

Abstract
Waves and dynamic patterns in chemical and physical systems have long interested experimentalists and theoreticians alike. Here we investigate a recent example within the context of cell biology, where waves of actin (a major component of the cytoskeleton) and its regulators (nucleation promoting factors, NPFs) are observed experimentally. We describe and analyze a minimal reaction diffusion model depicting the feedback between signalling proteins and filamentous actin (F-actin). Using numerical simulation, we show that this model displays a rich variety of patterning regimes. A relatively recent nonlinear stability method, the Local Perturbation Analysis (LPA), is used to map the parameter space of this model and explain the genesis of patterns in various linear and nonlinear patterning regimes. We compare our model for actin waves to others in the literature, and focus on transitions between static polarization, transient waves, periodic wave trains, and reflecting waves. We show, using LPA, that the spatially distributed model gives rise to dynamics that are absent in the kinetics alone. Finally, we show that the width and speed of the waves depend counter-intuitively on parameters such as rates of NPF activation, negative feedback, and the F-actin time scale.
Keywords: actin waves, signalling, nucleation promoting factors, reaction diffusion systems, pattern formation, nonlinear stability analysis, local perturbation analysis
1. Introduction
The actin cytoskeleton is an important determinant of cell shape, function, and motility. Hence, the dynamics and assembly of filamentous actin (F-actin) from its subunits (G-actin) is highly regulated in living cells. Recent experimental observations have shown remarkable spontaneously generated spatiotemporal patterns of F-actin in cells such as Dictyostelium discoideum [1, 2], neutrophils [3], fibroblasts [4] and mast cells [5]. The patterns include stationary fronts, moving spots, and moving waves, providing a rich set of “benchmark dynamics” that models for actin organization should address. An interesting theoretical question is what minimal, biologically-grounded mechanism suffices to explain such waves and patterns.
Actin interacts with other proteins that regulate its nucleation and assembly. Experimental data supports the role of these regulatory proteins (often denoted actin “nucleation promoting factors,” abbreviated NPFs) in producing the dynamic patterns of actin [6, 7, 8, 9]. Such observations have led to physical and biophysical models that aim to understand the interactions between these signalling proteins and actin [3, 10, 11, 12]. However little effort so far has been devoted to using mathematical methods to investigate these phenomena, motivating our exploration in this paper.
In a previous paper [13], we considered signalling proteins such as small GTPases (Rac, Rho, and Cdc42) as putative NPFs. Upstream of actin, small GTPases have been observed to undergo spontaneous symmetry breaking leading to wave dynamics. Polarization of Rac was shown to be required for formation of Hem1 / F-actin waves in [3]. Machacek et al [14] provided quantitative evidence for the correlation of Cdc42, Rac1, and RhoA wave dynamics with F-actin mediated edge protrusion. In [5], waves of Cdc42 were shown to be correlated in both space and time with waves of F-actin.
We describe the interactions of F-actin and its regulators by a system of reaction-diffusion equations (RDEs) and view the formation of waves and other dynamics as a pattern formation process. More broadly, reaction-diffusion models have been used to investigate pattern formation in physical or physicochemical processes [15, 16, 17], morphogenesis and development [18, 19], or distributions of populations in a changing environment [20, 21, 22, 23]. A common sufficient (and usually necessary) condition for patterning in these cases is different rates of diffusion. The system discussed here, like many biochemical regulatory systems, is characterized by not just different, but vastly different rates of diffusion, which we exploit to simplify model analysis.
To search the parameter space of this model and investigate the structure responsible for initiation of patterning in different regimes of behavior, we utilize a relatively new nonlinear stability analysis method for such systems, the Local Perturbation Analysis (LPA) [24]. This method takes advantage of the disparity between slow and fast diffusion timescales to reduce the system of RDE’s to a system of ODE’s whose linear and nonlinear stability properties mimic those of the RDE’s. Results of this analysis in conjunction with linear stability analysis and numerical simulation demonstrate a rich collection of patterning behaviors distinct from those observed in other systems. We identify the mathematical structure responsible for these distinct dynamics and show that this is a minimal model capable of producing such dynamics.
In our previous paper [13], we presented preliminary analysis of the actin wave model, together with some simulation results. In this follow-up paper we have two distinct aims. (1) To understand the regimes of behavior of the waves in greater detail and to characterize parameter dependence of wave attributes such as speed and width. (2) To highlight and explain features of LPA and its mathematical properties. Our detailed (though informal) exposition is meant to enable the reader to use similar methods on related problems.
2. Biological and modelling background
Many previous models of actin waves [3, 10, 11, 25, 26, 12], reviewed in [27, 13], are based on F-actin interacting with nucleation promoting factors such as Hem1 [3] and WASP or Scar/Wave [6, 7, 8, 9]. Some of these models consider the length or angular distributions of F-actin, and some depict the interactions of individual actin filaments [25].
In contrast, we consider a simplified model that can be analyzed mathematically. We focus on signalling proteins (small GTPases), coupled to a minimal F-actin representation. Our motivation for so doing is as follows: (a) Small GTPases (such as Cdc42 and Rac) are known to regulate the assembly of F-actin [14, 28, 29, 30, 31] and participate in the propagation of actin waves [3, 14, 5]. (b) A minimal model of small GTPases [32, 33] exhibits wave-based symmetrybreaking and formation of a simple polarization pattern. (c) There is evidence for the existence of feedback between F-actin and the small GTPases; see experimental work in [34, 35, 36] on the PI3K pathway and [37, 38] on integrins. (d) Combining the minimal GTPase model with a simplified F-actin model leads to a range of interesting patterns, as we have already described in [13]. As we sought a model of sufficient simplicity to yield mathematical insights, this minimal model proved to be ideal.
Briefly, small GTPases are proteins that cycle between an active state A (bound to the cell membrane) and an inactive state I (in the cytosol, the fluid cell interior). GTPases are activated by GEFs and inactivated by GAPs. In previous work, we investigated the role of these proteins in cell polarization, arriving at a simple caricature in [32] that was analyzed mathematically in [33]. We showed that the A, I model sustains waves that decelerate and stop inside the cell domain; this behavior was termed “wave-pinning”. Here we ask whether a small extension of the A, I model to include feedback to and from F-actin would engender any patterns similar to those observed experimentally. As we show in this paper, this proves to be the case.
2.1. The regulatory sub-model and its “wave-pinning” behavior
In this section, we briefly describe the regulatory sub-model. Let A, I represent active and inactive forms of a small GTPase protein with A bound to the membrane and I freely diffusing in the cell interior. This A, I system plays a regulatory role for F-actin assembly (analogous to the role of nucleation promoting factors, abbreviated NPFs) as well as being affected by feedback from F-actin. Equations for A, I, modified slightly from [32], are:
| (1a) |
| (1b) |
with D̃A ≪ D̃I and no flux boundary conditions. Note that we used A, I here to denote the amount of active and inactive NPFs. In the interconversion of A and I (which corresponds to activation and inactivation), the total amount A + I is conserved and A enhances its own activation. This positive feedback is represented by a Hill function (with magnitude γ, sharpness controlled by n ≥ 2, and Ã0 the typical level of A at which positive feedback “turns on”). The parameter k0 is the basal rate of activation and δ̃ sets the overall rate of dynamics. Details for the choice of this function are motivated in [39, 40, 41, 32] based on known and hypothesized properties of the small GTPases.
In [42], we noted two regimes of behavior. In one regime, arbitrarily small noise can destabilize a homogeneous steady state (HSS) and lead to patterning, as in classical Turing pattern formation. In the second regime, the HSS is linearly stable, and only a sufficiently large perturbation can lead to patterning. This perturbation produces a region of high activity that spreads then stalls (“wave-pinning”). The properties of this model responsible for wave-pinning include 1) D̃A ≪ D̃I (resulting from membrane bound vs free diffusing character), and 2) conservation of the total A + I in the domain. Heuristically, auto-activation of A induces a region of locally high activity. Diffusion causes this region to spread. Depletion of I as the wave spreads causes the wave to slow and stall in the interior of the domain.
3. Model for the full system: regulators and F-actin with positive and negative feedback
We asked whether linking the A, I signalling model to an F-actin equation could account for some of the dynamic F-actin patterns and waves. In experimentally observed actin waves, it is seen that a localized wave of actin nucleators leads and a localized F-actin wave follows [3]. This observation suggests negative feedback from F-actin to NPFs. These ideas led us to adopt a framework similar to the FitzHugh Nagumo [43] model, with nonlinear wave generation coupled to slow feedback as depicted in Fig. 1(a). We investigate how this coupling leads to both stationary and wave dynamics.
Figure 1.
