indices of renal injury, evidence of damage was moderate to severe in the males treated with l-NAME and mild to moderate in l-NAME-treated females (Table 1; Fig. 3).
Table 1.
Evidence of renal injury in l-NAME-treated males and l-NAME-treated females
| Male | Male + l-NAME | Female | Female + l-NAME | |
|---|---|---|---|---|
| Interstitial Artery | ||||
| Thickening | 0 | 2.4 ± 0.2† | 0 | 1.3 ± 0.3*† |
| Necrosis | 0 | 2.4 ± 0.2† | 0 | 1.0 ± 0.1*† |
| Thrombosis | 0 | 2.6 ± 0.2† | 0 | 1.0 ± 0.1*† |
| Glomerulus | ||||
| Tuft necrosis | 0 | 1.4 ± 0.2† | 0 | 0 |
| Hyalinosis | 0 | 1.4 ± 0.2† | 0 | 0.3 ± 0.3* |
| Thrombosis | 0 | 1.4 ± 0.2† | 0 | 0.3 ± 0.3* |
Values are means ± SE; n =5. Renal injury scores assessed by an independent, blinded observer. Score: 0, none; 1, mild; 2, moderate; 3, severe. l-NAME, NG-nitro-l-arginine methyl ester.
Significant difference from males;
significant difference from same sex control, P < 0.05.
Fig. 3.

Analysis of PASH (A–D) and Masson's trichrome staining (E–H) in the renal cortex of male and female SHR following vehicle or l-NAME treatment at 7 mg·kg−1·day−1 for 2 wk. l-NAME-induced renal injury in both male and female SHR; however, the degree of injury was more severe in male SHR, ×20 magnification; n = 5.
Urinary protein, albumin, and nephrin excretion were also measured before and after l-NAME treatment. l-NAME significantly increased urinary protein, albumin, and nephrin excretion in both sexes (Table 2). Male SHR had greater protein (P < 0.01) and albumin (P = 0.05) excretion than females at baseline, and protein excretion remained higher in males following l-NAME treatment (P = 0.07). There were no significant differences in nephrin excretion between control or l-NAME-treated male and female SHR or in albumin excretion following l-NAME between the sexes.
Table 2.
Urinary excretion data to assess renal injury
| Protein Excretion, mg/day | Albumin Excretion, mg/day | Nephrin Excretion, mg/day | ||
|---|---|---|---|---|
| Male SHR | Control | 20 ± 2 | 0.4 ± 0.1 | 0.05 ± 0.01 |
| l-NAME | 116 ± 23* | 2.2 ± 0.6* | 0.35 ± 0.10* | |
| l-NAME+ TTx | 18 ± 2† | 1.1 ± 0.1† | 0.15 ± 0.01† | |
| Female SHR | Control | 3 ± 1 | 0.3 ± 0.03 | 0.04 ± 0.01 |
| l-NAME | 66 ± 4* | 2.8 ± 0.7* | 0.30 ± 0.04* | |
| l-NAME+ TTx | 3 ± 1† | 0.5 ± 0.1† | 0.17 ± 0.02† | |
Values are means ± SE. SHR, spontaneoulsy hypertensive rats. TTx, triple therapy: reserpine, hydralazine, and hydrochlorothiazide.
Difference from same sex vehicle-control;
significant difference from l-NAME.
Chronic l-NAME Treatment: Triple Therapy
NOS inhibition or deficiency is associated with increases in renal immune cell infiltration (17, 18, 26). To determine the relative contribution of NOS inhibition versus BP elevation on renal T cell profiles in male and female SHR, additional SHR received triple therapy (TTx) in conjunction with l-NAME to block increases in BP. TTx abolished l-NAME hypertension in male and female SHR; average mean arterial pressure over the treatment period was 131 ± 2 mmHg in male SHR and 126 ± 2 mmHg in female SHR. TTx blocked l-NAME-induced increases in protein excretion in male and female SHR and attenuated l-NAME-induced increases in albuminuria and nephrinuria (Table 2). Urinary protein and albumin excretion were significantly less in female SHR than in males following l-NAME + TTx (P < 0.01), and albuminuria was comparable between the sexes.
