

Abstract
Glioblastoma cells secrete extra-cellular vesicles (EVs) containing microRNAs (miRNAs). Analysis of these EV miRNAs in the bio-fluids of afflicted patients represents a potential platform for biomarker development. However, the analytic algorithm for quantitative assessment of EV miRNA remains under-developed. Here, we demonstrate that the reference transcripts commonly used for quantitative PCR (including GAPDH, 18S rRNA, and hsa-miR-103) were unreliable for assessing EV miRNA. In this context, we quantitated EV miRNA in absolute terms and normalized this value to the input EV number. Using this method, we examined the abundance of miR-21, a highly over-expressed miRNA in glioblastomas, in EVs. In a panel of glioblastoma cell lines, the cellular levels of miR-21 correlated with EV miR-21 levels (p<0.05), suggesting that glioblastoma cells actively secrete EVs containing miR-21. Consistent with this hypothesis, the CSF EV miR-21 levels of glioblastoma patients (n=13) were, on average, ten-fold higher than levels in EVs isolated from the CSF of non-oncologic patients (n=13, p<0.001). Notably, none of the glioblastoma CSF harbored EV miR-21 level below 0.25 copies per EV in this cohort. Using this cut-off value, we were able to prospectively distinguish CSF derived from glioblastoma and non-oncologic patients in an independent cohort of twenty-nine patients (Sensitivity=87%; Specificity=93%; AUC=0.91, p<0.01). Our results suggest that CSF EV miRNA analysis of miR-21 may serve as a platform for glioblastoma biomarker development.
Introduction
Glioblastoma is the most common form of primary brain cancer and remains one of the deadliest of human cancers [1]. Timely diagnosis and sensitive therapeutic monitoring remain major challenges in the treatment of this disease. Clinically, response evaluations are largely based on clinical examination and Magnetic Resonance Imaging (MRI) [2]. However, both clinical examination and MRI are insensitive measures of disease status. For instance, the lowest resolution for reliable detection by MRI is on the order of millimeters [3]. Considering the size of a tumor cell, this limitation in resolution translates into a delay of at least ten cell divisions before therapeutic resistance can be detected [4]. While repeated post-treatment biopsies constitute a monitoring option, this practice is associated with significant morbidity [5,6]. In this context, less invasive platforms for therapeutic monitoring are needed.
Recent studies suggest that glioblastoma cells secrete extracellular vesicles (EVs) containing genetic materials that mirror the intracellular tumor milieu, including tumor specific mutations and patterns of RNA expression [7–11]. These EVs are released into the local environment and transgress anatomic compartments into cerebrospinal fluid (CSF) and the systemic blood circulation [7,8,12,13]. Analyses of these EVs offer a potential platform for monitoring tumor presence, phenotypic/genotypic features, and pathophysiology [7,9,14,15]. The EVs shelter the tumor-specific genetic material from the extracellular environment that is replete with RNAses [16,17] and preserve the integrity of these materials. Importantly, the genetic materials within EVs appear highly enriched for RNAs in the size range of microRNAs (miRNAs) [9].
Since non-neoplastic cells secrete EVs [18–20] and they out-number neoplastic cells by several orders of magnitude in a patient [21], tumor-specific EVs remain a rarity in clinical samples [22]. As such, sensitive methods of amplification, such as quantitative polymerase chain reactions (qPCR), are required for detection and quantitation of RNA in EVs [7,8,23]. In the frequently used relative CT method of qPCR [24], the cellular abundance of a “query” transcript is assessed by determining the number of cycles of PCR required to reach a threshold quantity [25]. To correct for the quality and quantity of the input material, this cycle number is then normalized to the PCR cycles required for a reference transcript to reach the same threshold [26]. This method assumes abundance and near-homogeneity of the reference transcript level per cell [27,28].
Here we report that the commonly used reference transcripts for quantitation of cellular RNA, including GAPDH, 18S rRNA, and hsa-miR-103, are present in extremely low and highly varied levels within EVs (on the order of 1 copy per 103 to 104 EVs). Moreover, their relative abundance in EVs bears no correlation to the total EV number or RNA yield. In this context, we characterized the abundance of EV miRNAs in absolute terms and normalized this value to the input EV particle numbers. Using this method, we studied the level of hsa-miR-21, a miRNA that is highly over-expressed in glioblastoma cells [29]. Our results indicate that miR-21 levels were significantly elevated in EVs isolated from the CSF of glioblastoma patients relative to those derived from non-oncologic patients.
Materials and Methods
Clinical specimen collection
All research performed were approved by IRB boards at University of California, San Diego Human Research Protections Program and were in accordance with the principles expressed at the declaration at Helsinki. Each patient was consented by a dedicated clinical research specialist prior to collection. Written consent was obtained for each patient. The consent process was approved by the ethics committee, and all records were documented in our electronic record system. The written consent from patients was also scanned into our electronic filing system. The serum and CSF specimens for the initial, exploratory studies were collected at the University of California San Diego Medical Center under IRB 120345X. The sera collection and an initial training CSF collection were performed by CCC and BSC at the time of surgical procedure. The CSF was collected by ventricular/lumbar drain placement or cisternal aspiration at the time of craniotomy. Blood was collected using an 18 Gauge-needle venipuncture into clot-activating blood collection tubes with gel separator (BD vacutainer catalog #366450). Attention was paid to minimize mechanical tube agitation. The samples were processed by spinning at 1,500 x g within 30 minutes of collection and the snap frozen [30]. The clinical diagnosis of the non-oncologic patients who contributed blood samples were: severe head trauma (n=2), subarachnoid hemorrhage (n=2), and normal pressure hydrocephalus (n=1). The clinical diagnosis of the non-oncologic patients who contributed CSF samples were: trauma (n=2), subarachnoid hemorrhage (n=8), normal pressure hydrocephalus (n=2), arteriovenous malformation (n=2). All diagnoses of glioblastoma were histologically confirmed. CSF specimens were snap frozen in -80 upon receipt and analyzed without further centrifugation. The CSF specimens for the validation study were generously provided by Dr. Santosh Kesari (University of California, San Diego; IRB 110551X) and Dr. Erwin Van Meri (Emory University, IRB 642-2005). 1-5 mL of CSF was collected from each patient through lumbar or ventricular puncture.
EV free media preparation
EV-depleted medium was prepared by ultracentrifugation of DMEM supplemented with 20% FBS at 120,000 x g for 18 hours at 4°C. The medium was then diluted to a final concentration of 10% FBS and used to culture cell lines as described.
Cell lines and cell culture
Eleven human glioblastoma cell lines (A1207, A172, LN18, LN340, LN464, T98G, U118, U373, U87MG, LN229, and LN235) [31] and 3 non-glioblastoma cell lines (A549, U20S, and 293T) [32–34] were cultured in DMEM supplemented with 10% FBS. At 60-70% confluency, the standard culture medium was replaced with EV depleted medium. The cells were cultured for an additional 72 hours before EV collection from the cell-free supernatants. 9 neurosphere lines (1123, 326, 83, 30, AC17, AC20, 84, BT70, and CMK3) were cultured in DMEM F12 supplemented with growth factors as described previously [35–37]. Cell free supernatant was collected three days after culturing for EV isolation.
Extracellular vesicle (EV) Isolation
The effect of freeze-thaw cycle on the stability of vesicular content has not been well studied. In order to avoid degradation of vesicles, all samples were processed promptly upon thawing of serum and CSF samples. The EV fraction was isolated by differential centrifugation [38]. Briefly, conditioned media or diluted bio-fluids were first centrifuged at 2,000 x g for 20 minutes to remove cellular debris. The supernatant was collected and further centrifuged at 10,000 x g for 30 minutes. The resultant supernatant was then transferred to ultracentrifuge tubes for ultracentrifugation at 120,000 x g for 2 hours. The supernatant was discarded and the EV pellets were re-suspended in 150μL of PBS for storage at -80°C prior to RNA isolation. All centrifugation steps were performed at 4°C. This protocol was designed to enrich for EVs in the 50-250 nm size range (Figure 5 ).
Figure 5. Discrimination of glioblastoma disease status by CSF EV miR-21 analysis.

