

Significance
Adults and children conform to members of their social groups and expect others to do the same, but the earlier development of this expectation is unknown. Through looking-time experiments with 7- to 12-month-old infants, we show that preverbal infants expect members of social groups to act alike. Our findings further reveal that this expectation is specific to social groups, as infants did not expect similar actions from socially unrelated individuals or from groups of inanimate objects. Thus, an expectation that social affiliates will share behaviors arises early in human development, prior to language or to extensive experience with different social groups.
Keywords: social development, infancy
Abstract
The short ontogenetic time courses of conformity and stereotyping, both evident in the preschool years, point to the possibility that a central component of human social cognition is an early developing expectation that social group members will engage in common behaviors. Across a series of experiments, we show that by 7 months of age preverbal infants differentiate between actions by individuals that are and are not consistent with the actions of their social group members. Infants responded to group-inconsistent actions only in a social context: they failed to distinguish the same behavioral differences when presented with collections of nonsocial agents or inanimate objects. These results suggest that infants expect social group membership and behavior to covary, before extensive intergroup experience or linguistic input. This expectation is consistent with the socially motivated imitation and stereotyping evident in toddlers and preschoolers, and may play a role in the early emergence of one or both of these aspects of social behavior and cognition.
Human behavior is often tied to the social affiliations of the actor. These ties are evident both in the ways we choose to act and the way we predict and perceive others’ actions. Individuals are biased to conform to the conventions of their social groups, maintaining cultural variations in behavior even in the face of intergroup contact and migration (1, 2). More transiently, we tend to mimic the actions and expressions of our partners in social interaction, especially when we want to be liked (3, 4). Stereotypes lead us to expect that others will act like their own social affiliates as well. We perceive social groups as sharing behavioral tendencies and traits that we use to predict and explain the behavior of individual group members (5, 6).
Socially motivated imitation or conformity and stereotyping share an early developmental onset (7, 8). Many cultural behaviors, including social gestures and language, are acquired in the first few years, suggesting early biases for adopting the typical behaviors of one's own social group. In the laboratory, preschool-aged children adopt labels for objects endorsed by the majority of informants and conform to consensus perceptual judgments and actions, and they copy the actions of ingroup over outgroup members when group membership is conveyed by native vs. foreign speech (9–13). Children even show an understanding of group-based social norms in their reactions to others’ behavior, protesting when ingroup members, although not outgroup members, break conventions (14, 15). Stereotyping is similarly robust at early ages; within North American populations, for example, preschool children are influenced by the gender and racial stereotypes of their culture (7, 16). In fact, in laboratory studies young children often rely on social category membership more than older children or adults when making predictions about individuals (17–19).
Conformity, imitation, and stereotyping differ in some respects. Stereotypes are beliefs about stable attributes of group members, often thought to be inherent in the individual members themselves (19, 20). Although conformity can be constituted by long-term adherence to group norms, it also occurs in the context of temporary situations and transient judgments, as does socially motivated mimicry (3, 4, 21). Especially in these latter cases, the actions engaged in during conformity and imitation are not determined by the actor’s own traits or values but rather by the presence and actions of his social partners. Still, despite these differences, stereotyping and conformity both center on an expectation that social group members will or should act alike.
Understanding how children come to make these connections between social categories and common behavior will be critical in determining why human groups develop and maintain conventional behaviors and stereotypes. By the preschool years, parents and other adults have begun to instruct children in the conventional behavior of their own groups, and they talk about the behaviors of social groups in generic terms (22). Children may generalize from this input and come to expect that groups are likely to be characterized by particular behaviors and that adherence to those behaviors is normative (23). Children might also learn from interacting with others that conventional behaviors can be socially rewarding, correct, or useful. Such a trajectory of acquisition would imply that the social-learning mechanisms that underlie cumulative culture are themselves the product of learning from instruction, experience with the success of past learning strategies, or both (24). Alternatively, the early development of socially motivated imitation and stereotyping may be predicated on an inherent sensitivity to shared behavior as a feature of social group membership. The latter view predicts that even preverbal infants, presented with novel social groups, may expect the group members to engage in common actions.
One complication in designing an experiment to test for this expectation is that little is known about how, or even whether, infants represent social groups. Research on face perception has established that, by 4–6 months of age, infants categorize faces along a variety of dimensions, including sex, age, and race, and prefer to look at faces from more familiar instances of these categories (25–28). This research demonstrates infants’ ability to use perceptual information to distinguish some of the categories recognized by children and adults, but it is not clear whether infants endow these categories with social meaning. Although people can be categorized in many ways, we generally expect social groups to be characterized by affiliation between their members rather than solely by shared attributes (29). It is an open question whether infants under a year of age can represent such social relationships and use cues to affiliation to define social groups that can then be used in the service of inference about their members.
The current research simultaneously asks whether infants recognize social groups on the basis of affiliative cues and, if so, whether they expect social group members to engage in the same actions and make the same choices. Infants were presented with animated depictions of two novel social groups, delineated via the spatial proximity and synchronous movements of their members, and were provided with information about the covariation of behavior with group membership for two of the three members of each group. Then we used two preferential-looking methods—a violation of expectancy method (Experiments 1–4) and a habituation method (Experiment 5)—to probe whether infants expected the remaining individuals to engage in the same behavior as the other members of their groups.
To investigate whether infants' expectations of within-group consistency apply both to shared behavior and shared choices or preferences, separate experiments tested infants' inferences about groups that performed different actions (Experiment 1) and groups that directed similar actions toward different objects and locations (Experiments 2–5). To investigate whether infants' expectations are specific to social groups, we compared infants’ responses to social characters and interactions with responses to similar events presented in the context of groups of inanimate entities (Experiments 2 and 5) and of animate agents stripped of information about social relationships (Experiment 3). We tested for the role of shared appearance in establishing and maintaining social groups by varying the presence of appearance cues to group membership (Experiments 4 and 5). Across these experiments, we tested for the robustness of infants’ reasoning about the shared actions of social groups by testing infants' inferences at different ages and with different experimental designs.