Chronic l-NAME Treatment: T Cell Profile
Blood.
l-NAME increased circulating CD3+ and CD4+ cells in male and female SHR relative to vehicle-treated controls with no changes in Tregs or Th17 cells (Table 3). TTx blunted l-NAME-induced increases in circulating CD3+ and CD4+ cells and decreased Th17 cells below vehicle control in both sexes (P < 0.05). Although not altered by l-NAME alone TTx increased circulating Tregs relative to both l-NAME and vehicle controls in male and female SHR (P < 0.05). There were no sex differences in CD3+ or Th17 absolute cell counts in whole blood, although females had more circulating CD4+ cells and fewer Tregs under all treatments (Table 4).
Table 3.
Flow cytometry analysis of whole blood
| CD3+ | CD4+ | Foxp3+ | ROR-γ+ | |
|---|---|---|---|---|
| Male SHR | ||||
| Control | 68 ± 1 | 38 ± 2 | 5 ± 1 | 25 ± 1 |
| l-NAME | 77 ± 1* | 52 ± 2* | 4 ± 1 | 28 ± 1 |
| l-NAME+ TTx | 72 ± 1*† | 41 ± 1† | 11 ± 1*† | 22 ± 1† |
| Female SHR | ||||
| Control | 72 ± 1 | 51 ± 2 | 4 ± 1 | 30 ± 0.7 |
| l-NAME | 83 ± 1* | 56 ± 1* | 2 ± 1 | 34 ± 2 |
| l-NAME+ TTx | 73 ± 1† | 48 ± 1† | 10 ± 1*† | 21 ± 2*† |
Values are means ± SE.
Difference from same sex vehicle-control;
indicates significant difference from l-NAME. CD3+ cells are expressed as % of total blood cells. CD4+ cells are expressed as % of CD3+ cells. Foxp3+ (Tregs) and ROR-γ+ cells are expressed as % CD3+CD4+ cells.
Table 4.
Absolute cell counts of flow cytometry analysis in whole blood
| CD3+ | CD4+ | Foxp3+ | ROR-γ+ | |
|---|---|---|---|---|
| Male SHR | ||||
| Control | 68 ± 1 | 25 ± 1 | 1.3 ± 0.1 | 6.5 ± 0.3 |
| l-NAME | 77 ± 1* | 40 ± 1* | 1.7 ± 0.1 | 11.2 ± 0.5* |
| l-NAME+ TTx | 72 ± 1† | 29 ± 1† | 3.2 ± 0.3*† | 6.5 ± 0.3† |
| Female SHR | ||||
| Control | 72 ± 1 | 37 ± 1 | 1.5 ± 0.2 | 10.8 ± 0.2 |
| l-NAME | 83 ± 2* | 46 ± 1* | 1.0 ± 0.1* | 15.7 ± 0.8 |
| l-NAME+ TTx | 73 ± 1† | 35 ± 1† | 3.6 ± 0.3*† | 7.4 ± 0.6*† |
Values are means ± SE; All numbers × 103. All cell counts in whole blood are from an analysis of 100,000 cells in total.
Difference from same sex vehicle-control;
significant difference from l-NAME-treated rats.
Kidney.
l-NAME increased renal CD3+ and Th17 cells in kidneys from male and female SHR compared with vehicle control (P < 0.001) and TTx mitigated this increase; however, CD3+ and Th17 cells in TTx groups remained significantly greater than in same sex vehicle controls (Figs. 4, A and D and 5, A and D; P < 0.05). TTx attenuated l-NAME-induced increases in renal CD4+ cells in male SHR and blocked the increase in females (Figs. 4B and 5B; P < 0.001). l-NAME decreased Tregs in kidneys of male and female SHR compared with same-sex vehicle controls (Figs. 4C and 5C; P < 0.05). In male SHR, TTx increased Tregs compared with same sex vehicle and l-NAME-treated rats (P < 0.001), but in female SHR TTx only increased Tregs compared with same sex vehicle rats (P < 0.001). Although the overall impact of l-NAME and TTx was similar on renal T cell profiles in male and female SHR, the magnitude of the changes differed. l-NAME resulted in greater increases in CD3+ and Th17 cells in female SHR (% increase: 226% and 596%) than in males (124% and 220%) compared with same-sex vehicle controls. Female SHR also had a greater decrease in Tregs after l-NAME treatment than males (% decrease: 86% vs. 49%). As a result, after l-NAME treatment female SHR had more renal CD3+ (P = 0.0009), CD4+ (P = 0.0006), and Th17 cells (P = 0.0009) than male SHR. These results are supported by an analysis of absolute numbers of renal T cells (Table 5).
Fig. 4.