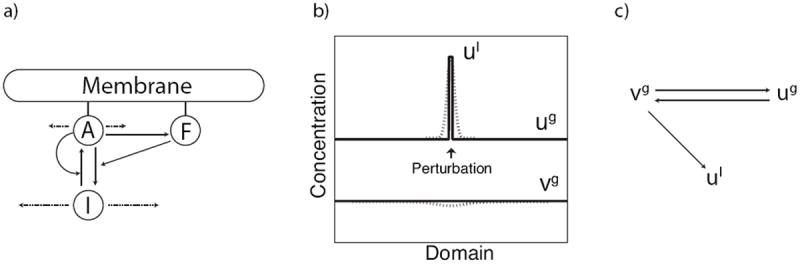
(a) A schematic diagram of interactions between F-actin (F) and its regulatory proteins (NPFs). A is active, membrane-bound protein and I is its inactive cytosolic form. We assume that A promotes actin nucleation and assembly. We also assume positive feedback from A and negative feedback from F to the rate activation of A. Total A + I is conserved. Dashed arrow represent slow or fast diffusion (not to scale). (b) A spatially localized perturbation showing the idealization that leads to the LPA system of local and global variables (solid), and the actual behavior in the full RD system (dotted). (c) Schematic representation of interactions of the local and global LPA variables. ug, υg interact exactly as in the well-mixed system. ul is local, and influenced by (but does not afffect) the global variables.
The model, schematically outlined in Fig. 1(a), represents the interactions of the active (A) and inactive (I) regulatory protein (NPF) with F-actin (F). The equations of the full model (after nondimensionalization, see Appendix B) are:
| (2a) |
| (2b) |
| (2c) |
We refer to this as the full model or as the F-actin regulation model. Neumann boundary conditions are assumed for A, I. The regulatory kinetics f are as in (1), with the modification that F-actin exerts a net negative feedback on its own regulators. Here we have chosen to depict this feedback with a Michaelis Menten actin-dependent rate of inactivation of A. This saturating response prevents over-damping of the entire system. The parameters s1 and s2 represent the relative contributions of basal inactivation, and inactivation due to feedback from F.
We are particularly interested in understanding how the balance between NPF activation and actin-feedback mediated inactivation affect wave formation and propagation. We thus focus on the parameters k0 and s2 that affect this balance. Other combinations of (in)activation parameters yield similar results. That is, an analysis of the parameter space structure with respect to γ and s2, for example, is qualitatively the same. The parameter ε determines the relative timescales of actin and regulatory protein dynamics, respectively, and will be considered as well. We assume that the F-actin feedback occurs on a slower timescale, so that ε ≪ 1. In models for wave propagation in neurons, variables that play a similar role to F are often denoted “slow”, or “recovery” or “refractory”. We will use similar terminology here. The balance between feedback and activation, and the effect of relative timescales will be investigated in subsequent sections.
We have made a few standard simplifying assumptions in formulating the model. We assume that the cell has spread onto a flat surface and that its depth is uniformly thin. To leading order, the concentration of I in the cell interior is then uniform in the depth direction. We further neglect structures such as the nucleus and consider a 1D strip of uniform width along the diameter of the cell. A, I, F are then functions of a single spatial variable (x) and time. The membrane bound / freely diffusing proteins differ only in their rates of diffusion, avoiding the issue of multiple physical compartments. See [42, 13] for details and justification. It is easily seen that A, I in Eqs. 2 satisfy a conservation law
4. Methods
Here we define terminology and introduce the techniques used to analyze the model. Consider a system of reaction-diffusion equations
| (3a) |
| (3b) |
that has been nondimensionalized appropriately, so that f, g = O(1) with no flux boundary conditions on [−1, 1]. Here, p represents a vector of system parameters and in general, u, υ could be vector valued with Du and Dυ being diagonal matrices encoding diffusion coefficients. We further restrict attention to a class of RDEs involving fast and slow diffusing variables so that Du ≪ 1 ≪ Dυ. For example, u = (A, F) represents membrane bound or stationary proteins and υ = (I) represents cytosolic proteins. We also frequently refer to the “well-mixed”, i.e., spatially homogeneous variant of this system:
| (4) |
The Local Perturbation Analysis (LPA) is an effective method for analyzing fast / slow systems of the form (3) [24]. This method was conceived by Marée and Grieneisen [44] and has been used extensively in [39, 45, 42, 13] to parameterize and explore models of signalling networks.
LPA is an approximation of the dynamics of a pulse under the limit where Du → 0 (u is infinitely slow diffusing) and Dυ → ∞ (υ is infinitely fast diffusing, reminiscent of a “shadow system” [46]). Let ul(t) denote the amplitude of a spatially localized perturbation and ug(t), υg(t) the essentially homogeneous background levels of u, υ. Then ul will grow or decay independently of ug. Inhomogeneities of υ will be instantly smoothed out, leaving only a global quantity υg. Assuming a perturbation with negligible total mass (i.e., whose “area” vanishes with Du), υg will be affected only by ug. (See Fig. 1(c)). Under this limiting approximation, the dynamics of the narrow pulse-like perturbation can be approximated by ODEs for (ul(t), ug(t), υg(t)):
| (5a) |
| (5b) |
| (5c) |
We refer to this system as the LPA-ODE’s, and to the analysis of its behavior as the Local Perturbation Analysis (LPA).
Before applying this method to the actin-regulatory model, we collect a few results about LPA and the type of information it is capable of providing. Details of the reasoning and validation of a number of results are provided in the Appendix.
Let (us, υs) denote the homogeneous steady state of (3). Then (ug, υg, ul) = (us, υs, us) is a fixed point of the LPA-ODE’s (5). Thus stability of the HSS to a localized perturbation (LPA stability) is determined by eigenvalues of
| (6) |
In the Appendix we provide arguments to support the following properties.
Property 1
LPA recapitulates linear stability results for the well-mixed system (4); that is to say, eigenvalues of the linearized well-mixed system about (us, υs) are contained in the spectrum of (6). These will be referred to as the well-mixed eigenvalues of JLP.
Property 2
LPA recapitulates linear stability results for the full reaction-diffusion system (Turing stability analysis). More precisely, the remaining eigenvalues of JLP approximate Turing eigenvalues obtained by linearizing the full system of RDE’s about (us, υs). We will refer to these as the “local” eigenvalues of .
In Appendix A, we show Property 2 in a 2 × 2 system. In the 3 × 3 (A, I, F) system, we show this property numerically. In higher order systems, we conjecture (but do not prove) that the same property holds.
The advantages of the LPA method are as follows:
LPA mirrors predictions of other stability analysis methods; it detects the number, values, and stability of spatially homogeneous steady states (HSS).
LPA predicts the response of the system to large amplitude perturbations. This property will be illustrated by example.
Since LPA leads to a set of ODE’s (5) and harnesses standard bifurcation techniques and software, it is substantially simpler to implement than other nonlinear, weakly nonlinear [47], or asymptotic methods [48].
Further, LPA is scalable to systems with many interacting components provided their components are separable into fast and slow diffusion classes. (Within a class, diffusivities need not be the same.) While this scalability will not be illustrated here (given the relatively few components of the actin waves model), we refer the reader to [42] for an example of a complex system with many interacting components to which this method was successfully applied.
While LPA makes useful predictions under specific conditions, it also has limitations as follows:
LPA applies only to RDEs with fast-slow variables, and not to reaction-advection PDEs.
LPA does not approximate the full dynamics of RDEs nor the evolution of their solutions, only stability properties.
The LPA approximation breaks down if the perturbation becomes very large or spreads in space. (This leads to interactions between local and global variables and boundary layer issues).
LPA provides no information about wave numbers of destabilizing periodic perturbations (predicted by Turing analysis), since spatial scales are absent in the asymptotic limit.
With these caveats in mind, we next apply LPA to the actin waves model.
5. Analysis of the (A, I, F) model
Parameter values for the underlying A, I dynamics are based on [32]. In absence of conclusive experimental data, feedback from F-actin is chosen to be in a range that is neither too strong nor too weak. All parameters are nondimensionalized as discussed in Appendix B and listed in Table 1.
Table 1.
Default nondimensional parameter values used in simulations of the full model. These values produce a reflecting pulse. Bracketed values produce a wave train. The abbreviation NPF represents the regulatory protein. Values for most dimension-carrying NPF parameters are based on small GTPases in [32], and lead approximately to the above dimensionless values based on scaling discussed in Appendix B.
| Parameter | Value | Description |
|---|---|---|
| DA | 0.00033 | Diffusion coefficient of A |
| DI | 0.033 | Diffusion coefficient of I |
| k0 | 0.05 (0.2) | Basal rate of activation of NPF |
| γ | 1 | Magnitude of NPF positive feedback |
| A0 | 0.4 | Level of active NPF for positive feedback to occur |
| s1 | 0.5 | Basal NPF inactivation rate |
| s2 | 0.6 (1.2) | F-actin-mediated NPF inactivation rate |
| kn | 2.0 | Rate of F-actin nucleation by NPF |
| ks | 0.25 | F-actin disassembly rate |
| ε | 0.1 | F-actin dynamics rate parameter |
| TN P F | 1 | Total amount of NPF |
We apply the local perturbation reduction to (2) and analyze the resulting LPA-ODE’s to determine regimes of pattern initiation. We focus on how the basal activation rate (k0) and the negative feedback from F-actin (s2) influence dynamics. (Other parameters were tested and found to produce similar results. For example, the behaviour relative to s1 is reciprocal to that relative to k0. Results for γ were similar, and are omitted for brevity.) After the LPA, we explore the long-term dynamics of the patterns with full RDE simulations. See Appendix D for details about numerical implementation.
