

Abstract
Anthropogenic fire is a form of ecosystem engineering that creates greater landscape patchiness at small spatial scales: such rescaling of patch diversity through mosaic burning has been argued to be a form of niche construction, the loss of which may have precipitated the decline and extinction of many endemic species in the Western Desert of Australia. We find evidence to support this hypothesis relative to one keystone species, the sand monitor lizard (Varanus gouldii). Paradoxically, V. gouldii populations are higher where Aboriginal hunting is most intense. This effect is driven by an increase in V. gouldii densities near successional edges, which is higher in landscapes that experience extensive human burning. Over time, the positive effects of patch mosaic burning while hunting overwhelm the negative effects of predation in recently burned areas to produce overall positive impacts on lizard populations. These results offer critical insights into the maintenance of animal communities in the desert, supporting the hypothesis that the current high rate of endemic species decline among small animals may be linked to the interaction between invasive species and mid-century removal of Aboriginal niche construction through hunting and patch mosaic burning.
Keywords: anthropogenic fire, ecosystem engineering, hunter–gatherers, trophic interactions, facilitation, coevolution
1. Introduction
Despite the increasing attention to ecosystem engineering as an important constructive force in the ecology and evolution of biotic communities [1,2], humans are widely perceived primarily through their destructive effects on vulnerable prey communities. While there are many examples of negative effects, such as population depression, extinction and niche destruction [3,4]; positive effects, such as enhancing another species' population growth or constructing new niches to support a wider range of other species, are difficult to find. Yet humans, particularly hunter–gatherers, do have the potential to be significant ‘ecosystem engineers [5]’: organisms that physically modify the environment in ways that benefit other species as well as themselves [6–8]. When hunter–gatherers employ patch mosaic burning over large spatial extents, landscape heterogeneity is enhanced, which may stabilize species interactions [9–11] and provide rescaling and habitat protection effects for species that require a variety of habitats for both food and shelter [12–14]. When hunter–gatherers have broad-spectrum diets and hunt many different prey species at multiple trophic levels [15], food web stability might increase, allowing more species to persist with more stable populations than in the absence of such generalized predation [16].
Biotic communities characterized by substantial histories of such engineering should eventually become adapted to these niche-constructing activities, even dependent upon them. These kinds of evolutionary feedbacks are commonly observed through removal experiments, either natural or artificial: the removal of a potent ecosystem engineer should have cascading effects on many other species [17]. For example, sea otters prevent urchins from denuding kelp beds and promote the return of sea grasses [18,19], providing habitat for many species of fish and shellfish: in the absence of otters, vegetation disappears as well as the fish that depend on it. Such effects have been demonstrated for a variety of other species, but evidence for human niche construction has been limited primarily to the introduction of novel species, mainly through domestication [7].
Yet, plant domestication is not the only type of niche construction: hunter–gatherers also have the potential to have positive impacts on ecosystems, especially where their life ways have persisted in situ for millennia. Australia is one continent where we are likely to find such evidence. In the arid desert grasslands that cover much of the continent, human foraging has been part of the landscape for at least the last 36 000 years or more [20,21], and in some places, still persists. Desert foraging strategies (at least as documented ethnohistorically) have commonly incorporated the use of fire to increase hunting returns. Hunting effort of both men and women was focused around the acquisition of small-to-medium-sized animals, both mammals and herpetofauna, with the occasional larger prey (kangaroo, Macropus robustus and Macropus rufus and emu, Dromaius novaehollandiae) [22–24]. Throughout the desert, fires were used primarily during the cool-dry season to clear areas of mature spinifex (Triodia spp.) grassland in sandplain and dune country to facilitate search for burrowed prey, especially sand monitor (Varanus gouldii) and other herpetofauna, but also small mammals, such as bilby (Macrotis lagotis, now rare), burrowing bettong (Bettongia lesueur, extinct in the Australian mainland) and rufous hare-wallaby (Lagorchestes hirsutus, highly endangered). Spot fires for flushing prey were used while hunting for larger monitors (Varanus giganteus and Varanus panoptes), brushtail possum (Trichosurus vulpecula, now rare in the desert) and the feral house-cat (Felis domesticus). Plants found at high density only in recently burned areas were dietary staples: Solanum diversiflorum and Solanum centrale, and grass seeds, such as Eragrostis and Panicum. Desert foragers moved across an extensive landscape, concentrating their hunting and burning around reliable water, and moving on to new camps when hunting returns declined.
