

Abstract
Activated polymorphonuclear neutrophils play an important role in the pathogenesis of vaso-occlusive painful sickle cell crisis. Upon activation, polymorphonuclear neutrophils can form neutrophil extracellular traps. Neutrophil extracellular traps consist of a meshwork of extracellular DNA, nucleosomes, histones and neutrophil proteases. Neutrophil extracellular traps have been demonstrated to be toxic to endothelial and parenchymal cells. This prospective cohort study was conducted to determine neutrophil extracellular trap formation in sickle cell patients during steady state and painful crisis. As a measure of neutrophil extracellular traps, plasma nucleosomes levels were determined and polymorphonuclear neutrophil activation was assessed measuring plasma levels of elastase-α1-antitrypsin complexes in 74 patients in steady state, 70 patients during painful crisis, and 24 race-matched controls using Enzyme Linked Immunosorbent Assay. Nucleosome levels in steady state sickle cell patients were significantly higher than levels in controls. During painful crisis levels of both nucleosomes and elastase-α1-antitrypsin complexes increased significantly. Levels of nucleosomes correlated significantly to elastase-α1-antitrypsin complex levels during painful crisis, (Sr = 0.654, P<0.001). This was seen in both HbSS/HbSβ0-thalassemia (Sr=0.55, P<0.001) and HbSC/HbSβ+-thalassemia patients (Sr=0.90, P<0.001) during painful crisis. Levels of nucleosomes showed a correlation with length of hospital stay and were highest in patients with acute chest syndrome. These data support the concept that neutrophil extracellular trap formation and neutrophil activation may play a role in the pathogenesis of painful sickle cell crisis and acute chest syndrome.
Introduction
Sickle cell disease (SCD) is characterized by recurrent acute painful vaso-occlusive crisis (VOC), accounting for the vast majority of SCD-related hospital admissions.1–3 VOC-related complications, such as acute chest syndrome, stroke and multi-organ failure are associated with high morbidity and mortality.4 The exact pathogenesis of acute painful VOC remains to be clarified. Alongside the crucial role for sickle erythrocytes in this, it encompasses an inflammatory response as evidenced by endothelial activation, coagulation activation and enhanced cellular adhesion, finally all contributing to microvascular occlusion.
Leukocytes play an important role in the development of microvascular obstruction and sickle cell disease-related complications. In steady state sickle cell patients, leukocytosis is associated with severity of disease.5 Clinical studies show that leukocytosis is a risk factor for major sickle cell-related complications such as stroke,6 acute chest syndrome7 and early death.8 Additionally, the clinical benefit of hydroxycarbamide in sickle cell patients has partly been attributed to a reduction in polymorphonuclear neutrophil (PMN) cell count9 and reduced PMN adhesion.10In vitro studies have demonstrated that PMN isolated from sickle cell patients are primed as evidenced by an increased expression of adhesion molecules,11–13 rendering them more susceptible to inflammatory stimuli as compared to PMN from healthy controls.14 Moreover, activation of PMN, e.g. upon interaction with red blood cells,15 leads to the production of toxic reactive oxygen species (ROS), contributing to oxidative stress.16In vitro studies as well as in vivo studies in SCD mice models demonstrate P- and E-selectin interactions with integrins17–19 to be crucial for the adherence of leukocytes to endothelial and sickle red blood cells, contributing to the complex process of vaso-occlusion.20,21 This identifies PMN activation and adhesion as important processes in the pathogenesis of vaso-occlusion in SCD.
Recently, activated PMN have been demonstrated to form neutrophil extracellular traps (NET).22 During NET formation, DNA and DNA-binding proteins are extruded from the neutrophils exposing a mesh consisting of nucleosomes, histones and neutrophil proteases such as elastase. These NET are regarded to be part of the innate immune response system.23 However, their function is considered to be a double-edged sword. On one hand, NET formation is an efficient strategy to kill invading micro-organisms, like bacteria and fungi. On the other hand, NET can become harmful for the host since its exposed compounds (e.g. the mesh of DNA, histones and neutrophil proteases) are toxic to endothelial cells and parenchymal tissue.24–26 NET formation has been reported to be pro-coagulant in inflammatory models and is thought to contribute to the development of disseminated intravascular coagulation, and hence to morbidity and mortality in sepsis.27–29 Circulating nucleosomes and markers of neutrophil activation have been reported to be suitable markers for NET formation in plasma in baboons and humans.29–31 Nucleosomes consist of a core octamer of two copies each holding the histones H2A, H2B, H3 and H4, around which a segment of helical DNA of 146 base pairs is wrapped.32 Nucleosomes can be actively released into the circulation from dead cells as a result of the activity of factor-VII activating protease (FSAP).33 Circulating cell-free DNA in the form of nucleosomes has been reported to correlate with organ dysfunction, disease severity and mortality in sepsis patients and children suffering from meningococcal sepsis.34–36
So far, no data are available on NET formation in sickle cell patients. Since white blood cell counts have been shown to correlate with morbidity of sickle cell patients, and since PMN activation seems to play an important role in the development of sickle cell painful vaso-occlusive crisis, we hypothesized that NET formation may be involved in these processes. The aim of this prospective cohort study, therefore, was to measure plasma levels of circulating nucleosomes and PMN activation as evidenced by human neutrophil elastase-α1-antitrypsin (EA) complexes as a measure of NET formation in plasma in sickle cell patients both during steady state and painful VOC, and to evaluate their correlation with crisis severity.
