

Abstract
Artificial APCs (aAPCs) genetically modified to express selective costimulatory molecules provide a reproducible, cost-effective, and convenient method for polyclonal and Ag-specific expansion of human T cells for adoptive immunotherapy. Among the variety of aAPCs that have been studied, acellular beads expressing anti-CD3/anti-CD28 efficiently expand CD4+ cells, but not CD8+ T cells. Cell-based aAPCs can effectively expand cytolytic CD8+ cells, but optimal costimulatory signals have not been defined. 4-1BB, a costimulatory molecule expressed by a minority of resting CD8+ T cells, is transiently up-regulated by all CD8+ T cells following activation. We compared expansion of human cytolytic CD8+ T cells using cell-based aAPCs providing costimulation via 4-1BB vs CD28. Whereas anti-CD3/anti-CD28 aAPCs mostly expand naive cells, anti-CD3/4-1BBL aAPCs preferentially expand memory cells, resulting in superior enrichment of Ag-reactive T cells which recognize previously primed Ags and efficient expansion of electronically sorted CD8+ populations reactive toward viral or self-Ags. Using HLA-A2-Fc fusion proteins linked to 4-1BBL aAPCs, 3-log expansion of Ag-specific CD8+ CTL was induced over 14 days, whereas similar Ag-specific CD8+ T cell expansion did not occur using HLA-A2-Fc/anti-CD28 aAPCs. Furthermore, when compared with cytolytic T cells expanded using CD28 costimulation, CTL expanded using 4-1BB costimulation mediate enhanced cytolytic capacity due, in part, to NKG2D up-regulation. These results demonstrate that 4-1BB costimulation is essential for expanding memory CD8+ T cells ex vivo and is superior to CD28 costimulation for generating Ag-specific products for adoptive cell therapy.
Clinical trials have demonstrated beneficial effects of adoptive immunotherapy in viral infection (1, 2), HIV infection (3), EBV-associated lymphoproliferative disorder (4), EBV-associated nasopharyngeal carcinoma (5), Hodgkin's disease (6), and malignant melanoma (7, 8). In murine melanoma, combining adoptive T cell therapy with vaccination and cytokine therapy can induce durable regression of sizable tumors whereas similar results cannot be achieved with vaccination and/or cytokine administration alone (9). Thus, adoptive immunotherapy holds promise for the treatment of viral and neoplastic disease. Currently, most investigators generate cell products for clinical trials of adoptive immunotherapy by repetitive TCR-based stimulation of PBLs using one of a variety of APCs. No consensus exists regarding the optimal cell type or costimulatory signals needed to generate these products, therefore a variety of APCs have been used including EBV-immortalized lymphoblast cell lines (LBL)3 (2, 10), CD40-activated B cells (11), autologous monocytes (12), autologous PBMCs (13), and autologous dendritic cells (14, 15). Although each of these approaches has been demonstrated capable of expanding Ag-specific T cells, careful studies to identify the optimal costimulatory signals for generating CTL products have not been undertaken. Thus, it remains unclear which, if any, APC is superior to the others.
The CD28:B7 pathway is widely considered the most important costimulatory pathway for TCR activation (16) and, as a result, most approaches used to expand T cells have emphasized a critical role for CD28 costimulation. However, bead-based anti-CD3/CD28 artificial APCs (aAPCs) induce brisk expansion of CD4+ populations but not CD8+ T cells (17–19), demonstrating that the CD28 costimulation is not sufficient for expansion of CD8+ CTL. Using genetically modified autologous monocytes, Bukczynski et al. (12) identified a critical role for 4-1BBL in expanding Ag-specific CTL, and Maus et al. (19) demonstrated that the incorporation of 4-1BBL into an aAPC greatly augmented the capacity for Ag-specific expansion of CD8+ T cells ex vivo. Thus, these results provided evidence that 4-1BB signaling augmented ex vivo expansion of CD8+ CTL. However, the APCs used in both of these studies also provided CD28 signaling, and therefore the studies did not clarify whether 4-1BB costimulation was sufficient to induce expansion in the absence of CD28 signaling.
In this study, we used genetically modified aAPCs to directly compare CD28 vs 4-1BB costimulation for expanding cytolytic CD8+ cells. Surprisingly, we observed efficient expansion of CD8+ CTL using 4-1BB costimulation in the absence of CD28 signaling, thus demonstrating that CD28 costimulation is not necessary for efficient expansion of memory CD8+ CTL. Furthermore, we demonstrate that 4-1BB costimulation preferentially expands memory CD8+ T cells, and that 4-1BB signaling augments cytolytic function both directly, and indirectly, via up-regulation of NKG2D. Thus, 4-1BB is an essential costimulatory molecule for efficient ex vivo expansion of Ag-specific CD8+ CTL.
Materials and Methods
Donors and cells
HLA-A0201+ PBLs were obtained from healthy donors by leukapheresis following informed consent according to National Institutes of Health Clinical Center and/or the University of Pennsylvania Institutional Review Board approved protocols. CD8 + T cells were isolated by negative selection according to the manufacturer's instructions via a MACS CD8 T cell isolation kit (Miltenyi Biotec). Resulting populations consisted of >90% CD8+ T cells. P815 and T2 (American Type Culture Collection (ATCC)) cell lines were cultured and maintained in RPMI 1640 containing 10% FCS (Harlan), 2 mM L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin, and 50 μg/ml 2-ME at 37°C, 5% CO2.
aAPCs and cell expansion cultures
K32 (20) and K32/4-1BBL (19) aAPCs were generated and maintained as described previously. Briefly, the parent K562 cell line (ATCC) was transfected with CD32 to generate the K32 aAPC, which is maintained in culture under G418 selection. The K32/4-1BBL aAPC was generated by transfecting the K32 line with 4-1BBL, and this line is maintained in culture under G418 and hygromycin selection. The procedures used to generate the KT series of cell lines have been detailed elsewhere (21). Briefly, K562 cells (ATCC) were transduced with lentiviral vectors to introduce genes for CD32, CD80, 4-1BBL, IL-15, and sorted into single-cell clones using high-speed MoFlo sorting (Cytomation) to generate KT-4-1BBL that express CD32 and 4-1BBL, KT80-4-1BBL that expresses CD32, CD80, and 4-1BBL, KTIL-15-4-1BBL-CD80 that secretes IL-15 and expresses CD32, 4-1BBL, and CD80, and KT80 that expresses CD32 and CD80. All aAPCs were irradiated (10,000 rad) and cryopreserved in 10% DMSO and 90% FCS then thawed before initiation of aAPC:T cell coculture.
Upon thawing, the aAPCs were suspended at a concentration of 107 cells/ml and coincubated with the designated mAbs: anti-CD3 (OKT3; Ortho Biotech), and/or anti-CD28 (clone 9.3; kindly provided by Dr. R. Gress, Experimental Transplantation and Immunology Branch, National Cancer Institute, Bethesda, MD), at 1 μg/ml for 15 min at room temperature, then washed and resuspended before coincubation with the CD8+ cells undergoing expansion. For experiments using HLA-A2-Fc fusion proteins, thawed K32 and K32/4-1BBL were maintained at cell concentrations of <107/ml and incubated with the HLA-A2-Ig-Fc fusion protein (BD Biosciences) at 1 μg/ml for 5 min at room temperature. The designated HLA-A2-binding peptide was then added at a concentration of 10 μM for an additional 10 min. The peptide-loaded HLA-A2-Ig-aAPCs were then directly placed into coculture without washing. CD8+ T cells were incubated with the cell based aAPCs at a 2:1 CD8:aAPC ratio or with anti-CD3/anti-CD28 beads (Dynal Biotech) at a 1:3 CD8:bead ratio. All cell expansion cultures used 10% human AB serum in AIM-V (Invitrogen Life Technologies) medium containing 20 IU/ml IL-2 (Chiron) and 10 ng/ml IL-7 (Cytheris).