5.1. The well-mixed system
A standard bifurcation analysis of the well-mixed variant of the system (2) with respect to k0 leads to Fig. 2(a), where we display the steady state A. There is a single unique HSS that is stable for all k0 values, given the default parameter set.
Figure 2.
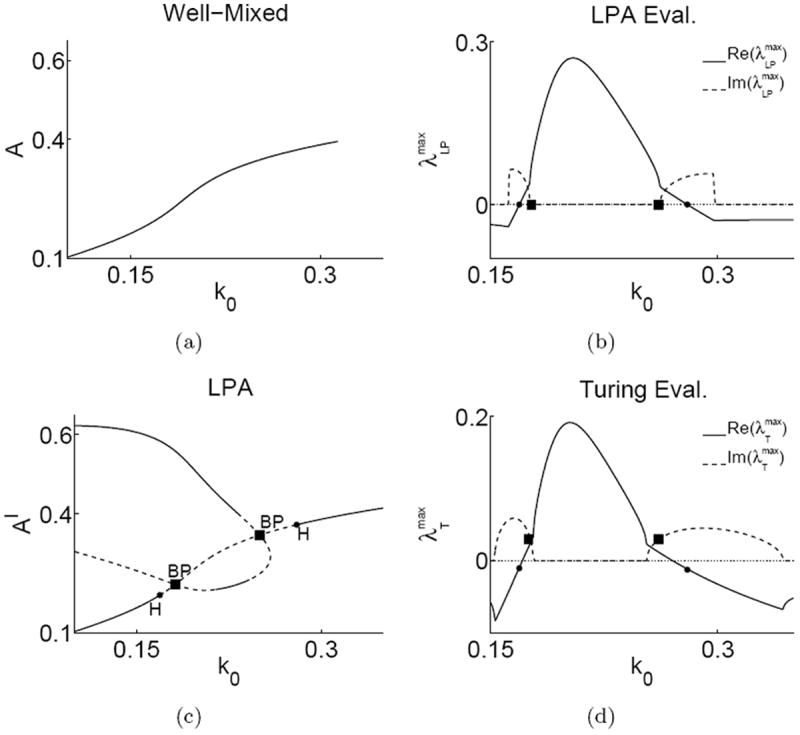
Bifurcation analysis and eigenvalues in the actin-wave model for s1 = 0.7, s2 = 0.7, ε̃ = 0.1 and other parameters at default values. Left panels: Analysis of well mixed (a) and LPA (c) ODE’s plotting steady states of A and Al respectively versus the basal activation rate k0 (Solid = stable, dashed = unstable). The well-mixed model has a single stable steady state (a). The LPA system (c) has the same global branch, and additional local branches. ■, BP = branch points, ●, H = Hopf bifurcation. Right panels: A comparison of maximal (with largest real parts) LPA (b) and Turing (d) eigenvalues versus k0. (Solid = real, dashed = imaginary parts). Symbols ■, ●, mark the corresponding k0 values in (c). Hopf points correspond to zero real part and branch points to zero imaginary parts of . The eigenvalues in (b) and (d) are similar and the Hopf and BP bifurcations closely relate to bifurcations of the RDE model.
5.2. LPA analysis
We set up LPA equations for the full actin-regulatory protein model (see the Eqs. (C.1)). In doing so, we treat the non-diffusive F-actin and the active protein A as slow variables and inactive protein I (rapidly diffusing in the cytosol) as a fast variable. Consequently, the LPA equations have variables Ag, Ig, Fg, Al, Fl.
A bifurcation analysis of the LPA equations (C.1) is shown for Al versus k0 in Fig. 2(c). Comparing this diagram to the diagram in Fig. 2(a), the monotonically increasing well-mixed branch (hereafter denoted the “global branch”) appears in both as expected. We also find new features in the LPA diagram, including a second “local branch” emanating from branch points (BP). This local branch represents additional states that only the local variables can attain.
Interpreting the LPA bifurcation diagram of Fig. 2(c) from left to right, we first find that for small k0, the global branch is stable to small perturbations, whereas the local branch has both unstable (dashed) and stable (solid) regions. Taken together, the global and local branches demonstrate a response threshold: a local perturbation large enough to elevate Al above the (dashed) threshold will be attracted to the upper (stable) local branch while the global variables remain at their unique fixed point (lowest, global branch). Significantly, as the background rate of activation of regulatory protein increases, the size of the stimulating pulse needed to produce a patterned response decreases. This result implies that the system becomes more sensitive to stimulation.
In the LPA diagram, we also find Hopf bifurcations (H) at which the global branch loses stability. Here, the three “well-mixed” eigenvalues have negative real parts (indicating stability). Thus the instability must arise from the two remaining “local” eigenvalues, in this case complex conjugates whose real and imaginary parts plotted in Fig. 2(b). At H, these eigenvalues cross the imaginary axis. This fact indicates that oscillations arise not from the kinetics of the wellmixed system (where no interesting bifurcations take place) but rather from the interactions of the local variables Al, Fl.
We compared eigenvalues of the LPA system and of the full RDEs in the limit Du → 0, Dυ → ∞. From Figs. 2(b), 2(d), we see that LPA eigenvalues provide a reasonable asymptotic approximation of Turing eigenvalues. This confirms that Result 2 in Appendix A holds for this three component system. The significance of this result is that the full model exhibits responses that depend on spatial aspects of the system that are absent in homogeneous (well-mixed) variants of the model.
Further notice that between the branch points (BP) in Figs. 2(c), 2(b), the local eigenvalues are real and positive, indicating simple Turing instability. Between the Hopf and branch points, eigenvalues are complex conjugates with positive real part, corresponding to a traveling wave (oscillatory Turing) instability, as found in [49, 50] for example. The LPA thus predicts four stability regimes. (a) Between k0 = 0 and the bifurcation labelled, H, sufficiently large perturbations induce patterning (nonlinear instability). (b) In the range from H to BP, a wave instability is present. (c) Between the two BP bifurcations, a stationary Turing instability is present. (d) From H to k0 = ∞, no patterning is possible.
5.2.1. Role of conservation
Consider for the moment the actin-NPF model Eqs. (2) with the quantity I fixed as a parameter. Then the remaining A, F system has an activator-inhibitor structure that can be shown to oscillate. This is the source of the Hopf bifurcation and corresponding oscillatory dynamics of the local variables Al, Fl, for which Ig plays the role of a parameter (since I is fixed at its steady state value). The global variables Ag, Ig, Fg, on the other hand, satisfy the conservation law Ag + Ig = 1 and evolve according to the kinetics of the well mixed system, which exhibits a single stable steady state without oscillatory dynamics. We thus see that a minimum of two slow variables (A, F) are required to produce the complex pair of local eigenvalues corresponding to oscillations on a local scale. The third fast variable (I) is necessary to suppress these oscillations on the global scale and stabilize the well mixed system. In this way, the A, I, F model is a minimal system demonstrating the type of dynamics observed in our spatial model.
5.3. Two-parameter bifurcation analysis
In order to investigate the interplay between the activation parameter k0 and the F-actin negative feedback s2, we next perform a two-parameter bifurcation analysis, tracking both the Hopf (H) and branch point (BP) bifurcations, Fig. 4. Interesting dynamics lie along a diagonal region in Fig. 4(a), where the activation and feedback are roughly balanced. While Fig. 2(c) identifies both wave and Turing instabilities for s2 = 0.7, there is a region at higher s2 values where only wave instabilities occur. As s2 is increased further, the Turing region shrinks and terminates. This observation indicates that (a) stationary Turing instabilities are prominent at lower s2 values, (b) oscillatory wave instabilities take over at intermediate s2 values, and (c) yet higher s2 values stabilize the HSS.
Figure 4.