Entangled within the practice of foraging is a body of knowledge concerning the nature of the relationships between humans and other species, and how those relationships were maintained: the Dreaming, what Aboriginal people in the Western Desert refer to as Jukurr. The Jukurrpa is described by Aboriginal people variously as ‘Law’, ‘Business’ or ‘University’, as cosmology, philosophy and natural history. It explains ‘not just what animals do and where plants grow, but why they live the way they do and how they are related to each other and to the ancestral beings’ (P. Taylor 2003, personal communication). It contains information about the origins of life and the mechanisms of its maintenance, including detailed information about ecological interactions, down to the minutia of which species of skink returns to the same location to defaecate and which mouse prefers recently burnt spinifex. Critical to the perpetuation of life is the proper adherence to Jukurrpa, which frames the importance of fire within the notion that the country must be used if life is to continue. As one elder puts it, to stop hunting and to stop burning would mean ‘the end of the world’ (P. Biljabu 2002, personal communication). Many Aboriginal people believe that if they do not continue to re-enact the Jukurrpa through emulating the creative forces of the ancestral beings across the landscape—hunting, collecting, burning and caring for family—those plants and animals that depend on their actions will cease to exist. This belief has important material ecological expression.
Most of the last nomadic groups of desert foragers left the desert regions between the late 1950s and early 1970s, moving towards missions and pastoral stations around the desert's margins. During that time, somewhere between 10 and 20 native species went extinct, 43 more went into sharp decline, and the landscape was dominated by extensive lightening fires [25,26]. Analysis of aerial photography and satellite imagery covering a large tract (241 210 ha) of Western Desert grassland demonstrated significant shifts in mean fire size, from 64 ha in 1953, when Aboriginal foragers were present, to more than 52 000 ha in 1984 [26]. In 1984, a group of Manyjiljarra, Kartujarra and Warnman-speaking desert dwellers (Martu) returned to their traditional country in the Little Sandy Desert after a 20-year absence and immediately lamented the state of the land and especially its perceived scarcity of edible foods (F. Walsh 2008, unpublished data). They began to burn small fires near camps and along tracks, paths and roads leading to hunting locations and water sources. They adopted new technologies (vehicles, rifles, etc.) to increase foraging efficiency in a degraded landscape and imported supplemental food sources (purchased goods). Since the 1980s, Aboriginal fires in localized regions have restored a finer-grained, more diverse successional mosaic, which prevents the spread of large lightening fires and actually protects more mature vegetation from burning. In regions without Aboriginal fires, far from communities and hunting camps and tracks, lightening is still the major source of fire ignition: fires are much larger, farther apart and more habitat is burnt [27,28]. Because Martu burn when they hunt small prey, hunting intensity is correlated with the number and size of fires on the landscape, leading to a finer-grained mosaic of patches in different stages of regrowth (successional patch) compared with lightening-dominated mosaics (see the electronic supplementary material). As use of a region by Martu hunters increases, the mean number of successional patches increases, causing successional patch diversity (Simpson's Diversity Index), edge density (the metres of contrast between two different fire ages per square metre) and the amount of unburned area to increase [27,28].
The solution to the mystery of species extinctions in the Western Desert may lie at least in part, in linking Dreamtime logic to these ecological consequences. The belief that it is not hunting that causes prey decline, but the lack of it, seems paradoxical, and to some, evidence of a religiously mandated mythology that has little to do with ecological reality [29]. Here, we propose one ecological hypothesis that is consistent with (if not part of) Dreamtime logic: that the creation of finer-grained habitat mosaics through the application of fire (patch mosaic burning) in the course of hunting acts as a type of ecosystem engineering that provides net positive effects to some species, including humans themselves. Over time, these ecosystem engineering effects change the availability of habitat niches, supporting a range of species that would not otherwise persist in this desert region: such evolutionary consequences would turn ecosystem engineering into a form of niche construction [17]. These positive effects for some species could come about because finer-grained habitat mosaics increase returns for high-ranked resources through reducing search or pursuit costs, or reduce mortality rates from non-human predators (including invasive cats and foxes, which are now primary sources of mortality) by reducing distance between refuge and foraging locations. The fact that today, the landscape is segregated into areas of intense versus no human influence, provides a natural ‘removal’ experiment through which we can test for the ecosystem engineering effects on species distributions and human foraging returns.
Here, we investigate the interactive effects of patch mosaic burning and Martu hunting on sand monitor (V. gouldii) population density. The half-kilogram sand monitor is a generalist forager that pursues a wide variety of animal prey, especially smaller lizards, insects and arachnids [30]. Over 50% of contemporary Martu foraging time is devoted to hunting for V. gouldii. Foraging efficiency is constrained by visibility in spinifex-dominated sandplain, so foragers either target recently burned areas, or burn their own patches for immediate search. Herpetofauna enter a period of brumation during the cool-dry season (May to September) and most hunting fires are set during this time in habitat preferred for winter burrows. Hunters (more often women) work alone, or in small cooperative groups to burn an area of oldergrowth spinifex, search for fresh burrows, then probe with a long, narrow digging stick to search for the resting chamber, which usually lies 10–20 cm below the ground surface. Summer season hunts when varanids are more mobile target early successional habitat with good track visibility. While human hunting may provide direct negative effects on V. gouldii populations, it may also provide indirect positive effects either by increasing habitat suitability through patch burning or by removing other predators, such as giant perenties (V. giganteus), argus monitors (V. panoptes) and feral cats [31].