Methods
Patients
This study followed a prospective design in which patients with sickle cell anemia (HbSS) and compound heterozygous states HbSβ0-thalassemia, HbSβ+-thalassemia and sickle-hemoglobin C (HbSC) patients were eligible for inclusion. Diagnosis of hemoglobinopathy was confirmed by means of high performance liquid chromatography in combination with measurement of erythrocyte mean corpuscular volume. Consecutive sickle cell patients, 18 years of age or over, attending the outpatient clinic (steady state) or being admitted for a painful crisis to the Academic Medical Center or the Slotervaart Hospital, Amsterdam, The Netherlands, were invited to participate. A painful crisis was defined as musculo-skeletal pain not otherwise explained, recognized as such by the patient and requiring medical treatment. Samples during painful crisis were obtained within the first 24 h of admission. Patients with painful crisis within four weeks and/or blood transfusion within three months prior to the evaluation for the present study were excluded from inclusion. Other exclusion criteria were: pregnancy, inflammatory autoimmune disease or any acute infection within three months prior to study participation. Information concerning complications during admission was collected from medical records. An acute chest syndrome was defined as a new infiltrate (on admission or during hospitalization) on chest X-ray associated with one or more new symptoms of chest pain, fever, tachypnea, wheezing, cough or hypoxemia.37 Samples from race-matched volunteers were taken for control reference measurements. Written informed consent was obtained from all participants before any study procedure was performed. The study protocol was approved by the local medical ethical committee and conducted in agreement with the 1975 Declaration of Helsinki as revised in 2008.
Blood sample collection and laboratory analysis
Blood samples were taken by venipuncture. Blood vials were centrifuged at 4°C for 15 min at 3000 rpm, and serum and plasma was stored in small aliquots at −80°C until further analysis. Hematology parameters, nucleosome and EA levels were measured in EDTA-anticoagulated plasma. Soluble vascular adhesion molecule1 (sVCAM1) levels were determined in serum (R&D Systems; Minneapolis, USA). Lactate hydrogenase (LDH) and bilirubin levels were measured with spectrophotometry in heparinized plasma (P800 Modular, Roche, Switzerland). Plasma levels of the long pentraxin-3 (PTX3) were determined using sandwich ELISA.38 Antigen levels of von Willebrand factor (vWFag) were assessed by ELISA using antibodies from Dako (Glostrup, Denmark). Nucleosome levels were measured using ELISA as described previously.35,39 Neutrophil activation in form of elastase-α1-antitrypsin (EA) complexes was measured by an ELISA as previously described.30,40
Statistical analysis
For statistical analysis, patients were divided primarily into two groups: patients with the relatively severe genotypes HbSS and HbSβ0-thalassemia were grouped together (HbSS/HbSβ0-thal) and patients with the relatively milder HbSC and HbSβ+-thalassemia genotypes were put in the other group (HbSC/HbSβ+-thal).41,42 We used a commercial statistical package (IBM SPSS Statistics 19, SPSS Inc., Hong Kong, PRC) for data analysis. Since results were not normally distributed, they are expressed as median with interquartile range. Unless stated otherwise, P<0.05 was considered statistically significant. Bonferroni’s correction was applied for multiple testing (P<0.004 was considered statistically significant).
Results
Patients
Seventy-four patients in steady state (49 HbSS/HbSβ0-thal and 25 HbSC/HbSβ+-thal), 70 patients during painful crisis (53 HbSS/HbSβ0-thal and 17 HbSC/HbSβ+-thal), and 24 healthy race-matched controls (HbAA) were included in the study. Patients’ characteristics are summarized in Table 1. Twenty-four percent of sickle cell patients during steady state and a similar percentage (23%) of patients during painful crisis were on hydroxycarbamide treatment. Of the patients included with painful crisis, one (HbSS/HbSβ0-thal) patient was admitted with acute chest syndrome (ACS) and 5 (4 HbSS/HbSβ0-thal and one HbSC/HbSβ+-thal) patients developed an ACS 48–60 h after admission. No infection was reported for any of the patients with an acute chest syndrome.
Table 1.
Baseline characteristics.