Experiment 1
In Experiment 1, 12-month-old infants saw an animated display featuring two groups of identical geometrical figures, one consisting of three orange stars and the other of three purple trapezoids, each with a pair of eyes. The figures composing each group initially appeared in close proximity to one another in the top two corners of the screen, where they took turns “dancing”; the figures in each group made a series of synchronized, small, semicircular movements around a larger, circular path, producing a characteristic sound at six points throughout their path of motion (Fig. 1A and Movie S1).
Fig. 1.
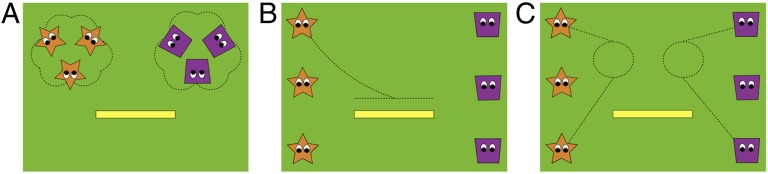
Scenes from the animation presented to infants in Experiment 1, including (A) the introductory sequence, (B) a familiarization trial featuring a sliding event, and (C) a group reminder scene. The dashed lines represent paths of motion.
Following this introductory sequence, the figures spread out down the sides of the screen, and infants saw three rounds of familiarization and test trials. The first round began with four familiarization trials featuring the top two figures from each group. In each trial, one figure moved toward and landed on a platform at the center of the screen and then either jumped up and down or slid back and forth three times, with action type covarying with group membership and accompanied by the group's characteristic sound (Fig. 1B). These familiarization trials were followed by two test trials in which the remaining figures, one from each group, performed the same action. Half of the participants saw both figures jump, whereas half saw both slide. For one figure, this action matched that of its group and this trial was thus designated as a “consistent” trial, whereas for the other figure the action contrasted with that of its group, resulting in an “inconsistent” trial. Familiarization and test trials were infant directed; after completing its motion, each figure remained stationary on the platform while the infant’s cumulative looking time to the display was measured until the infant looked away for 2 consecutive seconds.
The second and third rounds each consisted of two familiarization trials, one for each group, and a pair of consistent and inconsistent test trials. These trials maintained both the same group–action pairing and the identities of the consistent and inconsistent figures. The rounds were separated by a group reminder scene in which infants saw the test figure and a familiarization figure from each group move toward and circle one another and then return to their original locations (Fig. 1C).
To test whether group-inconsistent trials violated infants’ expectations regarding the figures’ behavior, we compared the percentages of looking time directed to the two types of test trials. A repeated-measures ANOVA including trial type (consistent vs. inconsistent) as a within-subject factor and test order (inconsistent trial first vs. second), and action assignment (which group jumped and which slid during familiarization) as between-subjects factors found only a significant main effect of trial type [F(1,20) = 8.461, P < 0.01]. Infants looked longer at the inconsistent trials (55.0%) than the consistent trials (45.0%; Fig. 2; analyses were initially performed with sex as a between-subjects factor, but it was removed upon confirmation that there were no main effects or interactions involving this variable).
Fig. 2.

Looking preferences for consistent and inconsistent events in Experiments 1–4. Infants looked significantly longer at the inconsistent trials in both Experiment 1 (n = 24) and the animate condition of Experiment 2 (n = 24), where groups were defined by social affiliation among similar-looking, animate individuals (*P < 0.05). In contrast, looking to consistent and inconsistent trials did not differ when groups were composed of inanimate entities (Experiment 2, inanimate, n = 24) or animate but socially unrelated characters (Experiment 3, n = 24). In Experiment 4 (n = 16), when social but no featural cues to group membership were available, infants differentiated between trial types but looked significantly longer at consistent trials. Error bars represent SEM.
These results provide initial evidence that infants use social groups as the basis for predicting the action of individual group members, even when the infants themselves are not members of either social group. Nevertheless, they also raise immediate questions. First, did infants perceive the characters in the events as animate and social, or did they simply categorize them as objects of particular shapes and colors and learn their feature–action relationships? Second, if infants indeed perceived the characters as animate and expected them to engage in the same actions as members of their social group, at what level did they expect such group consistency? Do infants expect consistency only at the level of action, or do they, like adults, expect groups to be homogeneous at the level of attitudes, preferences, or goals? Third, can such expectations be observed earlier in the first year of life? Experiment 2 addressed these questions.
Experiment 2
In Experiment 2, 8-month-old infants saw an animated display again featuring two groups of identical geometrical figures, here consisting of three red circles and three yellow triangles. The platform in the middle of the screen was replaced by two stationary, distinctively colored, and patterned boxes. One-half of the participants were assigned to an “animate” condition, in which the figures had eyes like those in Experiment 1 (Fig. 3A), and one-half to an “inanimate” condition, in which the shapes composing the figures’ eyes were rearranged into a symmetrical pattern that did not resemble a face (Fig. 3B). The structure of the experiment—an introductory sequence, followed by three rounds of infant-directed familiarization trials and test trials, separated by group reminder scenes—was the same for both conditions, and mirrored the chain of events in Experiment 1.
Fig. 3.

Depictions of the introductory scenes for (A) Experiment 2, animate condition; (B) Experiment 2, inanimate condition; (C) Experiment 3; and (D) Experiment 4.
The animate condition began with the same introductory sequence as Experiment 1, with the groups of figures taking turns dancing in synchrony. The members of each group then spread out on the sides of the screen, and in each of the subsequent familiarization trials one of the top two figures from each group moved toward, circled around, and landed on one of the two central boxes. Because all four characters engaged in the same circling motion, the landing location was the key variable, with figures belonging to one group landing on the blue box on the left, and figures belonging to the other group landing on the purple box on the right (pairings of groups and boxes were counterbalanced across infants). The test trials featured the two remaining figures, one from each group, both approaching, circling, and landing on the same box (either left for both or right for both). For one figure this landing location matched that of its group (consistent trial), whereas for the other figure it did not (inconsistent trial). The group reminder scenes were the same as in Experiment 1 (Movie S2).
