

Abstract
Since the 1990s we have known of the fascinating ability of a complex set of professional antigen presenting cells (APCs; dendritic cells, monocytes/macrophages, and B lymphocytes) to mediate HIV-1 trans infection of CD4+ T cells. This results in a burst of virus replication in the T cells that is much greater than that resulting from direct, cis infection of either APC or T cells, or trans infection between T cells. Such APC-to-T cell trans infection first involves a complex set of virus subtype, attachment, entry, and replication patterns that have many similarities among APC, as well as distinct differences related to virus receptors, intracellular trafficking, and productive and nonproductive replication pathways. The end result is that HIV-1 can sequester within the APC for several days and be transmitted via membrane extensions intracellularly and extracellularly to T cells across the virologic synapse. Virus replication requires activated T cells that can develop concurrently with the events of virus transmission. Further research is essential to fill the many gaps in our understanding of these trans infection processes and their role in natural HIV-1 infection.
1. Introduction
The uptake of human immunodeficiency virus 1 (HIV-1) by professional antigen presenting cells (APCs) and subsequent transfer of virus to CD4+ T cells can result in explosive levels of virus replication in the T cells. This could be a major pathogenic process in HIV-1 infection and development of acquired immunodeficiency syndrome (AIDS). This process of trans (Latin; to the other side) infection of virus going across from the APC to the T cell is in contrast to direct, cis (Latin; on this side) infection of T cells by HIV-1. Note that this is a narrow definition of trans infection, as direct, cell-to-cell spread of virus is a classic phenomenon in viral infections [1], including HIV-1 [2, 3]. In fact, T cell-to-T cell infection involves many factors that are part of APC-to-T cell trans infection, including expression of CD4 on T cells and formation of a virologic synapse [4, 5] and evasion of neutralizing antibody [6] and the viral inhibitory effects of antiretroviral treatment (ART) [7, 8]. The use of trans infection in this review, however, refers to the in vitro model where infection of APC with low levels of HIV-1 leads to replication of virus in T cells, that is, orders of magnitude more than cis infection of either APC or T cells, and T cell-to-T cell or APC-to-APC trans infections.
The outcome of the APC-to-T cell trans infection process has been considered to be central to sexual transmission of HIV-1 at mucosal (anal and vaginal) and epidermal (foreskin) sites [9–11]. A further, potentially critical feature is its role in progression of HIV-1 infection. In either case, the initial phase of the HIV-1 trans infection process involves unique, cis interactions and replication cycles of virus in the major types of professional APC, that is, subsets of dendritic cells (DC), monocytes/macrophages, and B lymphocytes. Recent reviews have focused on DC-T cell [12] and macrophage-T cell [13] trans infections. This review will focus on the role of each type of APC in HIV-1 trans infection, how these infected cells transfer virus to the CD4+ T cell, and the outcome of this “kiss of death.” Note that this review does not cover information on trans infection of simian immunodeficiency virus (SIV) in nonhuman primates. The reader is referred to recent reviews on SIV infection for this information [14, 15].
2. Langerhans Cells (LC), Dermal DC (dDC), and Interdigitating DC (idDC)
2.1. Cis Infection: Introduction
LC serve among a family of cellular sentries detecting microorganisms that enter the epidermis, and most pertinent to HIV-1, the vagina, ectocervix, and male foreskin [16]. They recognize pathogens through C-type lectin receptor (CLR), Toll-like receptor (TLR), and other pattern recognition receptors [17]. LC express langerin (CD207), a CLR that binds microorganisms for endocytosis, and have Birbeck granules that are involved in endosomal recycling. LC are negative for the DC-specific intercellular adhesion molecule-3-grabbing nonintegrin (DC-SIGN; CD209) CLR. LC are therefore primarily distinguished from dDC and idDC in that the latter are langerin negative and express DC-SIGN. Note that in this review, idDC refers to the conglomeration of tissue and interstitial myeloid DC, which are understudied in HIV-1 infection. These distinguishing properties are important in how DC subsets can interact with HIV-1 and result in cis and trans infections. Figure 1 presents basic phenotypic characteristics of LC, dDC, and idDC that could be involved in HIV-1 cis and trans infection during mucosal transmission. The complexity of human DC is becoming more evident, as detailed in a recent, comprehensive study comparing blood and skin DC for a plethora of markers [18].