Flow cytometry and FACS sorting
T cells (∼106/tube) were removed from culture, washed with FACS medium (PBS containing 0.5% FCS and 2 mM EDTA), collected by centrifugation, and resuspended in 100 μl of FACS medium. Cells were stained with the Abs indicated (fluorochrome-labeled Abs were purchased from Caltag Laboratories, unless specified otherwise) for phenotypic analysis on ice for 30 min. Anti-4-1BB-FITC, anti-IL-15-FITC, anti-IL-15Rα, CD107a-PE, and NKG2D-PE were purchased from BD Pharmingen. For tetramer-binding assays, cells were stained with the designated HLA-A2+ tetramers (Beckman Coulter) for 30 min at room temperature. Apoptosis was assayed using Annexin VFITC (BD Pharmingen) surface staining according to the manufacturer's manual. Flow cytometry was performed using a FACSCalibur and CellQuest software (BD Biosciences) and analyzed using FlowJo 8.3 (Tree Star). FACS sorting for CMV+ and Flu+ cells was performed using a FACSVantage (BD Immunocytometry Systems).
Naive and memory CD8+ T cell proliferative assay
CD8 T cells, isolated as described above, were separated into naive (CD45RO−) and memory (CD45RO+) subsets using CD45RO microbeads, according to the manufacturer's instructions (Miltenyi Biotec). The resulting population consisted of >90% CD45RO− (collected negatively) or CD45RO+ (collected positively) T cells. Subsequently, naive and memory CD8 T cells (105 each per well) were added to a 96-well flat-bottom plate (Nunc), which was previously coated with OKT3 at 1 μg/ml and variable doses of anti-4-1BB (R&D Systems) and anti-CD28 as indicated. Cells were incubated for 5 days at 37°C in a final volume of 200 μl/well with 10% human AB serum and AIM-V medium. Proliferation was measured by adding 1 μCi of [3H]thymidine/well (ICN Pharmaceuticals) 16 –18 h before the cells were harvested and thymidine incorporation (expressed as cpm) was measured using a gamma counter (Beckman Packwood).
Cytolytic assays
Where designated, cultures were harvested, cells were counted, and lytic activities were measured using conventional 4-h 51Cr-release assays using T2 target cells that were first pulsed with either 10 μM Flu M1 peptide (GILGFVFTL), CMV pp65 peptide (NLVPMVATV), or the carcinoem-bryonic Ag-derived HLA-A2-binding peptide CAP1 (GVLVGVALI; New England Peptide), then labeled with 51Cr (PerkinElmer). Radioactivity in the supernatant was measured and the percentage-specific 51Cr lysis was calculated using the following equation: percent-specific lysis = (test release − spontaneous release)/(maximal release − spontaneous release) × 100. Cytolysis was also measured using CD107a expression as a marker of degranulation as previously described (22).
For the T cell-redirected cytolytic assay, P815 cells (1 × 106) were labeled with 51Cr as described above, then coupled with anti-CD3 (Ortho-clone), anti-4-1BB (R&D Systems), anti-CD28 (BD Biosciences), and/or anti-NKG2D (R&D Systems) where designated for 15 min at room temperature. Expanded T cells were assayed for their ability to lyse 51Cr-labeled Ab-bound P815 target cells in an Ag-independent manner using a conventional 4-h 51Cr-release assay as described above. One lytic unit was defined as 30% lysis of 5 × 103 target cells. Blockade of lysis following coincubation with the anti-MHC class I mAb W6/32 (Leinco Tech) was used to confirm MHC-restricted killing.
Statistical analysis
Differences between values were evaluated using a two-tailed unpaired Student's t test. Significance was determined as p < 0.05.
Results
TCR stimulation induces transient 4-1BB expression on all circulating CD8+ T cells
4-1BB is not expressed by resting naive or memory peripheral blood CD8+ T cells, but is up-regulated following activation (23). To assess 4-1BB expression on normal human CD8+ PBL following anti-CD3/anti-CD28 vs anti-CD3/anti-4-1BB-based stimulation, CD8+ T cells obtained from healthy donors were stimulated with plate-bound anti-CD3/anti-CD28 or anti-CD3/anti-4-1BB. In both cultures, 4-1BB was transiently up-regulated on essentially all CD8+ cells from normal donors with peak expression between 8 and 10 h following stimulation (Fig. 1A). A similar time course and degree of 4-1BB expression was also observed following anti-CD3 stimulation alone (data not shown). Therefore, although 4-1BB is expressed by only a minority of resting peripheral blood CD8+ T cells, T cell activation induces rapid, transient up-regulation of 4-1BB on all circulating CD8+ T cells, raising the possibility that 4-1BBL could serve as an effective costimulatory molecule for ex vivo expansion of CD8+ cells.
Figure 1.

K32 cell-based aAPCs are superior to bead-based aAPCs for CD8+ expansion. A, CD8 PBL were purified and stimulated using plate-bound anti-CD3 plus anti-4-1BB (top row) or anti-CD3 plus anti-CD28 (bottom row) for the duration indicated. CD3+ CD8+ cells were gated by flow cytometry and evaluated for 4-1BB expression. Blue is binding of anti-4-1BB mAb at time 0, red is binding of anti-4-1BB mAb at time points indicated, and green is binding of anti-4-1BB mAb following preincubation with 4-1BBL-μ Ig Fc fusion protein at the time points indicated. This is a representative of three separate experiments using cells from different normal donors. B, Expansion of human CD8 T cells stimulated on with K32/anti-CD3/4-1BBL, K32/anti-CD3/anti-CD28, or anti-CD3/anti-CD28-coated magnetic beads as described in Materials and Methods. Fold cell number increase after 7 days is shown. Results are representative of more than seven different experiments, each using cells from a different normal donor. C, CD8+ cells on day 7 following expansion using anti-CD3/anti-CD28 beads demonstrate higher apoptosis compared with cells expanded using K32/anti-CD3/anti-CD28 or K32/anti-CD3/4-1BBL-based aAPCs, as evidenced by flow cytometric binding of FITC-labeled annexin V. Results are a representative of five experiments using cells from different normal donors. D, Both K32 and K32/4-1BBL aAPCs produce IL-15 and express IL-15Rα. Intracellular IL-15 and surface IL-15Rα analysis was performed on K32 and K32/4-1BBL aAPCs, as well as EBV LBL serving as a negative control for IL-15 and 5838 tumor cells serving as a negative control for IL-15Rα. E, Anti-IL-15 partially blocks CD8+ cell proliferation induced by K32/anti-CD3/4-1BBL. CD8+ T cells were coincubated with aAPC using anti-CD/4-1BBL at various ratios in the presence of anti-IL-15 mAb and isotype Ab as a control. The doses of Abs used were indicated in the figure. Proliferation was measured by [3H]thymidine incorporation during the last 16 ∼ 18 h of a 5-day culture. Results were a representative of three different experiments, using cells from different individual donors.