We tease out the linked effects of hunting and patch mosaic burning by examining landscapes at varying travel distances from Martu communities: given that Martu are central-place foragers, as travel distance increases, both use of fire and intensity of hunting on V. gouldii should decrease. However, hunting V. gouldii is linked to the immediate presence of fire: in heavy-use landscapes, burned patches are likely to be where Martu have removed prey. If there is a net positive effect of patch mosaic burning on V. gouldii, we should see an increase in prey density with landscape use (figure 1). Alternatively, if there is a net negative effect of hunting, we would expect to see a net decrease in prey density with use. However, it is likely that there are both positive and negative effects at different spatial scales and that the strength of the negative effect depends on whether a local area has been burnt and prey removed by human hunters. If so, we should see an overall increase in V. gouldii density with use in unburned patches (reflecting the positive effects of ecosystem engineering through patch mosaic burning), but depending on the balance of positive and negative forces, either a decline or an intermediate peak in density in burned patches, reflecting the negative effects of human hunting. Martu landscape use should show positive feedback effects to human hunters: here we measure such effects as % of harvests with no prey captured, harvest size (kcals of V. gouldii), and return rate (kcals V. gouldii per hour spent in search and pursuit). All of these metrics should increase consistently or show an intermediate peak with use if burning is a form of ecosystem engineering.
Figure 1.
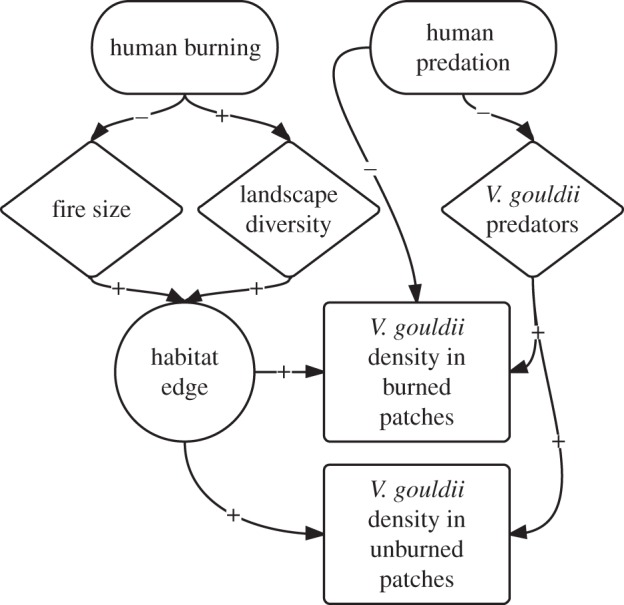
Flowchart illustrating the hypothetical process through which Martu hunting and burning affects sand monitor population density. Over the long term, human burning reduces mean fire size and increases diversity of regrowth patches across the landscape, increasing habitat edge density and providing positive feedbacks to V. gouldii populations. At the same time, human predation removes V. gouldii from recently burned areas (providing a negative feedback effect), but also removes V. gouldii predators (providing an indirectly positive effect).
2. Material and methods
The study region includes more than 46 000 km2 of the Great and Little Sandy Deserts bioregion of Western Australia (see the electronic supplementary material, figure S1). Within this area, there are four primary ecological communities: (i) spinifex (Triodia schinzii and Triodia basedowii) and Acacia (Acacia pachycarpa and Acacia ligulata, among others) dominated sandplains and dunes covering 85.6% of the total land area, (ii) lateric uplands and clay-dominated soils with mulga (Acacia aneura) woodland (2.4%) and Cassia shrubland (1.1%), (iii) Triodia-dominated but poorly vegetated rocky ranges (7.3%) and (iv) Eucalyptus (mainly Eucalyptus victrix and Eucalyptus microtheca)-dominated watercourse margins and floodplains (3.2%). This area includes a large portion of the Martu Aboriginal Native Title and Karlamilyi (Rudall River) National Park in which two Martu communities (Parnngurr and Punmu) are located.
Martu foraging data are available from 347 sample days and 4461 focal person-hours of search and pursuit in hunting and collecting (all seasons, June 2000–September 2010). The hunting returns used in the analysis below come from this dataset, in which on each sample day, we accompanied a foraging group from the community (Parnngurr or Punmu), usually in a vehicle, and recorded the location of the ‘dinner camp’, a centrally located cooking hearth which served as the temporary base for the foraging group and followed at least one hunter as they walked on foot from the dinner camp, which marked the beginning and end of each individual foraging bout. There were a total of 266 separate dinner camps for which spatial information and foraging data are complete. Each foraging bout (n = 1811 adult bouts, 104 different foragers) includes the number, type and weight of all resources acquired by each participant in the foraging party, total time each spent in search, pursuit and processing of each resource type, as well as time spent in burning and the location of ignition points. In our sample, Martu harvested 0.83 V. gouldii per adult forager per day hunting occurred. This sums to 1754 V. gouldii over 315 foraging days (1882 person-days), captured in 705 hunts. Energetic returns are calculated as net energy gained per forager per hour in search and pursuit. Net energy calculations are estimated based on a standard value of 3 kcal min–1, the cost of walking by a 60 kg individual at 3.2 km h–1.