The inanimate condition differed from the animate condition in the appearance of the figures and in the events presented in the introductory sequence and group reminder scenes, which were modified to remove all impression of animate characters engaged in a social interaction. During the introductory sequence, these figures moved in straight paths at constant speeds along a triangular frame (Fig. 3B), producing a characteristic sound at six points of motion. The group reminder scenes still featured the test figure and a familiarization figure from each group moving toward one another, but in this case the motion was initiated by pokers that appeared briefly in each corner of the screen, and the figures bounced off of one another rather than circling each other. The events in the familiarization and test trials were identical to those used in the animate condition, with familiarization trials in which figures from the different groups landed on different boxes and test trials that depicted both consistent and inconsistent box-landing events (Movie S3).
Percentages of looking time to consistent and inconsistent trials were analyzed with a repeated-measures ANOVA including condition (animate vs. inanimate), test order (inconsistent first vs. second), and familiarization distance (which of the two boxes each group landed on) as between-subjects factors. A significant condition by trial type interaction [F(1,40) = 6.515, P < 0.05] indicated that assignment to either the animate or inanimate condition affected infants’ relative responses to the consistent and inconsistent trials. Paired-sample t tests revealed that infants in the animate condition, like those in Experiment 1, looked significantly longer to inconsistent trials (56.5% of looking time) than to consistent trials [43.5%; t(23) = 2.87, P < 0.01; this difference was also reflected in a strong trend toward a main effect of trial type in the ANOVA, F(1,40) = 3.974, P = 0.053]. In contrast, for infants in the inanimate condition the difference between looking times to inconsistent (49.3%) and consistent trials (50.7%) was not statistically significant [t(23) = 0.33, P > 0.7; Fig. 2].
Might the interaction between the conditions stem from a tendency for participants in the animate condition to pay better attention to the events? To test this possibility, we compared the participants’ average looking time to all familiarization trials in the animate condition [mean (M): 9.9 s] to that observed in the inanimate condition (M: 8.5 s), and found that these times did not differ [t(46) = 1.03, P > 0.3]. Average looking time to both test trial types in the inanimate condition (inconsistent: 5.9 s; consistent: 6.4 s) was also similar to that observed for the consistent trials in the animate condition (6.4 s), whereas looking time to the inconsistent trials of the animate condition was higher (7.6 s).
As in Experiment 1, the results of the animate condition are consistent with the possibility that infants use social group membership to infer the likely actions of individuals, given information about the typical behavior of other group members. Such a hypothesis can explain why inconsistent trials violated their expectations, resulting in longer looking times to those trials than to the consistent trials. In addition, these data help to address some of the questions raised by Experiment 1 by showing that these category-based inferences begin earlier in the first year of life and apply not only to specific actions but to the goals or outcomes of those actions. Although the two groups of figures in Experiment 1 each engaged in a different action, those in Experiment 2 all engaged in the same action of circling around and landing on a box. All that distinguished the groups in the latter experiment was the identity and location of the box that each figure chose to circle around. This distinction was sufficient to establish an expectation that individuals would behave like those in their own group.
The findings of Experiment 2 also suggest that infants’ longer looking time to the inconsistent trials cannot be explained by a learned association between the appearances of the figures and the properties of the actions. Although the two conditions involved the same familiarization and test events, and infants in both conditions thus experienced the same feature–motion correlations (e.g., the tendency of red circles to land on the left box), the change in context in the inanimate condition eliminated infants’ tendency to use this correlation to infer the likely actions of additional group members at test. This interaction between the two conditions thus begins to suggest that expectations of behavioral consistency are confined to social beings and their interactions.
Nevertheless, one may question whether infants' behavioral inferences are confined to social categories. The two conditions of Experiment 2 were designed to differ not only in the presence of social affiliations between the figures but also in cues to the figures' animacy. Infants’ expectations of shared behavior therefore might have resulted from reasoning about animate categories formed on the basis of the perceptual similarity of the figures, with no appeal to social information. To test whether cues to animacy are sufficient to establish categories that will support the sorts of behavioral inferences seen above, Experiment 3 investigated whether infants use animate categories to guide expectations about behavior when the animate characters do not interact socially.
Experiment 3
In Experiment 3, a new group of 8-month-old infants was presented with the same two sets of identical figures used in the animate condition of Experiment 2, after eliminating the key indicators of a social relationship between the group members: spatial proximity and synchronized, dancing motions. Instead of appearing in separate, proximal groups, the three figures of each type were interspersed in a horizontal row across the screen (Fig. 3C). Each figure moved individually, traveling out and back along a curved path similar to the paths taken by each individual in a group in Experiments 1 and 2, and making the same sounds used in those experiments (Movie S4). Infants then saw the same type of familiarization trials (featuring the two leftmost figures of each kind) and test trials (featuring the rightmost figures of each kind) as in Experiment 2, but without the group reminder scenes, as there were no social relationships to reemphasize between test trial pairs.
Looking times to consistent and inconsistent test trials were coded as in Experiment 2. A repeated-measures ANOVA including trial type as a within-subject factor and familiarization and test order as between-subjects factors found no significant main effects or interactions (all P > 0.4). The percentages of time infants spent looking to consistent (50.7%) and inconsistent (49.3%) trials were similar (Fig. 2). A 2 (trial type) × 2 (condition) ANOVA comparing Experiment 3 to the animate condition from Experiment 2 revealed a significant trial type by condition interaction [F(1,44) = 4.434, P < 0.05]. Infants showed a reliably greater looking preference for the inconsistent test events in the animate, social condition of Experiment 2 than in the animate, nonsocial condition of Experiment 4.
The results of Experiment 3 suggest that animate motion, facial features, and similarity of appearance together are not sufficient to lead infants to expect that individuals will make the same behavioral choices. Without the cues to social affiliation present in Experiments 1 and 2, infants’ looking times no longer differentiated the consistent and inconsistent trials. One caveat, however, is that infants may have had a more difficult time representing similar looking individuals as a coherent category when they were not grouped together in proximity. Given that proximity is likely to be a potent cue that individuals are socially related, it cannot be equated across social and nonsocial conditions involving animate objects (30, 31), so one goal of the next experiments was to test further whether social information is central to the expectations of behavioral consistency demonstrated in Experiments 1 and 2. Experiment 4 asked whether social information alone would be sufficient for infants to delineate groups and evaluate behavioral consistency, even in the absence of any shared perceptual features.