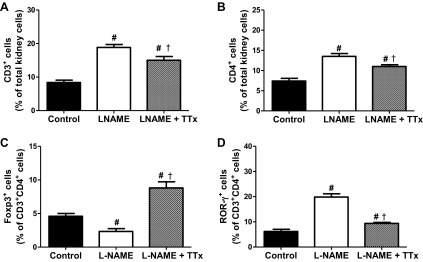
Renal T cell profiles in control, l-NAME, and l-NAME + triple therapy (TTx: reserpine, hydralazine, and hydrochlorothiazide)-treated male SHR after 2 wks of 7 mg·kg−1·day−1 l-NAME. CD3+ (A) and CD4+ cells (B) are expressed as percentage of total kidney cells, Foxp3+ (Tregs; C) and ROR-γ+ (Th17; D) cells are expressed as percentage of CD3+CD4+ cells. l-NAME increased renal pro-inflammatory cells and decreased Tregs in male SHR; TTx mitigated these changes. #Difference from same sex vehicle-control; †significant difference from l-NAME.
Fig. 5.
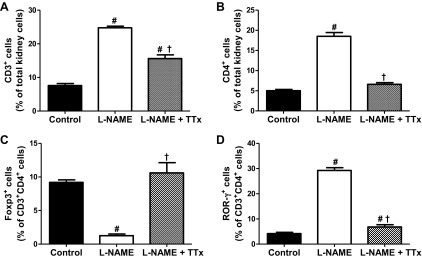
Renal T cell profiles in control, l-NAME, and l-NAME + TTx-treated female SHR after 2 wks instead of 14 days for consistency of 7 mg·kg−1·day−1 l-NAME. CD3+ (A) and CD4+ cells (B) are expressed as percentage of total kidney cells Foxp3+ (Tregs; C), and ROR-γ+ (Th17; D) cells are expressed as percentage of CD3+CD4+ cells. l-NAME increased renal pro-inflammatory cells and decreased Tregs in female SHR; TTx mitigated these changes. #Difference from same sex vehicle-control; †significant difference from l-NAME.
Table 5.
Absolute cell counts of flow cytometry analysis in kidneys
| CD3+ | CD4+ | Foxp3+ | ROR-γ+ | |
|---|---|---|---|---|
| Male SHR | ||||
| Control | 69 ± 6 | 28 ± 1 | 1.3 ± 0.1 | 1.8 ± 0.2 |
| l-NAME | 154 ± 7 * | 79 ± 1* | 1.8 ± 0.3* | 15.6 ± 1.0* |
| l-NAME+ TTx | 123 ± 9*† | 49 ± 1 † | 4.3 ± 0.5*† | 4.6 ± 0.2*† |
| Female SHR | ||||
| Control | 53 ± 4 | 24 ± 2 | 2.2 ± 0.1 | 1.0 ± 0.1 |
| l-NAME | 173 ± 3* | 102 ± 2* | 1.3 ± 0.3* | 29.8 ± 1.1* |
| l-NAME+ TTx | 109 ± 8*† | 54 ± 2 † | 5.7 ± 0.8*† | 3.7 ± 0.5*† |
Values are means ± SE. All numbers × 105. For the kidney, in males we counted 82 × 106 ± 5 × 106 total kidney cells while in females we counted 70 × 106 ± 5 × 106 total kidney cells.
Difference from same sex vehicle-control;
significant difference from l-NAME-treated rats.
Chronic l-NAME Treatment: Adhesion Molecules
NO inhibits adhesion molecule expression in vitro and in vivo and decreases vascular monocyte adhesion (9, 15). Additional experiments assessed intercellular adhesion molecule (ICAM) and vascular cell adhesion molecule (VCAM) mRNA expression in the renal cortex and medulla of male and female SHR (Figures 6 and 7). l-NAME increased renal cortical ICAM and VCAM mRNA expression in male and female SHR, and TTx abolished l-NAME induced increases in adhesion molecule expression (Fig. 6, P < 0.05). Female SHR displayed greater increases in cortical ICAM and VCAM expression (109% and 66%) than male SHR (39% and 34%) in response to l-NAME. Medullary ICAM and VCAM were significantly increased only in male SHR with l-NAME (240% and 127%; Fig. 7). TTx did not alter medullary ICAM expression in either sex but abolished l-NAME-induced increases in VCAM in male SHR.
Fig. 6.