Martu are central-place foragers, and as such, the intensity with which any region is searched (forager-hours per square kilometre) should be, all else being equal, a function of travel costs from the central place. Prey depletion is thus more likely in regions that have lower (time) costs of travel. To determine how human activity and predation rates vary across the landscape, we mapped vehicle tracks used during foraging trips with a hand-held GPS and used them to construct 12 separate buffer regions at varying distances from roads and communities. We then analysed the spatial distribution of the 266 foraging camps across each of the 12 buffers. Out of all foraging camps 50.2% were established within 20 km of the community (about 10–20 min one-way travel time) and less than 500 m from an established vehicle track: buffers within this distance were classified rank 4. A total of 38.4% of camps (rank 3) were located close to tracks between 20 and 50 km from a community, or close to the community but between 500 and 1500 m from a track. A total of 10.4% of all camps (rank 2) were within 20–50 km from a community and between 500 and 1500 m from a track, and more than 50 km and between 1500 and 5000 m from a track. The regions with the lowest human impact (rank 1, less than 1% of all camps) were those at all distances from communities that were more than 5000 m from a track. The intensity of use of each ranked region is calculated as forager-days per square kilometre observed in our sample (see electronic supplementary material, figure S1).
To survey V. gouldii populations, we conducted a series of ten 10 × 10 m linear pedestrian transects roughly perpendicular to vehicle tracks at varying distances from Parnngurr and Punmu communities during June and July of 2003. Each transect was subdivided into plots of 100 m2 in which we enumerated all visible new burrows of V. gouldii and estimated the proportion of spinifex cover and successional stage. There were 368 plots in use-rank 4, 2921 in rank 3, 4282 in rank 2, and 607 in rank 1. Adjacent plots across transects exhibited no significant autocorrelation. Only fresh, likely burrow sites were counted (e.g. those with a loose mound of soil at the entrance): feeding holes (e.g. no or minimal soil mound at the entrance) were not enumerated. The criterion for burrow identification was developed by one of us, N.T., who is a Martu hunter herself. As this survey was conducted in the Austral winter, V. gouldii were brumating in their winter burrows. In this analysis, we use only plots falling entirely in spinifex sandplain and dunefield. Each plot fell entirely within a single successional stage: for this analysis, we collapse patch time since fire into two stages, approximately less than 5 years since fire (burned) and approximately more than 5 years since fire (unburned). This successional stage classification corresponds to the Martu distinction between ‘regrowing’ (waru-waru and nyukura) and ‘burnable’ (manguu and kunarka) vegetation. Analysis was conducted in JMP, v. 10, SAS Institute Inc.
3. Results
(a). Effects of use on lizard density
Paradoxically, lizards are denser where they are hunted most often. The mean percentage of plots with lizards present increases from 7.6 ± 1.8% (95% CI) in lowest use landscapes (use rank 1), to 9.9 ± 0.9 in rank 2, to 11.9 ± 1.1 in rank 3, to 13.0 ± 3.1% in landscapes under highest human hunting pressure (rank 4) (logistic regression, n = 8178, χ2 = 16.10, p = 0.0011). Successional stage does not have a significant effect independently of use rank (χ2 = 2.33, p = 0.1302). When we consider burned and unburned plots separately, use rank is a highly significant predictor of burrow presence in both (GLM binomial logit, maximum-likelihood estimation, burned: n = 6429, χ2 = 14.22, p = 0.0026; unburned: –n = 1748, χ2 = 22.86, p < 0.0001). However, burned and unburned plots exhibit different responses to use rank (figure 2). Lizard densities peak under intermediate use in burned plots, with densities under high and low use not significantly different. However, there is no intermediate optimum for unburned plots: densities increase to a peak under highest use of a region. Differences between burned and unburned plots are significant only for the highest use rank. If V. gouldii are indeed at higher density in more diverse habitat, we should expect to see more burrows at the edges of patches, where lizards have easy access both to mature and early or mid-successional habitat. In the transect survey, there were 1748 late successional plots: 294 of these were within 20 m of an earlier burn. Controlling for the covarying effects of use rank, edge plots in late successional habitat were 1.51 times more likely to have burrows present compared with non-edge late successional plots (likelihood ratio test, χ2 = 4.74, p = 0.0294). Use rank significantly predicts measures of fire patch size and landscape diversity (see electronic supplementary material, figure S2).
Figure 2.