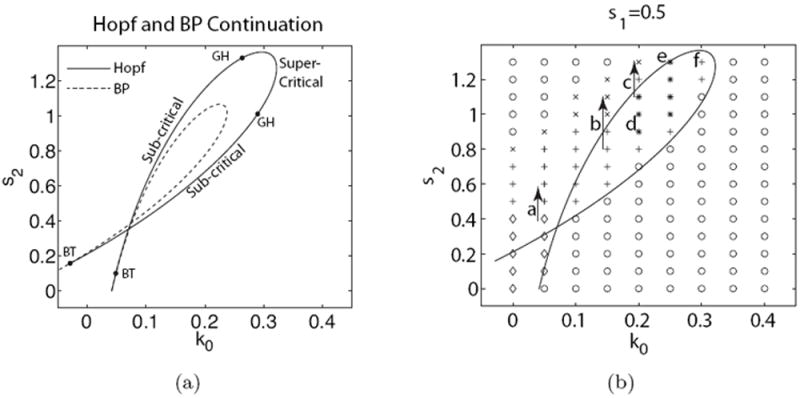
Left: The s2, k0 two-parameter continuation of the H and BP bifurcations seen in Fig. 2(c). GH and BT represent co-dimension 2 Generalized Hopf and Bogdanov Takens bifurcations. At BT, the solid branch switches from being a neutral saddle to a Hopf bifurcation. At GH, the criticality of the Hopf curve changes. s1 = 0.5, ε = 0.1 and all other parameters as in Table 1. Right: The same parameter plane showing classification of the resulting long-term dynamics into one of several patterns: no pattern (o), boundary localized (◇), reflecting pulse (+), pulse trains (*), and a single pulse (×). Arrows and letters indicate the parameter values corresponding to long term simulations in Fig. 5. The LPA s2, k0 two-parameter continuation of the Hopf bifurcations (curves) is superimposed.
For low values of s2, k0, in Fig. 4(a) the Hopf bifurcations are sub-critical and large amplitude oscillations result immediately upon crossing these bifurcation points. When s2, k0 increase, the Hopf branch undergoes a Generalized Hopf bifurcation (GH), becoming super-critical. Bogdanov-Takens (BT) bifurcations represent the intersection of the Hopf and BP curves. For s2 below these bifurcation points, the H branch represents neutral saddles rather than an oscillatory bifurcation. As s2 is increased, the neutral saddles cross through the BP curve becoming true Hopf points.
5.4. Long term evolution of patterns
In order to gain insight about how predictions of LPA and standard stability analysis compare with actual long-term evolution of patterns, we carried out full numerical simulations of the model equations (2). We explored the simulated behavior (with default parameters as in Table 1), and compared the actual patterns observed to the regimes obtained in the LPA two-parameter continuation of Fig. 4(a). To do so, we used an automated classification algorithm [13] for broad pattern classification, as shown in Fig. 4(b).
For each s2, k0 grid point, a simulation is run and we indicate one of five basic behaviors: no pattern (o), boundary localized (◇), reflecting pulse (+), pulse trains (*), and a single pulse (×). As suggested by the LPA continuation curves, interesting dynamics occur along the diagonal in the s2, k0 plane, where activation (k0) and feedback mediated inactivation (s2) of the regulatory protein roughly balance. The labelled arrows in Fig. 4(b) correspond to simulations in Fig. 5.
Figure 5.
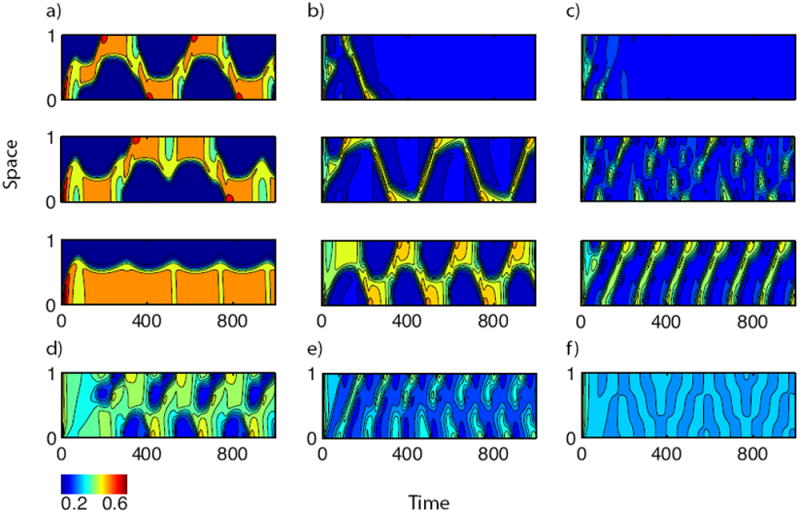
Simulations of full model, showing A(x, t) as intensity plots versus x (vertical) and t (horizontal) axes. (a-c): transitions represented by arrows (a-c) in Fig. 4B, (s2 increases from bottom to top in each column). (d-f) Exotic patterns at the boarders of regimes, as in Fig. 4B. (a) k0 = 0.05, s2 = 0.4, 0.56, 0.6, (b) k0 = 0.15, s2 = 0.8, 1.02, 1.1, (c) k0 = 0.2, s2 = 1.1, 1.24, 1.3, (d) k0 = 0.2, s2 = 0.83, (e) k0 = 0.25, s2 = 1.3, (f) k0 = 0.3, s2 = 1.36. (See also Table 1).
We examined in greater detail some of the transitions between pattern types. Here we comment on several examples, indicated by the span and direction of arrows in Fig. 4(b). In each case, we explain how the pattern changes over a parameter sweep (base to tip of arrow). Then, we show these transitions (bottom to top) in correspondingly labelled panels in Fig. 5. The value s2 = 0 corresponds to pure wave-pinning and hence, to a polarized state. (a) For relatively low values of s2 and k0, a transition from static to dynamic patterns is seen: As s2 increases, the polarized state first undergoes local oscillations at the boundary, and then transitions smoothly into a reflecting pulse solution. (b) At larger k0, s2 values, the width of the reflecting pulse decreases. Eventually, a single pulse traverses the domain once and terminates. (c) For larger s2, an oscillating amplitude is superimposed on the traveling wave train. In both (b) and (c), the pulse is suppressed smoothly with increased s2, eventually leading to transient dynamics. A few additional exotic patterns (d,e,f) are also shown in Fig. 5. We found that at higher values of s2 (not shown), there is complete suppression of dynamics.
Further comparing long-term behavior with the LPA bifurcation structure, we see that different wave regimes can be identified with distinct regimes depicted in the LPA. Wave trains are associated with a Turing bifurcation in which a pair of complex conjugate eigenvalues have a positive real part (“non-stationary Turing bifurcation”). The oscillatory nature of these waves arises as an instability of the HSS, and is predicted by linear stability analysis. In this sense, the wave trains occur “spontaneously” in response to small perturbations or noise. In contrast, reflecting waves occur either in 1) a stationary Turing regime (a single real positive eigenvalue) or 2) linearly stable regime with a threshold that can be breached by a sufficiently large perturbation. In the stationary Turing regime, noise initiates patterning but without any propagation dynamics. Once this pattern grows, nonlinear effects become important and lead to propagation. Thus, reflecting waves are inherently a nonlinear phenomenon, and they can be either spontaneous or excitable. In both regimes 1) and 2), the dynamics of reflecting waves cannot be predicted based on linear stability analysis. We conclude that, while the LPA does not directly predict long-term patterns, it allows some inferences about how stability properties correlate with dynamics. In particular, LPA suggests that reflecting waves and wave trains are initiated in fundamentally distinct ways.
5.5. Pulse Width / Velocity
To further characterize how patterns change as the parameters of the model are varied, we investigated pulse width and velocity (for reflecting pulses and pulse trains). Results for k0 and s2 are shown in Fig. 6. Pulse width was computed at the half maximum and velocity was computed by tracking the location of the maximal value of A as a function of time and computing the slope of the resulting line. This width increases as the activation rate k0 increases (or as the negative feedback parameter s2 decreases), as expected. Wave speed decreases as k0 is increased (or as s2 is decreased) which is unusual, since wave velocities typically increase with activation rates. Both patterns, reflecting pulses and pulse trains, exhibit similar parameter dependence.
Figure 6.
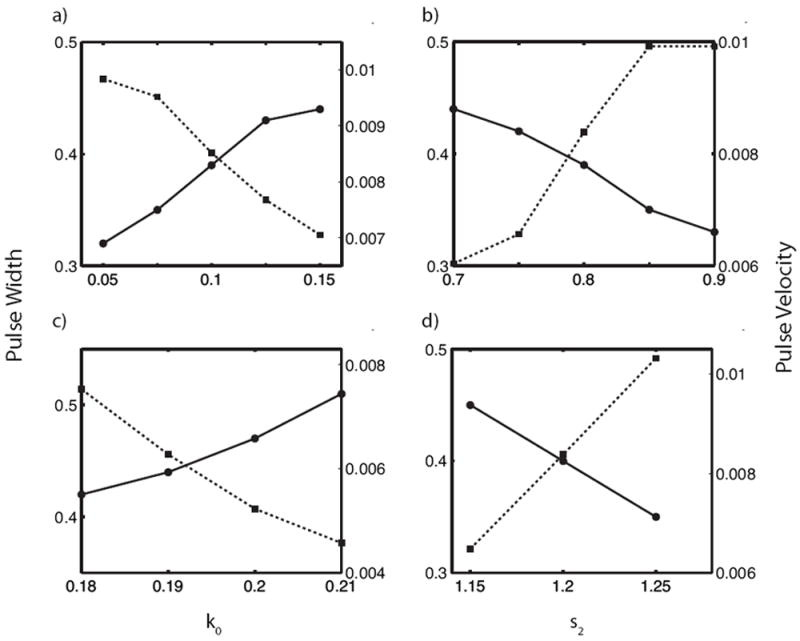
Pulse width (solid) and velocity (dashed) versus k0 (left) and s2 (right) for reflecting pulses (top) and pulse trains (bottom). Width is measured at the pulse half maximum for A, and velocity is determined by plotting the position of the pulse maximum as a function of time and computing the slope. Parameters: (a) s2 = 0.8 (b) k0 = 0.1 (c) s2 = 1.0 (d) k0 = 0.25. All other parameters are as in Table 1.