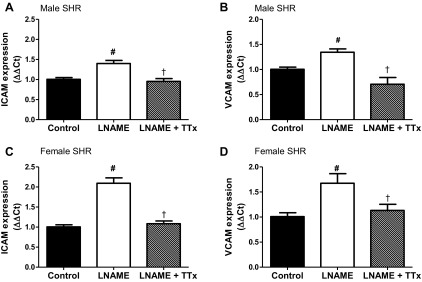
ICAM (A and C) and VCAM (B and D) mRNA expression in the renal cortex of male and female SHR. l-NAME-induced increases in adhesion molecule expression in male and female SHR is blocked by TTx. #Difference from same sex vehicle-control; †significant difference from l-NAME.
Fig. 7.

ICAM (A and C) and VCAM (B and D) mRNA expression in the renal medulla of male and female SHR. l-NAME increases medullary adhesion molecule expression in male, but not female SHR. TTx blocks l-NAME-induced increase in VCAM but not ICAM. #Difference from same sex vehicle-control; †significant difference from l-NAME.
DISCUSSION
The primary novel findings of the current study were that BP in female SHR is more sensitive to NOS inhibition than in males and NOS inhibition sensitizes BP in female SHR, but not males, to a second exposure to l-NAME. Moreover, the loss of the sex difference in BP in male and female SHR with NOS inhibition was associated with greater increases in renal pro-inflammatory T cells in female SHR than in males. Finally, l-NAME-induced increases in renal T cells were dependent on both increases in BP and NOS inhibition.
Consistent with experimental and epidemiological studies indicating greater NO bioavailability in females relative to males (11, 14, 23), female SHR in the current study were more sensitive to l-NAME-induced alterations in BP than males. This was illustrated by a number of results: female SHR exhibited a greater increase in BP to low-dose l-NAME than males thereby abolishing the sex difference in basal BP; BP in female SHR did not return to basal values following the discontinuation of l-NAME; and prior exposure to l-NAME sensitized female SHR to increases in BP to chronic NOS inhibition. SHR are very sensitive to NOS inhibition-induced increases in BP. Although BP plateaued at each dose in the dose-response period, we cannot rule out that continuing treatment of lower doses of l-NAME longer than 4 days may have resulted in additional increases in BP. However, for the chronic phase of the study we were interested in using a dose closer to those more commonly used in the literature to induce l-NAME hypertension in SHR (5, 24). There is little consensus in the literature regarding the impact of sex on BP responses to NOS inhibition. In contrast to our findings in SHR, male Sprague-Dawley and Wistar-Kyoto rats exhibited larger increases in BP to l-NAME and Nω-nitro-l-arginine (l-NNA) than females (19, 30, 35). However, these studies were all of longer duration (up to 24 wk) than the current study. Consistent with our study, female Wistar are more sensitive to l-NAME-induced increases in BP than males over a 10-wk treatment period, and BP in females remained elevated to a greater extent than in males following the termination of l-NAME treatment (36). However, 2 wk of l-NAME treatment has also been reported to result in comparable increases in BP in male and female Wistar rats (38). Inconsistencies in the literature are likely related to the dose of NOS inhibitor used, duration of treatment, age and strain of the animals, and the method used to measure BP. Our study is the first to examine the contribution of NOS to BP control in hypertensive males and females using telemetry.
Our data suggest that once the NOS system is compromised in female SHR, they lose the ability to mitigate BP increases and exhibit a BP response to NOS inhibition comparable to males. Not only did BP remain elevated only in female SHR following the termination of the l-NAME dose-response experiments in group 1, but this sex-specific carryover effect on BP reactivity in females extended for more than 10 days into the secondary l-NAME infusion. There is indirect evidence supporting a sex difference in the contribution of NOS to BP control in SHR. Maternal supplementation with the NOS substrate citrulline from day 7 of gestation until 6 wk of age resulted in a sustained decrease in systolic BP in female SHR, although BP in males was comparable to untreated controls by 24 wk of age (21). One limitation of the current study is that it is not known if sex differentially impacts the metabolism, efficacy, or intestinal absorption of l-NAME. As such, we cannot rule out the possibility that at the same dose of l-NAME female SHR have a greater degree of global NOS inhibition relative to males.