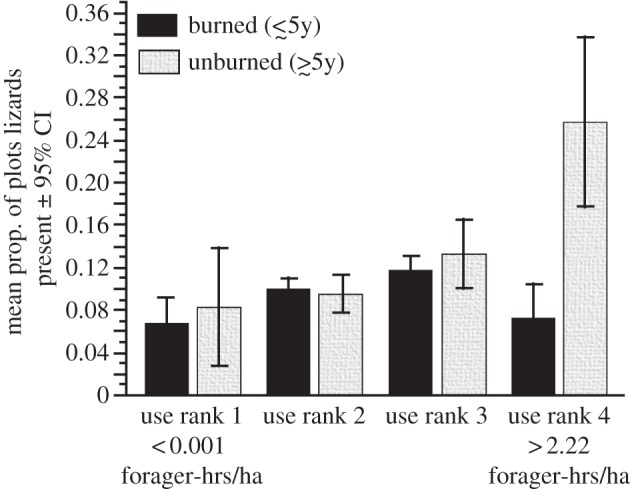
Sand monitor density increases with use rank in unburned patches, but reaches an intermediate peak in burned patches. Varanus gouldii density is measured as the mean (±95% CI) proportion of sample plots in both previously burned (within 5 years) and unburned (greater than 5 years since fire) patches with at least one burrow present. Density is plotted against use rank (ranked by forager-hours per square kilometre). There were a total of 368 plots located in regions assigned to use rank 4, 2921 in use rank 3 regions, 4282 in use rank 2 regions and 607 in use rank 1 regions.
(b). Effects on Martu hunting returns
Landscape-level effects on lizard density have measurable impacts on Martu hunting return rates. Lizard hunting returns are poorer and more uncertain in landscapes unmodified by human activity. In comparison to use rank 1, a lizard hunter's odds of harvest failure are 6.04p<0.0001, 6.23p=0.0001 and 5.42p=0.0003 times lower in use ranks 2, 3 and 4, respectively (logistic regression, controlling for sex, season, cooperation and precipitation, model χ2 = 54.63, p < 0.0001). Harvest sizes (per forager-day) in regions experiencing the lowest human predation intensity are halved compared with intermediate use ranks (table 1). This effect is driven by the rate at which foragers encounter successional patch diversity while hunting: an increase in diversity from 1 to 2 successional stages encountered per hour raises net foraging returns from 541 ± 827 to 1256 ± 675 kcals h−1 (least-square (LS) mixed model (forager ID as random effect), model r2 = 0.279, β = 727, model F ratio = 7.55, p = 0.0104, n = 31). The impetus to burn is owing to the impact of fire on foraging returns at the patch scale. Foragers gain their highest returns in the winter, when pursuits involve mainly burrow-spotting, by burning late successional vegetation: fire increases net foraging returns from 25 kcal h−1 to 1552 (table 2). In the summer, when lizards are pursued by tracking, access to ground burned earlier in the season is critical: returns are lower with more plant cover as tracks become more difficult to see.
Table 1.
Least-squares mean net return rates for sand monitor hunting (s.e., 95% CIs) derived from LS mixed model with forager ID as random effect, controlling for rainfall, season and cooperation, n = 750 bouts; use rank effect test F-ratio = 4.16, p = 0.0062. Values connected by the same letter are not significantly different (Student's post-hoc t-test, p < 0.05).
| use rank | LS mean (kcal d–1) | s.e. | 95% CI |
|---|---|---|---|
| 4 | 1344bc | 144 | 1059–1629 |
| 3 | 1654a | 135 | 1385–1923 |
| 2 | 1632ab | 179 | 1279–1985 |
| 1 | 850c | 288 | 283–1416 |
Table 2.
LS mean net return rates by habitat type and season. LS means derived from LS regression mixed nested model (forager random effect) n = 170 patches, model r2 = 0.217, PursuitType (nested within successional stage) effect test, F-ratio = 2.40, p = 0.0706, successional stage effect test F-ratio = 5.70, p = 0.0010. Values connected by the same letter are not significantly different (post-hoc Student's t-test, p < 0.05).
| type of pursuit | successional stage | LS mean (kcal h–1) | s.e. | 95% CI |
|---|---|---|---|---|
| tracking summer |
early | 1838a | 446 | 930–2746 |
| mid | 248b | 555 | −870 to 1368 | |
| late | 76b | 711 | −1339 to 1492 | |
| burrow-spotting winter |
early | 13b | 616 | −1224 to 1252 |
| mid | 403b | 380 | −551 to 1358 | |
| late | 25b | 407 | −929 to 981 | |
| burn late | 1552a | 326 | 634–2469 |
4. Discussion
Martu patch mosaic burning while hunting V. gouldii and other prey creates habitat patchiness that provides net positive benefits to lizard populations, feeding back to increase human hunting returns in regions of intermediate hunting intensity. Plots in unburned patches show an increase in V. gouldii density with use, whereas plots in burned patches show an intermediate optima. The difference between burned and unburned plots under heaviest human use reflects the extent to which hunting reduces local populations; the lack of a significant difference between burned plots under heavy use and burned plots under no human use suggest that the negative effects of hunting at the patch scale are balanced by the positive effects of human ecosystem engineering at the landscape scale. That this effect is owing to the rescaling of patch size through patch mosaic burning is indicated by the significant effect of edge contrasts on burrow presence. Burrows are more concentrated in patches of mature vegetation within a short distance of a previous burn. However, Martu patch mosaic burning is not designed to create more ideal habitat for V. gouldii per se: hunters burn primarily in the winter months when search and pursuit involves burrow-spotting because their net foraging returns increase more than 62 times. Sand monitor probably benefit because burning creates more diverse plant and animal communities across smaller scales on the landscape, increasing V. gouldii access to high-ranked prey [32,33].