Cell-based aAPCs induce greater CD8 expansion than bead-based aAPCs
To determine the relative efficiency of 4-1BBL and CD28 costimulation for expanding human CD8+ T cells, enriched peripheral blood CD8+ T cells were enumerated following two consecutive stimulations (days 0 and 7) with aAPCs comprising K32 cell-based anti-CD3/4-1BBL, K32 cell-based anti-CD3/anti-CD28, and anti-CD3/anti-CD28-coated magnetic beads (Fig. 1B). Anti-CD3/4-1BBL and anti-CD3/anti-CD28 stimulation delivered via the K32 aAPCs induced rapid growth with a mean 7-fold expansion at 7 days, whereas anti-CD3/anti-CD28-coated beads induced only modest CD8+ T cell expansion during the same period. Similar relationships were observed on day 14 with K32-based expansion cultures generating a median of 50-fold expansion compared with ∼10-fold expansion using bead-based aAPCs (data not shown). These results confirm previous data demonstrating that bead-based aAPCs do not efficiently expand CD8+ T cells (17–19). To determine whether the differences in CD8+ T cell expansion induced by cell- vs bead-based aAPCs resulted from different apoptotic rates, we measured the percentage of cells expressing annexin V throughout the first 7 days of culture under the various conditions. Bead-based anti-CD3/anti-CD28-stimulated CD8+ T cells demonstrated more apoptosis compared with cells expanded using K32-based aAPCs (Fig. 1C), indicating that programmed cell death limits expansion of CD8+ T cells when using anti-CD3/anti-CD28-coated beads. Because cell-based aAPCs outperform the bead-based aAPCs for CD8 expansion, we investigated whether the K32 cells might express IL-15 and/or IL-15Rα, which could exert an antiapoptotic effect on expanding CD8+ T cells (24). Indeed, both K32 and K32/4-1BBL aAPCs express IL-15Rα and produce IL-15 (Fig. 1D), a key survival factor for activated and memory CD8+ T cells. Furthermore, neutralization of IL-15 in the cell-based aAPC:CD8+ cocultures substantially diminished CD8+ T cell proliferation, providing evidence that the augmented effectiveness of the cell-based aAPCs in this system results, at least in part, from the production and presentation of IL-15 by the K32 cells (Fig. 1E).
aAPCs providing both CD3 and 4-1BB signaling selectively enrich for CD28− memory CTLs
Adoptive immunotherapy typically seeks to eradicate large antigenic burdens in viral reservoirs or tumor cells in hosts already primed toward the Ag to be targeted. Therefore, one goal of adoptive immunotherapy is to increase the number of primed, Ag-specific CTL beyond that generated naturally in vivo. To compare the capacity for 4-1BBL vs anti-CD28-based costimulation to expand memory CD8+ CTL, we compared the frequency of flu- and CMV-specific CTL in HLA-A0201+ peripheral blood CD8+ T cells following expansion using anti-CD3/anti-CD28 vs anti-CD3/4-1BBL K32 aAPCs. As shown in Fig. 2 A, baseline flu matrix protein (FluM1)- and CMV (pp65)-specific cell frequencies were 0.02 and 0.04%, respectively; following anti-CD3/4-1BBL-based expansion, flu- and CMV-specific cells increased in frequency to 0.4 and 0.6%, respectively, whereas anti-CD3/anti-CD28 expansion did not enrich viral-specific cells in the expanded populations (Fig. 2A). An HLA-A2-binding peptide derived from ERBB2 (Her2Neu), a self Ag which is not expected to induce immune priming in healthy donors, served as a control because the low frequency of ERBB2-specific cells did not change following either anti-CD3/anti-CD28 or anti-CD3/4-1BBL-based expansion. The enrichment observed using tetramer enumeration is functionally significant because anti-CD3/4-1BBL expanded CD8+ PBL demonstrated lytic activity against the FluM1- and pp65-pulsed targets whereas the anti-CD3/anti-CD28-expanded CD8+ PBL did not (Fig. 2B). Therefore, anti-CD3/4-1BBL expansion rapidly enriches peripheral blood CD8+ populations for memory CTL, which retain functional activity.
Figure 2.
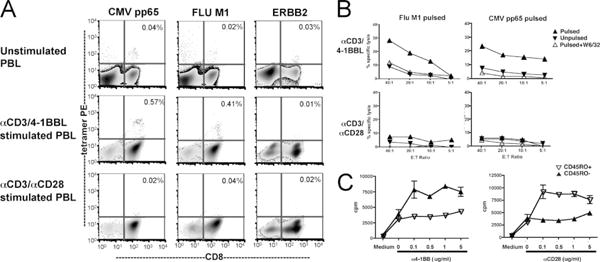
PBL costimulated with 4-1BBL, but not CD28, are enriched for memory CTL. A, CD8 PBL were unstimulated (top panel) or stimulated with K32/anti-CD3/4-1BBL (middle panel) or K32/anti-CD3/anti-CD28 (lower panel) for 7 days, then expanded cells were assayed using HLA A*0201 tetramers loaded with CMV pp65, Flu M1 peptide, and ERBB2 peptides. Anti-CD3/4-1BBL-expanded cells, but not anti-CD3/anti-CD28-expanded cells show increased frequencies of cells responding to viral recall Ags. Percentages reflect the percent of tetramer binding of CD8+ cells. B, CD8 PBL stimulated with K32/anti-CD3/4-1BBL, but not K32/anti-CD3/anti-CD28, demonstrate cytotoxicity to FluM1- and pp65-pulsed T2 targets on day 7. Results were representative of five different experiments, using cells from different individual donors. C, 4-1BB costimulation preferentially expands memory CD8+ cells but anti-CD28 costimulation preferentially expands naive CD8 cells. CD8 T cells were separated into naive (CD45RO−) and memory (CD45RO+) subsets using anti-CD45RO microbeads, then placed into a 96-well flat-bottom plate coated with anti-CD3 (1 μg/ml) and anti-CD28 or anti-4-1BB at the concentrations indicated. Proliferation was measured by [3H]thymidine incorporation during the last 16∼18 h of a 5-day culture. This is a representative result of three different experiments.
These results suggested that 4-1BBL costimulation preferentially expands memory CTL whereas anti-CD28 costimulation preferentially expands naive CD8+ T cells. To directly address this, CD8 PBL were immunomagnetically separated into CD45RO+ and RO− subsets (purity >90%) and then cultured on plates coated with a submitogenic dose of OKT3 (1 μg/ml) and titrated doses of anti-4-1BB or anti-CD28. Whereas the combination of anti-CD3/anti-CD28 preferentially expands naive CD8 subsets, anti-CD3/anti-4-1BB costimulation preferentially expanded memory CD8 subsets (Fig. 2C). These results confirm the 4-1BBL preferentially expands memory CTL and further demonstrate that the differential results shown in Fig. 2A are not due to a previously unrecognized difference between the K32 and the K32/4-1BBL cell lines, because this study used plate-bound Abs instead of K32-based cellular aAPCs. Rather, the results are consistent with the known physiologic role for 4-1BB costimulation in sustaining expansion of Ag-specific memory CD8+ T cells in vivo during the evolution of an immune response, as demonstrated previously in other model systems (24–27).