(A) Comparable levels of miR-21 level in sera EVs derived from 24 glioblastoma patients and 5 non-oncologic patients. (B) Elevated level of miR-21 in CSF EV derived from 13 glioblastoma patients relative to 14 non-oncologic patients. (C) miR-21 is detected in CSF EV but not in EV depleted CSF in samples isolated from five independent glioblastoma patients. Patient 1, 2, 3, 4, and 5 correspond to T2, T7, T10, T12, and T13 respectively from the UCSD glioblastoma cohort.
EV Quantification and Assessment
The number of vesicles recovered was determined by Nanoparticle Tracking Analysis (NTA) on a Nanosight LM-10HS in accordance to the manufacturer’s instructions (Nanosight, Wiltshire, UK). Resuspended vesicles were diluted 1:40 to 1:200 with PBS before analysis. The purity of the EV isolated were assessed using electron microscopy as previously described [38] (Figure 1 ).
Figure 1. Relative abundance of GAPDH, 18S rRNA, and miR-103 in glioblastoma cell line derived EVs.

(A) GAPDH, 18S rRNA, and hsa-mir-103 levels in EVs isolated from 11 glioblastoma and 3 non-glioblastoma adherent cell lines, cultured under serum conditions. Transcript copy number was plotted against total RNA yield extracted from the EVs. (B) GAPDH, 18S rRNA, and hsa-mir-103 transcript copy number was plotted against the total number of EVs isolated for RNA extraction. (C) Cellular GAPDH transcript number tightly correlated with the number of cells collected for RNA extraction and (D) the amount of RNA recovered.
Quantitative Reverse Transcriptase-Polymerase Chain Reaction (qRT-PCR)
RNA from the EV fraction was extracted using the mirRCURY RNA Isolation Kit (Exiqon, Vedbaek, Denmark) per manufacturer’s protocol. RNA concentration and quality were determined using the NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Waltham, MA). For profiling GAPDH, 18S rRNA, miR-21, and miR-103 expression, cDNA was synthesized with the miRCURY LNATM Universal RT microRNA PCR system (Exiqon). The resultant cDNA were diluted 20x for qRT-PCR. (See Table 1 for primer information).
Table 1. Abundance of GADPH mRNA, 18s RNA, and hsa-miR-103 in cultured cell lines and EVs derived from cultured cell lines.
|
GAPDH
|
18S RNA
|
miR-103
|
|||||
|---|---|---|---|---|---|---|---|
| Cell line | Copies/cell | Copies per EV | Copies/cell | Copies per EV | Copies/cell | Copies per EV | |
| Adherent lines | U87MG | 6.97 x 102 | 1.14 per 105 | 1.29 x 106 | 1.71 per 103 | 8.31 x 103 | 2.13 per 104 |
| T98G | 3.21 x 102 | 2.02 per 106 | 2.80 x 105 | 1.87 per 10 | 4.42 x 104 | 7.23 per 104 | |
| LN229 | 4.52 x 102 | 1.46 per 103 | 1.09 x 105 | 1.81 per 102 | 1.02 x 105 | 6.74 per 103 | |
| LN464 | 3.27 x 102 | 8.63 per 107 | 1.21 x 106 | 6.36 per 104 | 8.65 x 104 | 1.26 per 103 | |
| A172 | 2.91 x 102 | 2.58 per 106 | 9.32 x 105 | 1.35 per 104 | 1.90 x 104 | 4.47 per 104 | |
| LN340 | 5.10 x 102 | 6.24 per 105 | 5.13 x 105 | 1.28 per 103 | 2.58 x 104 | 2.82 per 104 | |
| A549 | 1.03 x 103 | 1.30 per 104 | 1.34 x 106 | 3.19 per 104 | 3.92 x 104 | 3.35 per 104 | |
| U2OS | 9.06 x 102 | 1.84 per 104 | 1.41 x 106 | 3.84 per 104 | 7.42 x 104 | 6.79 per 104 | |
| Neurospheres | 1123 | 3.96 x 102 | 3.93 per 106 | 3.99 x 106 | 5.75 per 105 | 3.68 x 103 | 2.47 per 104 |
| 326 | 5.14 x 102 | 3.36 per 105 | 6.36 x 106 | 6.38 per 103 | 2.13 x 103 | 6.95 per 104 | |
| 83 | 5.62 x 102 | 6.39 per 106 | 6.63 x 106 | 2.63 per 103 | 2.15 x 103 | 9.23 per 105 | |
| 30 | 3.60 x 102 | 9.24 per 106 | 4.66 x 106 | 1.74 per 103 | 5.16 x 103 | 8.42 per 104 | |
| AC17 | 2.92 x 102 | 9.00 per 106 | 9.96 x 105 | 3.85 per 105 | 3.24 x 103 | 6.82 per 105 | |
| AC20 | 1.42 x 102 | 1.08 per 105 | 6.13 x 105 | 5.01 per 104 | 7.62 x 103 | 4.03 per 104 | |
| 84 | 2.37 x 102 | 2.93 per 105 | 1.03 x 106 | 1.40 per 103 | 3.41 x 103 | 8.16 per 105 | |
| BT70 | 5.30 x 102 | 5.43 per 105 | 2.23 x 106 | 3.88 per 104 | 4.11 x 103 | 3.36 per 105 | |
| CMK3 | 2.12 x 102 | 2.95 per 104 | 6.72 x 105 | 2.41 per 103 | 6.53 x 103 | 1.75 per 104 | |
Determination of copy number
To determine the absolute copy numbers of GAPDH and 18S RNA in the cellular cytoplasm and in the EVs, a standard curve for each gene was generated using serial dilutions of known quantity of U87MG genomic DNA. The copy number of GAPDH and 18S RNA within the U87 genome was previously determined by integrated sequencing efforts and comparative genomic hybridization [39]. To determine copy number of miRNA (miR-103, miR-21), standard curves were generated by serial dilution of known quantities of miRNA mimic (Qiagen, Germantown, MD), followed by cDNA synthesis using the miRCURY LNATM Universal RT microRNA PCR system under conditions that allow one round of cDNA synthesis [40]. The cDNAs were then used for the generation of standard curve.
Statistical Analysis
All statistical analyses were performed using the GraphPad Prism Software version 5 (GraphPad, La Jolla, CA). Correlation between GAPDH, 18S RNA, and miR-103 transcript level, EV RNA yield, and EV numbers were determined using Pearson’s Correlation [41]. Difference in hsa-miR-21 level between glioblastoma and non-oncologic clinical samples was determined using the student’s t-test (2 tailed) [42]. Sensitivity, specificity, positive predictive value (PPV), and negative predictive value (NPV), Receiver Operating Characteristic (ROC) curve and Area Under the Curve (AUC) were determined as previously described [43].
Results
GAPDH, 18S rRNA, and hsa-miR-103 levels in glioblastoma cell line derived EVs
Housekeeping genes, such as GAPDH, 18S rRNA, and miR-103, are expressed at high levels across cell types and exhibit little cell-to-cell variability relative to most query genes. As such, they are considered well-validated reference genes in the expression profiling of cellular contents [27,28,44]. The absolute copy number of these transcripts in EVs, however, remains poorly characterized. Since the mechanisms by which genetic materials are transported from the cell into the EVs remain largely unknown [45], it is conceivable that EV transcript levels differ from their cellular expression levels. Despite this knowledge gap, GADPH mRNA, 18s rRNA, and miR-103 transcripts are frequently used as reference genes for quantitative analysis of EV-derived genetic materials [46–48] via the relative CT qPCR method [49–53].
We first determined the relative abundance of GAPDH mRNA, 18S rRNA, and hsa-miR-103 in EVs derived from 6 glioblastoma and 2 non-glioblastoma adherent cell lines. All three transcripts were abundant in the cellular cytoplasm, with copy numbers that ranged from 300 to 1,000 copies per cell for GAPDH, 100,000 to 1,000,000 copies for 18S rRNA, and 8,000 to 80,000 copies for hsa-miR-103 (Table 1 ). These copy numbers are comparable to those previously published [54–57].