Nucleosomes and neutrophil activation
Plasma levels of nucleosomes were significantly higher during painful crisis (20.0 U/mL; IQR 7.9–107.3) as compared to those in steady state (6.4 U/mL; 3.5–9.7, P<0.001) (Figure 1A). This was seen in both HbSS/HbSβ0-thal (20.2 U/ml; 8.9–129.0 vs. 6.0 U/mL; 3.0–9.8, P<0.001) and HbSC/HbSβ+-thal (11.7 U/mL; 5.1–67.7 vs. 7.1 U/mL; 4.6–9.6, P=0.045) patients (Figure 1B). Plasma levels of nucleosomes in healthy controls were just above the detection limit of the assay (5.0 U/ml; 3.0–6.5). In steady state sickle cell patients, plasma levels of nucleosomes were significantly higher than levels in healthy controls (P=0.031). In the analysis for the two genotype groups separately, the same was seen for HbSC/HbSβ+-thal patients in steady state (P=0.020) while plasma nucleosome levels in HbSS/HbSβ0-thal patients in steady state were comparable with those in healthy controls (P=0.089).
Figure 1.

Levels of nucleosomes (A and B) and elastase-α1-antitrypsin complexes (C and D) in healthy controls and in sickle cell patients in steady state and during painful crisis. Results are shown for all patients (A and C) and for the subgroups containing HbSS/HbSβ0-thalassemia and HbSC/HbSβ+-thalassemia patients (B and D). The number of patients in each group is indicated above the x-axis. Patients who developed an acute chest syndrome during admission were among those with the highest nucleosome and EA levels. The corresponding levels are indicated in black. Results are indicated as median with interquartile range. Comparison of the groups was performed by means of Mann Whitney rank sum test. P<0.05 was considered statistically significant.
Plasma levels of EA were significantly higher during painful crisis (73.6 ng/mL; 54.9–100.8) as compared to those in steady state (46.2 ng/mL; 34.3–65.6, P<0.001) (Figure 1C). This was seen in HbSS/HbSβ0-thal patients (75.1 ng/mL; 56.5–102.4 vs. 45.7 ng/mL; 34.7–59.7, P<0.001), while in HbSC/HbSβ+-thal patients, the increment did not reach statistical significance (62.0 ng/mL; 48.0–96.7 vs. 50.2 ng/ml; 33.3–67.7, P=0.051) (Figure 1D). Plasma levels of EA in healthy controls (39.9 ng/mL; 31.5–62.2) were comparable to those in steady state sickle cell patients (P=0.330).
During painful crisis, levels of nucleosomes correlated significantly with levels of EA (Sr = 0.654, P<0.001). This was seen in both HbSS/HbSβ0-thal (Sr = 0.55, P<0.001) as well as in HbSC/HbSβ+-thal patients (Sr = 0.90, P<0.001). During steady state, the correlation between levels of nucleosomes and EA was significant but weak (Sr = 0.236, P=0.043). The correlation in HbSC/HbSβ+-thal patients in steady state remained significant (Sr = 0.63, P=0.001), while no correlation was found between levels of nucleosomes and EA in HbSS/HbSβ0-thal patients in steady state (Sr = 0.043, P=0.77).
There was no difference in levels of nucleosomes and EA between patients with and patients without documented infection during painful crisis (data not shown). The use of hydroxycarbamide had no effect on levels of nucleosomes or EA in sickle cell patients whether in steady state or during painful crisis (data not shown).
In a paired analysis of 25 patients (total 36 painful crises), significant increments were observed during painful crisis in plasma levels of both nucleosomes (from 5.0 U/ml; 3.0–10.8 to 20.2 U/ml; 6.8–94.3, P<0.001) and EA (47.9 ng/ml; 36.0–67.6 to 70.6 ng/ml; 55.9–101.4, P<0.001) as compared to those in steady state (Figure 2A and B).
Figure 2.