The dependencies of pulse velocity and width on parameters are particularly interesting as they depart from expectation. In classical wave propagation models such as FitzHugh Nagumo (compared to our model in Section 6), propagation velocity increases with an activation rate, in contrast to what we find here. We can explain this by noting that in our model, the rate of activation depends not only on the rate parameters k0, γ, but also on the available pool of inactive form I. As that pool is depleted, propagation velocity falls [32]. Thus there is an inverse relationship between pulse width and velocity, larger pulses deplete the inactive pool further, slowing propagation. So paradoxically, increasing the rate of activation (k0) increases the depletion of this pool and slows propagation. Similarly, it is depletion of the inactive protein limits the width of the activated regions.
Figure 7 supports this relationship. Figure 7(a) shows that as the total pool of regulatory protein, TN P F, is increased, the pulse width increases commensurately. This increase in pulse width depletes the pool of inactive NPF to roughly the same level at each value of TN P F (data not shown). For this reason, while pulse velocity increases, it does so by a smaller fraction since the net activation rate remains roughly the same.
Figure 7.
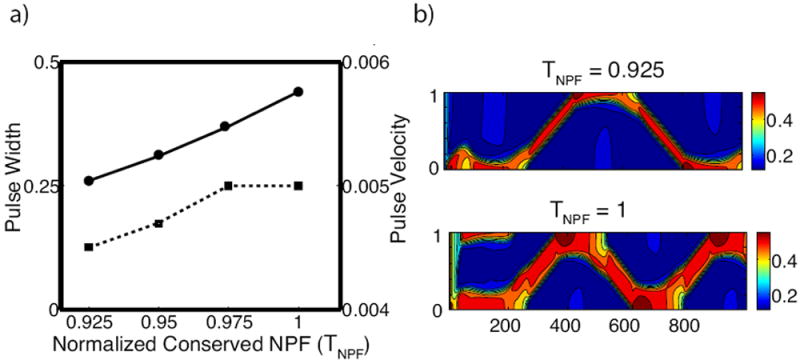
The effects of the total amount of regulatory protein TN P F on pulse propagation. (a) Pulse width (solid) and velocity (dashed) as a function of TN P F. (b) Intensity plot of A at the two endpoints TN P F = 0.925, 1 respectively. Parameters are k0 = 0.15, s1 = 0.7, s2 = 0.6 with all others as in Table 1.
5.6. The F-actin (refractory) time scale, ε
So far, we have explored the roles of activation rate and negative feedback parameters. Small values of ε correspond to a slow F-actin response. Varying this parameter will increase or decrease the influence of the F-actin on its regulators, which corresponds to a change in the timescale for the slow (refractory) negative feedback. It is well known that the timescale of refractory negative feedback is an important determinant of oscillatory dynamics. Hence, we here explore the role of ε and characterize the conditions required for these oscillatory bifurcations to occur.
In Appendix C.1 and Appendix C.2, we show that ε does not affect the values of the steady state of the LPA-system q* = (Āg, Īg, F̄g, Āl, F̄l) but does affect stability. Briefly, the LPA Jacobian is in the from
| (7) |
J0 is the well mixed Jacobian (A.1), and Jloc is the Jacobian of the equations involving the local variables. Both Hi (Eqn. (C.7)) are independent of ε. Jext does not affect eigenvalues due to the block lower triangular structure. Since eigenvalues of J0 are negative, Jloc alone governs the appearance of Hopf bifurcations and oscillations, and its eigenvalues are
| (8) |
(These explicitly depend on ε.) The conditions for a Hopf bifurcation are now
| (9) |
which can be reformulated as
| (10) |
For a fixed set of parameters excluding ε, the above implies that there is a maximum value ε* where B1 ≤ B2, for which a Hopf bifurcation exists. As an illustration, we plot ε = B1,2(k0) for two parameter sets in Fig. 8 (a,c) and LPA diagrams for ε = 0.3 in Fig. 8 (b,d). A Hopf bifurcation exists wherever the curve ε = B1 lies below the curve ε = B2. Where B1 > B2, the B1 curve represents saddle points without oscillatory instabilities. The associated LPA diagrams in panels (b, d) showing the presence (or absence) of Hopf bifurcations are consistent with the results of this eigenvalue analysis.
Figure 8.
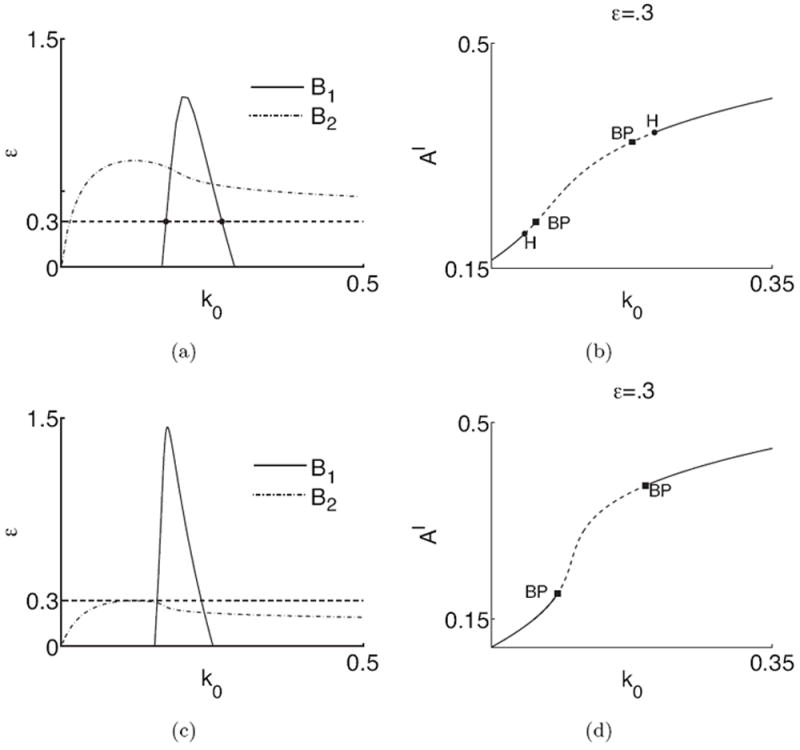
The timescale of F-actin dynamics affects the presence and locations of Hopf bifurcations in the LPA system and oscillatory dynamics in the original RDEs. Left: Plot of B1(k0) and B2(k0) of (10) for (a) s1 = s2 = 0.7, (c) s1 = 0.9 and s2 = 0.3 (Other parameters as in Table 1). Horizontal dashed lines are values of ε corresponding to LPA diagram (b,d). Hopf bifurcations correspond to (k0, ε) values on the B1 curve below B2. Points on the B1 curve above B2 represent saddles. Heavy dots on the B1 curve in (a) correspond to Hopf points in (b). In (b,d), we show LPA bifurcation plots for the parameter sets corresponding to (a,c) respectively with ε = 0.3.
Thus, as the relative timescale of the refractory negative feedback τ = 1/ε decreases, oscillatory dynamics are suppressed. Intuitively, the refractory timescale should be long enough that large deviations of Al driven by activation are achieved before suppression by feedback. This timescale dependence often occurs in single refractory feedback systems, such as the FitzHugh Nagumo (FHN) model. While these conclusions are based on LPA, the fact that local eigenvalues approximate eigenvalues of the linearized RDEs, means that oscillatory linear instabilities would be seen in the original RDE system.
This refractory timescale does not affect the branch point (BP) bifurcations since those result purely from the presence of local branches representing ε independent steady states.
We explored the effect of ε on long term behavior. Keeping other parameters at their default values (Table 1), we varied ε from 0.001 to 0.5 in Fig. 9a. When ε is small, F-actin responds slowly, somewhat decoupling the regulatory protein equations. Consequently, that system exhibits wave pinning dynamics, and static patterns result (panel a, top left). For large values of ε, actin polymerization and regulatory protein dynamics occur on the same timescale, also resulting in static patterns (panel a, bottom right). Patterning is most dynamic between these extremes.
Figure 9.