We next analyzed the level of reference transcripts in EVs secreted by these adherent cell lines. EVs were isolated by ultracentrifugation and their numbers were determined by Nanoparticle Tracking Analysis [58–61]. While all three transcripts were detectable, their abundance in the EVs were approximately 105 -1010 fold lower than the levels detected in the cellular cytoplasm. At most, an average of one copy of GAPDH transcript was detected in 1,000 EVs; an average of one 18S rRNA transcript was found in 40 EVs; an average of one copy of miR-103 transcript was present in 750 EVs (Table 1 ).
To determine whether reference housekeeping genes would serve as surrogate markers for the total RNA yield or the total number of EVs, we analyzed the levels of GAPDH mRNA, 18S rRNA, and hsa-miR-103 in EVs isolated from 11 glioblastoma and 3 non-glioblastoma adherent cell lines (cultured under serum conditions). We observed that the level of the potential reference transcripts varied by at least an order of magnitude in EVs isolated from different lines. The expression of the 18S rRNA and hsa-miR-103 demonstrated low correlation with EV RNA yield (Figure 1A ) or EV number (Figure 1B ). While EV GADPH transcript level demonstrated correlation with EV RNA yield and number, these correlations were not as robust as those seen between cellular GAPDH transcript level and cell number (Figure 1C ) or the RNA yield from cell extracts (Figure 1D ).
As cell lines cultured under neurosphere and serum-free conditions may better recapitulate certain aspects of glioblastoma biology [62,63], we quantitated the three potential reference transcripts in the EVs isolated from nine glioblastoma neurosphere lines [35–37]. While GAPDH mRNA, 18S rRNA, and hsa-miR-103 were abundant in the cellular cytoplasm, their abundance in the EVs was 108 fold less relative to the cellular content (Table 1 ). In contrast to the results obtained using adherent glioblastoma lines, GADPH mRNA levels in EVs derived from neurosphere lines did not correlate with EV RNA yield (Figure 2A ) or EV number (Figure 2B ). Instead, correlations were observed between EV hsa-miR-103 and 18s rRNA and EV number/EV RNA yield (Figures 2A and 2B ). Careful analysis of the correlation plot, however, suggests that the correlations may have been driven by a small subset of samples. In EVs derived from cultured cells, there was a strong agreement between RNA yield from EV preparation and the total EV number (R2=0.74, p=0.003) (Figure 3A ).
Figure 2. Relative abundance of GAPDH, 18S rRNA, and miR-103 in neurosphere glioblastoma line derived EVs.

(A) GAPDH, 18S rRNA, and hsa-mir-103 levels in EVs isolated from 9 glioblastoma neurophere lines, cultured under serum-free conditions. Transcript copy number was plotted against total RNA extracted from the EVs. (B) GAPDH, 18S rRNA, and hsa-mir-103 transcript copy number was plotted against the total number of EVs isolated for RNA extraction.
Figure 3. Relative abundance of GAPDH, 18S rRNA, and miR-103 in EV derived from CSF of glioblastoma and non-oncologic patients.

(A) GAPDH, 18S rRNA, and hsa-mir-103 levels in EVs isolated from the CSF of 13 glioblastoma and 14 non-oncologic patients. Transcript copy number was plotted against total RNA extracted from the EVs. (B) GAPDH, 18S rRNA, and hsa-mir-103 transcript copy number was plotted against the total number of EVs isolated for RNA extraction. The relative abundance of these transcripts in CSF EVs was approximately 10 fold lower than those of sera EV with abundance ranging between 1 transcript in 1 EV to 1 transcript in 105 EVs.
GADPH mRNA, 18S rRNA, and hsa-miR-103 levels in EVs derived from patient sera and cerebrospinal fluid (CSF)
We next tested whether the abundance of GAPDH mRNA, 18S rRNA, and hsa-miR-103 correlated with RNA yield or EV number in clinical specimens from glioblastoma patients. The purity of the EVs isolated from clinical sera or CSF was verified by electron microscopy (Figure 1 ). There was no convincing or consistent correlation between GADPH, 18s rRNA, or miR-103 transcripts and EV particle number in serum (Figure 2 ) or CSF (Figure 3 ). There was little correlation between EV number and RNA yield in EVs derived from sera derived EVs (R2=0.14, p=0.04). Modest correlation between RNA yield and EV number was observed in CSF derived EVs (R2=0.60, p<0.001; Figure 3B and C ).
In summary (Table 2 ), EV GAPDH mRNA levels correlated with EV numbers in glioblastoma lines cultured under adherent conditions, but this correlation was not observed in lines cultured under neurosphere conditions. On the other hand, EV miR-103 and 18s RNA levels correlated with EV number in glioblastoma lines cultured under neurosphere conditions, but not in lines cultured under adherent conditions. In clinical sera or CSF, there was no correlation between these reference transcripts and EV numbers. These discrepancies create significant uncertainties as to the appropriate reference transcript for quantitative EV analysis in clinical bio-fluids. Given these discrepancies and the lack of correlation between EV GAPDH mRNA, 18S rRNA, or miR-103 and EV number/EV RNA yield, we propose that EV RNA be quantitated in absolute terms and normalized to the input EV number. We reasoned that this method is analogous to determining absolute quantities of cellular miRNA and normalizing to the input cell number. Our proposed method prevents arbitrary changes in miRNA quantitation secondary to random fluctuation in the levels of the reference transcript.
Table 2. Overview of robustness of reference transcripts in EVs derived from cell lines and bio-fluids.
|
Correlation with RNA yield
|
Correlation with EV number
|
|||||
|---|---|---|---|---|---|---|
| Sample type | GAPDH | 18S rRNA | miR-103 | GAPDH | 18S rRNA | miR-103 |
| Adherent lines | + | - | - | - | - | - |
| Neurosphere lines | - | + | + | - | + | + |
| Serum | NT | NT | NT | - | - | - |
| Cerebrospinal Fluid | - | - | - | - | - | - |
No correlation
Significant correlation (R2 >0.5 and p<0.05)
NT: Not testedmiR-21 levels in EVs derived from glioblastoma cell lines
Using absolute measurements of miRNA normalized to input EV numbers, we characterized the levels of miR-21, a highly over-expressed miRNA in glioblastoma [29], in EVs isolated from glioblastoma neurosphere lines. We reasoned that if EV miRNA contents reflect the internal milieu of the secreting cell [34], then miR-21 should be one of the more abundant species in glioblastoma EVs. Indeed, microarray based miRNA profiling of EVs derived from two glioblastoma cell lines revealed that miR-21 is one of the most abundant miRNAs in glioblstoma EVs [7].
We first characterized EV miR-21 using nine glioblastoma neurosphere lines. Consistent with published literature, miR-21 was highly expressed in these lines, with quantities that ranged from 700 to 20,000 copies per cell (Figure 4A , Table 3 ). miR-21 levels per EV were significantly lower, ranging from 1 copy per 100 to 2,000 EVs (Figure 4B ). Nevertheless, miR-21 was consistently detectable in EVs isolated from all nine lines. Moreover, the EV miR-21 level correlated with the cellular miR-21 level (R2=0.50; p<0.05), suggesting that EV miRNA analysis may afford a window into the physiology of the secreting cell (Figure 4C ).
Figure 4. Detection of miR-21 in glioblastoma cell lines secreted EVs.
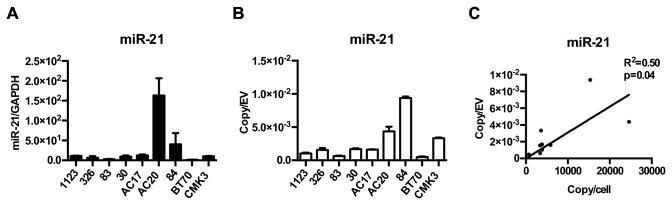
(A) The expression level of miR-21 was quantitatively assessed in nine neurosphere glioblastoma lines. (B) The abundance of miR-21 was assessed in EVs derived from nine neurosphere glioblastoma lines. (C) EV miR-21 levels correlate tightly with cellular miR-21 levels (R2 = 0.50, p<0.01).