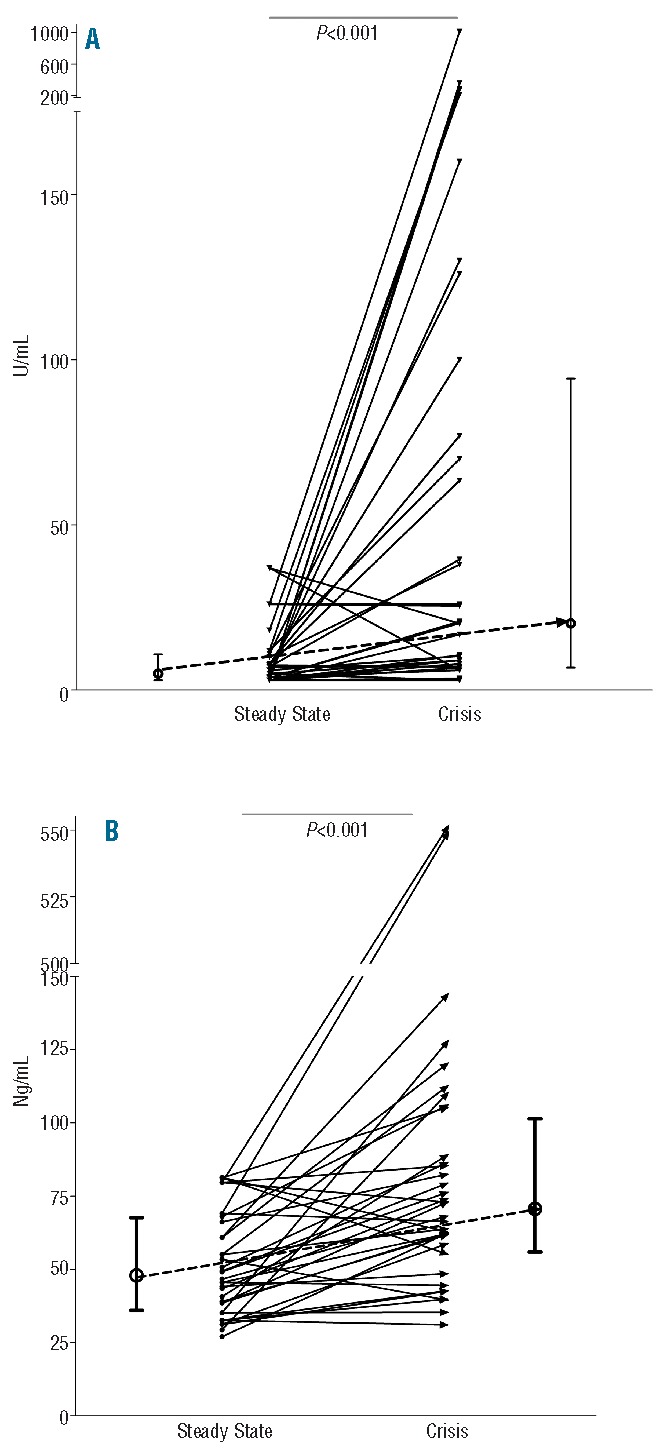
Paired analysis of levels of nucleosomes (A) and elastase-α1-antitrypsin complexes (B) in 36 painful crisis in 25 patients included both in steady state and in painful crisis. For comparison between related samples the Wilcoxon’s signed rank test was used. P<0.05 was considered statistically significant. Results are indicated as median with interquartile range.
Nucleosomes and EA in association with markers of endothelial activation, hemolysis and inflammation
While nucleosome levels in steady state HbSS/HbSβ0-thal patients correlated significantly with vWF:Ag (Sr = 0.452, P=0.001) and sVCAM-1 (Sr = 0.421, P=0.003) they only correlated significantly with PTX3 (Sr = 0.623, P=0.001) during painful crisis. In the same patient group during painful crisis, EA levels just failed to reach a statistical significant correlation with PTX3 levels (Sr = 0.529, P=0.008).
Leukocyte counts did not correlate with levels of nucleosomes or EA. In addition, neutrophil count did not correlate with levels of nucleosomes or levels of EA, neither when results of patients were pooled, nor when they were evaluated separately in the different subgroups. No association was found between markers of hemolysis (hemoglobin, LDH and bilirubin) and levels of nucleosomes or EA.
Association with acute chest syndrome and duration of hospitalization
Acute chest syndrome: the 6 patients who developed an ACS were among those with the highest nucleosome (359, 274.8, 190, 130, 128 and 100 U/mL, respectively) and EA levels (549.9, 120.8, 91.8, 86.7, 75.1, and 63.9 ng/mL, respectively) (Figures 1A–D). In these 6 sickle cell patients with ACS, nucleosome levels were significantly higher than those in patients during painful crisis without ACS (n= 64; 160.0 U/mL; 121.0–295.9 vs. 20.07 U/mL; 7.9–107.3, P=0.002).
Hospitalization duration: nucleosome levels, but not EA levels, correlated significantly with duration of hospital stay in all sickle cell patients during painful crisis (Sr = 0.441, P<0.001). Excluding the patients with acute chest syndrome, the correlation remained statistically significant (Sr = 0.385, P=0.002). When analyzing the correlation for HbSS/HbSβ0-thal patients, the correlation between nucleosome levels and duration of hospital stay was stronger (Sr = 0.530, P<0.001). Figure 3 shows the association between levels of nucleosomes and duration of hospitalization.
Figure 3.