These results demonstrate that 4-1BBL costimulation can modestly enrich viral-specific CTL following polyclonal expansion, however, many clinical applications seek to use a product with much higher levels of antigenic specificity than achieved using the polyclonal expansion approach shown above. We therefore investigated whether the K32 aAPCs delivering signals via anti-CD3 and 4-1BBL could effectively expand highly enriched Ag-reactive enriched CD8+ T cells from T cell populations which were electronically sorted based upon tetramer binding (Fig. 3A). Peripheral blood CD8+ cells from donors immune to influenza were first expanded for 10 days using anti-CD3/4-1BBL aAPCs to increase the frequency of flu-specific cells above that found at baseline. CD8 cells binding FluM1 tetramers were then electronically sorted and placed back into coculture with anti-CD3/4-1BBL aAPCs. Fourteen days following stimulation of the sorted populations, flu-specific CD8 cells comprised >85% of the culture and the cells had undergone 1 log expansion. The cells demonstrated robust IFN-γ production and degranulation following exposure to FluM1 peptide-pulsed targets but not targets pulsed with irrelevant peptide. In three similar experiments, a mean of 1 log expansion was observed in 14 days using electronically sorted populations. Therefore, anti-CD3/4-1BBL aAPCs can be used to generate sizable populations of highly enriched Ag-specific CTL from peripheral blood, which retain specificity and function. Furthermore, not only does this data demonstrate that CD28 costimulation is not necessary to expand memory CTL, but also CD28 costimulation was inferior to 4-1BBL in expanding these populations because K32 aAPCs delivering anti-CD3/anti-CD28 expanded the electronically sorted cultures but they did not retain Ag specificity or function (data not shown). These results suggest that either the small numbers of naive cells contaminating the sorted cell population were preferentially expanded via CD28 costimulation, resulting in a subsequent loss of Ag specificity and/or CD28 costimulation induced anergy in these populations as described previously (18).
Figure 3.

Generation of FluM1 and MART-1-specific CTL using K32/4-1BBL aAPCs. A, CD8 cells from a normal HLA-A2+ donor underwent polyclonal expansion using anti-CD3/4-1BBL resulting in enrichment of FluM1-specific cells to 0.9% by day 10. Cells were then stained with FluM1 tetramers, electronically sorted, collected, and re-expanded using anti-CD3/4-1BBL aAPCs. Ag-specific cell numbers increased from 1.4 × 104 cells on day 10 presort to 2.4 × 105 cells on day 24. The expanded cells demonstrated specificity and functional capacity as evidenced by CD107a expression (bold) and intracellular IFN-γ production (bold) in response to FluM1- but not irrelevant peptide (CAP1)-pulsed T2 targets. Isotype controls are shown in gray. This is representative of more than nine experiments sorted using both CMV- and flu-specific cells. B, Left, Expansion of Mart-1-enriched CTLs using KT32/anti-CD3 aAPCs expressing 4-1BBL, 4-1BBL + CD80, or CD80. Mart-1-specific CD8+ T cell frequency, assessed using tetramer-based flow cytometry, was 10.5% on day 0, and 13.7 and 12.8% on day 9 in the KT32/anti-CD3/4-1BBL and KT32/anti-CD3/4-1BBL/CD80 aAPC-stimulated cultures, respectively. KT32/anti-CD3/CD80 aAPCs did not expand cells after day 4, and no viable CTLs were recovered on day 8, which prevented further phenotype/functional analysis for this condition. Middle, Mart-1 CTL were tested for cytotoxicity using a 4-h chromium release assay on day 9 post-stimulation with either KT32/anti-CD3/4-1BBL (square) or KT32/anti-CD3/4-1BBL/CD80 (diamond) aAPCs. Cr-labeled T2 targets were pulsed with Mart-1 (solid) or control irrelevant peptide (open). There were insufficient expanded CTLs to study cells cocultured with KT32 aCD3/CD80 aAPCs. Right, Mart-1-enriched CTLs were repetitively stimulated (days 0, 9, and 14) using KT32/anti-CD3/IL-15/4-1BBL/CD80 aAPCs for long-term expansion. Mart-1-specific cells ranged from 10 to 20% during culture (data not shown).
aAPCs providing both CD3 and 4-1BB signaling support expansion of cytolytic MART-1-reactive effector CTL
Because T cells used in tumor-directed adoptive cell therapy often seek to target tumor-associated Ags which represent self Ags (28), we also sought to determine the relative efficiency of expanding self-reactive memory cells using this approach. For these experiments, the KT32 cells lines were used as these represent the product currently being developed for clinical application. A MART-1 reactive CTL line was generated from an HLA-A201+ donor by three cycles of stimulation with peptide-loaded autologous APC. After three cycles of stimulation, the culture comprised 10% MART-1-specific CD8+ T cells, as measured by tetramer flow cytometry. The cell line was then restimulated with the KT32 aAPC expressing anti-CD3 and 4-1BBL and/or CD80 as a source of costimulation (Fig. 3B). Whereas MART-1 cells expanded and maintained viability following stimulation with anti-CD3/4-1BBL and anti-CD3/4-1BBL/CD80 aAPCs, no viable MART-1-reactive CD8 T cells were recovered in the cells stimulated aCD3/CD80 aAPCs. Furthermore, the MART-1 cells recovered from the 4-1BBL-containing culture exhibited potent peptide-specific cytotoxicity (Fig. 3B, middle panel). In addition, three successive cycles of stimulation of the MART-1 cell line with an cell-based aAPC genetically manipulated to express IL-15, CD80, 4-1BBL, and CD32 (used to load anti-CD3) induced robust expansion of the MART-1-reactive CTL, to potentially therapeutic numbers (Fig. 3B, right panel).
Selective expansion of Ag-specific cells from polyclonal populations using MHC-Fc fusion peptides and K32/4-1BBL aAPCs
Because electronic cell sorting is expensive, labor intensive, and requires large numbers of starting cells, we sought to determine whether the preferential expansion of memory cells induced by 4-1BBL costimulation could lead to selectively enriched Ag-specific memory CTL from polyclonal T cell populations without the requirement for electronic cell sorting. To that end, the CD32-expressing aAPCs described above were coupled to an HLA-A2 Ig-Fc fusion protein, which was subsequently pulsed with the CMV pp65 antigenic peptide to expand memory CD8 cells reactive toward CMV. Ag-specific T cell expansion was compared when the costimulatory signal was provided by anti-CD28 vs 4-1BBL. Fig. 4 shows the results of cell cultures stimulated in this manner weekly for 3 wk. On day 22 of culture, 4-1BBL expressing aAPC induced substantial enrichment of CMV-specific T cells (7% CMV positive) (Fig. 4A) which retained functionality and specificity, (Fig. 4B), whereas the K32 aAPC, bearing the same pp65 A2-Ig Fc but delivering anti-CD28 costimulation, did not expand CMV-specific T cells. Cytolytic activity quantitated based upon degranulation and measured using CD107a expression (22) suggest that the tetramer assay in Fig. 4A underestimated the true frequency of pp65-specific CTL. In a series of three consecutive experiments, 10- to 100-fold enrichment and >2 log expansion of viral Ag-specific CD8+ T cells was observed within 14 days using cells from three different normal donors stimulated with HLA-A2-Fc/4-1BBL aAPCs (Table I). Similar Ag-specific expansion was also accomplished using FluM1-pulsed HLA-A2-Fc molecules to expand flu-specific CTL (data not shown).
Figure 4.

Selective expansion of Ag-specific CTL using K32/4-1BBL/HLA-A2-Fc aAPCs. A, CD8 cells were stimulated using K32/4-1BBL/pp65-HLA-A2-Fc, K32/anti-CD28/pp65-HLA-A2-Fc, or K32/pp65-HLA-A2-Fc. On day 22 after three stimulations, the cells were harvested, counted, and analyzed by FACS. K32/4-1BBL/pp65 aAPCs expanded Ag-specific cells but not cells binding an irrelevant tetramer while K32/anti-CD28pp65 and K32/pp65 did not expand Ag-specific cells. B, K32/4-1BBL/pp65, but not K32/anti-CD28/pp65, expanded CTL demonstrate Ag-specific cytotoxicity as assessed by CD107a expression. CD8+ cells stimulated with K32/pp65 did not produce sufficient cell numbers to perform the assay. This is a representative result of three experiments using Ag-specific expansion from different HLA-A2+ donors.