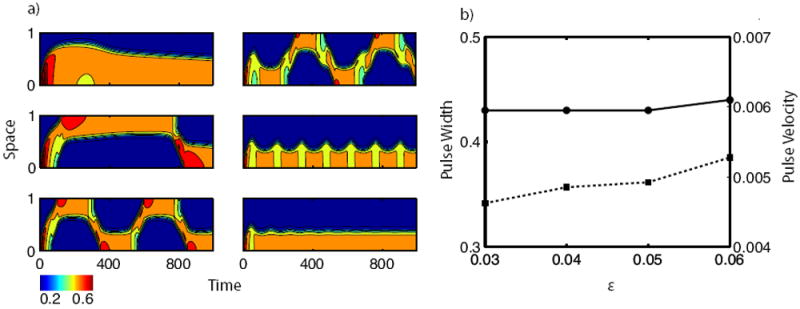
The effect of the refractory feedback rate ε. (a) Intensity plots of A. The ε values from top to bottom, left to right are 0.001, 0.01, 0.05, 0.11, 0.19, 0.29. Other parameters as for reflecting pulses in Table 1. (b) Pulse width (solid) and velocity (dashed) as a function of ε for reflecting waves.
Finally, we asked how ε affects pulse width and velocity. Results for reflecting pulses are shown in Fig. 9b. Similar results were found for pulse trains (not shown). Increasing ε by a factor of 2 results in a 20% increase in the velocity, as expected, but hardly affects the width of the pulse (a mere 2% change). The latter result is counterintuitive, since a larger feedback rate ε should suppress the trailing edge faster, leading to smaller activated regions.
6. Comparison to FitzHugh Nagumo Model
It is illuminating to compare our F-actin regulation model with the FitzHugh Nagumo model [43], a well-known elementary model for excitation waves. Both models have a nonlinear wave generator, and a refractory negative feedback.
In the FHN system, one equation consist of nonlinear (“cubic”) kinetics that engenders bistability and hysteresis. A second linear equation describes the evolution of a “recovery” (or refractory) variable whose feedback leads to cycling around the hysteretic loop. The system has limit cycle oscillations even in the space-independent case. Diffusion spreads the excitation in space, spawning a travelling wave-front separating regions of low and high activity that sweeps entirely through the domain. Refractory negative feedback suppresses the trailing edge of that wave, producing a travelling pulse with a spatially localized high state separating low states on either side. For this model, pulse velocity increases with respect to an activation rate that resembles the activation parameter k0, and pulse width decreases as an ε like refractory rate parameter increases.
In the F-actin regulation model, our results indicate that stability properties and long term dynamics of (2) are not merely inherited from the behavior of the space-free system. In the parameter range of interest, and in absence of feedback (s2 = 0), that underlying system has a single steady state. Turning on the refractory feedback (s2 > 0) does not induce limit cycle oscillations. Rather, diffusion plays a vital role in determining the wave and pulse behavior. So, in contrast to the standard FHN system, the reaction kinetics alone do not sustain oscillatory dynamics; in fact, diffusion is required for wave formation. The reason for this is that in contrast to the FHN system, instead of traditional bistability associated waves, the wave generating A, I system of proteins sustain a “wave pinning” behavior, for which diffusion is a key ingredient. Refractory feedback interacts with this form of wave generation to produce travelling wave and Turing bifurcations of the full RDEs. Conservation of A + I is a key feature that gives rise to these interesting stability properties. With I held fixed, A and F exhibit typical activator/inhibitor oscillations on a local spatial scale (as demonstrated by the local variables). The conservative dynamics of I on the other hand stabilize the spatially homogeneous system.
Furthermore, as shown above, the long-term dynamics of the F-actin regulation model and those of FHN differ significantly. In our model, we find both persistent and transient dynamics in linearly stable parameter regimes, whereas FHN shows only transient single pulse dynamics. Second, polarized patterns, boundary oscillations, and reflecting wave dynamics are not exhibited by the FHN system. Third, pulse velocities depend inversely on the regulatory protein activation parameter k0 rather than directly. Fourth, the size of an activated region is insensitive to the refractory timescale ε. Finally, our results on the relationship between the total pool of regulatory protein, TN P F, and pulse width and velocity (Figure 7) also demonstrate the importance of conservation of the total amount of regulating protein (TN P F). This is the key structural property differentiating out model from FHN.
7. Discussion
At present, it is not certain whether actin waves are secondary phenomena, or whether they have functional significance to cell behavior. It has been speculated that the waves of F-actin aid cells to explore their environment [3, 51], engulf particles [52], and/or organize cell edge protrusion [53]. Whether or not such waves have functional significance, their existence informs models for F-actin and its regulators. Here we have shown that a minimal model, combining a caricature of small GTPases (the “wave pinning” A, I system) with feedback to/from F-actin is capable of producing a rich diversity of patterns: static polarization, reflecting waves, wave trains, transient pulses, and persistent boundary oscillations, with a variety of smooth transitions between them. Further, it is one of the simplest models exhibiting such behavior.
In analyzing regimes of behavior of the model, we introduced and applied the local perturbation analysis (LPA), a method for determining nonlinear stability properties. This method rests on the observation that for systems with a large diffusion disparity, the response to an arbitrarily tall but spatially localized perturbation can be approximated by a set of ODEs for global (fast) and local (slow) variables. LPA bifurcation plots (Figs. 2, 4a) then provide useful information about growth or decay of that perturbation. While the predictions are (like all stability analyses) short-term, they help to highlight how patterning is initiated: whether through arbitrarily small amplitude stimuli, or as threshold responses to perturbations of some finite size. In this sense, LPA provides a combination of linear and nonlinear stability information.
It is worth mentioning that LPA is uniquely suited to analyze biologically motivated systems. For one thing, it is capable of detecting both linear and nonlinear patterning phenomena. Linear and nonlinear methods can in some cases be combined to provide similar or even more complete information. However this is usually possible only in specific situations where functional forms are amenable to analysis. Furthermore, such methods are usually only applicable in systems involving a small number of variables. We have applied LPA to substantially larger and more complex systems without particular difficulty. Examples can be found in [42, 39, 45]. Since LPA takes advantage of existing bifurcation software, the technical/computational “cost” of its implementation is low compared to other methods.
Applied to the actin-waves model, LPA reveals two distinct linearly unstable regimes (both stationary Turing and oscillatory wave instabilities, in agreement with classical linear analysis), and a third regime whose homogeneous steady state is unstable to a sufficiently large perturbation (not detected by linear methods). Simulations of the full model confirmed those LPA predictions. For example, the two-parameter LPA bifurcation diagram (Fig. 4) predicts that interesting dynamics occur when there is a rough balance between activation (k0) and inactivation (s2) parameters, in agreement with simulations. Wave trains were found to correlate with the wave instability detected by the LPA, and reflecting waves with the stationary Turing instability and threshold patterning regimes. Further analysis showed oscillatory instabilities require the actin feedback rate (ε) to be smaller than the protein activation rate.
While aspects of the model were motivated by the refractory and bistable components of the FitzHugh Nagumo model, we showed that its behavior contrasts in important ways. For one thing, our model has no inherent oscillatory features when the system is well-mixed and the spatial distribution is essential. Second, pattern regimes are significantly more diverse than those found in the FHN model. Third, pulse width and velocities depend in counterintuitive ways on parameters. Finally, the conservation of A + I in our model appears to be a distinguishing feature that is vitally important in determining these dynamics.
We can use several features of the model to suggest experimental tests. Among these are the role of conservation of the regulatory proteins and the predicted inverse relationship between pulse width and velocity. First, we predict that any perturbation that increases NPF activation would lead to wider, slower waves. Consequently, increasing GEF activation or overexpressing a GEF (which corresponds to increasing the basal activity k0, of NPF’s such as Cdc42 or Rac) should produce this effect. Conversely, overexpressing or increasing GAP activity (identified with increasing the rate of inactivation of GTPases, s1) should have the opposite effect, producing thinner, faster waves. Perturbations that magnify F-actin mediated feedback, indicative of an increase in s2 should produce this result. Overexpression of Arp2/3, WASP, or the Wave complex (identified with an increase in kn) would, by enhancing F-actin production, also lead to thinner, faster waves. Finally, overexpressing GTPases such as Cdc42 or Rac, which is equivalent to increasing TN P F should lead to wider waves while having a modest effect on propagation velocity.
Figure 3.
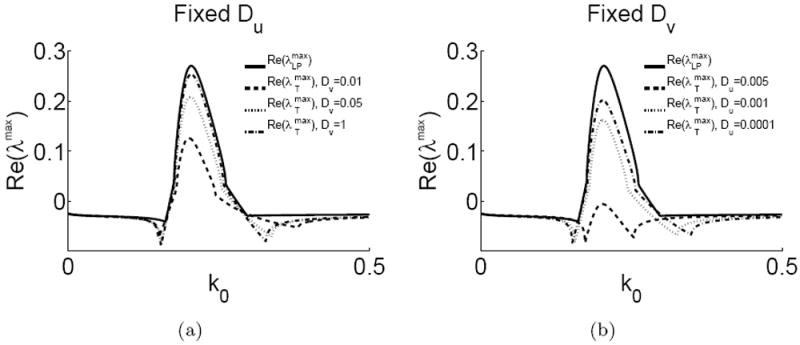
In the limits (a) Dυ (i.e. DI) → ∞, (b) Du (i.e. DA) → 0, Turing maximal eigenvalues (dashed and dotted) approach LPA eigenvalues (solid). The real parts of λmax is plotted against k0. In the given limits, the LPA and Turing eigenvalues agree well.