Greater BP sensitivity to NOS inhibition in female SHR was paralleled by greater increases in protein and albumin excretion; however, the degree of structural damage following chronic l-NAME was greater in male SHR. TTx normalized l-NAME-induced increases in protein and nephrin excretion in female SHR, suggesting that the greater increase in protein and nephrin excretion in females results from greater increase in BP. The acute rise in BP in females may result in a larger insult on the functional integrity of the glomerulus that is not yet evident histologically. Greater structural alterations in kidneys of male SHR likely reflects the fact that kidneys of male SHR are exposed to a greater pressure load for a longer period of time compared with kidneys of females. Contrary to our results, male Wistar rats display a greater increase in proteinuria following chronic l-NAME (30), and male Sprague-Dawley rats are more sensitive to l-NNA-induced proteinuria compared with females (35). However, in both of these studies, the male rats also have a larger increase in BP in response to chronic NOS inhibition which would be consistent with greater renal injury.
l-NAME hypertension increases vascular and renal monocyte/macrophage infiltration in male SHR and contributes to end-organ damage (17, 18), yet little is known regarding the impact of l-NAME hypertension on T cells. In the current study, chronic l-NAME significantly increased pro-inflammatory T cells in the kidneys of SHR, although the increase was greater in females. It should be noted that female SHR have smaller kidneys than males and a smaller number of total cells were counted for the flow cytometric analysis, therefore, our results may underestimate the absolute magnitude of the sex difference in the pro-inflammatory immune cell profile following l-NAME. Although not directly tested, we hypothesize that the greater increase in pro-inflammatory T cells in female SHR is related to the greater BP sensitivity and increases in renal injury. This is consistent with the finding that the lymphocyte suppressant MMF attenuates l-NAME hypertension and renal damage in male Sprague-Dawley rats (26). However, coadministration of MMF with l-NAME in male Munich-Wistar rats did not alter l-NAME-induced hypertension but abolished l-NAME-induced increases in T cell infiltration and attenuated renal injury (12), suggesting that l-NAME-induced increases in BP are independent of T cells, although renal injury is not.
To differentiate the relative contribution of increases in BP and loss of NO on modulating T cells, additional rats were treated with TTx, which implicated both a BP-dependent and a NOS-dependent component to the T cell profile in SHR with l-NAME hypertension. Interestingly, l-NAME-induced increases in CD3+ and CD4+ T cells in male SHR were primarily NO dependent, whereas increases in CD4+ cells in female SHR were BP dependent, suggesting sex-specific mechanisms regulate renal T cells. The finding that TTx abolished l-NAME-induced increases in BP despite minimal decreases in total CD3+ cells and CD4+ renal T cells calls into question the role of T cells in l-NAME hypertension. However, l-NAME resulted in a BP-dependent increase in renal Th17 cells in both sexes, and Th17 cells have been directly implicated in increases in BP (22). Therefore, measuring total CD3+ or CD4+ T cells alone may be insufficient to fully understanding the impact of T cells on BP control. Indeed, TTx also increased renal Tregs, which limit increases in BP in male rats (2). Therefore, greater increases in Tregs may mitigate Th17 cell-induced increases in BP. Renal T cell profiles were only assessed after 14 days of l-NAME treatment in the current study, and l-NAME results in a rapid rise in BP in SHR, which is likely independent of the renal T-cell responses. However, increases renal T cells could have taken place at a much earlier time point coinciding with initial increases in BP, consistent with both BP and NO independently influencing the renal T-cell profile. A limitation of the current study is that TTx itself may impact the T cell profile. Hydralazine has been shown to reduce the number of adherent and migrating leukocytes in the vasculature of male SHR independent on an effect on BP (29).
To begin to assess a mechanism by which NO may impact T cell infiltration, adhesion molecule expression was examined. Endothelial cells modulate inflammatory cell adhesion and migration via adhesion molecules including VCAM and ICAM (6), and NO inhibits adhesion molecule expression (9, 15). ICAM and VCAM contribute to T cell adhesion and infiltration (25) in male rats with acute renal allograft rejection (28) and in mice with lupus nephritis (39). Consistent with greater increase in T cell infiltration, female SHR exhibited greater increases in renal cortical adhesion molecule expression than males in the current study. Female SHR also possessed lower basal levels of renal adhesion molecule expression, which may contribute to sex difference in the T cell profile in control rats. In contrast, only male SHR exhibited increases in medullary VCAM or ICAM, suggesting that there are sex differences in the impact of TTx on immune cells. In both sexes TTx blocked l-NAME-induced increases in cortical adhesion molecule expression, BP, and protein excretion but did not totally block T cell infiltration. These data suggest that BP drives the increase in VCAM and ICAM expression and that additional adhesion factors account for the remaining increases in CD3+ and Th17 cells in rats treated with TTx and for the robust increase in Tregs in male SHR. Alternatively, the current study did not assess the portion of the renal T cell profile that was due to recruitment of T cells into the kidney versus differentiation of resident naïve T cells.