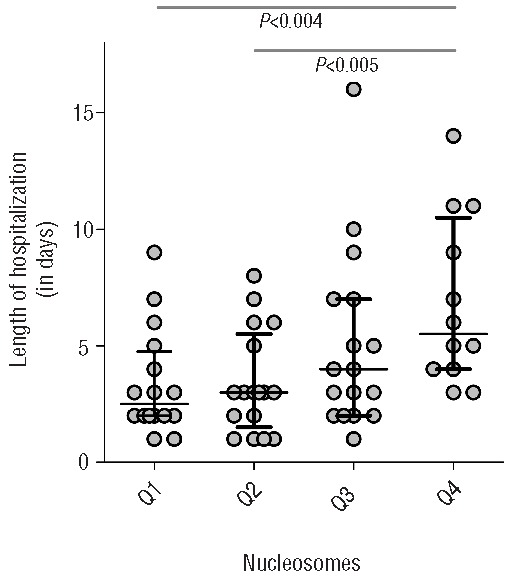
Association between levels of nucleosomes in all SCD patients and length of hospital stay (LOS). Median stay is 3 days (IQR 2–7 days). Display of LOS when nucleosome level is divided in quartiles. An increasing LOS can be seen when levels of nucleosomes increase.
Discussion
In the present study, we demonstrate that during painful vaso-occlusive crisis sickle cell patients have significantly higher levels of circulating nucleosomes and neutrophil activation, as shown by increased EA complexes, as compared to sickle cell patients in steady state. Results of the paired analyses support the findings of the between-group analyses. We show that patients developing the severe and potentially life-threatening complication of acute chest syndrome were among those with the highest nucleosome and EA levels. Moreover, we found that nucleosome levels correlate with duration of hospitalization. Nucleosome and EA levels correlate significantly with each other during painful crisis. Together, our data provide indirect evidence for NET formation in patients with sickle cell disease suffering from VOC.
Our results are in line with a previous study in sickle cell patients demonstrating significantly increased amounts of circulating cell-free DNA, as determined by quantitative PCR amplification, in sickle cell patients during painful crisis as compared to levels in steady state.43 Interestingly, in the current study, nucleosome levels in sickle cell patients with acute chest syndrome were comparable to levels measured in patients with severe sepsis using the same assay.34,35 In these patients with sepsis, circulating cell-free DNA in form of nucleosomes correlated with morbidity and mortality.34,35
This study has several limitations that need to be taken into account when interpreting the data collected. Firstly, we have not performed sequential nucleosome and EA analysis during admission for painful crisis in our patients, limiting the findings to a single measurement at presentation with a painful crisis. The observation that nucleosome and EA levels taken at presentation were highest in patients developing an ACS during admission are nevertheless in line with previous findings suggesting neutrophils to be an important player in the pathogenesis of ACS.44,45 The observation that neutrophil count does not correlate with EA or nucleosome levels in this study in sickle cell patients is supported by observations from studies in patients with severe sepsis.40 While EA results reflect general PMN activation, it is likely that during vaso-occlusive crisis neutrophil count is a measure for circulating (‘countable’) neutrophils while it does not reflect neutrophils migrated to tissue or adherent to activated endothelial cells, the latter process being observed in vaso-occlusive crisis in mice models for sickle cell disease.20,21
Secondly, the correlation during painful crisis between nucleosome levels with EA levels and PTX3, both being localized in NET46,47 (while the latter has previously been demonstrated to be increased during sickle cell painful vaso-occlusive crisis)48 support the hypothesis of PMN as an important nucleosome source via NET formation, at least during vaso-occlusive complications. This is also in line with the publications reporting circulating nucleosomes with or without markers for neutrophil activation to be a good measure for NET formation in circulation.29,31 However, the ELISAs detecting nucleosomes are not specific for nucleosomes released by PMN, and we can not, therefore, exclude the possibility that nucleosomes released into the circulation by other cell types, such as endothelial and parenchymal cells, are detected as well. The statistically significant correlation between nucleosome levels and markers of endothelial activation, vWFag and sVCAM-1, in steady state HbSS/HbSβ0-thal patients might be indirect evidence that damaged endothelial cells contribute to the circulating nucleosomes. Whether this endothelial cell damage is a consequence of local PMN activation, e.g. in the form of NET formation being cytotoxic to endothelial cells, remains to be established.
Thirdly, results of the analyses performed on the specified genotype groups sometimes diverge from the analyses carried out when pooling data and considering the patient population as one group. This may be due to a limited number of patients in the respective subgroups causing a lack of power to show statistically significant findings. Moreover, the influence of the interaction of sickle cell erythrocytes with PMN on NET formation and the role of the genotype in this interaction has yet to be established.
In conclusion, we demonstrate for the first time elevated levels of circulating nucleosomes and neutrophil activation in sickle cell patients with painful crisis suggesting NET formation in these patients. NET, consisting of nucleosomes, proteases and histones, may promote endothelial activation and contribute to longer and more severe sickle cell crisis. The role of NET in the prediction of clinical complications in sickle cell disease painful crisis and as a potential therapeutic target deserves further study.
