

Abstract
Aims
We investigate native and introduced populations of Solanum rostratum, an annual, self-compatible plant that has been introduced around the globe. This study is the first to compare the genetic diversity of Solanum rostratum between native and introduced populations. We aim to (1) determine the level of genetic diversity across the studied regions; (2) explore the likely origins of invasive populations in China; and (3) investigate whether there is the evidence of multiple introductions into China.
Methods
We genotyped 329 individuals at 10 microsatellite loci to determine the levels of genetic diversity and to investigate population structure of native and introduced populations of S. rostratum. We studied five populations in each of three regions across two continents: Mexico, the U.S.A. and China.
Important Findings
We found the highest genetic diversity among Mexican populations of S. rostratum. Genetic diversity was significantly lower in Chinese and U.S.A. populations, but we found no regional difference in inbreeding coefficients (F IS) or population differentiation (F ST). Population structure analyses indicate that Chinese and U.S.A. populations are more closely related to each other than to sampled Mexican populations, revealing that introduced populations in China share an origin with the sampled U.S.A. populations. The distinctiveness between some introduced populations indicates multiple introductions of S. rostratum into China.
Introduction
Studying the genetic diversity and structure of introduced populations is a key component to understand the potential of introduced species to establish and spread in the novel range [1,2]. For example, severe reductions in genetic diversity may limit the ability of introduced populations to quickly adapt to novel environmental conditions as rapid evolution in invasive species is expected to occur from existing genetic variation [3]. Nevertheless, there are examples of successful invasive species in the absence of significant amounts of neutral genetic diversity [4,5], and even small populations may maintain genetic variation in quantitative traits [6]. In contrast, multiple introductions may be important not only for maintaining variation in introduced populations [7], but also in bringing about novel combinations of genetic variation not seen in the native range. It is thus not surprising that investigating the population genetics of invasive species continues to be at the heart of many ecological, evolutionary and conservation studies.
Solanum rostratum is a self-compatible species that produces nectarless hermaphroditic flowers with dimorphic anthers, and is pollinated by bees [8]. Individual plants can grow to approximately 1m in height and produce up to 1915 fruits (72.7± 52.2; mean ± SD, range: 12-1915), each containing an average of 41 seeds; single plants have been recorded to produce in excess of 78000 seeds [9]. Thought to be originated from a region centred on the Mexican highlands [10,11], Solanum rostratum has spread to the U.S.A., Canada [12], Europe, Australia [13], the former Soviet Union [10], South Korea, and China [14,15]. In many of these areas, S. rostratum is treated as a noxious weed as it grows aggressively following habitat disturbance [15,16], and livestock is discouraged from grazing on vegetation where it grows as thorns cover all the plant except the flowers and can cause poisoning if ingested [16].
One of the most recent invasions of S. rostratum has occurred in China during the last 30 years, with the first record made in ChaoYang city of the Liaoning province in 1981 [17]. Despite being a relatively new arrival, S. rostratum has spread across a large area in northern China, namely in the provinces of Beijing, Hebei, Jilin, Liaoning, Shanxi provinces and Xinjiang Uygur Autonomous Region [9,18-20]. As in other invaded regions, Chinese populations of S. rostratum usually grow in open, disturbed habitats, such as roadsides, fallow fields and along train tracks. Previous studies have shown on-going dispersal towards the north of China, and indicate that S. rostratum is at potential risk of an outbreak [13].
We use recently developed genetic tools [21] to investigate the genetic diversity and population structure of native and introduced populations of S. rostratum. We studied five populations in each of three regions across two continents: Mexico, the U.S.A. and China. We genotyped individuals using 10 microsatellite loci to address the following questions: (1) What is the level of genetic diversity across the studied regions and to what extent is genetic diversity reduced in introduced populations? (2) What are the likely sources of origin of invasive populations in China inferred from the genetic relationships among samples? (3) Is there evidence of multiple introductions into China? Our study represents the first attempt to characterise the genetic diversity and population structure of native and introduced populations of S. rostratum and offers a unique insight into the historical pathways of dispersal of this invasive weed.
Materials and Methods
Population sampling
We randomly collected fresh leaves of from individuals of Solanum rostratum. The sampling sites were located in road sides, banks of rivers, waste land. These sites didn't belong to a national park or other protected area of land and the relevant regulatory body concerned with protection of wildlife, and they also didn't belong to private land. We confirm that the field studies did not involve endangered or protected species. A total of 15 populations were sampled from July 2010 to June 2011. Five populations were sampled from each of the following three regions: central Mexico, the Kansas-Oklahoma region in the U.S.A. where records date back at least to the 1880's (Kansas State University Herbarium) and northern China (Table 1, Figure 1). These three regions were chosen to represent a range of residence histories for S. rostratum from the native range in Mexico, to the Kansas-Oklahoma region where S. rostratum has been reported for at least 130 years, to the recent introduction of this species to China in the last 30 years.