Perspectives and Significance
There is a growing body of literature examining the role of T cells as a determining factor in the development and progression of numerous cardiovascular disorders in males. We recently published that immune suppression decreases BP in female SHR (34), yet there remains a scarcity of data on the role of T cells in hypertension in females or on how T cells are regulated between the sexes. Although women are more likely than men to develop innate immunological disorders, including rheumatoid arthritis and systemic lupus erythematosus (37), both of which have an increased risk of cardiovascular disease (8), little is known regarding the adaptive immune responses in women with hypertension. We propose that sex differences in the immune cell profiles underlie sex differences in the development and progression of many cardiovascular diseases including hypertension. Moreover, greater NO levels in females may offer cardioprotection by suppressing the activation and infiltration of effector T cells thereby limiting end-organ damage.
GRANTS
The authors acknowledge funding from the American Heart Association and the National Institutes of Health (American Heart Association Scientist Development Grant to A. A. Elmarakby; American Heart Association predoctoral fellowship to K. N. Brinson; and 1R01 HL-093271-01A1 and Scientist Development Grant to J. C. Sullivan).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: K.N.B., A.A.E., B.B., and J.C.S. conception and design of research; K.N.B., A.J.T., and G.R.C. performed experiments; K.N.B., A.A.E., A.J.T., G.R.C., T.Y., B.B., and J.C.S. analyzed data; K.N.B., T.Y., B.B., and J.C.S. interpreted results of experiments; K.N.B. and J.C.S. drafted manuscript; K.N.B., A.A.E., A.J.T., G.R.C., T.Y., B.B., and J.C.S. edited and revised manuscript; K.N.B., A.A.E., A.J.T., G.R.C., T.Y., B.B., and J.C.S. approved final version of manuscript; B.B., J.C.S., A.J.T., and K.N.B. prepared figures.
ACKNOWLEDGMENTS
The authors acknowledge the assistance of Vanessa Kemp and Margaret Zimmerman.
REFERENCES
- 1.Baban B, Chandler PR, Johnson BA, 3rd, Huang L, Li M, Sharpe ML, Francisco LM, Sharpe AH, Blazar BR, Munn DH, Mellor AL. Physiologic control of IDO competence in splenic dendritic cells. J Immunol 187: 2329–2335, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Barhoumi T, Kasal DA, Li MW, Shbat L, Laurant P, Neves MF, Paradis P, Schiffrin EL. T regulatory lymphocytes prevent angiotensin II-induced hypertension and vascular injury. Hypertension 57: 469–476, 2011 [DOI] [PubMed] [Google Scholar]
- 3.Baylis C. Nitric oxide deficiency in chronic kidney disease. Am J Physiol Renal Physiol 294: F1–F9, 2008 [DOI] [PubMed] [Google Scholar]
- 4.Baylis C, Mitruka B, Deng A. Chronic blockade of nitric oxide synthesis in the rat produces systemic hypertension and glomerular damage. J Clin Invest 90: 278–281, 1992 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Benter IF, Yousif MH, Anim JT, Cojocel C, Diz DI. Angiotensin-(1–7) prevents development of severe hypertension and end-organ damage in spontaneously hypertensive rats treated with l-NAME. Am J Physiol Heart Circ Physiol 290: H684–H691, 2006 [DOI] [PubMed] [Google Scholar]
- 6.Chen C, Mobley JL, Dwir O, Shimron F, Grabovsky V, Lobb RR, Shimizu Y, Alon R. High affinity very late antigen-4 subsets expressed on T cells are mandatory for spontaneous adhesion strengthening but not for rolling on VCAM-1 in shear flow. J Immunol 162: 1084–1095, 1999 [PubMed] [Google Scholar]
- 7.Cheng S, Xanthakis V, Sullivan LM, Vasan RS. Blood pressure tracking over the adult life course: patterns and correlates in the framingham heart study. Hypertension 60: 1393–1399, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chogle AR, Chakravarty A. Cardiovascular events in systemic lupus erythematosus and rheumatoid arthritis: emerging concepts, early diagnosis and management. J Assoc Physicians India 55: 32–40, 2007 [PubMed] [Google Scholar]