Here, we have shown that at least one species is more abundant in anthropogenically modified environments, but there is at least circumstantial evidence to support the hypothesis that human ecosystem engineering has evolutionary consequences for a broader range of species. The fact that native mammal decline and extinction corresponded with an increase in invasive mesopredator populations and the departure of Martu and other linguistic groups from the desert between the 1930s and 1970s [25] suggests an even stronger argument for the niche construction aspects of Martu patch mosaic burning. The ecosystem engineering effects of burning that positively impact V. gouldii may also have supported a wide range of small mammals that also benefit from smaller fire mosaics. Rufous hare-wallabies (Lagorchestes hirsutus, now highly endangered) are browsers that feed on plants in many different successional stages and require mature spinifex hummocks for nesting and predator protection. Prior to the 1950s, they were abundant and hunted frequently by Martu throughout the spinifex sandplains, but their continued persistence may be dependent on patch mosaic burning to maintain access to early successional habitat adjacent to mature spinifex which provides refuge from predators [34–36]. Likewise, the brushtail possum (T. vulpecula) seems to have been able to persist in regions only where its habitat, riparian eucalyptus woodlands, is still protected from devastating fire through Aboriginal patch mosaic burning [37]. It was not human hunting of these small mammals that led to their extinction but the loss of human hunting. The loss of human hunting and burning may have led to an increase in predation (especially from invasive cats and foxes) and a reduction in the food supply that caused many of these animals adapted to smaller scale fire mosaics to go extinct or abandon the desert for more favourable regions.
The loss of Martu ecosystem engineering has had a negative impact on people as well. Facing an environment in which prey density is much lower than that in the past, foragers have adopted vehicles to reduce the costs of travelling between patches, and rely in part on imported foods. However, in doing so, Martu face new trade-offs. Using vehicles means that most hunting and burning is constrained to regions near tracks and roads, limiting the spatial extent that Martu can affect through fire, and creating vast landscapes interior to tracks where lightening fires burn unimpeded. The increased reliance on imported foods, which are expensive and of low quality has had damaging effects on indigenous health and well-being [38]. The logic of the Dreaming—that a healthy people and a healthy country are linked through fire and the use of resources—reflects a sophisticated ecological and evolutionary understanding of the ways that human activity functions as a keystone process in this desert climate. An environment that can support both healthy people and a wide range of now critically endangered species may now be linked to the continued presence and traditional hunting activities of Aboriginal people. Such effects are likely to be found in other environments in which indigenous patch mosaic burning and hunting provides a significant engineering effect.
Finding such effects requires recognizing the complexity of human–environment interactions: that there are both positive and negative feedbacks to humans and to other species, which are felt both directly, and indirectly via their effects on some other organism (e.g. removing a competitor), and which are characterized by spatially varying degrees of intensity. This complexity is further enhanced by heterogeneity in species responses to human predation and landscape modification: some species, like the sand monitor lizard described here, with fast life histories and a generalized diet, may respond positively to anthropogenic landscapes; others, such as more specialized, slowly reproducing species, may not.
The extent to which humans do end up embedded within ecosystems as a keystone species is likely to be a function of a long history of coevolutionary interaction between cultural practice and particular ecological communities, where ecoevolutionary feedbacks to human decision-making processes are the main force of cultural evolutionary change. This would be more likely where cultural evolutionary change is not complicated by the flow of practices and technologies between individuals living in widely varying environments, where individuals cannot simply move away to a new region or new environment when things get bad locally, and where populations can make it through the initial instability caused by practices that are not sustainable. There are certainly many examples of populations moving into new environments (for example, small oceanic islands) and importing practices that cause ecosystems to collapse, with subsequent negative effects on the human populations themselves. However, given enough time and given that human effects are spatially variable, we should see a cultural practice which is highly fine-tuned to local environments, where activities that produce net negative feedbacks to humans result in a decline in population, whereas those that produce net positive feedbacks increase populations. Beliefs such as the Jukurr are the institutionalization of such histories of ecological practice.
Acknowledgements
The authors are grateful to the Martu communities of Parnngurr and Punmu for hosting, supporting and carrying out this research project, to Christopher Parker and Brooke Scelza for assisting in data collection, and to Eric Pianka, Lisa Curran, Neil Burrows, Don Hankins, and two anonymous reviewers for comments on earlier versions of this paper.