Table I. Ag-specific expansion of CMV-reactive PBL using K32/4-1BBL/CMVpp65-HLA-A2-Fca.
| Total Number of pp65 Tetramer-Binding Cells (% Tetramer Binding) | ||||
|---|---|---|---|---|
|
| ||||
| Donor | Day 0 | Day 7 | Day 14 | Day 21 |
| 1 | 540 (0.05) | 61,440 (1.3) | 347,060 (5.2) | ND |
| 2 | 8,800 (0.9) | 25,350 (1.7) | 200,420 (9.1) | 313,560 (8.7) |
| 3 | 5,920 (0.37) | 12,100 (1.1) | 227,200 (7.1) | ND |
CD8+ PBL were stimulated on days 0, 7, and 14 with K32/4-1BBL bound to CMVpp65-pulsed HLA-A2-Fc fusion protein as described in Materials and Methods. Enumeration shown in the table was performed just prior to subsequent stimulation. ND, Third stimulation not performed in these cultures.
Anti-CD3/4-1BB signaling mediated up-regulation of NKG2D expression and enhancing cytolysis
Ag-specific CD8+ T cells that persist in hosts with high viral or tumor burdens often lack effective cytolytic activity (29–32) and, in some models, this is associated with NKG2D down-regulation, which can limit the capacity for cytolytic T and NK cells to kill MIC-expressing tumor cells (33, 34). Therefore, adoptive immunotherapy also seeks to augment the functional capacity of cells generated ex vivo, in addition to increasing numbers of Ag-specific T cells available to the host. We previously reported that CD8+ T cells expanded polyclonally with aAPCs using anti-CD3/anti-CD28 and anti-CD3/4-1BBL could mediate MHC-unrestricted cytotoxicity (35). We therefore sought to determine whether NKG2D, a killer-activating receptor (KAR) which can mediate MHC-unrestricted killing (36–38), was expressed on anti-CD3/4-1BBL-activated CD8+ T cells. Resting CD8+ PBL express high levels of NKG2D, but CD8+ PBL maintained in culture for 7 days or expanded using anti-CD3/anti-CD28 aAPC-based costimulation showed minimal NKG2D expression. In contrast, CD8+ T cells expanded using 4-1BBL costimulation maintain substantial NKG2D expression (Fig. 5A). To determine whether the NKG2D expression induced by anti-CD3/4-BBL-based expansion conferred NKG2D-signaling capacity and augmented cytotoxic function, we used a T cell-redirected cytotoxicity assay. Although neither fresh resting CD8+ T cells nor anti-CD3/anti-CD28-expanded CD8+ T cells were cytolytic in this assay (data not shown), anti-CD3/4-1BBL-expanded CD8+ PBL showed baseline cytolytic function as evidenced by lytic activity following cross-linking with anti-MHC class I, used as a negative control in this assay. Furthermore, cytolytic activity was substantially increased when either anti-NKG2D (p = 0.02) or anti-CD3 was added to the culture (p = 0.03), demonstrating that both NKG2D and CD3 signaling mediated a cytolytic signal in the expanded populations. Although anti-4-1BB signaling alone did not induce augmented cytolytic activity (p = 0.13), the highest cytolytic activity was observed when CD3 signaling was combined with anti-4-1BB or anti-NKG2D (Fig. 5B). Therefore, 4-1BB costimulation not only facilitates survival of memory populations in vivo as reported previously (39–41), and expansion of memory cells ex vivo as shown in this report, but also augments cytolytic function directly and indirectly via up-regulation of NKG2D.
Figure 5.

4-1BB signaling augments cytolytic activity of expanded CTL. A, Anti-CD3/4-1BBL aAPC-expanded CD8+ T cells up-regulate NKG2D expression. Purified human CD8 T cells were unstimulated, or stimulated with anti-CD3/4-1BBL or anti-CD3/anti-CD28 aAPC, then assessed for NKG2D expression (bold) on days 0 and 7. Isotype control is shown in gray. This is representative of three experiments from different donors. B, Cytolytic function of human CD8 T cells stimulated with anti-CD3/4-1BBL aAPCs 7 days prior was assayed using 51Cr-labeled P815 target cells cross-linked with the Abs indicated for 4 h. The bar graph is from eight separate experiments with individual donors. Resting CD8 cells and anti-CD3/anti-CD28 aAPC CD8 cells showed no detectable cytolytic activity (data not shown). CD3 and NKG2D signaling alone augment cytolytic function of anti-CD3/4-1BBL aAPC-expanded CD8+ T cells (p < 0.05) whereas cytolytic function after 4-1BB signaling alone was not significantly different than observed with anti-MHC class I. Combining 4-1BB and CD3 signaling or NKG2D and CD3 signaling significantly increases cytolytic activity compared with CD3 signaling alone (**, p = 0.008).
Discussion
Ex vivo T cell expansion for adoptive immunotherapy seeks to rapidly generate large numbers of functional T cells to amplify Ag-specific effector function in vivo. Several groups have demonstrated that autologous APCs, generated from a variety of cell types, can effectively expand cytolytic T cells (2, 10, 14, 15, 42). Although autologous APCs are ideal in principle because they are natural APCs, they are difficult to standardize, expensive to generate, and are not optimal for identifying critical costimulatory pathways that are most effective in inducing CTL expansion. aAPCs (aAPC), off-the-shelf reagents that eliminate the requirement for generating patient-specific APCs to provide TCR stimulation and costimulation to induce ex vivo T cell expansion, represent a potentially cost-effective and less labor-intensive approach to developing cytolytic T cell products for adoptive immunotherapy. In general, the vast majority of aAPCs under study have incorporated CD28 ligands as the primary costimulatory signal (43–47). However, we and others have noted that anti-CD3/anti-CD28 mAbs conjugated to magnetic beads do not effectively expand CD8+ T cells (17, 19, 48), and it is well-known that CD8+ populations rapidly down-regulate CD28 upon acquisition of effector function (49). Alternatively, several studies have also demonstrated a role for 4-1BB costimulation in CD8+ T cell expansion, although each of these studies also delivered CD28 signals during the phase of CD8+ T cell expansion (12, 19, 20). 4-1BB, a member of TNFR family expressed primarily on activated lymphocytes (23), promotes CD8+ T cell proliferation, differentiation, and inhibits programmed cell death (50–53). The 4-1BB ligand is expressed by activated B cells, dendritic cells, and monocytes/macrophages and likely serves to augment survival of cytolytic CD8+ T cells during the evolution of an immune response (39–41).
In this report, we used a system of genetically modified aAPCs to directly compare the efficacy of CD28 vs 4-1BB signaling in the expansion of CD8+ CTL. Because K32 cells are not natural APCs, they do not endogenously express a host of stimulatory and inhibitory ligands, and thus allowed us to directly assess whether 4-1BB costimulation was sufficient to induce CD8+ CTL expansion. The data demonstrate K32 cell-based aAPCs, which deliver either anti-CD3/anti-CD28 signals or anti-CD3/4-1BBL signals, promote rapid expansion of peripheral blood CD8 T cells with a median 1.5 log expansion in 14 days. Such expansion was greater than that observed using bead-based APCs, in part, due to the production of IL-15 and expression of IL-15Rα expression on the K32-based aAPCs. Interestingly, the quantitative increases in CD8 expansion could not be reproduced by adding IL-15 to the bead-based cell cultures, perhaps due to inadequate presentation in the absence of IL-15Rα. CD8 expansion was numerically similar following 4-1BBL or CD28 stimulation despite vastly different levels of expression of CD28 and 4-1BB on resting T cells. However, our studies uncovered substantial qualitative differences in the CD8+ populations expanded using 4-1BB vs CD28 costimulation. Whereas anti-CD3/4-1BBL aAPCs preferentially expand memory CD8 T cells, resulting in an increased frequency of cells responding to viral recall Ags in the expanded cultures from healthy donors, anti-CD3/anti-CD28 aAPCs preferentially expand naive CD8+ cells and therefore do not enrich for viral-specific CTL. Interestingly, we did not detect any difference in the frequency or extent of 4-1BB expression following anti-CD3 signaling on naive vs memory CD8+ T cells (data not shown), therefore the basis for the selective expansion of memory cells via 4-1BB signaling cannot be ascribed to differential expression of 4-1BB on these two populations. Furthermore, while we observed increased short-term expansion of polyclonal memory populations compared with naive populations using 4-1BB costimulation as shown in Fig. 2C, it remains possible that there are also significant survival differences between naive and memory populations that contribute to the results reported here. Indeed, ample data in murine models demonstrate that 4-1BB signaling plays a critical role in survival of memory CD8+ T cells during the later stages of the immune response (24–27).