- 9.De Caterina R, Libby P, Peng HB, Thannickal VJ, Rajavashisth TB, Gimbrone MA, Jr, Shin WS, Liao JK. Nitric oxide decreases cytokine-induced endothelial activation. Nitric oxide selectively reduces endothelial expression of adhesion molecules and proinflammatory cytokines. J Clin Invest 96: 60–68, 1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.De Miguel C, Guo C, Lund H, Feng D, Mattson DL. Infiltrating T lymphocytes in the kidney increase oxidative stress and participate in the development of hypertension and renal disease. Am J Physiol Renal Physiol 300: F734–F742, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Forte P, Kneale BJ, Milne E, Chowienczyk PJ, Johnston A, Benjamin N, Ritter JM. Evidence for a difference in nitric oxide biosynthesis between healthy women and men. Hypertension 32: 730–734, 1998 [DOI] [PubMed] [Google Scholar]
- 12.Fujihara CK, Avancini Costa Malheiros DM, de Lourdes Noronha II, De Nucci G, Zatz R. Mycophenolate mofetil reduces renal injury in the chronic nitric oxide synthase inhibition model. Hypertension 37: 170–175, 2001 [DOI] [PubMed] [Google Scholar]
- 13.Gamboa A, Shibao C, Diedrich A, Choi L, Pohar B, Jordan J, Paranjape S, Farley G, Biaggioni I. Contribution of endothelial nitric oxide to blood pressure in humans. Hypertension 49: 170–177, 2007 [DOI] [PubMed] [Google Scholar]
- 14.Glushkovskaya-Semyachkina OV, Anishchenko TG, Sindyakova TA, Leksina OV, Berdnikova VA. Sex-related differences in nitric oxide content in healthy and hypertensive rats at rest and under stress conditions. Bull Exp Biol Med 142: 9–11, 2006 [DOI] [PubMed] [Google Scholar]
- 15.Harrison CB, Drummond GR, Sobey CG, Selemidis S. Evidence that nitric oxide inhibits vascular inflammation and superoxide production via a p47phox-dependent mechanism in mice. Clin Exp Pharmacol Physiol 37: 429–434, 2010 [DOI] [PubMed] [Google Scholar]
- 16.Harrison DG, Guzik TJ, Lob HE, Madhur MS, Marvar PJ, Thabet SR, Vinh A, Weyand CM. Inflammation, immunity, and hypertension. Hypertension 57: 132–140, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hsieh NK, Wang JY, Liu JC, Wang SD, Chen HI. Nitric oxide inhibition accelerates hypertension and induces perivascular inflammation in rats. Clin Exp Pharmacol Physiol 31: 212–218, 2004 [DOI] [PubMed] [Google Scholar]
- 18.Ikeda H, Tsuruya K, Toyonaga J, Masutani K, Hayashida H, Hirakata H, Iida M. Spironolactone suppresses inflammation and prevents l-NAME-induced renal injury in rats. Kidney Int 75: 147–155, 2009 [DOI] [PubMed] [Google Scholar]
- 19.Kamari Y, Peleg E, Leibowitz A, Grossman E. Blunted blood pressure response and elevated plasma adiponectin levels in female Sprague Dawley rats. Am J Hypertens 25: 612–619, 2012 [DOI] [PubMed] [Google Scholar]
- 20.Kang KT, Sullivan JC, Spradley FT, d'Uscio LV, Katusic ZS, Pollock JS. Antihypertensive therapy increases tetrahydrobiopterin levels and NO/cGMP signaling in small arteries of angiotensin II-infused hypertensive rats. Am J Physiol Heart Circ Physiol 300: H718–H724, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Koeners MP, van Faassen EE, Wesseling S, de Sain-van der Velden M, Koomans HA, Braam B, Joles JA. Maternal supplementation with citrulline increases renal nitric oxide in young spontaneously hypertensive rats and has long-term antihypertensive effects. Hypertension 50: 1077–1084, 2007 [DOI] [PubMed] [Google Scholar]
- 22.Madhur MS, Lob HE, McCann LA, Iwakura Y, Blinder Y, Guzik TJ, Harrison DG. Interleukin 17 promotes angiotensin II-induced hypertension and vascular dysfunction. Hypertension 55: 500–507, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Majmudar NG, Robson SC, Ford GA. Effects of the menopause, gender, and estrogen replacement therapy on vascular nitric oxide activity. J Clin Endocrinol Metab 85: 1577–1583, 2000 [DOI] [PubMed] [Google Scholar]