Data accessibility
All data referenced here are in the possession of the primary author and can be obtained upon request.
Funding statement
The research we report here was supported by grants from the National Science Foundation, the Leakey Foundation and the Woods Institute for the Environment, Stanford University.
References
- 1.Stachowicz JJ. 2001. Mutualism, facilitation, and the structure of ecological communities. BioScience 51, 235–246 (doi:10.1641/0006-3568(2001)051[0235:MFATSO]2.0.CO;2) [Google Scholar]
- 2.Kikvidze Z, Callaway RM. 2009. Ecological facilitation may drive major evolutionary transitions. BioScience 59, 399–404 (doi:10.1525/bio.2009.59.5.7) [Google Scholar]
- 3.Bodmer RE, Eisenberg JF, Redford KH. 1997. Hunting and the likelihood of extinction of Amazonian mammals. Conserv. Biol. 11, 460–466 (doi:10.1046/j.1523-1739.1997.96022.x) [Google Scholar]
- 4.McDaniel CN, Borton DN. 2002. Increased human energy use causes biological diversity loss and undermines prospects for sustainability. BioScience 52, 929–936 (doi:10.1641/0006-3568(2002)052[0929:IHEUCB]2.0.CO;2) [Google Scholar]
- 5.Jones CG, Lawton JH. 1994. Organisms as ecosystem engineers. Oikos 69, 373–386 (doi:10.2307/3545850) [Google Scholar]
- 6.Laland KN, Odling-Smee J. 2010. How culture shaped the human genome: bringing genetics and the human sciences together. Nat. Rev. Genetics 11, 137–148 (doi:10.1038/nrg2734) [DOI] [PubMed] [Google Scholar]
- 7.Smith BD. 2011. General patterns of niche construction and the management of ‘wild’ plant and animal resources by small-scale pre-industrial societies. Phil. Trans. R. Soc. B 366, 836–848 (doi:10.1098/rstb.2010.0253) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rowley-Conwy P. 2011. Foraging and farming as niche construction: stable and unstable adaptations. Phil. Trans. R. Soc. B 366, 849–862 (doi:10.1098/rstb.2010.0307) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Roff DA. 1974. Spatial heterogeneity and the persistence of populations. Oecologia 15, 245–258 (doi:10.1007/BF00345181) [DOI] [PubMed] [Google Scholar]
- 10.Holt RD. 1984. Spatial heterogeneity, indirect interactions, and the coexistence of prey species. Am. Nat. 377–406 (doi:10.1086/284280) [DOI] [PubMed] [Google Scholar]
- 11.Roxburgh SH, Shea K, Wilson JB. 2004. The intermediate disturbance hypothesis: patch dynamics and mechanisms of species coexistence. Ecology 85, 359–371 (doi:10.1890/03-0266) [Google Scholar]
- 12.Futuyma D. 1988. The evolution of ecological specialization. Annu. Rev. Ecol. Syst. 19, 207–233 (doi:10.1146/annurev.ecolsys.19.1.207) [Google Scholar]
- 13.Marvier M, Kareiva P, Neubert MG. 2004. Habitat destruction, fragmentation, and disturbance promote invasion by habitat generalists in a multispecies metapopulation. Risk Anal. 24, 869–878 (doi:10.1111/j.0272-4332.2004.00485.x) [DOI] [PubMed] [Google Scholar]
- 14.Wiggins N, McArthur C. 2006. Diet switching in a generalist mammalian folivore: fundamental to maximising intake. Oecologia 147, 650–657 (doi:10.1007/s00442-005-0305-z) [DOI] [PubMed] [Google Scholar]
- 15.Redford KH, Robinson JG. 1987. The game of choice: patterns of Indian and colonist hunting in the neotropics. Am. Anthropol. 89, 650–667 (doi:10.1525/aa.1987.89.3.02a00070) [Google Scholar]
- 16.Gross T, Rudolf L, Levin SA. 2009. Generalized models reveal stabilizing factors in food webs. Science 325, 747–750 (doi:10.1126/science.1173536) [DOI] [PubMed] [Google Scholar]
- 17.Odling-Smee J, Erwin DH, Palkovacs EP, Feldman MW, Laland KN. 2013. Niche construction theory: a practical guide for ecologists. Q. Rev. Biol. 88, 3–28 (doi:10.1086/669266) [DOI] [PubMed] [Google Scholar]
- 18.Estes JA, Palmisano JF. 1974. Sea otters: their role in structuring nearshore communities. Science 185, 1058–1060 (doi:10.1126/science.185.4156.1058) [DOI] [PubMed] [Google Scholar]
- 19.Hughes BB, Eby R, Dyke EV, Tinker MT, Marks CI, Johnson KS. 2013. Recovery of a top predator mediates negative eutrophic effects on seagrass. Proc. Natl Acad. Sci. USA 110, 15 313–15 318 (doi:10.1073/pnas.1302805110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.O'Connell JF. 2004. Dating the colonization of Sahul (Pleistocene Australia–New Guinea): a review of recent research. J. Archaeol. Sci. 31, 835–853 (doi:10.1016/j.jas.2003.11.005) [Google Scholar]