Highlights.
We propose a model for F-actin interacting with its nucleation promoting factors
The model consists of 3 PDES of the reaction-diffusion type
The model exhibits novel dynamics of waves and propagating patterns.
A non-linear, local perturbation analysis is used to investigate pattern initiation.
Our model depicts the minimal set of interactions that can produce these dynamics.
Acknowledgments
The authors acknowledge support from NSERC discovery grant to LEK and NSERC postgraduate fellowship to MD. LEK and WRH have also been partly supported by NIH grant R01 GM086882 (to Anders Carlsson) and WRH has been partially supported by NIH grant P50GM76516.
Appendix A
Comparison of Local Perturbation Analysis (LPA) and linear stability (Turing) Analysis
Here we provide details for the comparison of the nonlinear LPA stability analysis method to the linear Turing stability analysis. Consider the homogeneous steady state, (us, υs) of (3). Linear stability to spatially nonuniform perturbations is determined by eigenvalues of
| (A.1) |
These eigenvalues are growth rates of a small periodic perturbation of the form cos(kπx). Define to be those eigenvalues.
Note that (ug, υg, ul) = (us, υs, us) is a fixed point of the LPA-ODE’s (5). Thus stability of the HSS to a localized perturbation (LPA stability) is determined by eigenvalues of (6), reproduced below
| (A.2) |
Property 1
LPA recapitulates linear stability results for the well-mixed system (4).
Verification
Notice that (5a), (5b) decouple and represent the well-mixed system (4). As a result, (A.2) is block lower triangular with the top left block representing J0 (A.1). So are eigenvalues of (A.2) [54] and any instabilities associated with the k = 0 mode (well-mixed instabilities) are found by LPA. From here on, we refer to as well-mixed eigenvalues.
Property 2
LPA recapitulates linear stability results for the full reaction-diffusion system (Turing stability analysis).
Verification for two variable systems
Note that the remaining eigenvalues of JLP are determined precisely by the eigenvalues of fu (us, υs; p). Define these to be . We compare the Turing eigenvalues with the remaining LPA eigenvalues in the extreme diffusion regime where the LPA is valid. A simplified setting with one fast and one slow diffusing variable is considered. In this setting, the eigenvalues , are roots of the characteristic polynomial
| (A.3) |
In the LPA regime, Du ≪ 1 ≪ Dυ, these roots satisfy
| (A.4) |
Then clearly has negative real part independent of the system parameters and does not contribute to stability properties. Consider instead the O(1) eigenvalue. Substituting into (A.3)
| (A.5) |
Thus, in the relevant limit. This Turing eigenvalue contributes to stability and as we have shown here, is approximated by an LPA eigenvalue. We will refer to as “local” eigenvalues. For the 3 × 3 (A, I, F) system, where this verification does not apply, we show numerically that Result 2 still holds.
Appendix B
Nondimensionalizing the model
The equations of the full actin waves model are:
| (B.1a) |
| (B.1b) |
| (B.1c) |
with
| (B.1d) |
| (B.1e) |
Equations for A, I imply that
| (B.2) |
is constant.
We translate the model into dimensionless form by rescaling the variables as follows:
| (B.3) |
After making these substitutions and dropping thē, the resulting nondimensional equations become (2) with
| (B.4) |
and
| (B.5) |
Note that while total protein amounts have been normalized to one, we will allow TN P F to vary in order to investigate the effects of protein amounts on dynamics. In this way, TN P F parameterizes total protein levels relative to a typical amount T̃N P F. Further note that several rate parameters from the original model (k0, γ, s1, s2, kn, ks) were already dimensionless as a result of aggregating the timescale information into δ̃, ε̃.
Appendix C
LPA equations for the model
The LPA equations for the model are
| (C.1a) |
| (C.1b) |
| (C.1c) |
with kinetics as defined previously in (2).
Appendix C.1. LPA system steady states
The steady state of the LPA-system q* = (Āg, Īg, F̄g, Āl, F̄l) is independent of ε. It satisfies
| (C.2) |
| (C.3) |
| (C.4) |
Further, using conservation (Ig + Ag = TN P F), we can eliminate Ig. Then, after some algebra, the steady states satisfy
| (C.5) |
Clearly one solution satisfies Āl = Āg, F̄l = F̄g as expected, corresponding to the HSS.
Appendix C.2. LPA system stability
We compute the Jacobian of the LPA system Eqs. (C.1) and find
| (C.6) |
J0 is the well mixed Jacobian (Eqn. (A.1)), Jext is extraneous and does not effect eigenvalues, and
| (C.7) |
| (C.8) |
Appendix D
Numerical Implementation
To explore stability regimes and compare predictions of the analytic methods, we carried out a bifurcation analysis of the ODEs corresponding to the LPA and the well-mixed system. This computation is carried out using the bifurcation package Matcont [55] of Matlab (MathWorks Inc). Computation of Turing eigenvalues, LPA eigenvalues, and quantities associated with the refractory timescale ε were performed using Mathematica (Wolfram Research). To track the long-term evolution of these patterns we carried out numerical simulations of the full RDEs, (2) in 1D using a Crank-Nicholson discretization scheme in Matlab.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Vicker M. F-actin assembly in dictyostelium cell locomotion and shape oscillations propagates as a self-organized reaction-diffusion wave. FEBS Lett. 2002;510:5–9. doi: 10.1016/s0014-5793(01)03207-0. [DOI] [PubMed] [Google Scholar]
- 2.Vicker M. Eukaryotic cell locomoation depends on the propagation of self-organized reaction-diffusion waves and oscillations of actin filament assembly. J Expt Cell Res. 2002;275:54–66. doi: 10.1006/excr.2001.5466. [DOI] [PubMed] [Google Scholar]
- 3.Weiner O, Marganski W, Wu L, Altschuler S, Kirschner M. An actin-based wave generator organizes cell motility. PLoS biology. 2007;5:e221, 2053–2063. doi: 10.1371/journal.pbio.0050221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Millius A, Dandekar S, Houk A, Weiner O. Neutrophils establish rapid and robust wave complex polarity in an actin-dependent fashion. Current Biology. 2009;19:253–259. doi: 10.1016/j.cub.2008.12.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wu M, Wu X, De Camilli P. Calcium oscillations-coupled conversion of actin travelling waves to standing oscillations. Proceedings of the National Academy of Sciences. 2013;110:1339–1344. doi: 10.1073/pnas.1221538110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bretschneider T, Diez S, Anderson K, Heuser J, Clarke M, Mueller-Taubenberger A, Koehler J, Gerisch G. Dynamic actin patterns and Arp2/3 assembly at the substrate-attached surface of motile cells. Curr Biol. 2004;14:1–10. doi: 10.1016/j.cub.2003.12.005. [DOI] [PubMed] [Google Scholar]
- 7.Gerisch G, Bretschneider T, Müller-Taubenberger A, Simmeth E, Ecke M, Diez S, Anderson K. Mobile actin clusters and traveling waves in cells recovering from actin depolymerization. Biophysical journal. 2004;87:3493–3503. doi: 10.1529/biophysj.104.047589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bretschneider T, Anderson K, Ecke M, Müller-Taubenberger A, Schroth-Diez B, Ishikawa-Ankerhold H, Gerisch G. The three-dimensional dynamics of actin waves, a model of cytoskeletal self-organization. Biophysical journal. 2009;96:2888–2900. doi: 10.1016/j.bpj.2008.12.3942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xiong Y, Huang C, Iglesias P, Devreotes P. Cells navigate with a local-excitation, global-inhibition-biased excitable network. Proc Natl Acad Sciences. 2010;107:17079–17086. doi: 10.1073/pnas.1011271107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Doubrovinski K, Kruse K. Cytoskeletal waves in the absence of molecular motors. Europhysics Letters. 2008;83:18003p1–6. [Google Scholar]
- 11.Whitelam S, Bretschneider T, Burroughs N. Transformation from spots to waves in a model of actin pattern formation. Physical review letters. 2009;102:198103. doi: 10.1103/PhysRevLett.102.198103. [DOI] [PubMed] [Google Scholar]