Table 1. Localities and number of individuals successfully genotyped for each of the 15 populations analysed in this study.
| Code | Region and Population | State or Province | Latitude | Longitude | Altitude (m) | Individuals genotyped |
|---|---|---|---|---|---|---|
| China | ||||||
| BC | Baicheng | Jilin | 45.352° | 122.501° | 154 | 24 |
| CY | Chaoyang | Liaoning | 41.273° | 120.186° | 190 | 23 |
| WSL | Zhangjiakou | Hebei | 40.454° | 114.552° | 729 | 24 |
| MY | Miyun | Beijing | 40.239° | 116.504° | 85 | 24 |
| TZ | Tongzhou | Beijing | 39.451° | 116.435° | 17 | 24 |
| U.S.A. | ||||||
| HAY | Hays | Kansas | 38.915° | -99.315° | 645 | 24 |
| BOT | Cheyenne Bottoms | Kansas | 38.423° | -98.573° | 548 | 24 |
| ROL | Roll | Oklahoma | 35.836° | -99.730° | 677 | 24 |
| CHE | Cheyenne | Oklahoma | 35.675° | -99.679° | 614 | 24 |
| WIC | Wichita (Snyder) | Oklahoma | 34.631° | -98.791° | 403 | 24 |
| Mexico | ||||||
| VDU | Vicente Guerrero | Durango | 23.744° | -103.996° | 1926 | 19 |
| SLP | San Antonio del Rul | San Luis Potosí | 22.640° | -101.140° | 1873 | 15 |
| SLG | San Luis de la Paz | Guanajuato | 21.310° | -100.514° | 2050 | 22 |
| QSJ | San Juan del Río | Querétaro | 20.377° | -99.993° | 1955 | 16 |
| TEM | San Juan Teotihuacán | Estado de México | 19.684° | -98.859° | 2277 | 18 |
Figure 1. Map showing the location of the 15 sampled populations of Solanum rostratum used in this study.

Left panel: Mexican (native, green circles) and U.S.A. populations from the U.S.A. (residence time >130 years, blue circles). Right panel: Chinese populations (residence time <31 years, red circles). Details of localities and population codes are provided in Table 1.
Sample preparation and genotyping
In each population, fresh leaves were collected from 20—30 randomly chosen individuals and quickly dried in plastic or paper bags with silica gel. Between September and November 2011, DNA was extracted from dried leaves using TIANGEN plant genomic DNA kit (Tiangen Biotech, Beijing, China) following manufacturer’s instructions. Individuals were genotyped at ten microsatellite loci previously developed for S. rostratum [21]. Estimates of allelic dropout, false alleles, and null allele frequencies for a sample of Mexican populations are given in Vallejo-Marín et al. [22]. All loci were amplified in a multiplex PCR using QIAGEN Type-it Microsatellite PCR Kit (Qiagen, Shanghai, China), 100µM concentration of each primer (labelled with one of 6-FAM, VIC, PET, or NED fluorescent dyes; Life Technologies, Shanghai, China), and DNA template. PCR program was as follows: one cycle of 95°C for 5min, 30 cycles of 95°C for 30s, 58°C for 180s, and 72°C for 30s, followed by a final step at 60°C for 30min. Fragment analysis of PCR products was done using an ABI3730xl capillary sequencer with a 80-500bp size standard. Fluorescent profiles were analysed and binned in GeneMapper V3.0 (Applied Biosystem 2002, Foster City, CA, USA). Data are available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.mk58d.
Genetic Analysis
Genetic variation
For each population-locus combination, the following indices were calculated using GENALEX 6.4 [23]: number of alleles (N a); number of effective alleles (N e); expected heterozygosity (H E); observed heterozygosity (H O); inbreeding coefficient (F IS). In addition, for each population we also calculated the proportion of polymorphic loci (P), allelic richness averaged across loci (R S), and the number of private alleles. The significance of F IS was calculated using 150000 randomizations, and adjusting the significance threshold for multiple comparisons using a Bonferroni correction in FSTAT 2.9.3.2 [24].
To test for overall differences in genetic variation between the three geographic regions (Mexico, the U.S.A. and China), we compared allelic richness, H O, H E, F IS and F ST, and relatedness using 1000 permutations in FSTAT [24]. We also tested the hypothesis that populations in the U.S.A. and China have lower levels of genetic variation than Mexican populations using a one-tailed test based on 1000 permutations.
Population Genetic Structure
To assess the level of population genetic structure we used a series of complementary approaches. We calculated pairwise F ST using FSTAT, and used these statistics to compare the average levels of between-population differentiation in the three studied regions. In addition, we calculated pairwise genetic distances between populations using Nei’s (1978) standardized genetic distance Ds. We used the resulting distance matrix to generate an UPGMA tree using TFPGA1.3 [25], and assessed the support for each node using 1000 bootstrap replicates implemented in POPULATIONS 1.2.30 [26].
We also used an analysis of molecular variance (AMOVA, [27]) implemented in Arlequin 3.5.1.3 [28] to partition genetic variation across nested levels: within populations, between populations within geographic regions, and between geographic regions. For the AMOVA, we used the number of different alleles as a measure of genetic variation (FST-like option in Arlequin) and 1000 permutations to test for statistical significance.
In addition, we used InStruct [29] to jointly assign individuals to groups and calculate inbreeding coefficients per population. InStruct is similar to the program STRUCTURE, in that it uses a multi-locus clustering method to probabilistically assign individuals to K groups that minimize the amount of linkage-disequilibrium [30]. Because the assignment of each individual to a given group is done probabilistically, this approach can detect admixed or migrant individuals. Importantly, a difference between STRUCTURE and InStruct is that the latter allows for inbreeding to occur within populations. Incorporating inbreeding in this analysis is important as S. rostratum has a mixed mating system in both native and introduced populations ([22]; L. Yu and A. Lou unpublished data). The InStruct analysis was conducted in a nested fashion. We first analysed the entire data set to determine the number of clusters across the three geographic regions. We ran InStruct with K-values between 1 and 15 (the number of studied populations), using a 100000 burn-in and 100000 MCMC iterations with trimming every 100 generations, and estimating both admixture and cluster's selfing rate. Each MCMC chain was re-run three times. Because accurately determining the optimal number of clusters can be difficult when regions display strong genetic structure, we then ran InStruct separately for the populations in the two identified clusters (see Results): Mexico (K-values 1—5) and USA/China (K-values 1—10), using the same parameter values. The optimal number of clusters (K*) for each analysis was assessed using ∆K statistic [31].