Table 3. Abundance of miR-21 in cultured neurosphere lines and neurosphere derived EVs.
|
miR-21
| ||
|---|---|---|
| Cell line | Copies/cell | Copies per vesicle |
| 1123 | 3.87 x 103 | 1.05 per 103 |
| 326 | 3.36 x 103 | 1.57 per 103 |
| 83 | 3.40 x 103 | 6.28 per 104 |
| 30 | 3.83 x 103 | 1.68 per 103 |
| AC17 | 5.86 x 103 | 1.60 per 103 |
| AC20 | 2.46 x 104 | 4.37 per 103 |
| 84 | 1.54 x 104 | 9.38 per 103 |
| BT70 | 7.14 x 102 | 4.80 per 104 |
| CMK3 | 3.60 x 103 | 3.34 per 103 |
We also examined the effect of the culturing condition on the abundance of miR-21 in EVs. To this end, the CMK-3 neurosphere cell line was adapted to growth as adherent culture in the presence of fetal bovine serum. This experimental manipulation led to a nine-fold increase in the cellular level of miR-21. However, the abundance of miR-21 in secreted EVs was decreased by approximately ten fold (Figure 6 ). In a reciprocal experiment, adherent U87MG cells were cultured under neurosphere conditions. This adaptation led to decreased abundance of miR-21 in both cellular and EV contents. These results suggest that glioblastoma growth under neurosphere conditions is associated with decreased cellular levels of miR-21. While culturing conditions also significantly impact EV miR-21 level, these effects are less predictable.
Figure 6. Elevated levels of CSF EV miR-21 are detected in glioblastoma patients.

(A) Sensitivity, specificity, positive predictive, and negative predictive values associated with EV miR-21 level of <0.25 copy/EV as a discriminating threshold for glioblastoma disease status in the initial exploratory study. (B) Sensitivity, specificity, positive predictive, and negative predictive values associated with EV miR-21 level of <0.25 copy/EV as a discriminating threshold for glioblastoma disease status in a validation study. (C) Receiver Operating Characteristic Curve for EV miR-21 level of <0.25 copy/EV as a discriminating threshold for glioblastoma disease status. (D) Level of miR-21 in CSF EV from a glioblastoma patient at time of surgery and three months after a gross-total resection. The relative abundance of miR-21 in CSF EV was decreased by approximately 50-fold after surgical resection.
miR-21 levels in EVs isolated from glioblastoma and non-oncologic patients
Since miR-21 is present in glioblastoma secreted EVs, if these EVs eventually reach bio-fluids such as blood or CSF, then the elevated levels of EV miR-21 in these bio-fluids may serve to indicate the presence of tumor. This hypothesis pre-supposes that EVs secreted from normal cells harbor significantly lowered levels of miR-21 relative to those of glioblastoma EVs. As a first step toward testing this hypothesis, we determined whether miR-21 levels in clinical bio-fluids (sera and CSF) derived from glioblastoma patients differed from those of non-oncologic patients.
Since EVs bearing tumor specific RNAs have been detected in blood [7], we tested whether miR-21 levels differed in EVs derived from the sera of 24 glioblastoma patients differed from those of 5 non-oncologic patients (2 trauma, 2 subarachnoid hemorrhage, 1 normal pressure hydrocephalus). On average, 1.96x109 to 1.19x1011 EVs were isolated from 2-5cc of sera. The level of EV miR-21 ranged from 0.21 to 10.35 copies/EV in the glioblastoma patients and 0.21 to 14.30 copies/EV in the non-oncologic patients. No statistically significant difference was detected between these two groups (Figure 5A ).
We hypothesized that the secretion of EV miR-21 by hematopoietic cells [64] might obscure glioblastoma EV miR-21 in sera. Thus, we analyzed miR-21 levels in EVs from CSF of glioblastoma (n=13) and non-oncologic patients (n=14). CSF EV miR-21 ranged from 0.14 to 1.04 copies/EV in glioblastoma patients and 5.26x10-4 to 1.48x10-1 copies/EV in non-oncologic patients. The difference between these two groups was statistically significant at p<0.001 (Figure 5B ), suggesting that CSF EV may be a platform for therapeutic monitoring of tumor presence. We also noted that the bulk of miR-21 is located within EVs rather than freely floating in CSF, <10% of total CSF miR-21 (Figure 5C ) was detected in the EV depleted CSF fraction in samples collected from five independent glioblastoma patients. In our analysis, we noted that all of the CSFs derived from non-oncologic patients harbored EV miR-21 level of < 0.25 copy/EV. Of note, the number of EVs as well as the size distribution of EVs isolated from the bio-fluids of glioblastoma patients did not significantly differ from those isolated from non-oncologic patients (Figures 4 and 5 ).
CSF EV miR-21 analysis in an independent cohort
We next wished determined whether such a threshold can be utilized to discriminate CSF derived from glioblastoma patients from those of non-oncologic patients in an independent cohort. In our initial cohort, the sensitivity, specificity, positive predictive value (PPV), and negative predictive value (NPV) associated with EV miR-21 level of <0.25 copy/EV were 85%, 100%, 100%, and 93%, respectively (Figure 6A ). We calculated a need for CSF from 30 additional patients (with 15 glioblastoma patients and 15 non-oncologic patients) for statistical testing of our hypothesis, with type I error set at 0.05 and a power of 0.8 [65].
Through a collaborative network with Emory University and the University of California San Diego Neurology programs, 29 additional CSF samples were secured (with 15 glioblastoma specimens and 14 non-oncologic specimens). EVs were isolated from these specimens, and miR-21 levels were determined. In this second and independent cohort, setting EV miR-21 level of <0.25 copy/EV as the discriminating threshold for glioblastoma versus non-oncologic patients yielded sensitivity, specificity, PPV, and NPV of 87%, 93%, 87%, and 86%, respectively (Figure 6B ). Receiver Operating Characteristics (ROC) curves for CSF EV miR-21 as a biomarker of glioblastoma tumor presence show an AUC of 0.9 and p<0.01 (Figure 6C ). In aggregate, these results support CSF EV miR-21 as a feasible biomarker for the presence of glioblastoma.
In aggregate, these results suggest that CSF EV miR-21 constitutes a biomarker for the presence of glioblastoma cells. A corollary of this prediction is that surgical excision of glioblastoma should be associated with a decrease in CSF EV miR-21 level. We tested this prediction in a patient who underwent CSF sampling at the time of glioblastoma surgery and then at three months after a gross-total resection. Consistent with our hypothesis, the relative abundance of miR-21 in CSF EV was decreased by approximately 50-fold after surgical resection (Figure 6D ).
Discussion
Analysis of genetic material within glioblastoma secreted within EVs in bio-fluids represents a unique opportunity for diagnosis and therapeutic monitoring. EVs isolated from bio-fluids can encompass exosomes, microvesicles, retrovirus-like particles (RLPs) and apoptotic bodies [66,67]. While the mechanisms of their biogenesis may differ, genetic materials from their originating cells have been detected in all types of vesicles. Enrichment of miRNA in EV content [9] renders EVs a particularly attractive platform.
The quantitative assessment of EV miRNAs requires thoughtful considerations. Our study reveals that commonly used reference transcripts (GADPH mRNA, 18S rRNA, and hsa-miR-103) were present at extremely low and variable levels in EVs. Moreover, there were no consistent correlations between the level of these transcripts and EV RNA yield or EV particle numbers (Figure 3 and Figure 2 ). When comparing the EV particle numbers and the total amount of RNA extracted, we noted a strong correlation in systems such where there is only one source of EV secreting cell (e.g. neurosphere culture; Figure 3A ). Under these conditions, RNA yield serves as an effective normalization method in place of EV number (Figure 7A and 7B ). However, in more complex samples such as patient CSF, where EVs are likely secreted by multiple cells of origin, the correlation between EV number and total RNA yield are significantly lowered (Figure 3C ). Because of these observations, we recommend quantifying EV miRNA in absolute terms and normalizing to total EV number.
Using this method, we showed that EV miR-21 levels can be used to differentiate CSF isolated from glioblastoma patients and CSF from non-oncologic patients. Importantly, our result was validated using multiple independent CSF collections. It is important to note that CSF is not routinely collected during the course of treatment for glioblastoma patients in the U.S. Thus, while the total number of CSF samples analyzed here is relatively small, with a total of 28 glioblastoma CSF and 28 non-oncologic CSF samples analyzed, our study represents an exhaustive analysis of CSF specimens collected by four independent investigators over a multi-year period. The results presented here provide a sound basis for a prospective, multi-center study to validate CSF EV miR-21 as a biomarker for assessing the presence of glioblastoma.