Acknowledgments
The authors would like to thank B. Bottazzi and A. Mantovani (Istituto Clinico Humanitas, Rozzano, Milan, Italy) for their laboratory work on pentraxin-3.
Footnotes
The Online version of this paper contains a Supplementary Appendix.
The CURAMA study group is a collaborative effort studying sickle cell disease in Curaçao and The Netherlands. Participating centers: The Red Cross Blood Bank Foundation, Curaçao, The Antillean Institute for Health Research, Curaçao, The Department of Internal Medicine, Slotervaart Hospital, Amsterdam, The Netherlands; the Department of Vascular Medicine and the Department of Hematology, Academic Medical Center, Amsterdam, The Netherlands; the Department of Hematology, Erasmus Medical Center, Rotterdam, The Netherlands; the Department of Laboratory Medicine, University Medical Center Groningen, The Netherlands; the Department of Internal Medicine, Laboratory of Clinical Thrombosis and Hemostasis, and the Cardiovascular Research Institute, Academic Hospital Maastricht, The Netherlands.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Serjeant GR, Ceulaer CD, Lethbridge R, Morris J, Singhal A, Thomas PW. The painful crisis of homozygous sickle cell disease: clinical features. Br J Haematol. 1994; 87(3):586–91 [DOI] [PubMed] [Google Scholar]
- 2.Neonato MG, Guilloud-Bataille M, Beauvais P, Begue P, Belloy M, Benkerrou M, et al. Acute clinical events in 299 homozygous sickle cell patients living in France. French Study Group on Sickle Cell Disease. Eur J Haematol. 2000;65(3):155–64 [DOI] [PubMed] [Google Scholar]
- 3.Platt OS, Thorington BD, Brambilla DJ, Milner PF, Rosse WF, Vichinsky E, et al. Pain in sickle cell disease. Rates and risk factors. N Engl J Med. 1991;325(1):11–6 [DOI] [PubMed] [Google Scholar]
- 4.Steinberg MH. In the clinic. Sickle cell disease. Ann Intern Med. 2011;155(5):ITC31-15 [DOI] [PubMed] [Google Scholar]
- 5.Anyaegbu CC, Okpala IE, Akren’Ova YA, Salimonu LS. Peripheral blood neutrophil count and candidacidal activity correlate with the clinical severity of sickle cell anaemia (SCA). Eur J Haematol. 1998; 60(4):267–8 [DOI] [PubMed] [Google Scholar]
- 6.Ohene-Frempong K, Weiner SJ, Sleeper LA, Miller ST, Embury S, Moohr JW, et al. Cerebrovascular accidents in sickle cell disease: rates and risk factors. Blood. 1998; 91(1):288–94 [PubMed] [Google Scholar]
- 7.Castro O, Brambilla DJ, Thorington B, Reindorf CA, Scott RB, Gillette P, et al. The acute chest syndrome in sickle cell disease: incidence and risk factors. The Cooperative Study of Sickle Cell Disease. Blood. 1994; 84(2):643–9 [PubMed] [Google Scholar]
- 8.Platt OS, Brambilla DJ, Rosse WF, Milner PF, Castro O, Steinberg MH, et al. Mortality in sickle cell disease. Life expectancy and risk factors for early death. N Engl J Med. 1994;330(23):1639–44 [DOI] [PubMed] [Google Scholar]
- 9.Charache S, Barton FB, Moore RD, Terrin ML, Steinberg MH, Dover G, et al. Hydroxyurea and sickle cell anemia. Clinical utility of a myelosuppressive “switching” agent. The Multicenter Study of Hydroxyurea in Sickle Cell Anemia. Medicine (Baltimore). 1996;75(6):300–26 [DOI] [PubMed] [Google Scholar]
- 10.Almeida CB, Scheiermann C, Jang JE, Prophete C, Costa FF, Conran N, et al. Hydroxyurea and a cGMP-amplifying agent have immediate benefits on acute vaso-occlusive events in sickle cell disease mice. Blood. 2012;120(14):2879–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Okpala I, Daniel Y, Haynes R, Odoemene D, Goldman J. Relationship between the clinical manifestations of sickle cell disease and the expression of adhesion molecules on white blood cells. Eur J Haematol. 2002;69 (3):135–44 [DOI] [PubMed] [Google Scholar]
- 12.Lard LR, Mul FP, de Haas M, Roos D, Duits AJ. Neutrophil activation in sickle cell disease. J Leukoc Biol. 1999;66(3):411–5 [DOI] [PubMed] [Google Scholar]
- 13.Fadlon E, Vordermeier S, Pearson TC, Mire-Sluis AR, Dumonde DC, Philips J, et al. Blood polymorphonuclear leukocytes from the majority of sickle cell patients in the crisis phase of the disease show enhanced adhesion to vascular endothelium and increased expression of CD64. Blood. 1998; 91(1):266–74 [PubMed] [Google Scholar]