Finally, we used graph theory implemented in POPULATION GRAPH [32] to determine the minimum number of edges (connections) between populations. This approach uses the genetic data to determine the covariance relationships among populations independently of the geographic region of origin, and has been used in similar population genetic analyses [33,34]. An advantage of this method is that it does not rely on specific assumptions about Hardy-Weinberg equilibrium within populations or clusters [32].
Results
Genetic diversity across geographic regions
A total of 329 individuals belonging to 15 populations in Mexico, the U.S.A. and China were successfully genotyped using the panel of 10 microsatellite loci. A full description of genetic diversity parameters per locus per population is provided in Table S1. On average, the proportion of polymorphic loci across populations was high (0.95 ± 0.03; mean ± S.E.), while the average number of alleles per locus per population was relatively low (3.17 ± 0.20; for 21.93 ± 0.84 individuals genotyped per population) (Table 2). Two out of five populations in both China and Mexico had at least one monomorphic locus, while only one of five US populations had a single monomorphic locus (WIC; Table 2).There was a deficit in the observed number of heterozygotes across populations (Table 2), indicative of an average inbreeding coefficient of F IS= 0.252 ± 0.038. Individual populations showed a broad range of inbreeding levels, particularly in China where F IS varied widely (0.023—0.521) but also in the U.S.A. (F IS = 0.104—0.433) and to a lesser extent Mexico (F IS = 0.124—0.371) (Table 2).
Table 2. Genetic diversity estimates for sampled population of Solanum rostratum in China, the U.S.A. and Mexico.
| Region and Population | N | P | Na (range) | Ne (± S.E.) | Allelic richness | HO | HE | FIS FSTAT | Private Alleles |
|---|---|---|---|---|---|---|---|---|---|
| China | |||||||||
| BC | 24 | 0.60 | 1.90 (1–3) | 1.39 (±0.16) | 1.89 | 0.10 | 0.21 | 0.521*** | 1 |
| CY | 23 | 1.00 | 2.60 (2–4) | 1.84 (±0.15) | 2.60 | 0.36 | 0.42 | 0.174** | 0 |
| WSL | 24 | 1.00 | 2.70 (2–5) | 1.72 (±0.17) | 2.69 | 0.28 | 0.37 | 0.280*** | 1 |
| MY | 24 | 1.00 | 2.80 (2–5) | 1.65 (±0.14) | 2.75 | 0.35 | 0.35 | 0.023 | 1 |
| TZ | 24 | 0.90 | 2.30 (1–3) | 1.60 (±0.15) | 2.28 | 0.28 | 0.32 | 0.164* | 0 |
| Mean ± SE | 23.8 ± 0.20 | 0.90 ± 0.08 | 2.9 ± 0.16 | 1.6 ± 0.17 | 2.44± 0.16 | 0.27 ± 0.04 | 0.39 ± 0.04 | 0.6 ± 0.25 | |
| U.S.A. | |||||||||
| HAY | 24 | 1.00 | 2.90 (2–5) | 1.90 (±0.11) | 2.89 | 0.27 | 0.46 | 0.433*** | 0 |
| BOT | 24 | 1.00 | 3.10 (2–5) | 1.92 (±0.12) | 3.09 | 0.42 | 0.45 | 0.104* | 1 |
| ROL | 24 | 1.00 | 2.80 (2–6) | 1.79 (±0.27) | 2.79 | 0.22 | 0.36 | 0.415*** | 0 |
| CHE | 24 | 1.00 | 3.20 (2–7) | 1.88 (±0.27) | 3.18 | 0.29 | 0.4 | 0.303*** | 3 |
| WIC | 24 | 0.90 | 2.70 (1–6) | 1.89 (±0.29) | 2.69 | 0.29 | 0.39 | 0.269*** | 1 |
| Mean ± SE | 24 ± 0 | 0.98 ± 0.02 | 3.4 ± 0.09 | 1.9±0.10 | 2.93 ± 0.09 | 0.30 ± 0.03 | 0.45 ± 0.02 | 1.0 ± 0.55 | |
| Mexico | |||||||||
| VDU | 19 | 1.00 | 4.10 (2–6) | 2.39 (±0.26) | 3.49 | 0.36 | 0.55 | 0.371*** | 7 |
| SLP | 15 | 0.90 | 4.00 (1–8) | 2.24 (±0.22) | 3.5 | 0.44 | 0.51 | 0.165** | 4 |
| SLG | 22 | 1.00 | 4.70 (3–6) | 2.54 (±0.33) | 3.89 | 0.40 | 0.56 | 0.303*** | 5 |
| QSJ | 16 | 0.90 | 3.80 (1–8) | 2.14 (±0.36) | 3.12 | 0.40 | 0.44 | 0.124* | 4 |
| TEM | 18 | 1.00 | 3.90 (2–8) | 2.27 (±0.44) | 3.42 | 0.40 | 0.45 | 0.137** | 4 |
| Mean ± SE | 18 ± 1.22 | 0.98 ± 0.02 | 4.60 ± 0.16 | 2.30 ± 0.16 | 3.48 ± 0.12 | 0.39 ± 0.01 | 0.53 ± 0.02 | 4.80 ± 0.58 | |
| Grand Mean ± S.E. | 21.93±0.84 | 0.95±0.03 | 3.17±0.20 | 1.94±0.08 | 2.95±0.13 | 0.32 ±0.02 | 0.42±0.02 | 2.20±0.56 |
N = number of individuals successfully genotyped; P = Proportion of polymorphic loci; N a = Average number of alleles per locus; Ne= Average number of effective alleles per locus; R s = Allelic richness averaged across loci; H O = Observed heterozygosity averaged across loci; H E = Expected heterozygosity across loci (gene diversity); F IS = inbreeding coefficient calculated in F-STAT, with 150000 randomizations, and a nominal level of significance of 1/1000. * P <0.05; ** P< 0.01; *** P<0.001; bold represents significance after correcting for multiple comparisons.