Recent studies suggest that circulating miRNA can exist in two compartments: 1) outside of EV where they complex with Argonaut2, the catalytic component of the RNA-induced silencing complex (RISC) or high density lipoproteins; and 2) within EVs [68,69]. We add the finding that the majority of CSF miR-21 is of EV origin. Additionally, the available data suggests that genetic material within EVs are quite stable in the CSF [70]. Thus, isolating CSF EV may enhance the sensitivity of CSF miRNA based biomarker assays.
miR-21 was previously reported as a CSF biomarker for glioblastoma burden in a prior study [70], While our overall conclusion is generally consistent with this previous study, it is worthwhile noting the differences between the two. First, Teplyuk et. al. [70] measured the level of miR-21 in total CSF and not CSF EVs. Second, in the previous study, quantitative assessment of miRNA in the CSF was performed using the relative CT methods using miR-24 and miR-125 as reference transcripts. We tested whether miR-24 and miR-125 are adequate reference transcripts for quantitative EV miRNA analysis. While we observed modest to strong correlations between miR-24/miR-125 expression and EV RNA yield and/or total EV number in cells cultured under neurosphere conditions (Figure 8A ), such correlations were not found in clinical CSF specimens (Figure 8B ). Using criteria established in our study, these results would suggest that miR-24 and miR-25 are not adequate reference transcripts for the quantitative miRNA analysis of CSF EVs. Consistent with this proposition, Teplyuk et. al. reported that miR-24 and miR-25 were present in relatively low levels and varied between clinical specimens [70].
miR-21 expression is not unique to glioblastoma, it has been detected in endothelial cells [71], normal breast tissue [72], cervical tissue [73], and hematopoietic cells [74]. While the expression level of miR-21 in glioblastoma cells are generally one to two orders of magnitude higher than those found in normal tissue [34], it is certainly conceivable that the normal cellular secretion of EV miR-21 may mask the presence of a small number of glioblastoma EVs containing miR-21. When EVs isolated from sera were examined, this appeared to have been the case, as no significant differences in EV miR-21 levels in could be detected between patients with and without glioblastoma (Figure 5A ). However, Wang et. al. previously reported that plasma levels of miR-21 were significantly altered in glioblastoma patients compared to normal controls [75]. In the context of our study, it is possible that EV independent miR-21 significantly differed between glioblastoma and non-oncology patients. It is also worthwhile noting the technical difference in the method of miRNA quantitation between our study and this previous study [75]. In the previous study, Wang et al. [75] quantitated miR-21 using the relative CT method, with murine miR-29 as spike-in control. In contrast, our study quantitated miR-21 in absolute terms and normalized to the number of EVs. Both methods harbor inherent advantages and disadvantages [76]. The optimal method of miRNA quantitation in bio-fluids remains an unresolved issue and may be bio-fluid dependent.
In contrast to our sera derived results, miR-21 levels in the CSF of non-oncologic patients were extremely low by comparison. The amount of miR-21per EV derived from the CSF of glioblastoma patients was, on average, ten-fold higher than those derived from non-oncologic patients (Figure 5B ). These results suggest that the normal tissues in contact with CSF secrete less miR-21/EV as compared to brain tissue harboring glioblastoma cells.
Our studies raised several important questions. Although EVs in CSF appear to be enriched for miR-21, we have little data on the sub-speciation of these EVs in terms of size, zeta potential, shape, or type [67]. Equally uncertain is the origin of CSF EV miR-21, which may be from glioblastoma or associated endothelial, ependymal or inflammatory cells [77,78]. To the extent that we consistently observed the secretion of miR-21 containing EVs from all glioblastoma cells examined, we propose that these vesicles contribute to the presence of hsa-miR-21 in the CSF. However, this remains a point of uncertainty. Finally, with the emergence of the concept of glioblastoma subtypes [79], whether subtyping can be achieved through CSF EV miRNA analysis remains an important question,
In sum, our study delineates miR-21 levels in absolute terms as number of copies per EV and provides data suggesting the utility of CSF EV miRNA analysis as a biomarker for glioblastoma patients. It is likely that other glioblastoma EV miRNAs [80] may be similarly exploited as diagnostic biomarkers.
Supporting Information
Electron micrograph of EVs isolated from CSF. EVs were isolated from CSF by differential centrifugation and analyzed by transmission electron microscopy. Scale bar represents 200nm in (A) and 100nm in (B). EVs in the size range of 50-250nm were observed.
(EPS)
Relative abundance of GAPDH, 18S RNA, and miR-103 in EV derived from sera of glioblastoma and non-oncologic patients. GAPDH, 18S rRNA, and hsa-mir-103 transcript copy number was plotted against the total number of EVs isolated for RNA extraction. The relative abundance of these transcripts in EV ranged between 1 transcript in 7 EVs to 1 transcript in 106 EVs.
(EPS)
Correlation between EV number and RNA yield. For each sample, the RNA yield from EV preparations for (A) neurosphere cell lines, (B) Serum samples, and (C) CSF samples were plotted on the X-axis; the input EV number for these two datasets were plotted on the Y axis. There was excellent correlation between the RNA yield and the EV preparation in neurosphere lines, the correlation was poor in serum samples.
(EPS)
Abundance of EVs per cc of biofluids. (A) Abundance of EVs per cc of serum in glioblastoma and non-oncogenic patients. There was no significant difference between tumor and control samples. (B) Abundance of EVs per cc of CSF in glioblastoma and non-oncogenic patients. No significant differences were observed.
(EPS)
Average size profile of EVs isolated from biofluids. (A) Average size distribution of EVs isolated from serum of glioblastoma and non-oncogenic patients. There was no significant difference between tumor and control samples. (B) Average size distribution of EVs isolated from the CSF of glioblastoma and non-oncogenic patients. No significant size differences were observed.
(EPS)
Effects of culturing condition on miR-21 expression in cells and EVs. Glioblastoma cell lines U87MG and CMK3 were grown under adherent or neurosphere condition and the miR-21 level in cells or EVs were quantitated.
(EPS)
Normalization of EV miR-21 by RNA yield. (A) The expression level of miR-21 was quantitatively assessed in nine neurosphere glioblastoma lines using total RNA yield as normalization parameter. (B) Comparison of normalization method revealed good correlation between normalizing by EV particle number or total RNA yield in cell culure. (C) Levels of miR-21 in EVs isolated from glioblastoma and non-oncologic patients normalized by RNA yield. (D) Correlation between normalization by EV number or total RNA yield in CSF.
(EPS)
Relative abundance of miR-24 and miR-125 in EVs derived from neurosphere glioblastoma cell lines and CSF of glioblastoma and non-oncologic patients. has-miR-24 and has-miR-125 levels in EV isolated from from (A) 9 glioblastoma neurophere lines, cultured under serum-free conditions or (B) from CSF of 13 glioblastoma and 14 non-oncologic patients were plotted against total RNA extracted from the EVs or against the total number of EVs isolated for RNA extraction.
(EPS)
List of primers used.
(DOCX)
Acknowledgments
We would like to thank Jie Li, Ying Shen, David Gonda, Diahnn Futalan, Tyler Steed and Zack Taich for critical reading of the manuscript and helpful advice.