One theoretical advantage to the use of CD28-based costimulation might be a preferential expansion and/or retention of CD28-expressing CD8+ T cells, because multiple studies have demonstrated that CD28+ cells retain a greater proliferative potential and are more functional than the CD28− subset (49). We therefore analyzed the CD28+/CD28− ratio of CD8 T cells pre- vs post-expansion using either K32-based anti-CD3/anti-CD28 or anti-CD3/4-1BBL stimulation. Whereas normal human CD8+ PBL showed a CD28+/CD28− ratio of ∼2:1, T cells expanded by K32-based anti-CD3/anti-CD28 and anti-CD3/4-1BBL aAPCs studied in this report resulted in a postexpansion ratio of 1:2 with no difference in the percent of cells expressing CD28 following anti-CD28 vs 4-1BBL costimulation (data not shown). Therefore, we found no evidence to suggest that CD28 costimulation was superior to 4-1BB costimulation in retaining or expanding CD28+ CTL.
In addition to demonstrating that 4-1BB costimulation preferentially expands memory CTL and the demonstration that 4-1BB costimulation alone is sufficient to induce dramatic expansions of CD8+ CTL, this report is also the first to demonstrate that 4-1BB signaling directly augments cytotoxic function in human CD8+ T cells. Furthermore, the up-regulation of NKG2D on 4-1BBL-co-stimulated cells potentially represents an important secondary mediator of cytolysis which may be particularly pertinent for adoptive immunotherapy, because NKG2D down-regulation contributes to immune escape in murine models of cancer and chronic infection (33, 34). The capacity for 4-1BBL-expressing aAPCs to generate a cell product with high levels of NKG2D that can augment cytolytic capacity is potentially favorable to avoid tumor immune escape by down-regulation of either MHC or shedding of NKG2D ligands. Indeed, consistent with this notion, we demonstrated previously that CD8+ T cells expanded with anti-CD3/4-1BBL-mediated non-MHC-restricted killing of tumor targets and mediated antitumor activity in vivo in a xenograft model whereas anti-CD3/anti-CD28-expanded CD8+ T cells did not mediate antitumor activity in vivo (35).
In conclusion, these results provide the first evidence that CD28 costimulation is not essential for generating CD8+ T cells for adoptive immunotherapy. Rather, TCR and/or CD3 signals combined with 4-1BB costimulation can generate large numbers of cytolytic CD8+ T cells with enhanced effector functions that are suitable for adoptive cellular immunotherapy. The expansion of electronically sorted populations or enrichment of large numbers of Ag-specific cells from polyclonal peripheral blood CD8+ T cells using K32/4-1BBL-HLA-A2-Fc APCs provides opportunities for expanding large numbers of Ag-specific T cells with MHC-restricted cytolytic activity. Furthermore, this approach yields CD8+ T cells with enhanced MHC-restricted cytolytic activity and the potential for non-MHC-restricted cytolysis as a result of NKG2D up-regulation, which is potentially important for the elimination of infected and/or transformed cells which can down-regulate MHC expression. Considering the wide range of diseases that are potentially amenable to treatment given a robust, reproducible approach for ex vivo expansion of Ag-specific CTLs, 4-1BBL-expressing cell-based aAPCs hold promise for improving the effectiveness of adoptive immunotherapy.
Acknowledgments
We thank Drs. Terry Fry and Ron Gress for their careful and critical reviews of this manuscript. H. Zhang performed most of the experiments shown, analyzed data, and partially wrote the manuscript. K. M. Snyder and M. M. Suhoski performed experiments demonstrating the capacity for aAPCs to expand purified populations of Ag-specific cells. M. M. Suhoski and M. V. Maus created the aAPCs, performed experiments, and analyzed data. C. H. June provided the intellectual insight to use 4-1BBL as a costimulatory molecule in aAPCs and provided the financial support for the creation of these cells. C. L. Mackall provided intellectual and financial support for the experiments shown, analyzed data, and partially wrote the manuscript.
Footnotes
This work was supported, in part, by the Intramural Research Program of the National Cancer Institute.
Abbreviations used in this paper: LBL, lymphoblast cell line; aAPC, artificial APC.
Disclosures: M. V. Maus and C. H. June have applied for a patent related to the work that is described in the present study.
References
- 1.Walter EA, Greenberg PD, Gilbert MJ, Finch RJ, Watanabe KS, Thomas ED, Riddell SR. Reconstitution of cellular immunity against cytomegalovirus in recipients of allogeneic bone marrow by transfer of T-cell clones from the donor. N Engl J Med. 1995;333:1038–1044. doi: 10.1056/NEJM199510193331603. [DOI] [PubMed] [Google Scholar]
- 2.Leen AM, Myers GD, Sili U, Huls MH, Weiss H, Leung KS, Carrum G, Krance RA, Chang CC, Molldrem JJ, et al. Monoculture-derived T lymphocytes specific for multiple viruses expand and produce clinically relevant effects in immunocompromised individuals. Nat Med. 2006;12:1160–1166. doi: 10.1038/nm1475. [DOI] [PubMed] [Google Scholar]
- 3.Brodie SJ, Lewinsohn DA, Patterson BK, Jiyamapa D, Krieger J, Corey L, Greenberg PD, Riddell SR. In vivo migration and function of transferred HIV-1-specific cytotoxic T cells. Nat Med. 1999;5:34–41. doi: 10.1038/4716. [DOI] [PubMed] [Google Scholar]
- 4.Savoldo B, Huls MH, Liu Z, Okamura T, Volk HD, Reinke P, Sabat R, Babel N, Jones JF, Webster-Cyriaque J, et al. Autologous Epstein-Barr virus (EBV)-specific cytotoxic T cells for the treatment of persistent active EBV infection. Blood. 2002;100:4059–4066. doi: 10.1182/blood-2002-01-0039. [DOI] [PubMed] [Google Scholar]
- 5.Straathof KC, Bollard CM, Popat U, Huls MH, Lopez T, Morriss MC, Gresik MV, Gee AP, Russell HV, Brenner MK, et al. Treatment of nasopharyngeal carcinoma with Epstein-Barr virus-specific T lymphocytes. Blood. 2005;105:1898–1904. doi: 10.1182/blood-2004-07-2975. [DOI] [PubMed] [Google Scholar]