A comparison across geographic regions indicated significant differences in the pattern of genetic variation. Allelic richness, observed heterozygosity (H O), and gene diversity (H E), were significantly higher in Mexico than in the U.S.A. and China (Table 3). Furthermore, the number of private alleles per region showed a strong contrast with nearly 50% (44/89) of alleles across loci being restricted to Mexico, and only between 3%—6% being found only in either the U.S.A. or China (Fig. 2). In contrast, neither the inbreeding coefficient (F IS) nor population differentiation (F ST) was statistically different between geographic regions (Table 3). The U.S.A. population had the lowest value of population differentiation (F ST = 0.047), while Chinese populations had higher average genetic differentiation (F ST = 0.267). Calculation of F ST in China without the BC population reduced this value to F ST =0.100, intermediate between the values observed in the U.S.A. and Mexico (F ST = 0.159; Table 3). The mean number of private alleles of Mexican populations was nearly eight times larger than in China and four times larger than in the U.S.A. (Table 2). In summary, the highest levels of genetic diversity were observed in Mexican populations, while US populations had only marginally higher levels of diversity than China, while there were no statistically significant regional differences in either inbreeding within populations (F IS) or population differentiation (F ST) (Table 3).
Table 3. Comparison of genetic diversity across geographic regions.
| China | U.S.A. | Mexico | P-value | |
|---|---|---|---|---|
| Allelic richness | 2.162 | 2.56 | 3.483 | 0.001 |
| H O | 0.271 | 0.296 | 0.399 | 0.034 |
| H E | 0.343 | 0.425 | 0.521 | 0.001 |
| F IS | 0.209 | 0.302 | 0.234 | 0.548 |
| F ST | 0.267 | 0.047 | 0.159 | 0.167 |
| U.S.A.-China | Mexico | P-value | ||
| Allelic Richness: | 2.361 | 3.483 | 0.001 | |
| H O | 0.284 | 0.399 | 0.008 | |
| H E | 0.384 | 0.521 | 0.002 | |
| F IS | 0.261 | 0.234 | 0.623 | |
| F ST | 0.171 | 0.159 | 0.544 | |
P-value generated with 1000 randomizations of the data.
Figure 2. Venn diagram of the distribution of all distinct alleles across loci for the three geographic regions studied: Mexico, the U.S.A. and China.

Overlapping areas (intersections) denote shared alleles between two or three regions.
Population Genetic Structure
Pairwise F ST values, representing the degree of genetic differentiation between populations, ranged from 0.018 (CHE-HAY) to 0.399 (BC-TEM) in individual population pairs (Table S2). Among regions, the lowest average genetic differentiation between populations was observed in the U.S.A. (F ST=0.039, range 0.018–0.069), while Chinese populations had the highest average value (F ST=0.139, range 0.039–0.282). Mexican populations had an intermediate average F ST (0.100, range 0.040–0.141). The high genetic differentiation observed among Chinese populations is due in part to the distinctiveness of one population, BC. This population has an average pairwise F ST = 0.260 (0.224–0.282) with other Chinese populations and F ST=0.274 (0.224–0.399) when compared to all other 14 populations. Calculating pairwise F ST without the BC population gives a Chinese average of F ST = 0.059 ± 0.010, similar to the value observed for U.S.A. populations (Table S2). In pairwise comparisons between regions, average genetic differentiation with Mexican populations was F ST = 0.241 ± 0.010 for China, and 0.187 ± 0.005 for the U.S.A. In contrast, average differentiation between U.S.A. and Chinese populations was lower (F ST = 0.102 ± 0.005; F ST = 0.062 ± 0.012 when excluding BC).
The UPGMA analysis calculated from Nei's pairwise genetic distance (Ds; Table S3) showed a split between Mexican and U.S.A./Chinese populations, which is supported by a 61% bootstrap value (Fig. 3). In contrast, Chinese and U.S.A. populations were hardly differentiated from each other, showing shallow branches with low bootstrap support (Fig. 3). The exception to this limited level of genetic distance among populations was the Chinese population BC, which formed a well-supported clade separate from the rest (Fig. 3). With the exception of BC, Mexican populations showed higher levels of genetic distance (deeper branches in Fig. 3; Table S3) than U.S.A./Chinese populations. This pattern of genetic variation is consistent with a recent divergence between the U.S.A. and most Chinese populations studied.
Figure 3. Unweighted pair group method with arithmetic mean (UPGMA) tree depicting the relationships between 15 Solanum rostratum populations from Mexico, the U.S.A. and China.