- 21.Smith M, Williams A, Turney C. 2008. Human–environment interactions in Australian drylands: exploratory time-series analysis of archaeological records. Holocene 18, 389 (doi:10.1177/0959683607087929) [Google Scholar]
- 22.Gould RA. 1991. Arid-land foraging as seen from Australia: adaptive models and behavioral realities. Oceania 62, 12–33 [Google Scholar]
- 23.O'Connell JF, Latz PK. 1983. Traditional and modern plant use among the Alyawara of Central Australia. Econ. Bot. 37, 80–109 (doi:10.1007/BF02859310) [Google Scholar]
- 24.Cane S. 1987. Australian aboriginal subsistence in the western desert. Hum. Ecol. 15, 391–434 (doi:10.1007/BF00887998) [Google Scholar]
- 25.Burbidge AA, Johnson KA, Fuller PJ, Southgate RI. 1988. Aboriginal knowledge of the mammals of the central deserts of Australia. Wildl. Res. 15, 9–39 (doi:10.1071/WR9880009) [Google Scholar]
- 26.Burrows ND, Burbidge AA, Fuller PJ. 2006. Evidence of altered fire regimes in the Western Desert region of Australia. Conserv. Sci. West Aust. 5, 272–284 [Google Scholar]
- 27.Bliege Bird R, Bird DW, Codding BF, Parker CH, Jones JH. 2008. The ‘fire stick farming’ hypothesis: Australian Aboriginal foraging strategies, biodiversity, and anthropogenic fire mosaics. Proc. Natl Acad. Sci. USA 105, 14 796–14 801 (doi:10.1073/pnas.0804757105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bliege Bird R, Codding B, Kauhanen P. 2012. Aboriginal hunting buffers climate-driven fire-size variability in Australia's spinifex grasslands. Proc. Natl Acad. Sci. USA 109, 10 287–10 292 (doi:10.1073/pnas.1204585109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rolls M. 2005. Black is not green. Austral. Stud. 18, 41–65 [Google Scholar]
- 30.Pianka ER. 1994. Comparative ecology of Varanus in the Great Victoria desert. Austral J. Ecol. 19, 395–408 (doi:10.1111/j.1442-9993.1994.tb00505.x) [Google Scholar]
- 31.Bird DW, Bliege Bird R, Codding BF. 2009. In pursuit of mobile prey: Martu hunting strategies and archaeofaunal interpretation. Am. Antiquity 74, 3–29 [Google Scholar]
- 32.Letnic M, Dickman CR, Tischler MK, Tamayo B, Beh CL. 2004. The responses of small mammals and lizards to post-fire succession and rainfall in arid Australia. J. Arid Environ. 59, 85–114 (doi:10.1016/j.jaridenv.2004.01.014) [Google Scholar]
- 33.Pianka ER, Goodyear SE. 2012. Lizard responses to wildfire in arid interior Australia: long-term experimental data and commonalities with other studies. Austral Ecol. 37, 1–11 (doi:10.1111/j.1442-9993.2010.02234.x) [Google Scholar]
- 34.Lundie-Jenkins G. 1993. Ecology of the rufous hare-wallaby, Lagorchestes hirsutus Gould (Marsupialia: Macropodidae) in the Tanami Desert, Northern Territory. I. Patterns of habitat use. Wildlife Res. 20, 457–475 (doi:10.1071/WR9930457) [Google Scholar]
- 35.Lundie-Jenkins G, Corbett L. 1993. Ecology of the rufous hare-wallaby, Lagorchestes hirsutus Gould (Marsupialia: Macropodidae) in the Tanami Desert, Northern Territory. III. Interactions with introduced mammal species. Wildlife Res. 20, 495–511 (doi:10.1071/WR9930495) [Google Scholar]
- 36.Lundie-Jenkins G, Phillips C. 1993. Ecology of the rufous hare-wallaby, Lagorchestes hirsutus Gould (Marsupialia: Macropodidae) in the Tanami Desert, Northern Territory. II. Diet and feeding strategy. Wildlife Res. 20, 477–493 (doi:10.1071/WR9930477) [Google Scholar]
- 37.Kerle J, Foulkes J, Kimber R. 1992. The decline of the Brushtail Possum, Trichosurus vulpecula (Kerr 1798), in arid Australia. Rangel. J. 14, 107–127 (doi:10.1071/RJ9920107) [Google Scholar]
- 38.Garnett ST, Sithole B, Whitehead PJ, Burgess CP, Johnston FH. 2009. Healthy country, healthy people: policy implications of links between indigenous human health and environmental condition in tropical Australia. Austral. J. Public Admin. 68, 53–66 (doi:10.1111/j.1467-8500.2008.00609.x) [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data referenced here are in the possession of the primary author and can be obtained upon request.