- 6.Bollard CM, Aguilar L, Straathof KC, Gahn B, Huls MH, Rousseau A, Sixbey J, Gresik MV, Carrum G, Hudson M, et al. Cytotoxic T lymphocyte therapy for Epstein-Barr virus+ Hodgkin's disease. J Exp Med. 2004;200:1623–1633. doi: 10.1084/jem.20040890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yee C, Thompson JA, Byrd D, Riddell SR, Roche P, Celis E, Greenberg PD. Adoptive T cell therapy using antigen-specific CD8+ T cell clones for the treatment of patients with metastatic melanoma: in vivo persistence, migration, and antitumor effect of transferred T cells. Proc Natl Acad Sci USA. 2002;99:16168–16173. doi: 10.1073/pnas.242600099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dudley ME, Wunderlich JR, Robbins PF, Yang JC, Hwu P, Schwartzentruber DJ, Topalian SL, Sherry R, Restifo NP, Hubicki AM, et al. Cancer regression and autoimmunity in patients after clonal repopulation with antitumor lymphocytes. Science. 2002;298:850–854. doi: 10.1126/science.1076514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Overwijk WW, Theoret MR, Finkelstein SE, Surman DR, de Jong LA, Vyth-Dreese FA, Dellemijn TA, Antony PA, Spiess PJ, Palmer DC, et al. Tumor regression and autoimmunity after reversal of a functionally tolerant state of self-reactive CD8+ T cells. J Exp Med. 2003;198:569–580. doi: 10.1084/jem.20030590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Sun Q, Pollok KE, Burton RL, Dai LJ, Britt W, Emanuel DJ, Lucas KG. Simultaneous ex vivo expansion of cytomegalovirus and Epstein-Barr virus-specific cytotoxic T lymphocytes using B-lymphoblastoid cell lines expressing cytomegalovirus pp65. Blood. 1999;94:3242–3250. [PubMed] [Google Scholar]
- 11.von Bergwelt-Baildon MS, Vonderheide RH, Maecker B, Hirano N, Anderson KS, Butler MO, Xia Z, Zeng WY, Wucherpfennig KW, Nadler LM, Schultze JL. Human primary and memory cytotoxic T lymphocyte responses are efficiently induced by means of CD40-activated B cells as antigen-presenting cells: potential for clinical application. Blood. 2002;99:3319–3325. doi: 10.1182/blood.v99.9.3319. [DOI] [PubMed] [Google Scholar]
- 12.Bukczynski J, Wen T, Ellefsen K, Gauldie J, Watts TH. Costimulatory ligand 4-1BBL (CD137L) as an efficient adjuvant for human antiviral cytotoxic T cell responses. Proc Natl Acad Sci USA. 2004;101:1291–1296. doi: 10.1073/pnas.0306567101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hammer MH, Meyer S, Brestrich G, Moosmann A, Kern F, Tesfa L, Babel N, Mittenzweig A, Rooney CM, Hammerschmidt W, et al. HLA type-independent generation of antigen-specific T cells for adoptive immunotherapy. Eur J Immunol. 2005;35:2250–2258. doi: 10.1002/eji.200526230. [DOI] [PubMed] [Google Scholar]
- 14.Szmania S, Galloway A, Bruorton M, Musk P, Aubert G, Arthur A, Pyle H, Hensel N, Ta N, Lamb L, Jr, et al. Isolation and expansion of cytomegalovirus-specific cytotoxic T lymphocytes to clinical scale from a single blood draw using dendritic cells and HLA-tetramers. Blood. 2001;98:505–512. doi: 10.1182/blood.v98.3.505. [DOI] [PubMed] [Google Scholar]
- 15.Peggs K, Verfuerth S, Mackinnon S. Induction of cytomegalovirus (CMV)-specific T-cell responses using dendritic cells pulsed with CMV antigen: a novel culture system free of live CMV virions. Blood. 2001;97:994–1000. doi: 10.1182/blood.v97.4.994. [DOI] [PubMed] [Google Scholar]
- 16.Greenwald RJ, Freeman GJ, Sharpe AH. The B7 family revisited. Annu Rev Immunol. 2005;23:515–548. doi: 10.1146/annurev.immunol.23.021704.115611. [DOI] [PubMed] [Google Scholar]
- 17.Laux I, Khoshnan A, Tindell C, Bae D, Zhu X, June CH, Effros RB, Nel A. Response differences between human CD4+ and CD8+ T-cells during CD28 costimulation: implications for immune cell-based therapies and studies related to the expansion of double-positive T-cells during aging. Clin Immunol. 2000;96:187–197. doi: 10.1006/clim.2000.4902. [DOI] [PubMed] [Google Scholar]
- 18.Deeths MJ, Kedl RM, Mescher MF. CD8+ T cells become nonresponsive (anergic) following activation in the presence of costimulation. J Immunol. 1999;163:102–110. [PubMed] [Google Scholar]
- 19.Maus MV, Thomas AK, Leonard DG, Allman D, Addya K, Schlienger K, Riley JL, June CH. Ex vivo expansion of polyclonal and antigen-specific cytotoxic T lymphocytes by artificial APCs expressing ligands for the T-cell receptor, CD28 and 4-1BB. Nat Biotechnol. 2002;20:143–148. doi: 10.1038/nbt0202-143. [DOI] [PubMed] [Google Scholar]
- 20.Thomas AK, Maus MV, Shalaby WS, June CH, Riley JL. A cell-based artificial antigen-presenting cell coated with anti-CD3 and CD28 antibodies enables rapid expansion and long-term growth of CD4 T lymphocytes. Clin Immunol. 2002;105:259–272. doi: 10.1006/clim.2002.5277. [DOI] [PubMed] [Google Scholar]
- 21.Suhoski MM, Golovina TN, Aqui NA, Tai VC, Varela-Rohena A, Milone MC, Carroll RG, Riley JL, June CH. Engineering artificial antigen-presenting cells to express a diverse array of co-stimulatory molecules. Mol Ther. 2007;15:981–988. doi: 10.1038/mt.sj.6300134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Betts MR, Brenchley JM, Price DA, De Rosa SC, Douek DC, Roederer M, Koup RA. Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation. J Immunol Methods. 2003;281:65–78. doi: 10.1016/s0022-1759(03)00265-5. [DOI] [PubMed] [Google Scholar]
- 23.Pollok KE, Kim YJ, Zhou Z, Hurtado J, Kim KK, Pickard RT, Kwon BS. Inducible T cell antigen 4-1BB: analysis of expression and function. J Immunol. 1993;150:771–781. [PubMed] [Google Scholar]
- 24.Pulle G, Vidric M, Watts TH. IL-15-dependent induction of 4-1BB promotes antigen-independent CD8 memory T cell survival. J Immunol. 2006;176:2739–2748. doi: 10.4049/jimmunol.176.5.2739. [DOI] [PubMed] [Google Scholar]
- 25.DeBenedette MA, Wen T, Bachmann MF, Ohashi PS, Barber BH, Stocking KL, Peschon JJ, Watts TH. Analysis of 4-1BB ligand (4-1BBL)-deficient mice and of mice lacking both 4-1BBL and CD28 reveals a role for 4-1BBL in skin allograft rejection and in the cytotoxic T cell response to influenza virus. J Immunol. 1999;163:4833–4841. [PubMed] [Google Scholar]
- 26.Croft M. Co-stimulatory members of the TNFR family: keys to effective T-cell immunity? Nat Rev Immunol. 2003;3:609–620. doi: 10.1038/nri1148. [DOI] [PubMed] [Google Scholar]
- 27.Cannons JL, Lau P, Ghumman B, DeBenedette MA, Yagita H, Okumura K, Watts TH. 4-1Bb ligand induces cell division, sustains survival, and enhances effector function of CD4 and CD8 t cells with similar efficacy. J Immunol. 2001;167:1313–1324. doi: 10.4049/jimmunol.167.3.1313. [DOI] [PubMed] [Google Scholar]