Of course, even if infants’ expectations of behavioral consistency are specific to the social domain, they could fail to expect consistency among members of diverse groups. Despite being insufficient, similarities in appearance may be necessary to support infants’ use of affiliative cues to parse individuals into social groups. Indeed, several developmental theories assert that children’s initial social group representations depend on salient visual differences between members of contrasting groups (7, 32, 33). For example, social group representations have been proposed to build on a preexisting propensity to distinguish biological kinds (32, 33). In contrast to such views, others have argued that early developing social group representations are at their core representations of affiliation between individuals (29, 34). Experiment 4 addresses this controversy, in addition to the question of whether infants’ expectations of behavioral consistency center on animate or on social kinds.
Experiment 4
Experiment 4 investigated whether 8-month-old infants would distinguish consistent and inconsistent behaviors of individuals whose social groups were not visually marked. It presented the same social information as Experiment 2, with six unique characters. The individuals making up the two groups were all circular figures with eyes, but each was distinctive in coloring, mouth shape, and hairstyle (Fig. 3D). The events were the same as those presented in the animate condition in Experiment 2, beginning with the introductory sequence that used proximity and synchronous dancing to establish two different social groups, each composed of three members, and continuing with four familiarization trials that demonstrated which box members of each group typically chose to land on, as in the preceding experiments. These events were followed by a pair of consistent and inconsistent test trials, and then a group reminder scene, followed by two more familiarization and two more test trials (Movie S5). The third round of familiarization and test trials was omitted in this experiment.
A repeated-measures ANOVA comparing percentage of looking time to consistent vs. inconsistent trials, with test order and familiarization distance as between-subjects factors, found a significant main effect of trial type [F(1,12) = 5.675, P < 0.05]. In contrast to the results in Experiment 3, infants presented with groups delineated by social cues alone distinguished between the consistent and inconsistent trials. An examination of the mean looking times, however, showed that the infants looked longer to the consistent trials (57.2%) rather than to the inconsistent trials (42.8%; Fig. 2). This pattern differed significantly from that observed in the social conditions of Experiments 1 and 2, in which infants looked longer to inconsistent trials [F(1,62) = 17.078, P < 0.001].
The finding that infants differentiated the consistent and inconsistent trials suggests that social cues are sufficient for the formation of group representations that can support behavioral inferences. If infants had not maintained some representation of which individuals had interacted with one another, or if they did not consider this information relevant for evaluating the later actions of the individuals, then they would have had no basis on which to differentiate the consistent and inconsistent test events, as each featured the same behavior. Moreover, if infants had expected that perceptually dissimilar individuals would act differently, then again they would have had no basis for differentiating between the test displays, as each test character differed in appearance from the members of both groups.
In combination with the results of Experiment 3, these findings support the hypothesis that it is social groups, rather than animate categories or shared visual features more broadly, that underlie early category-based behavioral inferences of the sort seen in Experiments 1 and 2. Nevertheless, the reversal of the direction of the effect in Experiment 4 raises questions. Although the canonical hypothesis in looking time studies is that infants will look longer at novel events, many experiments have documented situations in which infants show the opposite preference. Preferential looking to familiar or expected event outcomes often occurs in contexts of greater complexity or with younger infants who presumably have more difficulty processing and extracting regularities in the events (e.g., refs. 35–37). In a recent attempt to provide a comprehensive explanation for infants' familiarity and novelty preferences, Kidd et al. (37) argued that infants’ looking is guided by interest in the most informative events and displays. When events presented earlier in a study lead infants to high-confidence predictions or expectations for subsequent events, then events that match this schema offer little new information. However, when preceding information leads to lower confidence predictions or less specific inferences, the more familiar or expected test event may in fact be more informative to the infant, insofar as it helps to confirm a cohesive view of the events in question.
In the current experiment, infants may not have been as certain about the social relationships between individuals as they were in the previous experiments. The appearance cues likely helped infants to maintain a representation of the social ties between individuals established in the introductory sequence—even adults have an easier time encoding social relationships when they are marked by a visual cue (38)—and they may even have provided information regarding affiliation when accompanied by the other social cues present in Experiments 1 and 2. Thus, in the current experiment, the behavioral consistency exhibited at test may have been more informative for infants, attracting more attention as it reconfirmed their representations of the social relationships among individuals by showing that they did, indeed, act alike. Nevertheless, it is important to determine whether the present findings are robust. To this end, Experiment 5 was undertaken to replicate and extend the findings of Experiments 2 and 4, with somewhat younger infants and a modified experimental design, to test further whether infants expect social group members to act alike and under what circumstances they respond by looking longer at individuals whose actions are inconsistent with their groups.
Experiment 5
Experiment 5 repeated several conditions from the earlier experiments with a younger age group and a method that increased infants' exposure to each group’s typical behavior. Instead of using an expectancy violation method with just three familiarization trials, the experiment used a habituation method, displaying the familiarization events repeatedly until infants' looking time declined to one-half of its initial level. The experiment consisted of three conditions that aimed to replicate the results of both the animate and inanimate conditions from Experiment 2 as well as the unexpected finding, from Experiment 4, that infants presented with social groups lacking any correlated visual attributes looked longer to consistent actions. Because the figures comprising each group in the replication of the animate condition from Experiment 2 were visually identical, we refer to that condition here as the “homogeneous group” condition, whereas the replication of Experiment 4, where members of a group could not be identified by their visual appearance, is referred to as the “heterogeneous group” condition. The inanimate condition retains the same label.
The displays in each of these conditions were altered to consist of a single habituation phase, followed by two pairs of test trials. This change in design resulted in the elimination of the group reminder scenes used in earlier experiments. The use of habituation also addressed the possibility that infants in the inanimate condition might come to generate expectations regarding the figures’ behavior if simply given more evidence.