- 14.Lum AF, Wun T, Staunton D, Simon SI. Inflammatory potential of neutrophils detected in sickle cell disease. Am J Hematol. 2004;76(2):126–33 [DOI] [PubMed] [Google Scholar]
- 15.Hofstra TC, Kalra VK, Meiselman HJ, Coates TD. Sickle erythrocytes adhere to polymorphonuclear neutrophils and activate the neutrophil respiratory burst. Blood. 1996;87(10):4440–7 [PubMed] [Google Scholar]
- 16.Nur E, Biemond BJ, Otten HM, Brandjes DP, Schnog JJ. Oxidative stress in sickle cell disease; pathophysiology and potential implications for disease management. Am J Hematol. 2011;86(6):484–9 [DOI] [PubMed] [Google Scholar]
- 17.Brittain JE, Knoll CM, Ataga KI, Orringer EP, Parise LV. Fibronectin bridges monocytes and reticulocytes via integrin alpha4beta1. Br J Haematol. 2008; 141(6):872–81 [DOI] [PubMed] [Google Scholar]
- 18.Hidalgo A, Chang J, Jang JE, Peired AJ, Chiang EY, Frenette PS, et al. Heterotypic interactions enabled by polarized neutrophil microdomains mediate thromboinflammatory injury. Nat Med. 2009; 15(4):384–91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chang J, Shi PA, Chiang EY, Frenette PS. Intravenous immunoglobulins reverse acute vaso-occlusive crises in sickle cell mice through rapid inhibition of neutrophil adhesion. Blood. 2008;111(2):915–23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Turhan A, Weiss LA, Mohandas N, Coller BS, Frenette PS. Primary role for adherent leukocytes in sickle cell vascular occlusion: a new paradigm. Proc Natl Acad Sci USA. 2002;99(5):3047–51 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Turhan A, Jenab P, Bruhns P, Ravetch JV, Coller BS, Frenette PS. Intravenous immune globulin prevents venular vaso-occlusion in sickle cell mice by inhibiting leukocyte adhesion and the interactions between sickle erythrocytes and adherent leukocytes. Blood. 2004;103(6):2397–400 [DOI] [PubMed] [Google Scholar]
- 22.Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532–5 [DOI] [PubMed] [Google Scholar]
- 23.Brinkmann V, Zychlinsky A. Beneficial suicide: why neutrophils die to make NETs. Nat Rev Microbiol. 2007;5(8):577–82 [DOI] [PubMed] [Google Scholar]
- 24.Gupta AK, Joshi MB, Philippova M, Erne P, Hasler P, Hahn S, et al. Activated endothelial cells induce neutrophil extracellular traps and are susceptible to NETosis-mediated cell death. FEBS Lett. 2010; 584(14):3193–7 [DOI] [PubMed] [Google Scholar]
- 25.Xu J, Zhang X, Pelayo R, Monestier M, Ammollo CT, Semeraro F, et al. Extracellular histones are major mediators of death in sepsis. Nat Med. 2009; 15(11):1318–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Saffarzadeh M, Juenemann C, Queisser MA, Lochnit G, Barreto G, Galuska SP, et al. Neutrophil extracellular traps directly induce epithelial and endothelial cell death: a predominant role of histones. PLoS One. 2012;7(2):e32366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Massberg S, Grahl L, von Bruehl ML, Manukyan D, Pfeiler S, Goosmann C, et al. Reciprocal coupling of coagulation and innate immunity via neutrophil serine proteases. Nat Med. 2010;16(8):887–96 [DOI] [PubMed] [Google Scholar]
- 28.von Bruhl ML, Stark K, Steinhart A, Chandraratne S, Konrad I, Lorenz M, et al. Monocytes, neutrophils, and platelets cooperate to initiate and propagate venous thrombosis in mice in vivo. J Exp Med. 2012;209(4):819–35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD, Jr, et al. Extracellular DNA traps promote thrombosis. Proc Natl Acad Sci USA. 2010;107 (36):15880–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.van Montfoort ML, Stephan F, Lauw MN, Hutten BA, Van Mierlo GJ, Solati S, et al. Circulating nucleosomes and neutrophil activation as risk factors for deep vein thrombosis. Arterioscler Thromb Vasc Biol. 2013;33(1):147–51 [DOI] [PubMed] [Google Scholar]