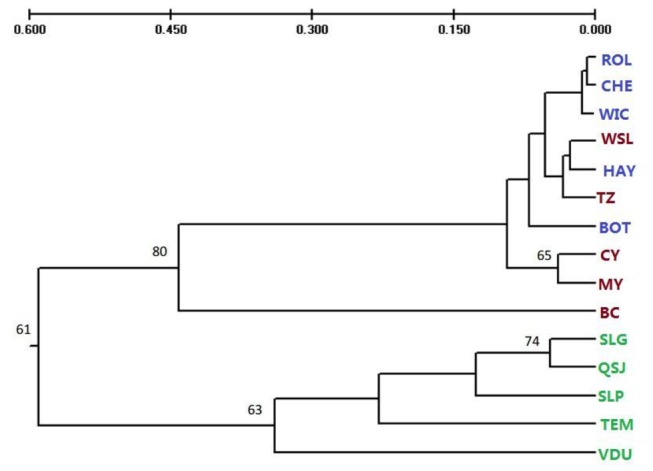
The tree is based on the standardized genetic distances among populations (Nei's D S) calculated from a matrix of genotypes at 10 microsatellite loci. The numbers above the nodes represent percentage of bootstrap support (1000 replications). Only bootstrap support above 50% is shown. Population codes as in Table 1.
The results of the nested AMOVA indicated that the majority (70%) of genetic variation in S. rostratum is contained within populations (Table 4; F ST= 0.299). The proportion of genetic variance partitioned among populations within regions was 14% (F SC= 0.164), while differences among geographic regions explained 16% of the variation (F CT= 0.162). All variance components were statistically significant (P-values < 0.001; Table 4).
Table 4. Nested analysis of molecular variance (AMOVA) of 329 individuals in 15 populations of Solanum rostratum in three geographic regions: Mexico, the USA, and China.
| Source of variation | d.f. | Sum of squares | Variance components | % variation | P-value |
|---|---|---|---|---|---|
| Among regions | 2 | 243.72 | 0.471 | 16.25 | <0.001 |
| Among populations within regions | 12 | 231.72 | 0.394 | 13.59 | <0.001 |
| Within populations | 643 | 1309.19 | 2.036 | 70.17 | <0.001 |
| Total | 657 | 1784.64 | 2.901 |
P-values were calculated using 1023 permutations, under the null hypothesis that a given variance component is equal to zero.
The InStruct analysis on the full data set found the strongest support for two clearly differentiated clusters (K* = 2) that corresponded to either Mexican or U.S.A./Chinese populations (Fig. 4). Increasing the number of clusters to K = 3 did not differentiate between US and Chinese populations, and instead individuals from populations in both geographic regions showed similar probabilities to be assigned to either of two clusters (Fig. 4). The BC population was the only one which was clearly assigned to a single of these clusters (Fig. 4). In the analysis within regions, the optimal number of clusters identified for Mexican populations was K* = 3 (Fig. 4). Populations VDU, QSJ and TEM showed little evidence of admixture at this level, while populations SLP and SLG contained individuals assigned to multiple clusters. The separate analysis of populations in the U.S.A. and China found an optimal number of clusters K* = 3 as well. Again, individuals in population BC were clearly differentiated from the rest forming a homogeneous and distinct cluster, but there was little evidence of population structure in other Chinese and U.S.A. populations (Figure 4).
Figure 4. Posterior probability of cluster assignment in nested InStruct analyses of 329 Solanum rostratum individuals genotyped at 10 microsatellite loci across 15 populations in three geographic regions: Mexico, the U.S.A. and China.
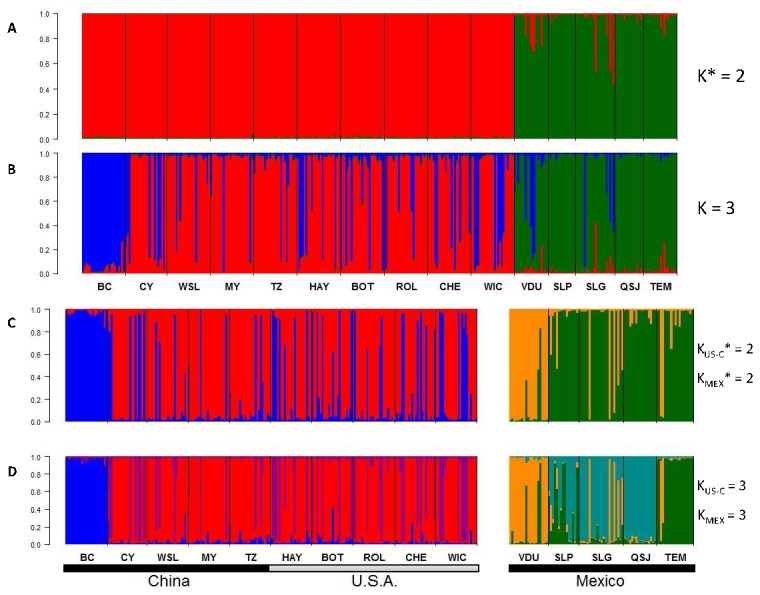
Each bar represents a single individual with populations separated by black lines and arranged from North to South within each region. Panel A depicts the uppermost level of population structure across regions. The optimal number of clusters (K* = 2) was calculated using Evanno’s et al. [31] ∆K statistic. For illustration, panel B shows the assignment probabilities for K = 3. Panel C represent two separate analyses of either U.S.A./Chinese or Mexican populations; the optimal number of clusters [31] in each of these separate analyses was K US-C * =K US-C * = 2. Panel D, shows assignment probabilities for K = 3, for each separate analysis.
In agreement with the UPGMA and InStruct results, the POPULATION GRAPH analysis showed two clearly differentiated groups composed of Mexican populations and China/U.S.A. populations (Fig. 5). The two groups were connected by two edges linking populations BC (China) and SLG and VDU (Mexico). The number of connections within each of these two groups was eight for the five Mexican populations, and 18 for the ten U.S.A./Chinese populations.
Figure 5. Network diagram of 15 populations of Solanum rostratum in Mexico (green circles), the U.S.A. (blue circles), and China (red circles) calculated in POPULATION GRAPH [32].