Seven-month-old infants were assigned to one of the three conditions. Displays in the homogeneous group condition (Movie S6), the inanimate condition (Movie S7), and the heterogeneous group condition largely replicated those in the animate and inanimate conditions of Experiment 2 and in Experiment 4, respectively. Each began with the same introductory sequence used in the relevant previous experiment, either synchronous dancing or synchronous movement around a frame. Then, in all three conditions, infants saw up to 12 familiarization trials in a row, again featuring two out of three members of each group circling and landing on one of the two center boxes. Presentation of familiarization trials was halted if a participant reached a criterion of looking less than one-half the amount of time to the last three trials that they looked to the first three. This habituation phase was followed by two consecutive pairs of consistent and inconsistent test trials, featuring the final figure from each group. The number of infants who reached the habituation criterion did not differ significantly across conditions [homogeneous: 13 of 16; inanimate: 10 of 16; heterogeneous: 12 of 16; Kruskal–Wallis test: H(2) = 0.86, P > 0.6], nor did the average number of habituation trials presented [homogeneous: 8.4; inanimate: 8.5; heterogeneous: 7.7; Kruskal–Wallis test: H(2) = 0.87, P > 0.6].
Looking times to the test trials were coded and analyzed in the same manner as in the previous experiments. A repeated-measures ANOVA including trial type as a within-subject factor and condition, test order, and familiarization distance as between-subjects factors found a significant interaction between trial type and condition [F(1,36) = 6.869, P < 0.01]. Using paired-sample t tests to compare proportional looking time to the different trial types within each condition, we replicated our previous results in each case (Fig. 4). (Tests were one-tailed for the homogeneous group and heterogeneous group conditions, where previous results led to a priori predictions regarding the expected direction of the difference between conforming and nonconforming trials, and two-tailed for the inanimate condition where there was no reason to expect any difference between trial types to be more likely in one direction than the other.) Infants in the homogeneous group condition looked significantly longer to inconsistent trials (60.1% of looking time) than to consistent trials [39.9%; t(15) = 4.41, P < 0.001]. For infants in the inanimate condition, the difference between looking times to inconsistent (49.4%) and consistent trials (50.6%) was not statistically significant [t(15) = 0.18, P > 0.8; the interaction between these two conditions alone, as seen in Experiment 2, was also replicated, F(1,30) = 6.886, P < 0.05]. Finally, infants in the heterogeneous group condition looked significantly longer to consistent trials (56.9%) than to inconsistent trials [43.1%; t(15) = 2.28, P < 0.05]. These results thus replicate the findings from Experiments 2 and 4 and provide evidence that 7-month-old infants consistently attend to the similarity or dissimilarity of behaviors engaged in by members of the same social group.
Fig. 4.

Looking preferences for consistent and inconsistent events in Experiment 5 (n = 48). Infants significantly differentiated between these events when groups were composed of animate, socially affiliated individuals [who either shared visual features (homogeneous groups) or shared no such features (heterogeneous groups)], but not when they were composed of inanimate entities (*P < 0.05). Error bars represent SEM.
The successful replication of the heterogeneous condition in Experiment 5 confirms that infants track behavioral consistency among social group members even when there are no visual cues to group membership. However, it does not provide a conclusive answer as to why the direction of looking preference reverses in the absence of visual cues. To provide positive evidence for the argument that these stimuli are more difficult to process, one would want to show that manipulations ameliorating that difficulty shift looking times back in the direction of longer looks to inconsistent events. We attempted to provide such evidence in two ways. First, we replicated Experiment 4 with a group of older infants, who should be better able to maintain representations of the social ties between the disparate individuals composing each group (35). Second, we tested a new group of 8-month-old infants using the displays and method of the heterogeneous condition of Experiment 4, but keeping proximity as a cue to social group membership not just during the introduction, but throughout the familiarization and test trials. In both cases, the changes produced significant shifts in the direction of infants' test trial looking preferences, toward longer looking to inconsistent trials, compared with those seen in Experiment 4 (Text S1). Thus, shared appearance cues help infants to keep track of social group relations when other cues to social affiliation are absent, rendering the homogeneous displays of Experiments 1, 2, and 5 easier to process than the heterogeneous displays. Nevertheless, shared appearance is neither sufficient (in Experiment 3) nor necessary (in Experiments 4 and 5) to establish social group relations.
Discussion
From a young age, children acquire stereotypes about social groups and are motivated to conform to members of their ingroup and of the majority consensus (7–19). Both of these aspects of social cognition rely on an expectation that social group members act alike. One of the major questions regarding the ontogeny of this expectation is whether children learn to use group membership to predict or guide behavior once they begin interacting broadly with others, or whether this expectation can emerge before substantial intergroup experience or linguistic input.
The results of the present experiments provide evidence supporting the latter view. We find that infants possess an early-developing expectation that social group members will or should act in similar ways, before they are able to engage in robust imitation of others or discussion of stereotypes. Infants in the second half of the first year of life expect that members of the same social group will both engage in similar actions and make similar choices. Affiliative cues to the existence of social relationships between animate figures were both necessary and sufficient to prompt infants to differentiate instances of behavioral consistency and inconsistency by those figures. These affiliative cues were only effective within the animate domain, as infants failed to show the same expectation of shared behavior when they were presented with the same proximity and synchrony cues in the context of inanimate figures (Experiments 2 and 5). The results also showed that visual correlates of category membership play a similar role for infants as they do for adults—such cues are neither necessary (Experiment 4) nor sufficient (Experiment 3) to prompt representations of social groups. Nevertheless, the removal of visual cues to social group membership produced a reversal in infants' looking preferences (Experiments 4 and 5), suggesting that visual cues substantially amplify the ability to identify and learn about groups of socially interacting individuals.
Together, these experiments provide evidence that infants represent the social relationships of others and that such representations drive domain-specific expectations of shared behavior, but more evidence is needed to link these expectations to stereotyping, conformity, or socially motivated imitation in children and adults. Stereotyping and conformity differ in their particulars. The application of stereotypes does not depend on the presence of additional group members, for example, and stereotypical behavior is assumed to spring from stable, inherent characteristics, perhaps a group essence, present in each individual member (19, 20). In contrast, to conform to or imitate group members, an actor must witness or otherwise be informed of the behavior or norm to which she is conforming. Although our findings reveal that infants expect group members to act alike, they do not reveal whether infants attribute either enduring, shared characteristics, or momentary perceptions, to such members. These are topics for further research.