Funding Statement
CCC is supported by the Doris Duke Charitable Foundation http://www.ddcf.org/, Sontag Foundation http://www.sontagfoundation.com/display.aspx?page=home, Burroughs Wellcome Fund http://www.bwfund.org/, Forbeck Foundation http://wgfrf.org/, and Kimmel Foundation http://www.kimmelfoundation.org/. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Van Meir EG, Hadjipanayis CG, Norden AD, Shu HK, Wen PY et al. (2012) Exciting new advances in neuro-oncology: the avenue to a cure for malignant glioma. CA Cancer J Clin 60: 166-193. PubMed: 20445000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Grossman SA, Batara JF (2004) Current management of glioblastoma multiforme. Semin Oncol 31: 635-644. doi: 10.1053/j.seminoncol.2004.07.005. PubMed: 15497116. [DOI] [PubMed] [Google Scholar]
- 3. Sorensen AG, Batchelor TT, Wen PY, Zhang WT, Jain RK (2008) Response criteria for glioma. Nat Clin Pract Oncol 5: 634-644. doi: 10.1038/ncponc1204. PubMed: 18711427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Teodori L, Albertini MC, Uguccioni F, Falcieri E, Rocchi MB et al. (2006) Static magnetic fields affect cell size, shape, orientation, and membrane surface of human glioblastoma cells, as demonstrated by electron, optic, and atomic force microscopy. Cytometry A 69: 75-85. PubMed: 16419064. [DOI] [PubMed] [Google Scholar]
- 5. Air EL, Leach JL, Warnick RE, McPherson CM (2009) Comparing the risks of frameless stereotactic biopsy in eloquent and noneloquent regions of the brain: a retrospective review of 284 cases. J Neurosurg 111: 820-824. doi: 10.3171/2009.3.JNS081695. PubMed: 19408980. [DOI] [PubMed] [Google Scholar]
- 6. Chen CC, Hsu PW, Erich Wu TW, Lee ST, Chang CN et al. (2009) Stereotactic brain biopsy: Single center retrospective analysis of complications. Clin Neurol Neurosurg 111: 835-839. doi: 10.1016/j.clineuro.2009.08.013. PubMed: 19765887. [DOI] [PubMed] [Google Scholar]
- 7. Skog J, Würdinger T, van Rijn S, Meijer DH, Gainche L et al. (2008) Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol 10: 1470-1476. doi: 10.1038/ncb1800. PubMed: 19011622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Balaj L, Lessard R, Dai L, Cho YJ, Pomeroy SL et al. (2011) Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat Commun 2: 180. doi: 10.1038/ncomms1180. PubMed: 21285958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Noerholm M, Balaj L, Limperg T, Salehi A, Zhu LD et al. (2012) RNA expression patterns in serum microvesicles from patients with glioblastoma multiforme and controls. BMC Cancer 12: 22. doi: 10.1186/1471-2407-12-22. PubMed: 22251860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Hong BS, Cho JH, Kim H, Choi EJ, Rho S et al. (2009) Colorectal cancer cell-derived microvesicles are enriched in cell cycle-related mRNAs that promote proliferation of endothelial cells. BMC Genomics 10: 556. doi: 10.1186/1471-2164-10-556. PubMed: 19930720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Rabinowits G, Gerçel-Taylor C, Day JM, Taylor DD, Kloecker GH (2009) Exosomal microRNA: a diagnostic marker for lung cancer. Clin Lung Cancer 10: 42-46. doi: 10.3816/CLC.2009.n.006. PubMed: 19289371. [DOI] [PubMed] [Google Scholar]
- 12. Camussi G, Deregibus MC, Bruno S, Grange C, Fonsato V et al. (2011) Exosome/microvesicle-mediated epigenetic reprogramming of cells. Am J Cancer Res 1: 98-110. PubMed: 21969178. [PMC free article] [PubMed] [Google Scholar]
- 13. Street JM, Barran PE, Mackay CL, Weidt S, Balmforth C et al. (2012) Identification and proteomic profiling of exosomes in human cerebrospinal fluid. J Transl Med 10: 5. doi: 10.1186/1479-5876-10-S3-O5. PubMed: 22221959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Shao H, Chung J, Balaj L, Charest A, Bigner DD et al. (2012) Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat Med 18: 1835-1840. doi: 10.1038/nm.2994. PubMed: 23142818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Taylor DD, Gercel-Taylor C (2008) MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol Oncol 110: 13-21. doi: 10.1016/j.ygyno.2008.04.033. PubMed: 18589210. [DOI] [PubMed] [Google Scholar]
- 16. Thery C (2011) Exosomes: secreted vesicles and intercellular communications. F1000. Biol Reprod 3: 15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Li L, Zhu D, Huang L, Zhang J, Bian Z et al. (2012) Argonaute 2 complexes selectively protect the circulating microRNAs in cell-secreted microvesicles. PLOS ONE 7: e46957. doi: 10.1371/journal.pone.0046957. PubMed: 23077538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Raposo G, Nijman HW, Stoorvogel W, Liejendekker R, Harding CV et al. (1996) B lymphocytes secrete antigen-presenting vesicles. J Exp Med 183: 1161-1172. doi: 10.1084/jem.183.3.1161. PubMed: 8642258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Blanchard N, Lankar D, Faure F, Regnault A, Dumont C et al. (2002) TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/zeta complex. J Immunol 168: 3235-3241. PubMed: 11907077. [DOI] [PubMed] [Google Scholar]
- 20. Taylor DD, Akyol S, Gercel-Taylor C (2006) Pregnancy-Associated Exosomes and Their Modulation of T Cell Signaling. J Immunol 176: 1534-1542. PubMed: 16424182. [DOI] [PubMed] [Google Scholar]
- 21. Alberts B, Johnson A, Lewis J (2002) Molecular Biology of the Cell. New York: Garland Sciences.
- 22. D'Souza-Schorey C, Clancy JW (2012) Tumor-derived microvesicles: shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes Dev 26: 1287-1299. doi: 10.1101/gad.192351.112. PubMed: 22713869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Hunter MP, Ismail N, Zhang X, Aguda BD, Lee EJ et al. (2008) Detection of microRNA expression in human peripheral blood microvesicles. PLOS ONE 3: e3694. doi: 10.1371/journal.pone.0003694. PubMed: 19002258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-408. doi: 10.1006/meth.2001.1262. PubMed: 11846609. [DOI] [PubMed] [Google Scholar]
- 25. VanGuilder HD, Vrana KE, Freeman WM (2008) Twenty-five years of quantitative PCR for gene expression analysis. BioTechniques 44: 619-626. PubMed: 18474036. [DOI] [PubMed] [Google Scholar]
- 26. Bustin SA (2000) Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol 25: 169-193. doi: 10.1677/jme.0.0250169. PubMed: 11013345. [DOI] [PubMed] [Google Scholar]
- 27. Suzuki T, Higgins PJ, Crawford DR (2000) Control selection for RNA quantitation. BioTechniques 29: 332-337. PubMed: 10948434. [DOI] [PubMed] [Google Scholar]
- 28. Thellin O, Zorzi W, Lakaye B, De Borman B, Coumans B et al. (1999) Housekeeping genes as internal standards: use and limits. J Biotechnol 75: 291-295. doi: 10.1016/S0168-1656(99)00163-7. PubMed: 10617337. [DOI] [PubMed] [Google Scholar]
- 29. Chan JA, Krichevsky AM, Kosik KS (2005) MicroRNA-21 Is an Antiapoptotic Factor in Human Glioblastoma Cells. Cancer Res 65: 6029-6033. doi: 10.1158/0008-5472.CAN-05-0137. PubMed: 16024602. [DOI] [PubMed] [Google Scholar]
- 30. Witwer KW, Buza ´s EI, Bemis LT, Lässer C, Lötvall J, et al. (In press) Standardisation of sample collection, isolation and analysis methods in extracellular vesicle research: A position paper arising from the 2012 ISEV Workshop on RNA in extracellular vesicles. Journal of Extracellular Vesicles. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ishii N, Maier D, Merlo A, Tada M, Sawamura Y et al. (1999) Frequent co-alterations of TP53, p16/CDKN2A, p14ARF, PTEN tumor suppressor genes in human glioma cell lines. Brain Pathol 9: 469-479. doi: 10.1111/j.1750-3639.1999.tb00536.x. PubMed: 10416987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Giard DJ, Aaronson SA, Todaro GJ, Arnstein P, Kersey JH et al. (1973) In vitro cultivation of human tumors: establishment of cell lines derived from a series of solid tumors. J Natl Cancer Inst 51: 1417-1423. PubMed: 4357758. [DOI] [PubMed] [Google Scholar]
- 33. Pontén J, Saksela E (1967) Two established in vitro cell lines from human mesenchymal tumours. Int J Cancer 2: 434-447. doi: 10.1002/ijc.2910020505. PubMed: 6081590. [DOI] [PubMed] [Google Scholar]