The diameter of each sphere is proportional to the amount of heterozygosity in each population. Black lines represent connections (edges) linking two populations.
Discussion
Genetic diversity in native and introduced populations
We found the highest genetic diversity (allelic richness, gene diversity, and number of private alleles) among Mexican populations of S. rostratum, and significantly reduced diversity in U.S.A. and Chinese populations (Fig. 2). Whalen (1979) suggested that the highlands around Mexico City are probably the area of origin of S. rostratum, and indicated that this species is most phenotypically variable in central Mexico. The genetic data presented here supports the idea that Mexico is the core of diversity of S. rostratum and that populations in the U.S.A. and China have partly lost that diversity (Figure 2). The reduction in genetic diversity outside Mexico is consistent with a population bottleneck acting on introduced populations as it has been shown for invasive populations of other species (e.g.[5,35,36]). Our results indicate that other processes that can prevent and even reverse the loss of diversity in invasive populations such as multiple introductions [37] have not been strong enough to erase the signature of range expansion in S. rostratum.
Although among populations we observed a wide range of variability in inbreeding rates (Table 2), we detected no dramatic differences in inbreeding across native and introduced regions (Table 3). This finding is perhaps surprising because successful long-distance dispersal may favour conditions leading to higher inbreeding including initially small population sizes, relatedness in the founding population, and selection for increased self-pollination [38-40]. In some cases, the most extreme form of inbreeding (self-pollination) is prevented in invasive species through self-incompatibility (e.g.[41,42]). However, as a self-compatible species, S. rostratum might rely on other aspects of its reproductive biology to maintain similar inbreeding levels in native and introduced populations. For example, the reproductive morphology of S. rostratum, which combines within-flower herkogamy, non-spontaneous pollen release requiring buzz-pollination, and enantiostyly may facilitate maintaining a relatively high outcrossing rate in introduced populations [22].
Origin and genetic structure of the recent invasion to China
All of our analyses showed a stronger genetic affinity of Chinese populations with the U.S.A. than with Mexico, which demonstrates that invasive Chinese populations share a common origin with U.S.A. material. This common origin could be explained in at least two ways. First, Chinese populations could be directly derived from genetic stock in the U.S.A. region. Second, both U.S.A. and Chinese populations could have been independently derived from populations located elsewhere (e.g. unsampled populations in northern Mexico). Distinguishing between these two possibilities would require more extensive sampling throughout North America. However, given the residence time of S. rostratum in the U.S.A. (>130 years), there has been ample opportunities for the U.S.A. to act as the source material for the Chinese populations.
The genetic differentiation between the Chinese populations at Baicheng in Jilin province (BC) raises the possibility that the invasion to China has occurred through multiple introductions. The BC population had the lowest genetic diversity and highest inbreeding coefficient of all populations, so part of this measured differentiation may be due to an extreme founding event [43]. However, another possibility is that the BC population comes from a separate introduction. There are some records of S. rostratum in the former Soviet Union [10], and is possible that BC was colonized from the north via Russia or eastern Mongolia.
Spread of introduced populations of S. rostratum
The relative rapid colonization of S. rostratum in northeast China raises the question as to how is it being dispersed. Using Chaoyang city, the first record of S. rostratum in China in 1981, as the focus for the spread of invasive populations and the location of the extant populations sampled here (at an approximate distance of 500km in both NE and SW directions), we can calculate that this species has dispersed at an average rate of 16km/year and probably much faster. This rate of dispersal is probably much too fast to be accounted naturally, especially given that this species lacks means for long-distance dispersal [10]. Earlier assertions (e.g.[44]) that the spiny fruits of this species could detach from the plant and become entangled with the fur of cattle and bison—incidentally one of the common names of S. rostratum is buffalo-bur—are incorrect. In reality, the fruits remain attached to the plant even after maturation and thus cannot be dispersed in isolation [10]. Instead, after the fruit matures, the spiny calyx surrounding the fruit partly opens and releases the small seeds (2.5mg ± 0.3 mg) which may travel short distances carried by wind or water [10]. In addition, Barrell ([45]; cited in [10]) also suggests that the congested habit of S. rostratum plants in the Great Plains could help them travel longer distances as tumble weeds.
We believe that the rapid dispersal of S. rostratum in China has been facilitated by both massive change in land use and accidental human transport. As a weed of disturbed habitats and roadsides, S. rostratum may have taken advantage of the alteration of the landscape by people and cattle movement, and expanded its range by spreading through newly opened habitats. It is also likely that dispersal of S. rostratum is further facilitated by accidental transport in contaminated grain or forage as it often grows on the margins of crop fields, as well as by transfer of plant and soil waste. In China, some populations may have been also dispersed by seasonally flooded streams and transported as stowaways along train tracks and roads allowing them to quickly travel long distances.
Conclusions
The genetic data presented here demonstrates that the centre of diversity of S. rostratum is in Mexico, and shows that the colonization of this species to other areas has been accompanied by a loss of genetic diversity without an increase in the level of inbreeding. The invasion of S. rostratum to China has probably originated in sampled populations of the U.S.A. or closely related to them. The distinctiveness of the BC population from other Chinese populations indicates multiple introductions of S. rostratum into China.
Supporting Information
Data per locus per population. Obtained with Genalex.
(DOCX)
Pairwise FST values between 15 sampled populations of Solanum rostratum.
(DOCX)
Nei’s Unbiased Identity (above diagonal) and Distance (Ds; below diagonal) among 15 sampled populations of Solanum rostratum derived from GenALEx.