- 28.Boon T, van der Bruggen P. Human tumor antigens recognized by T lymphocytes. J Exp Med. 1996;183:725–729. doi: 10.1084/jem.183.3.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zajac AJ, Blattman JN, Murali-Krishna K, Sourdive DJ, Suresh M, Altman JD, Ahmed R. Viral immune evasion due to persistence of activated T cells without effector function. J Exp Med. 1998;188:2205–2213. doi: 10.1084/jem.188.12.2205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rehermann B, Nascimbeni M. Immunology of hepatitis B virus and hepatitis C virus infection. Nat Rev Immunol. 2005;5:215–229. doi: 10.1038/nri1573. [DOI] [PubMed] [Google Scholar]
- 31.Pantaleo G, Koup RA. Correlates of immune protection in HIV-1 infection: what we know, what we don't know, what we should know. Nat Med. 2004;10:806–810. doi: 10.1038/nm0804-806. [DOI] [PubMed] [Google Scholar]
- 32.Barber DL, Wherry EJ, Masopust D, Zhu B, Allison JP, Sharpe AH, Freeman GJ, Ahmed R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature. 2006;439:682–687. doi: 10.1038/nature04444. [DOI] [PubMed] [Google Scholar]
- 33.Jinushi M, Hodi FS, Dranoff G. Therapy-induced antibodies to MHC class I chain-related protein A antagonize immune suppression and stimulate antitumor cytotoxicity. Proc Natl Acad Sci USA. 2006;103:9190–9195. doi: 10.1073/pnas.0603503103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wiemann K, Mittrucker HW, Feger U, Welte SA, Yokoyama WM, Spies T, Rammensee HG, Steinle A. Systemic NKG2D down-regulation impairs NK and CD8 T cell responses in vivo. J Immunol. 2005;175:720–729. doi: 10.4049/jimmunol.175.2.720. [DOI] [PubMed] [Google Scholar]
- 35.Zhang H, Merchant MS, Chua KS, Khanna C, Helman LJ, Telford B, Ward Y, Summers J, Toretsky J, Thomas EK, et al. Tumor expression of 4-1BB ligand sustains tumor lytic T cells. Cancer Biol Ther. 2003;2:579–586. doi: 10.4161/cbt.2.5.545. [DOI] [PubMed] [Google Scholar]
- 36.Bauer S, Groh V, Wu J, Steinle A, Phillips JH, Lanier LL, Spies T. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science. 1999;285:727–729. doi: 10.1126/science.285.5428.727. [DOI] [PubMed] [Google Scholar]
- 37.Groh V, Rhinehart R, Randolph-Habecker J, Topp MS, Riddell SR, Spies T. Costimulation of CD8αβ T cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat Immunol. 2001;2:255–260. doi: 10.1038/85321. [DOI] [PubMed] [Google Scholar]
- 38.Verneris MR, Karami M, Baker J, Jayaswal A, Negrin RS. Role of NKG2D signaling in the cytotoxicity of activated and expanded CD8+ T cells. Blood. 2004;103:3065–3072. doi: 10.1182/blood-2003-06-2125. [DOI] [PubMed] [Google Scholar]
- 39.Goodwin RG, Din WS, Davis-Smith T, Anderson DM, Gimpel SD, Sato TA, Maliszewski CR, Brannan CI, Copeland NG, Jenkins NA, et al. Molecular cloning of a ligand for the inducible T cell gene 4-1BB: a member of an emerging family of cytokines with homology to tumor necrosis factor. Eur J Immunol. 1993;23:2631–2641. doi: 10.1002/eji.1830231037. [DOI] [PubMed] [Google Scholar]
- 40.Pollok KE, Kim YJ, Hurtado J, Zhou Z, Kim KK, Kwon BS. 4-1BB T-cell antigen binds to mature B cells and macrophages, and costimulates anti-μ-primed splenic B cells. Eur J Immunol. 1994;24:367–374. doi: 10.1002/eji.1830240215. [DOI] [PubMed] [Google Scholar]
- 41.DeBenedette MA, Shahinian A, Mak TW, Watts TH. Costimulation of CD28− T lymphocytes by 4-1BB ligand. J Immunol. 1997;158:551–559. [PubMed] [Google Scholar]
- 42.Riddell SR, Watanabe KS, Goodrich JM, Li CR, Agha ME, Greenberg PD. Restoration of viral immunity in immunodeficient humans by the adoptive transfer of T cell clones. Science. 1992;257:238–241. doi: 10.1126/science.1352912. [DOI] [PubMed] [Google Scholar]
- 43.Levine BL, Bernstein WB, Connors M, Craighead N, Lindsten T, Thompson CB, June CH. Effects of CD28 costimulation on long-term proliferation of CD4+ T cells in the absence of exogenous feeder cells. J Immunol. 1997;159:5921–5930. [PubMed] [Google Scholar]
- 44.Papanicolaou GA, Latouche JB, Tan C, Dupont J, Stiles J, Pamer EG, Sadelain M. Rapid expansion of cytomegalovirus-specific cytotoxic T lymphocytes by artificial antigen-presenting cells expressing a single HLA allele. Blood. 2003;102:2498–2505. doi: 10.1182/blood-2003-02-0345. [DOI] [PubMed] [Google Scholar]
- 45.Latouche JB, Sadelain M. Induction of human cytotoxic T lymphocytes by artificial antigen-presenting cells. Nat Biotechnol. 2000;18:405–409. doi: 10.1038/74455. [DOI] [PubMed] [Google Scholar]
- 46.Dupont J, Latouche JB, Ma C, Sadelain M. Artificial antigen-presenting cells transduced with telomerase efficiently expand epitope-specific, human leukocyte antigen-restricted cytotoxic T cells. Cancer Res. 2005;65:5417–5427. doi: 10.1158/0008-5472.CAN-04-2991. [DOI] [PubMed] [Google Scholar]
- 47.Oelke M, Maus MV, Didiano D, June CH, Mackensen A, Schneck JP. Ex vivo induction and expansion of antigen-specific cytotoxic T cells by HLA-Ig-coated artificial antigen-presenting cells. Nat Med. 2003;9:619–624. doi: 10.1038/nm869. [DOI] [PubMed] [Google Scholar]
- 48.Deeths MJ, Mescher MF. B7-1-dependent co-stimulation results in qualitatively and quantitatively different responses by CD4+ and CD8+ T cells. Eur J Immunol. 1997;27:598–608. doi: 10.1002/eji.1830270305. [DOI] [PubMed] [Google Scholar]
- 49.Azuma M, Phillips JH, Lanier LL. CD28− T lymphocytes: antigenic and functional properties. J Immunol. 1993;150:1147–1159. [PubMed] [Google Scholar]
- 50.Smith CA, Farrah T, Goodwin RG. The TNF receptor superfamily of cellular and viral proteins: activation, costimulation, and death. Cell. 1994;76:959–962. doi: 10.1016/0092-8674(94)90372-7. [DOI] [PubMed] [Google Scholar]
- 51.Hurtado JC, Kim YJ, Kwon BS. Signals through 4-1BB are costimulatory to previously activated splenic T cells and inhibit activation-induced cell death. J Immunol. 1997;158:2600–2609. [PubMed] [Google Scholar]
- 52.Shuford WW, Klussman K, Tritchler DD, Loo DT, Chalupny J, Siadak AW, Brown TJ, Emswiler J, Raecho H, Larsen CP, et al. 4-1BB costimulatory signals preferentially induce CD8+ T cell proliferation and lead to the amplification in vivo of cytotoxic T cell responses. J Exp Med. 1997;186:47–55. doi: 10.1084/jem.186.1.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Takahashi C, Mittler RS, Vella AT. Cutting edge: 4-1BB is a bona fide CD8 T cell survival signal. J Immunol. 1999;162:5037–5040. [PubMed] [Google Scholar]