- 24.Ono H, Ono Y, Takanohashi A, Matsuoka H, Frohlich ED. Apoptosis and glomerular injury after prolonged nitric oxide synthase inhibition in spontaneously hypertensive rats. Hypertension 38: 1300–1306, 2001 [DOI] [PubMed] [Google Scholar]
- 25.Pober JS, Doukas J, Hughes CC, Savage CO, Munro JM, Cotran RS. The potential roles of vascular endothelium in immune reactions. Hum Immunol 28: 258–262, 1990 [DOI] [PubMed] [Google Scholar]
- 26.Quiroz Y, Pons H, Gordon KL, Rincon J, Chavez M, Parra G, Herrera-Acosta J, Gomez-Garre D, Largo R, Egido J, Johnson RJ, Rodriguez-Iturbe B. Mycophenolate mofetil prevents salt-sensitive hypertension resulting from nitric oxide synthesis inhibition. Am J Physiol Renal Physiol 281: F38–F47, 2001 [DOI] [PubMed] [Google Scholar]
- 27.Reckelhoff JF, Zhang H, Srivastava K. Gender differences in development of hypertension in spontaneously hypertensive rats: role of the renin-angiotensin system. Hypertension 35: 480–483, 2000 [DOI] [PubMed] [Google Scholar]
- 28.Renkonen R, Turunen JP, Rapola J, Hayry P. Characterization of high endothelial-like properties of peritubular capillary endothelium during acute renal allograft rejection. Am J Pathol 137: 643–651, 1990 [PMC free article] [PubMed] [Google Scholar]
- 29.Rodrigues SF, de Oliveira MA, dos Santos RA, Soares AG, de Cassia Tostes R, Carvalho MH, Fortes ZB. Hydralazine reduces leukocyte migration through different mechanisms in spontaneously hypertensive and normotensive rats. Eur J Pharmacol 589: 206–214, 2008 [DOI] [PubMed] [Google Scholar]
- 30.Sainz J, Osuna A, Wangensteen R, de Dios Luna J, Rodriguez-Gomez I, Duarte J, Moreno JM, Vargas F. Role of sex, gonadectomy and sex hormones in the development of nitric oxide inhibition-induced hypertension. Exp Physiol 89: 155–162, 2004 [DOI] [PubMed] [Google Scholar]
- 31.Sullivan JC, Bhatia K, Yamamoto T, Elmarakby AA. Angiotensin (1–7) receptor antagonism equalizes angiotensin II-induced hypertension in male and female spontaneously hypertensive rats. Hypertension 56: 658–666, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sullivan JC, Pardieck JL, Hyndman KA, Pollock JS. Renal NOS activity, expression, and localization in male and female spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol 298: R61–R69, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sullivan JC, Semprun-Prieto L, Boesen EI, Pollock DM, Pollock JS. Sex and sex hormones influence the development of albuminuria and renal macrophage infiltration in spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol 293: R1573–R1579, 2007 [DOI] [PubMed] [Google Scholar]
- 34.Tipton AJ, Baban B, Sullivan JC. Female spontaneously hypertensive rats have greater renal anti-inflammatory T lymphocyte infiltration than males. Am J Physiol Regul Integr Comp Physiol 303: R359–R367, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Verhagen AM, Attia DM, Koomans HA, Joles JA. Male gender increases sensitivity to proteinuria induced by mild NOS inhibition in rats: role of sex hormones. Am J Physiol Renal Physiol 279: F664–F670, 2000 [DOI] [PubMed] [Google Scholar]
- 36.Wang YR, Yen CH, Sun YF, Laun YT. Gender-dependent response in blood pressure changes following the inhibition of nitric oxide synthase. Chin J Physiol 46: 91–94, 2003 [PubMed] [Google Scholar]
- 37.Whitacre CC. Sex differences in autoimmune disease. Nature Immunol 2: 777–780, 2001 [DOI] [PubMed] [Google Scholar]
- 38.Wu Y, Huang A, Sun D, Falck JR, Koller A, Kaley G. Gender-specific compensation for the lack of NO in the mediation of flow-induced arteriolar dilation. Am J Physiol Heart Circ Physiol 280: H2456–H2461, 2001 [DOI] [PubMed] [Google Scholar]
- 39.Wuthrich RP. Vascular cell adhesion molecule-1 (VCAM-1) expression in murine lupus nephritis. Kidney Int 42: 903–914, 1992 [DOI] [PubMed] [Google Scholar]