In addition to further characterizing infants’ expectations about others’ group-consistent behavior, future research should also assess whether these expectations play any role in infants’ own behavior. Some evidence suggests that, like young children, infants aged 10–14 months adopt the behaviors of linguistic ingroup members, trying foods they endorse and imitating their actions (39–41). Past work on infants’ perception of and engagement in dyadic imitation suggests they see imitation as social and communicative (42): infants imitate actions they have seen in the presence of the person who modeled them but not in the presence of a new person, and they also use imitation to identify potential social partners (43, 44). Both the sociocommunicative nature of dyadic imitation and the early selection of ingroup members as behavioral models could relate to infants' expectation, demonstrated here, that social affiliates act alike. To date, however, no research links these phenomena.
Our findings have implications for other topics regarding the development of intergroup cognition. For example, one open question is whether early representations of social groups are initially organized dichotomously as “us” vs. “them,” without acknowledgment of outgroup divisions. The current research suggests that even infants in the first year of life can segregate others according to nonoverlapping social relationships in which the infants themselves have no role. This research also bears on questions concerning the nature of social categories. Although stereotypes associating various social groups with particular behaviors or traits seem similar to other sorts of category-based knowledge, there are findings, including those of neural specificity, that suggest cognitive processes that are devoted specifically to social categories (45, 46). The developmental pattern of selective inference demonstrated here provides suggestive evidence that even infants may engage in categorical reasoning processes that are specific to the social domain. More research is needed, however, to probe convergences and divergences between infants' reasoning about animate beings as agents, who act instrumentally on the world of objects, and as social partners, who engage with other animate beings by affiliating with them, communicating with them, and conforming to their actions and gestures.
A final unanswered question concerns the earlier development of the expectations of social group consistency that our studies reveal. It is possible that these expectations develop independently of any social experiences. Alternatively, the expectations that we find at 7 months and at older ages may be shaped by infants' earlier social and behavioral experiences interacting with others, as well as by their experiences observing the social interactions of the people around them. Experiments with younger infants are needed to distinguish these possibilities.
Over the last two decades, developmental psychologists have capitalized on the insights of the cognitive revolution, pursuing an understanding of the origins and ontogeny of our capacities to form representations in domains such as those of objects, space, causality, and number. These investigations have repeatedly revealed that central features of human cognition are evident in infancy and that studies of infants can shed light on why adults reason as we do. Social psychologists have an even longer tradition of studying the role of cognition in behavior, acknowledging since the middle of last century, for example, the critical role that categorization plays in our beliefs, attitudes, and actions toward others (5, 47). However, developmental psychologists have barely begun to investigate the extent to which the social cognitive capacities that guide adults are present in infancy and whether features of such nascent social cognition persist into adulthood (48, 49). Here, we report evidence that treating individuals as instances of social categories, one of the fundamental aspects of adult social reasoning (6), begins before humans can talk or tie their shoes and before most infants have even encountered a social group other than their families. By building on this foundation to characterize the origins of social cognition in infancy and its subsequent development, studies of young children may cast new light on how and why adults engage with others as we do.
Methods
Participants were full-term infants recruited from the greater Boston area (Experiment 1: n = 24, 10 females; mean age, 11 months, 28 d; range, 11:16–12:15; Experiment 2: n = 48, 24 females; mean age, 8:3; range, 7:17–8:15; Experiment 3: n = 24, 11 females; mean age, 7:29; range, 7:17–8:15; Experiment 4: n = 16, 8 females; mean age, 8:3; range, 7:17–8:15; Experiment 5: n = 48, 21 females; mean age, 7:0; range, 6:16–7:14). Parents were given travel reimbursement, and participants received a toy or other small reward for participating. Recruitment and experimental procedures were approved by the Committee on the Use of Human Subjects at Harvard University, and parents gave informed consent prior to their children's participation.
All displays were projected onto a 3.5′ × 4.5′ screen. Animations were created using Keynote ’09 software and featured figures ranging from 5.5” to 7” in width when projected onto the screen. Infants viewed the animations while seated on their parent’s laps, ∼60” from the screen, and parents were instructed to close their eyes for the duration of the experiment. Online blind coders viewed infants via a video feed from a camera positioned directly below the screen and continuously coded infants’ looking toward and away from the screen throughout the experiment.
In all experiments in which the figures moved synchronously as groups during the introductory sequence, the groups took turns following an ABBAAB pattern, with the order of the first group counterbalanced across infants. In Experiment 3, in which the introductory sequence featured individual movements, the figures took turns in sequential order according to their horizontal alignment across the screen, moving first left to right, then right to left, then left to right again, or vice versa. In all experiments, familiarization and test trials alternated between groups, with the order of the two groups' actions, as well as which action or location was paired with a particular group, counterbalanced across infants. The identities of the figures that performed the consistent and inconsistent actions were held constant across test pairs for a given participant but were counterbalanced across participants.
Before beginning data collection, we decided on a minimum of 0.5 s and a maximum of 120 s of cumulative looking per trial, and a 2-s continuous look away from the screen as the threshold for ending a trial. The experimenter controlling the animation also initiated recording of looking time to familiarization, habituation, and test trials on the basis of the sounds indicating the beginning of a jumping or sliding action (Experiment 1) or the landing of a figure on a box (Experiments 2–4). For Experiments 1 and 2, this experimenter could see the events of the animation on the laptop screen as the infant viewed them on the projection screen, but for Experiments 3–5 we covered the laptop screen so that this experimenter was also blind to the events being viewed by participants during data collection. Looking time software collecting data from the online coder(s) in the next room indicated when the experimenter should move on to the next trial. When possible, there were two online coders; videos of additional participants were recoded offline by two coders simultaneously to further confirm reliability. Over 25% of participants were double-coded, and agreement was over 94%.