- 34. DuBridge RB, Tang P, Hsia HC, Leong PM, Miller JH et al. (1987) Analysis of mutation in human cells by using an Epstein-Barr virus shuttle system. Mol Cell Biol 7: 379-387. PubMed: 3031469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Joshi K, Banasavadi-Siddegowda Y, Mo X, Kim SH, Mao P et al. (2013) MELK-dependent FOXM1 Phosphorylation is Essential for Proliferation of Glioma Stem Cells. Stem Cells, 31: 1051–63. PubMed: 23404835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Ng K, Nitta M, Hu L, Kesari S, Kung A et al. (2009) A small interference RNA screen revealed proteasome inhibition as strategy for glioblastoma therapy. Clin Neurosurg 56: 107-118. PubMed: 20214041. [PubMed] [Google Scholar]
- 37. Gu C, Banasavadi-Siddegowda YK, Joshi K, Nakamura Y, Kurt H et al. (2013) Tumor-specific Activation of the c-JUN/MELK Pathway Regulates Glioma Stem Cell Growth in a p53-dependent Manner. Stem Cells. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Théry C, Amigorena S, Raposo G, Clayton A (2006) Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol Chapter 3: Unit 3.22: Unit 3 22. PubMed: 18228490 [DOI] [PubMed] [Google Scholar]
- 39. Clark MJ, Homer N, O'Connor BD, Chen Z, Eskin A et al. (2010) U87MG Decoded: The Genomic Sequence of a Cytogenetically Aberrant Human Cancer Cell Line. PLOS Genet 6: e1000832 PubMed: 20126413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Trang P, Wiggins JF, Daige CL, Cho C, Omotola M et al. (2011) Systemic delivery of tumor suppressor microRNA mimics using a neutral lipid emulsion inhibits lung tumors in mice. Mol Ther 19: 1116-1122. doi: 10.1038/mt.2011.48. PubMed: 21427705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Stigler S (1989) Francis Galton's Account of the Invention of Correlation. Stat Sci 4: 7. [Google Scholar]
- 42. STUDENT (1908) THE PROBABLE ERROR OF A MEAN. Biometrika 6: 1-25. doi: 10.2307/2331554. [DOI] [Google Scholar]
- 43. Zweig MH, Campbell G (1993) Receiver-operating characteristic (ROC) plots: a fundamental evaluation tool in clinical medicine. Clin Chem 39: 561-577. PubMed: 8472349. [PubMed] [Google Scholar]
- 44. Wotschofsky Z, Meyer H-A, Jung M, Fendler A, Wagner I et al. (2011) Reference genes for the relative quantification of microRNAs in renal cell carcinomas and their metastases. Anal Biochem 417: 233-241. doi: 10.1016/j.ab.2011.06.009. PubMed: 21741950. [DOI] [PubMed] [Google Scholar]
- 45. Gonda DD, Akers JC, Kim R, Kalkanis SN, Hochberg FH et al. (2013) Neuro-oncologic Applications of Exosomes, Microvesicles, and Other Nano-Sized Extracellular Particles. Neurosurgery 72: 501-510. doi: 10.1227/NEU.0b013e3182846e63. PubMed: 23277376. [DOI] [PubMed] [Google Scholar]
- 46. Huan J, Hornick NI, Shurtleff MJ, Skinner AM, Goloviznina NA et al. (2013) RNA Trafficking by Acute Myelogenous Leukemia Exosomes. Cancer Res 73: 918-929. doi: 10.1158/0008-5472.CAN-12-2184. PubMed: 23149911. [DOI] [PubMed] [Google Scholar]
- 47. Rana S, Malinowska K, Zöller M (2013) Exosomal Tumor MicroRNA Modulates Premetastatic Organ Cells. Neoplasia 15: 281-295. PubMed: 23479506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Xiao D, Ohlendorf J, Chen Y, Taylor DD, Rai SN et al. (2012) Identifying mRNA, MicroRNA and Protein Profiles of Melanoma Exosomes. PLOS ONE 7: e46874. doi: 10.1371/journal.pone.0046874. PubMed: 23056502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. de Biase D, Visani M, Morandi L, Marucci G, Taccioli C et al. (2012) miRNAs Expression Analysis in Paired Fresh/Frozen and Dissected Formalin Fixed and Paraffin Embedded Glioblastoma Using Real-Time PCR. PLOS ONE 7: e35596. doi: 10.1371/journal.pone.0035596. PubMed: 22530056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Bangert A, Cristofanon S, Eckhardt I, Abhari BA, Kolodziej S et al. (2012) Histone deacetylase inhibitors sensitize glioblastoma cells to TRAIL-induced apoptosis by c-myc-mediated downregulation of cFLIP. Oncogene 31: 4677-4688. doi: 10.1038/onc.2011.614. PubMed: 22266862. [DOI] [PubMed] [Google Scholar]
- 51. Wang H, Shen W, Huang H, Hu L, Ramdas L et al. (2003) Insulin-like growth factor binding protein 2 enhances glioblastoma invasion by activating invasion-enhancing genes. Cancer Res 63: 4315-4321. PubMed: 12907597. [PubMed] [Google Scholar]
- 52. Jarboe JS, Johnson KR, Choi Y, Lonser RR, Park JK (2007) Expression of interleukin-13 receptor alpha2 in glioblastoma multiforme: implications for targeted therapies. Cancer Res 67: 7983-7986. doi: 10.1158/0008-5472.CAN-07-1493. PubMed: 17804706. [DOI] [PubMed] [Google Scholar]
- 53. Ehtesham M, Winston JA, Kabos P, Thompson RC (2006) CXCR4 expression mediates glioma cell invasiveness. Oncogene 25: 2801-2806. doi: 10.1038/sj.onc.1209302. PubMed: 16407848. [DOI] [PubMed] [Google Scholar]
- 54. Santangelo PJ, Alonas E, Jung J, Lifland AW, Zurla C (2012) Probes for intracellular RNA imaging in live cells. Methods Enzymol 505: 383-399. doi: 10.1016/B978-0-12-388448-0.00028-0. PubMed: 22289464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Barber RD, Harmer DW, Coleman RA, Clark BJ (2005) GAPDH as a housekeeping gene: analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol Genomics 21: 389-395. doi: 10.1152/physiolgenomics.00025.2005. PubMed: 15769908. [DOI] [PubMed] [Google Scholar]
- 56. Wada Y, Li D, Merley A, Zukauskas A, Aird WC et al. (2011) A multi-gene transcriptional profiling approach to the discovery of cell signature markers. Cytotechnology 63: 25-33. doi: 10.1007/s10616-010-9315-8. PubMed: 20972619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Visani M, de Biase D, Marucci G, Taccioli C, Baruzzi A et al. (2013) Definition of miRNAs expression profile in glioblastoma samples: the relevance of non-neoplastic brain reference. PLOS ONE 8: e55314. doi: 10.1371/journal.pone.0055314. PubMed: 23383149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Filipe V, Hawe A, Jiskoot W (2010) Critical evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the measurement of nanoparticles and protein aggregates. Pharm Res 27: 796-810. doi: 10.1007/s11095-010-0073-2. PubMed: 20204471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Wright M (2012) Nanoparticle tracking analysis for the multiparameter characterization and counting of nanoparticle suspensions. Methods Mol Biol 906: 511-524. PubMed: 22791460. [DOI] [PubMed] [Google Scholar]
- 60. Gercel-Taylor C, Atay S, Tullis RH, Kesimer M, Taylor DD (2012) Nanoparticle analysis of circulating cell-derived vesicles in ovarian cancer patients. Anal Biochem 428: 44-53. doi: 10.1016/j.ab.2012.06.004. PubMed: 22691960. [DOI] [PubMed] [Google Scholar]
- 61. Vallhov H, Gutzeit C, Johansson SM, Nagy N, Paul M et al. (2011) Exosomes containing glycoprotein 350 released by EBV-transformed B cells selectively target B cells through CD21 and block EBV infection in vitro. J Immunol 186: 73-82. doi: 10.4049/jimmunol.1001145. PubMed: 21106852. [DOI] [PubMed] [Google Scholar]
- 62. Galli R, Binda E, Orfanelli U, Cipelletti B, Gritti A et al. (2004) Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res 64: 7011-7021. doi: 10.1158/0008-5472.CAN-04-1364. PubMed: 15466194. [DOI] [PubMed] [Google Scholar]
- 63. Lee J, Kotliarova S, Kotliarov Y, Li A, Su Q et al. (2006) Tumor stem cells derived from glioblastomas cultured in bFGF and EGF more closely mirror the phenotype and genotype of primary tumors than do serum-cultured cell lines. Cancer Cell 9: 391-403. doi: 10.1016/j.ccr.2006.03.030. PubMed: 16697959. [DOI] [PubMed] [Google Scholar]
- 64. Caby MP, Lankar D, Vincendeau-Scherrer C, Raposo G, Bonnerot C (2005) Exosomal-like vesicles are present in human blood plasma. Int Immunol 17: 879-887. doi: 10.1093/intimm/dxh267. PubMed: 15908444. [DOI] [PubMed] [Google Scholar]
- 65. Chang A (2013) StatsToDo website. Available: www.statstodo.com. Accessed 2013 March 4.