- 12.Iglesias P, Devreotes P. Biased excitable networks: how cells direct motion in response to gradients. Current Opinion in Cell Biology. 2012;24:245–253. doi: 10.1016/j.ceb.2011.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Holmes W, Carlsson A, Edelstein-Keshet L. Regimes of wave type patterning driven by refractory actin feedback: transition from static polarization to dynamic wave behaviour. Physical Biology. 2012;9:046005. doi: 10.1088/1478-3975/9/4/046005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Machacek M, Hodgson L, Welch C, Elliott H, Pertz O, Nalbant P, Abell A, Johnson GL, Hahn KM, Danuser G. Coordination of Rho GTPase activities during cell protrusion. Nature. 2009;461:99–103. doi: 10.1038/nature08242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cross M, Hohenberg P. Pattern formation outside of equilibrium. Reviews of Modern Physics. 1993;65:851–1112. [Google Scholar]
- 16.Maini P, Painter K, Chau H. Spatial pattern formation in chemical and biological systems. Journal of the Chemical Society, Faraday Transactions. 1997;93:3601–3610. [Google Scholar]
- 17.Gollub J, Langer J. Pattern formation in nonequilibrium physics. Reviews of Modern Physics. 1999;71:396–403. [Google Scholar]
- 18.Turing A. The chemical theory of morphogenesis. Phil Trans Roy Soc. 1952;237:37–72. [Google Scholar]
- 19.Murray J. A pre-pattern formation mechanism for animal coat markings. Journal of Theoretical Biology. 1981;88:161–199. [Google Scholar]
- 20.Segel L, Jackson J. Dissipative structure: an explanation and an ecological example. Journal of Theoretical Biology. 1972;37:545–559. doi: 10.1016/0022-5193(72)90090-2. [DOI] [PubMed] [Google Scholar]
- 21.Okubo A, Levin S. Diffusion and ecological problems. Springer; 2002. [Google Scholar]
- 22.Cantrell R, Cosner C. Spatial ecology via reaction-diffusion equations. Wiley; 2004. [Google Scholar]
- 23.Yizhaq H, Gilad E, Meron E. Banded vegetation: biological productivity and resilience. Physica A: Statistical Mechanics and its Applications. 2005;356:139–144. [Google Scholar]
- 24.Walther GR, Marée AF, Edelstein-Keshet L, Grieneisen VA. Deterministic versus stochastic cell polarisation through wave-pinning. Bulletin of Mathematical Biology. 2012;74:2570–2599. doi: 10.1007/s11538-012-9766-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Carlsson A. Dendritic actin filament nucleation causes traveling waves and patches. Phys Rev Lett. 2010;104:228102. doi: 10.1103/PhysRevLett.104.228102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Doubrovinski K, Kruse K. Cell motility resulting from spontaneous polymerization waves. Physical Review Letters. 2011;107:258103. doi: 10.1103/PhysRevLett.107.258103. [DOI] [PubMed] [Google Scholar]
- 27.Carlsson A. Actin dynamics: from nanoscale to microscale. Annual review of biophysics. 2010;39:91–110. doi: 10.1146/annurev.biophys.093008.131207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Mackay D, Hall A. Rho GTPases. Journal of Biological Chemistry. 1998;273:20685–20688. doi: 10.1074/jbc.273.33.20685. [DOI] [PubMed] [Google Scholar]
- 29.Ridley A. Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking. Trends in cell biology. 2006;16:522–529. doi: 10.1016/j.tcb.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 30.Hall A. Rho GTPases and the actin cytoskeleton. Science. 1998;279:509–514. doi: 10.1126/science.279.5350.509. [DOI] [PubMed] [Google Scholar]
- 31.Sit S, Manser E. Rho GTPases and their role in organizing the actin cytoskeleton. Journal of Cell Science. 2011;124:679–683. doi: 10.1242/jcs.064964. [DOI] [PubMed] [Google Scholar]
- 32.Mori Y, Jilkine A, Edelstein-Keshet L. Wave-pinning and cell polarity from a bistable reaction-diffusion system. Biophysical Journal. 2008;94:3684–3697. doi: 10.1529/biophysj.107.120824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mori Y, Jilkine A, Edelstein-Keshet L. Asymptotic and bifurcation analysis of wave-pinning in a reaction-diffusion model for cell polarization. SIAM J Applied Math. 2011;71:1401–1427. doi: 10.1137/10079118X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Xu J, Wang F, Van Keymeulen A, Herzmark P, Straight A, Kelly K, Takuwa Y, Sugimoto N, Mitchison T, Bourne H. Divergent signals and cytoskeletal assemblies regulate self-organizing polarity in neutrophils. Cell. 2003;114:201–214. doi: 10.1016/s0092-8674(03)00555-5. [DOI] [PubMed] [Google Scholar]
- 35.Sasaki A, Chun C, Takeda K, Firtel R. Localized Ras signaling at the leading edge regulates PI3K, cell polarity, and directional cell movement. The Journal of cell biology. 2004;167:505–518. doi: 10.1083/jcb.200406177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sasaki A, Janetopoulos C, Lee S, Charest P, Takeda K, Sundheimer L, Meili R, Devreotes P, Firtel R. G protein–independent Ras/PI3K/F-actin circuit regulates basic cell motility. The Journal of cell biology. 2007;178:185–191. doi: 10.1083/jcb.200611138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Calderwood D, Shattil S, Ginsberg M. Integrins and actin filaments: reciprocal regulation of cell adhesion and signaling. Journal of Biological Chemistry. 2000;275:22607–22610. doi: 10.1074/jbc.R900037199. [DOI] [PubMed] [Google Scholar]
- 38.Abram C, Lowell C. The ins and outs of leukocyte integrin signaling. Annual review of immunology. 2009;27:339. doi: 10.1146/annurev.immunol.021908.132554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Marée A, Jilkine A, Dawes A, Grieneisen V, Edelstein-Keshet L. Polarization and movement of keratocytes: A multiscale modelling approach. Bulletin of Mathematical Biology. 2006;68:1169–1211. doi: 10.1007/s11538-006-9131-7. [DOI] [PubMed] [Google Scholar]
- 40.Jilkine A, Marée AF, Edelstein-Keshet L. Mathematical model for spatial segregation of the Rho-Family GTPases based on inhibitory crosstalk. Bulletin of Mathematical Biology. 2007;69:1943–1978. doi: 10.1007/s11538-007-9200-6. [DOI] [PubMed] [Google Scholar]
- 41.Dawes A, Edelstein-Keshet L. Phosphoinositides and rho proteins spatially regulate actin polymerization to initiate and maintain directed movement in a one-dimensional model of a motile cell. Miophysical Journal. 2007;92:744–768. doi: 10.1529/biophysj.106.090514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Holmes W, Lin B, Levchenko A, Edelstein-Keshet L. Modelling cell polarization driven by synthetic spatially graded Rac activation. PLoS Comput Biol. 2012;8:e1002366. doi: 10.1371/journal.pcbi.1002366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.FitzHugh R. Mathematical models of excitation and propagation in nerve. Vol. 1. McGraw-Hill; New York: 1969. pp. 1–85. [Google Scholar]
- 44.Grieneisen V. Ph D thesis. University of Utrecht; 2009. Dynamics of Auxin Patterning in Plant Morphogenesis. [Google Scholar]
- 45.Marée A, Grieneisen V, Edelstein-Keshet L. How cells integrate complex stimuli: The effect of feedback from phosphoinositides and cell shape on cell polarization and motility. Plos Computational Biology. 2012;8 doi: 10.1371/journal.pcbi.1002402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nishiura Y. Global structure of bifurcating solutions of some reaction-diffusion systems. SIAM J Appl Math. 1982;13:555–593. [Google Scholar]
- 47.Rubinstein B, Slaughter B, Li R. Weakly nonlinear analysis of symmetry breaking in cell polarity models. Physical Biology. 2012;9:045006. doi: 10.1088/1478-3975/9/4/045006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Iron D, Ward M. A metastable spike solution for a nonlocal reaction diffusion model. SIAM J Appl Math. 2000;60:778–802. [Google Scholar]
- 49.Yang L, Dolnik M, Zhabotinsky A, Epstein I. Pattern formation arising from interactions between turing and wave instabilities. Journal of Chemical Physics. 2002;117:7259–765. [Google Scholar]
- 50.Wang W, Liu Q, Jin Z. Spatiotemporal complexity of a ratio-dependent predator-prey system. Phys Rev E. 2007;75:051913. doi: 10.1103/PhysRevE.75.051913. [DOI] [PubMed] [Google Scholar]
- 51.Schroth-Diez B, Gerwig S, Ecke M, Hegerl R, Diez S, Gerisch G. Propagating waves separate two states of actin organization in living cells. HFSP journal. 2009;3:412–427. doi: 10.2976/1.3239407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gerisch G. Self-organizing waves that simulate phagocytic cup structures. PMC Biophysics. 2010;3:7. doi: 10.1186/1757-5036-3-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Driscoll MK, McCann C, Kopace R, Homan T, Fourkas JT, Parent C, Losert W. Cell shape dynamics: From waves to migration. PLoS Comput Biol. 2012;8:e1002392. doi: 10.1371/journal.pcbi.1002392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Nicholson W. Linear algebra with applications. Vol. 275. PWS Publishing Company; 1995. [Google Scholar]
- 55.Dhooge A, Govaerts W, Kuznetsov Y. Matcont: A Matlab package for numerical bifurcation analysis of ODEs. ACM TOMS. 2003;29:141–164. [Google Scholar]