(DOCX)
Acknowledgments
We thank C. Freeman, J. Fornoni and C. Dominguez for facilitating collecting plant material in Kansas and Mexico. R. Ness provided support in running the InStruct analysis.
Funding Statement
Support for this study was provided by a Royal Society of Edinburgh-Chinese National Science Foundation Joint Project to MVM and AL, a grant by the National Natural Science Foundation of China (Project 31070374) to AL, a Royal Society of London research grant (RG2010R1) to MVM, and a Horizon Ph.D. fellowship from the University of Stirling to LSM. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J et al. (2001) The population biology of invasive species. Annu Rev Ecol Syst 32: 305-332. doi: 10.1146/annurev.ecolsys.32.081501.114037. [DOI] [Google Scholar]
- 2. Facon B, Genton BJ, Shykoff J, Jarne P, Estoup A et al. (2006) A general eco-evolutionary framework for understanding bioinvasions. Trends Ecol Evol 21: 130-135. doi: 10.1016/j.tree.2005.10.012. PubMed: 16701488. [DOI] [PubMed] [Google Scholar]
- 3. Prentis PJ, Wilson JR, Dormontt EE, Richardson DM, Lowe AJ (2008) Adaptive evolution in invasive species. Trends Plant Sci 13: 288-294. doi: 10.1016/j.tplants.2008.03.004. PubMed: 18467157. [DOI] [PubMed] [Google Scholar]
- 4. Hollingsworth ML, Bailey JP (2000) Evidence for massive clonal growth in the invasive weed Fallopia japonica (Japanese knotweed). Bot J Linn Soc 133: 463-472. doi: 10.1006/bojl.2000.0359. [DOI] [Google Scholar]
- 5. Voss N, Eckstein RL, Durka W (2012) Range expansion of a selfing polyploid plant despite widespread genetic uniformity. Ann Bot 110: 585-593. doi: 10.1093/aob/mcs117. PubMed: 22730022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Willi Y, Van Buskirk J, Hoffmann AA (2006) Limits to the adaptive potential of small populations. Annu Rev Ecol Evol Syst 37: 433-458. doi: 10.1146/annurev.ecolsys.37.091305.110145. [DOI] [Google Scholar]
- 7. Novak SJ, Mack RN (1993) Genetic variation in Bromus tectorum (Poaceae): comparison between native and introduced populations. Heredity 71: 167-176. doi: 10.1038/hdy.1993.121. [DOI] [Google Scholar]
- 8. Vallejo-Marín M, Manson JS, Thomson JD, Barrett SC (2009) Division of labour within flowers: heteranthery, a floral strategy to reconcile contrasting pollen fates. J Evol Biol 22: 828-839. doi: 10.1111/j.1420-9101.2009.01693.x. PubMed: 19320798. [DOI] [PubMed] [Google Scholar]
- 9. Lin Y, Tan DY (2007) The potential and exotic invasive plant: Solanum rostratum . Acta Phytotaxonomiea Sinica 45: 675-685. doi: 10.1360/aps07010. [DOI] [Google Scholar]
- 10. Whalen MD (1979) Taxonomy of Solanum section Androceras . Gentes Herb (Ithaca) 11: 359-426. [Google Scholar]
- 11. Stern SR, Weese T, Bohs LA (2010) Phylogenetic relationships in Solanum section Androceras (Solanaceae). Syst Bot 35: 885-893. doi: 10.1600/036364410X539934. [DOI] [Google Scholar]
- 12. USDA (2012) The PLANTS Database. Available: http://plants.usda.gov. Accessed 2012 July 15.
- 13. Zhong GP, Shen WJ, Wan FH, Wang JJ (2009) Potential distribution areas of Solanum rostratum in China: A prediction with GARP niche model. Chin J Ecol 28: 162-166. [Google Scholar]
- 14. Cho YH, Kim W (1997) A new naturalized plant in Korea. Korean J Plants Tax 27: 277. [Google Scholar]
- 15. Gao F, Xu C, Zhou YL (2005) The evaluation of potential fatalness for a kind of exotic species Solanum rostratum strategies for its control. J Beijing Norm Univ; (Nat Sci) 41: 420-424. [Google Scholar]
- 16. Bassett I, Munro D (1986) The biology of Canadian weeds.78 Solanum carolinense L. and Solanum rostratum Dunal. Can J Plant Sci 66: 977-991. doi: 10.4141/cjps86-120. [DOI] [Google Scholar]
- 17. Guan GQ, Gao DC, Li WY, Ye J , Xin XG et al. (1984) A quarantine weed: Solanum rostratum . Plants Quarantine 4: 25-28. [Google Scholar]
- 18. Zhang SM, Han QZ (1997) Preliminary Study on the foreign plants in Dalian J Liaoning Norm Univ; (Nat Sci) 20: 323-330 [Google Scholar]
- 19. Wang WS, Zheng HQ, Zhu DM, Jiang ZH, Wang YF (2005) Investigation on harmful weed Solanum rostratum . Plants Quarantine 19: 247-248. [Google Scholar]
- 20. Zhao XY, Ma XD, Xu ZW (2007) Non-native Solanum rostratum and its ecological significance appeared in Urumqi, Xinjiang. Adv Earth Sci 22: 167-170. [Google Scholar]
- 21. Vallejo-Marín M, Solis-Montero L, Bacles CF, Lepais O (2011) Thirteen microsatellites developed by SSR-enriched pyrosequencing for Solanum rostratum (Solanaceae) and related species. Am J Bot 98: e296-e299. doi: 10.3732/ajb.1100159. PubMed: 21980166. [DOI] [PubMed] [Google Scholar]
- 22. Vallejo-Marín M, Solís-Montero L, Souto Vilaros D, Lee MY (2013) Mating system in Mexican populations of the annual herb Solanum rostratum Dunal (Solanaceae). Plant Biol, 15: 948–54. doi: 10.1111/j.1438-8677.2012.00715.x. PubMed: 23294438. [DOI] [PubMed] [Google Scholar]
- 23. Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6: 288-295. doi: 10.1111/j.1471-8286.2005.01155.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Goudet J (1995) FSTAT (version 1.2): a computer program to calculate F-statistics. J Hered 86: 485-486. [Google Scholar]
- 25. Miller MP (1997) Tools for population genetic analysis (TFPGA), version 1.3. Flagstaff: Department of Biological Sciences, Northern Arizona University. [Google Scholar]
- 26. Langella O (1999) Populations 1.2. 30. French National Center for Scientific Research (CNRS). Available: http://bioinformatics.org/~tryphon/populations/. Accessed 2012 November 10.