For each pair of test trials, we calculated looking time to the consistent and inconsistent trials as a percentage of the total looking time to that test pair, and then we averaged across test pairs to give the mean percentage of looking time to each trial type. If either trial in a pair failed to meet the criteria for inclusion (e.g., if cumulative looking time was less than 0.5 s or if participants in Experiments 2–5 were looking away as the figure in the trial landed on the relevant box), then the pair of trials was excluded. Participants with two or more invalid test pairs were designated as inattentive and excluded from the study. Additional participants also were excluded for fussiness or experimenter error (excluded participant tallies: Experiment 1, one fussiness; Experiment 2, six inattentiveness, four fussiness, two experimental error; Experiment 3, three inattentiveness, two fussiness; Experiment 4, 0; Experiment 5, five inattentiveness, seven fussiness, three experimental error).
Supplementary Material
Acknowledgments
We thank the Harvard Laboratory for Developmental Studies and the families who participated. This research was funded by National Institutes of Health Grant 5R01HD023103 (to E.S.S.).
Footnotes
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1304326110/-/DCSupplemental.
References
- 1.Boyd R, Richerson PJ. Culture and the Evolutionary Process. Chicago: The Univ of Chicago Press; 1985. [Google Scholar]
- 2.Henrich J, Boyd R. The evolution of conformist transmission and the emergence of between-group differences. Evol Hum Behav. 1998;19(4):215–241. [Google Scholar]
- 3.Chartrand TL, Bargh JA. The chameleon effect: The perception-behavior link and social interaction. J Pers Soc Psychol. 1999;76(6):893–910. doi: 10.1037//0022-3514.76.6.893. [DOI] [PubMed] [Google Scholar]
- 4.Lakin JL, Chartrand TL, Arkin RM. I am too just like you: Nonconscious mimicry as an automatic behavioral response to social exclusion. Psychol Sci. 2008;19(8):816–822. doi: 10.1111/j.1467-9280.2008.02162.x. [DOI] [PubMed] [Google Scholar]
- 5.Allport GW. The Nature of Prejudice. Cambridge, MA: Addison-Wesley Publishing; 1954. [Google Scholar]
- 6.Macrae CN, Bodenhausen GV. Social cognition: Thinking categorically about others. Annu Rev Psychol. 2000;51:93–120. doi: 10.1146/annurev.psych.51.1.93. [DOI] [PubMed] [Google Scholar]
- 7.Aboud FE. Children & Prejudice. Cambridge, MA: Basil Blackwell; 1988. [Google Scholar]
- 8.Over H, Carpenter M. Putting the social into social learning: Explaining both selectivity and fidelity in children’s copying behavior. J Comp Psychol. 2012;126(2):182–192. doi: 10.1037/a0024555. [DOI] [PubMed] [Google Scholar]
- 9.Corriveau KH, Fusaro M, Harris PL. Going with the flow: Preschoolers prefer nondissenters as informants. Psychol Sci. 2009;20(3):372–377. doi: 10.1111/j.1467-9280.2009.02291.x. [DOI] [PubMed] [Google Scholar]
- 10.Corriveau KH, Harris PL. Preschoolers (sometimes) defer to the majority in making simple perceptual judgments. Dev Psychol. 2010;46(2):437–445. doi: 10.1037/a0017553. [DOI] [PubMed] [Google Scholar]
- 11.Walker MB, Andrade MG. Conformity in the Asch task as a function of age. J Soc Psychol. 1996;136(3):367–372. doi: 10.1080/00224545.1996.9714014. [DOI] [PubMed] [Google Scholar]
- 12.Haun DBM, Tomasello M. Conformity to peer pressure in preschool children. Child Dev. 2011;82(6):1759–1767. doi: 10.1111/j.1467-8624.2011.01666.x. [DOI] [PubMed] [Google Scholar]
- 13.Kinzler KD, Corriveau KH, Harris PL. Children’s selective trust in native-accented speakers. Dev Sci. 2011;14(1):106–111. doi: 10.1111/j.1467-7687.2010.00965.x. [DOI] [PubMed] [Google Scholar]
- 14.Rakoczy H, Warneken F, Tomasello M. The sources of normativity: Young children’s awareness of the normative structure of games. Dev Psychol. 2008;44(3):875–881. doi: 10.1037/0012-1649.44.3.875. [DOI] [PubMed] [Google Scholar]
- 15.Schmidt MFH, Rakoczy H, Tomasello M. Young children enforce social norms selectively depending on the violator’s group affiliation. Cognition. 2012;124(3):325–333. doi: 10.1016/j.cognition.2012.06.004. [DOI] [PubMed] [Google Scholar]
- 16.Williams JE, Bennett SM, Best DL. Awareness and expression of sex stereotypes in young children. Dev Psychol. 1975;11(5):635–642. [Google Scholar]
- 17.Diesendruck G, HaLevi H. The role of language, appearance, and culture in children’s social category-based induction. Child Dev. 2006;77(3):539–553. doi: 10.1111/j.1467-8624.2006.00889.x. [DOI] [PubMed] [Google Scholar]
- 18.Berndt TJ, Heller KA. Gender stereotypes and social inferences: A developmental study. J Pers Soc Psychol. 1986;50(5):889–898. [Google Scholar]
- 19.Taylor M, Rhodes M, Gelman SA. Boys will be boys; cows will be cows: Children’s essentialist reasoning about human and animal development. Child Dev. 2009;80(2):461–481. doi: 10.1111/j.1467-8624.2009.01272.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yzerbyt V, Rocher S, Schadron G. In: The Social Psychology of Stereotyping and Group Life. Spears R, Oakes PJ, Ellemers N, Haslam SA, editors. Malden, MA: Blackwell Publishing; 1997. pp. 20–50. [Google Scholar]
- 21.Asch SE. Studies of independence and conformity: A minority of one against a unanimous majority. Psychol Monogr. 1956;70(9):1–70. [Google Scholar]
- 22.Gelman SA, Taylor MG, Nguyen S. Mother-child conversations about gender: Understanding the acquisition of essentialist beliefs. Monogr Soc Res Child Dev. 2004;69(1):1–142. [Google Scholar]