- 31.Fuchs TA, Kremer Hovinga JA, Schatzberg D, Wagner DD, Lammle B. Circulating DNA and myeloperoxidase indicate disease activity in patients with thrombotic microangiopathies. Blood. 2012; 120(6):1157–64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Amoura Z, Piette JC, Bach JF, Koutouzov S. The key role of nucleosomes in lupus. Arthritis Rheum. 1999;42(5):833–43 [DOI] [PubMed] [Google Scholar]
- 33.Zeerleder S, Zwart B, te Velthuis H, Stephan F, Manoe R, Rensink I, et al. Nucleosome-releasing factor: a new role for factor VII-activating protease (FSAP). FASEB J. 2008;22(12):4077–84 [DOI] [PubMed] [Google Scholar]
- 34.Zeerleder S, Zwart B, Wuillemin WA, Aarden LA, Groeneveld AB, Caliezi C, et al. Elevated nucleosome levels in systemic inflammation and sepsis. Crit Care Med. 2003;31(7):1947–51 [DOI] [PubMed] [Google Scholar]
- 35.Zeerleder S, Stephan F, Emonts M, de Kleijn ED, Esmon CT, Varadi K, et al. Circulating nucleosomes and severity of illness in children suffering from meningococcal sepsis treated with protein C. Crit Care Med. 2012;40(12):3224–9 [DOI] [PubMed] [Google Scholar]
- 36.Chen Q, Ye L, Jin Y, Zhang N, Lou T, Qiu Z, et al. Circulating nucleosomes as a predictor of sepsis and organ dysfunction in critically ill patients. Int J Infect Dis. 2012;16(7):e558–e64 [DOI] [PubMed] [Google Scholar]
- 37.Miller AC, Gladwin MT. Pulmonary complications of sickle cell disease. Am J Respir Crit Care Med. 2012;185(11):1154–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Muller B, Peri G, Doni A, Torri V, Landmann R, Bottazzi B, et al. Circulating levels of the long pentraxin PTX3 correlate with severity of infection in critically ill patients. Crit Care Med. 2001;29(7):1404–7 [DOI] [PubMed] [Google Scholar]
- 39.van Nieuwenhuijze AE, van Lopik T, Smeenk RJ, Aarden LA. Time between onset of apoptosis and release of nucleosomes from apoptotic cells: putative implications for systemic lupus erythematosus. Ann Rheum Dis. 2003;62(1):10–4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nuijens JH, Abbink JJ, Wachtfogel YT, Colman RW, Eerenberg AJ, Dors D, et al. Plasma elastase alpha 1-antitrypsin and lactoferrin in sepsis: evidence for neutrophils as mediators in fatal sepsis. J Lab Clin Med. 1992;119(2):159–68 [PubMed] [Google Scholar]
- 41.Ballas SK, Lewis CN, Noone AM, Krasnow SH, Kamarulzaman E, Burka ER, et al. Clinical, hematological, and biochemical features of Hb SC disease. Am J Hematol. 1982;13(1):37–51 [DOI] [PubMed] [Google Scholar]
- 42.Powars DR, Hiti A, Ramicone E, Johnson C, Chan L. Outcome in hemoglobin SC disease: a four-decade observational study of clinical, hematologic, and genetic factors. Am J Hematol. 2002;70(3):206–15 [DOI] [PubMed] [Google Scholar]
- 43.Vasavda N, Ulug P, Kondaveeti S, Ramasamy K, Sugai T, Cheung G, et al. Circulating DNA: a potential marker of sickle cell crisis. Br J Haematol. 2007;139(2):331–6 [DOI] [PubMed] [Google Scholar]
- 44.Ball JB, Khan SY, McLaughlin NJ, Kelher MR, Nuss R, Cole L, et al. A two-event in vitro model of acute chest syndrome: the role of secretory phospholipase A2 and neutrophils. Pediatr Blood Cancer. 2012;58(3):399–405 [DOI] [PubMed] [Google Scholar]
- 45.Haynes J, Jr, Obiako B, King JA, Hester RB, Ofori-Acquah S. Activated neutrophil-mediated sickle red blood cell adhesion to lung vascular endothelium: role of phosphatidylserine-exposed sickle red blood cells. Am J Physiol Heart Circ Physiol. 2006;291(4):H1679–H85 [DOI] [PubMed] [Google Scholar]
- 46.Savchenko AS, Inoue A, Ohashi R, Jiang S, Hasegawa G, Tanaka T, et al. Long pentraxin 3 (PTX3) expression and release by neutrophils in vitro and in ulcerative colitis. Pathol Int. 2011;61(5):290–7 [DOI] [PubMed] [Google Scholar]
- 47.Jaillon S, Peri G, Delneste Y, Fremaux I, Doni A, Moalli F, et al. The humoral pattern recognition receptor PTX3 is stored in neutrophil granules and localizes in extracellular traps. J Exp Med. 2007;204(4):793–804 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nur E, van Beers EJ, Martina S, Cuccovillo I, Otten HM, Schnog JJ, et al. Plasma levels of pentraxin-3, an acute phase protein, are increased during sickle cell painful crisis. Blood Cells Mol Dis. 2011;46(3):189–94 [DOI] [PubMed] [Google Scholar]