- 27. Excoffier L, Smouse PE, Quattro JM (1992) Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131: 479-491. PubMed: 1644282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Excoffier L, Lischer HE (2010) Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour 10: 564-567. doi: 10.1111/j.1755-0998.2010.02847.x. PubMed: 21565059. [DOI] [PubMed] [Google Scholar]
- 29. Gao H, Williamson S, Bustamante CD (2007) A Markov chain Monte Carlo approach for joint inference of population structure and inbreeding rates from multilocus genotype data. Genetics 176: 1635-1651. doi: 10.1534/genetics.107.072371. PubMed: 17483417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155: 945-959. PubMed: 10835412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14: 2611-2620. doi: 10.1111/j.1365-294X.2005.02553.x. PubMed: 15969739. [DOI] [PubMed] [Google Scholar]
- 32. Dyer RJ, Nason JD (2004) Population graphs: the graph theoretic shape of genetic structure. Mol Ecol 13: 1713-1727. doi: 10.1111/j.1365-294X.2004.02177.x. PubMed: 15189198. [DOI] [PubMed] [Google Scholar]
- 33. Lowry DB, Rockwood RC, Willis JH (2008) Ecological reproductive isolation of coast and inland races of Mimulus guttatus . Evolution 62: 2196-2214. doi: 10.1111/j.1558-5646.2008.00457.x. PubMed: 18637837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Dyer RJ, Nason JD, Garrick RC (2010) Landscape modelling of gene flow: improved power using conditional genetic distance derived from the topology of population networks. Mol Ecol 19: 3746-3759. doi: 10.1111/j.1365-294X.2010.04748.x. PubMed: 20723052. [DOI] [PubMed] [Google Scholar]
- 35. Henry P, Le Lay G, Goudet J, Guisan A, Jahodová Š et al. (2009) Reduced genetic diversity, increased isolation and multiple introductions of invasive giant hogweed in the western Swiss Alps. Mol Ecol 18: 2819-2831. doi: 10.1111/j.1365-294X.2009.04237.x. PubMed: 19500248. [DOI] [PubMed] [Google Scholar]
- 36. Zhang YY, Zhang DY, Barrett SC (2010) Genetic uniformity characterizes the invasive spread of water hyacinth (Eichhornia crassipes), a clonal aquatic plant. Mol Ecol 19: 1774-1786. doi: 10.1111/j.1365-294X.2010.04609.x. PubMed: 20529068. [DOI] [PubMed] [Google Scholar]
- 37. Bossdorf O, Auge H, Lafuma L, Rogers WE, Siemann E et al. (2005) Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 144: 1-11. doi: 10.1007/s00442-005-0070-z. PubMed: 15891837. [DOI] [PubMed] [Google Scholar]
- 38. Baker HG (1955) Self-compatibility and establishment after 'long-distance' dispersal. Evolution 9: 347-349. doi: 10.2307/2405656. [DOI] [Google Scholar]
- 39. Baker HG (1974) The evolution of weeds. Annu Rev Ecol Syst 5: 1-24. doi: 10.1146/annurev.es.05.110174.000245. [DOI] [Google Scholar]
- 40. Barrett SC (2011) Why reproductive systems matter for the invasion biology of plants. Fifty Years of Invasion Ecology: The Legacy of Charles Elton: 195-210. [Google Scholar]
- 41. Friedman J, Barrett SC (2008) High outcrossing in the annual colonizing species Ambrosia artemisiifolia (Asteraceae). Ann Bot 101: 1303-1309. doi: 10.1093/aob/mcn039. PubMed: 18387973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Lafuma L, Maurice S (2007) Increase in mate availability without loss of self-incompatibility in the invasive species Senecio inaequidens (Asteraceae). Oikos 116: 201-208. doi: 10.1111/j.0030-1299.2007.15220.x. [DOI] [Google Scholar]
- 43. Whitlock MC, McCauley DE (1990) Some population genetic consequences of colony formation and extinction: genetic correlations within founding groups. Evolution 44: 1717-1724. doi: 10.2307/2409501. [DOI] [PubMed] [Google Scholar]
- 44. Tower WL (1906) An investigation of evolution in chrysomelid beetles of the genus Leptinotarsa . Washington DC: Carnegie Institution of Washington. [Google Scholar]
- 45. Barrell J (1975) The Red Hills of Kansas: Crossroads of Plant Migrations: an Essay in Plant Geography. Rockford: Natural Land Institute. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data per locus per population. Obtained with Genalex.
(DOCX)
Pairwise FST values between 15 sampled populations of Solanum rostratum.
(DOCX)
Nei’s Unbiased Identity (above diagonal) and Distance (Ds; below diagonal) among 15 sampled populations of Solanum rostratum derived from GenALEx.
(DOCX)