- 23.Rhodes M, Leslie SJ, Tworek CM. Cultural transmission of social essentialism. Proc Natl Acad Sci USA. 2012;109(34):13526–13531. doi: 10.1073/pnas.1208951109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Heyes C. Grist and mills: On the cultural origins of cultural learning. Philos Trans R Soc Lond B Biol Sci. 2012;367(1599):2181–2191. doi: 10.1098/rstb.2012.0120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Quinn PC, Yahr J, Kuhn A, Slater AM, Pascalils O. Representation of the gender of human faces by infants: A preference for female. Perception. 2002;31(9):1109–1121. doi: 10.1068/p3331. [DOI] [PubMed] [Google Scholar]
- 26.Ramsey JL, Langlois JH, Hoss RA, Rubenstein AJ, Griffin AM. Origins of a stereotype: Categorization of facial attractiveness by 6-month-old infants. Dev Sci. 2004;7(2):201–211. doi: 10.1111/j.1467-7687.2004.00339.x. [DOI] [PubMed] [Google Scholar]
- 27.Kelly DJ, et al. Three-month-olds, but not newborns, prefer own-race faces. Dev Sci. 2005;8(6):F31–F36. doi: 10.1111/j.1467-7687.2005.0434a.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bar-Haim Y, Ziv T, Lamy D, Hodes RM. Nature and nurture in own-race face processing. Psychol Sci. 2006;17(2):159–163. doi: 10.1111/j.1467-9280.2006.01679.x. [DOI] [PubMed] [Google Scholar]
- 29.Shutts K, Pemberton CK, Spelke ES. Children’s use of social categories in thinking about people and social relationships. J Cogn Dev. 2013;14(1):35–62. doi: 10.1080/15248372.2011.638686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Campbell DT. Common fate, similarity, and other indices of the status of aggregates of persons as social entities. Behav Sci. 1958;3(1):14–25. [Google Scholar]
- 31.Hamilton DL, Sherman SJ, Lickel B. In: Intergroup Cognition and Intergroup Behavior. Sedikides C, Schopler J, Insko CA, editors. Mahwah, NJ: Psychology Press; 1998. pp. 47–74. [Google Scholar]
- 32.Hirschfeld LA. Race in the Making: Cognition, Culture and the Child’s Construction of Human Kinds. Cambridge, MA: MIT; 1996. [Google Scholar]
- 33.Medin DL, Atran S. The native mind: Biological categorization and reasoning in development and across cultures. Psychol Rev. 2004;111(4):960–983. doi: 10.1037/0033-295X.111.4.960. [DOI] [PubMed] [Google Scholar]
- 34.Rhodes M. Naïve theories of social groups. Child Dev. 2012;83(6):1900–1916. doi: 10.1111/j.1467-8624.2012.01835.x. [DOI] [PubMed] [Google Scholar]
- 35.Hunter MA, Ames EW. A multifactor model of infant preferences for novel and familiar stimuli. Adv Infancy Res. 1988;5:69–95. [Google Scholar]
- 36.Roder J, Bushnell EW, Sasseville AM. Infants’ preferences for familiarity and novelty during the course of visual processing. Infancy. 2000;1(4):491–507. doi: 10.1207/S15327078IN0104_9. [DOI] [PubMed] [Google Scholar]
- 37.Kidd C, Piantadosi ST, Aslin RN. The Goldilocks effect: Human infants allocate attention to visual sequences that are neither too simple nor too complex. PLoS One. 2012;7(5):e36399. doi: 10.1371/journal.pone.0036399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kurzban R, Tooby J, Cosmides L. Can race be erased? Coalitional computation and social categorization. Proc Natl Acad Sci USA. 2001;98(26):15387–15392. doi: 10.1073/pnas.251541498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kinzler KD, Dupoux E, Spelke ES. The native language of social cognition. Proc Natl Acad Sci USA. 2007;104(30):12577–12580. doi: 10.1073/pnas.0705345104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Shutts K, Kinzler KD, McKee CB, Spelke ES. Social information guides infants’ selection of foods. J Cogn Dev. 2009;10(1–2):1–17. doi: 10.1080/15248370902966636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Buttelmann D, Zmyj N, Daum M, Carpenter M, Selective imitation of in-group over out-group members in 14-month-olds. Child Dev 84(2):422–428. [DOI] [PubMed]
- 42. Meltzoff AN (2011) The Wiley-Blackwell Handbook of Childhood Cognitive Development, ed Goswami U (Wiley–Blackwell, Malden, MA), 2nd Ed, pp 49–75.
- 43.Meltzoff AN, Moore MK. Early imitation within a functional framework: The importance of person identity, movement, and development. Infant Behav Dev. 1992;15(4):479–505. doi: 10.1016/0163-6383(92)80015-M. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Meltzoff AN, Brooks R, Shon AP, Rao RPN. “Social” robots are psychological agents for infants: A test of gaze following. Neural Netw. 2010;23(8–9):966–972. doi: 10.1016/j.neunet.2010.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mitchell JP, Heatherton TF, Macrae CN. Distinct neural systems subserve person and object knowledge. Proc Natl Acad Sci USA. 2002;99(23):15238–15243. doi: 10.1073/pnas.232395699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Contreras JM, Banaji MR, Mitchell JP. Dissociable neural correlates of stereotypes and other forms of semantic knowledge. Soc Cogn Affect Neurosci. 2012;7(7):764–770. doi: 10.1093/scan/nsr053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Tajfel H. Human Groups and Social Categories: Studies in Social Psychology. Cambridge, UK: Cambridge Univ Press; 1981. [Google Scholar]
- 48.Olson KR, Dweck CS. A blueprint for social cognitive development. Perspect Psychol Sci. 2008;3(3):193–202. doi: 10.1111/j.1745-6924.2008.00074.x. [DOI] [PubMed] [Google Scholar]
- 49.Dunham Y, Olson KR. The importance of origins: Why cognitive development is central to a mature understanding of social cognition. Open Psychol J. 2008;1:59–65. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.