- 66. Raposo Ga Stoorvogel W (2013) Extracellular vesicles: Exosomes, microvesicles, and friends. J Cell Biol 200: 373-383. doi: 10.1083/jcb.201211138. PubMed: 23420871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Akers JC, Gonda D, Kim R, Carter BS, Chen CC (2013) Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neuro Oncol 113: 1-11. doi: 10.1007/s11060-013-1084-8. PubMed: 23456661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Arroyo JD, Chevillet JR, Kroh EM, Ruf IK, Pritchard CC et al. (2011) Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc Natl Acad Sci U S A 108: 5003-5008. doi: 10.1073/pnas.1019055108. PubMed: 21383194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Boon RA, Vickers KC (2013) Intercellular transport of microRNAs. Arterioscler Thromb Vasc Biol 33: 186-192. doi: 10.1161/ATVBAHA.112.300139. PubMed: 23325475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Teplyuk NM, Mollenhauer B, Gabriely G, Giese A, Kim E et al. (2012) MicroRNAs in cerebrospinal fluid identify glioblastoma and metastatic brain cancers and reflect disease activity. Neuro-Oncology 14: 689-700. doi: 10.1093/neuonc/nos074. PubMed: 22492962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Guduric-Fuchs J, O'Connor A, Cullen A, Harwood L, Medina RJ et al. (2012) Deep sequencing reveals predominant expression of miR-21 amongst the small non-coding RNAs in retinal microvascular endothelial cells. J Cell Biochem 113: 2098-2111. doi: 10.1002/jcb.24084. PubMed: 22298343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Qi L, Bart J, Tan LP, Platteel I, Sluis Tv et al. (2009) Expression of miR-21 and its targets (PTEN, PDCD4, TM1) in flat epithelial atypia of the breast in relation to ductal carcinoma in situ and invasive carcinoma. BMC Cancer 9: 163. doi: 10.1186/1471-2407-9-163. PubMed: 19473551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Deftereos G, Corrie SR, Feng Q, Morihara J, Stern J et al. (2011) Expression of Mir-21 and Mir-143 in Cervical Specimens Ranging from Histologically Normal through to Invasive Cervical Cancer. PLOS ONE 6: e28423. doi: 10.1371/journal.pone.0028423. PubMed: 22194833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Bhagat TD, Zhou L, Sokol L, Kessel R, Caceres G et al. (2013) miR-21 mediates hematopoietic suppression in MDS by activating TGF-beta signaling. Blood 121: 2875-2881. doi: 10.1182/blood-2011-12-397067. PubMed: 23390194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Wang Q, Li P, Li A, Jiang W, Wang H et al. (2012) Plasma specific miRNAs as predictive biomarkers for diagnosis and prognosis of glioma. J Exp Clin Cancer Res 31: 97. doi: 10.1186/1756-9966-31-97. PubMed: 23174013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Huggett J, Dheda K, Bustin S, Zumla A (2005) Real-time RT-PCR normalisation; strategies and considerations. Genes Immun 6: 279-284. doi: 10.1038/sj.gene.6364190. PubMed: 15815687. [DOI] [PubMed] [Google Scholar]
- 77. Tran Thang NN, Derouazi M, Philippin G, Arcidiaco S, Di Berardino-Besson W et al. (2010) Immune infiltration of spontaneous mouse astrocytomas is dominated by immunosuppressive cells from early stages of tumor development. Cancer Res 70: 4829-4839. doi: 10.1158/0008-5472.CAN-09-3074. PubMed: 20501837. [DOI] [PubMed] [Google Scholar]
- 78. Ghosh A, Chaudhuri S (2010) Microglial action in glioma: a boon turns bane. Immunol Lett 131: 3-9. doi: 10.1016/j.imlet.2010.03.003. PubMed: 20338195. [DOI] [PubMed] [Google Scholar]
- 79. Verhaak RG, Hoadley KA, Purdom E, Wang V, Qi Y et al. (2010) Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 17: 98-110. doi: 10.1016/j.ccr.2009.12.020. PubMed: 20129251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Møller HG, Rasmussen AP, Andersen HH, Johnsen KB, Henriksen M et al. (2013) A systematic review of microRNA in glioblastoma multiforme: micro-modulators in the mesenchymal mode of migration and invasion. Mol Neurobiol 47: 131-144. doi: 10.1007/s12035-012-8349-7. PubMed: 23054677. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Electron micrograph of EVs isolated from CSF. EVs were isolated from CSF by differential centrifugation and analyzed by transmission electron microscopy. Scale bar represents 200nm in (A) and 100nm in (B). EVs in the size range of 50-250nm were observed.
(EPS)
Relative abundance of GAPDH, 18S RNA, and miR-103 in EV derived from sera of glioblastoma and non-oncologic patients. GAPDH, 18S rRNA, and hsa-mir-103 transcript copy number was plotted against the total number of EVs isolated for RNA extraction. The relative abundance of these transcripts in EV ranged between 1 transcript in 7 EVs to 1 transcript in 106 EVs.
(EPS)
Correlation between EV number and RNA yield. For each sample, the RNA yield from EV preparations for (A) neurosphere cell lines, (B) Serum samples, and (C) CSF samples were plotted on the X-axis; the input EV number for these two datasets were plotted on the Y axis. There was excellent correlation between the RNA yield and the EV preparation in neurosphere lines, the correlation was poor in serum samples.
(EPS)
Abundance of EVs per cc of biofluids. (A) Abundance of EVs per cc of serum in glioblastoma and non-oncogenic patients. There was no significant difference between tumor and control samples. (B) Abundance of EVs per cc of CSF in glioblastoma and non-oncogenic patients. No significant differences were observed.
(EPS)
Average size profile of EVs isolated from biofluids. (A) Average size distribution of EVs isolated from serum of glioblastoma and non-oncogenic patients. There was no significant difference between tumor and control samples. (B) Average size distribution of EVs isolated from the CSF of glioblastoma and non-oncogenic patients. No significant size differences were observed.
(EPS)
Effects of culturing condition on miR-21 expression in cells and EVs. Glioblastoma cell lines U87MG and CMK3 were grown under adherent or neurosphere condition and the miR-21 level in cells or EVs were quantitated.
(EPS)
Normalization of EV miR-21 by RNA yield. (A) The expression level of miR-21 was quantitatively assessed in nine neurosphere glioblastoma lines using total RNA yield as normalization parameter. (B) Comparison of normalization method revealed good correlation between normalizing by EV particle number or total RNA yield in cell culure. (C) Levels of miR-21 in EVs isolated from glioblastoma and non-oncologic patients normalized by RNA yield. (D) Correlation between normalization by EV number or total RNA yield in CSF.
(EPS)
Relative abundance of miR-24 and miR-125 in EVs derived from neurosphere glioblastoma cell lines and CSF of glioblastoma and non-oncologic patients. has-miR-24 and has-miR-125 levels in EV isolated from from (A) 9 glioblastoma neurophere lines, cultured under serum-free conditions or (B) from CSF of 13 glioblastoma and 14 non-oncologic patients were plotted against total RNA extracted from the EVs or against the total number of EVs isolated for RNA extraction.
(EPS)
List of primers used.
(DOCX)