Figure 1.
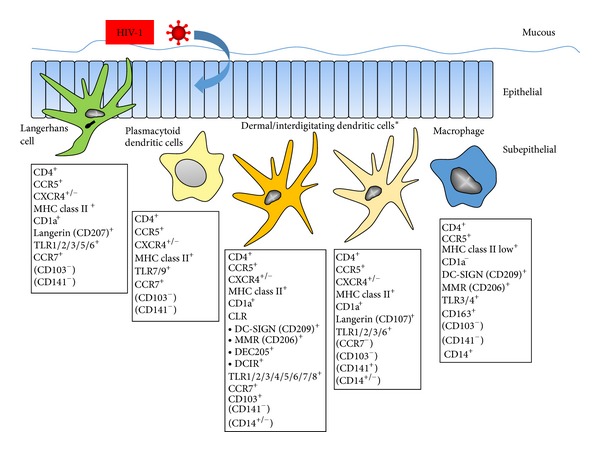
Major types of human, mucosal, professional APC that serve as targets for primary HIV-1 infection in the mucosa. Expression of the CXCR4 coreceptor can occur after DC activation in vivo and in vitro culture. *Note that dDC and idDC are undergoing continuous revision (hence the markers in parentheses) and could represent a continuum of a single type of DC regulated by local environmental influences. CD103 = integrin alpha 3; CD141 = blood dendritic cell antigen 3; CD163 = scavenger receptor.
2.2. Cis Infection: Early Studies
The first indication that DC were involved in HIV-1 infection was a report in 1984 that patients with AIDS had a significant reduction in the number of LC in the epidermis [19]. This was followed by a series of histopathological studies that either supported [20] or failed to support [21–23] that LC were infected with HIV-1 in vivo. There was also some in vitro evidence of HIV-1 infection of LC [24]. These early, contradictory findings were related to limitations in the clinical and laboratory study designs and available technologies to detect HIV-1 infected cells in vivo.
By 1996, a comprehensive investigation conducted by Compton et al. [25] was able to quantitate epidermal LC densities in serial biopsies from two burn patients over 11 years after they were infected with HIV-1 via contaminated blood transfusions. Immunostaining for intracellular p24 viral capsid antigen showed that HIV-1-infected LC constituted an extraordinary one-third to one-half of the epidermal LC over many years of asymptomatic infection. Interestingly, the number of epidermal LC in the HIV-1 infected persons was comparable to uninfected persons and occasionally increased. Other studies detecting viral antigen or HIV-1 RNA by polymerase chain reaction (PCR) supported that LC are infected in vivo with HIV-1 [26–28]. Epidermal LC isolated from AIDS patients have a range of 107 to 3,645 HIV-1 DNA copies per 105 LC, comparable to that found in CD4+ T cells [29]. However, on a direct cellular basis, the relative frequency of idDC harboring HIV-1 DNA from spleens of AIDS patients ranges from 1/720 to 1/18,000, which is 10–100 times less than CD4+ T cells (range: 1/17 to 1/190) [30]. Nevertheless, it is clear that a significant number of LC and idDC are targeted by HIV-1 in vivo during untreated HIV-1 infection.
Similar to the improved in vivo evidence, data from upgraded in vitro models reinforced that LC can be cis infected with HIV-1. LC purified from migrating cells of human epidermis explants express CD4, the primary receptor for HIV-1 [31], and coreceptor CCR5 [32, 33], which is required for infection by R5 tropic strains of HIV-1 [34]. Consequently, cultured LC can be productively infected with R5 HIV-1 [35].
2.3. Cis Infection: HIV-1 Receptors
Expression of the primary CD4 receptor and chemokine coreceptors for HIV-1 in dDC and idDC differs as to their mucosal, dermal, and tissue sites, cytokine/chemokine milieu, and study design (e.g., in situ, ex vivo, and in vitro) [36] (Figure 2). Thus, early studies failed to detect expression of CD4 on freshly isolated dDC [37], while others showed intense CD4 expression on tonsil idDC subsets [38]. Indeed, the distinction between dDC and idDC is further blurred in that various subsets of dDC appear to migrate to and populate draining lymph nodes [39]. Evidently CD4 is expressed on various if not all myeloid DC populations in vivo, although there is little understanding of its function besides serving as the primary receptor for HIV-1 [40].
Figure 2.
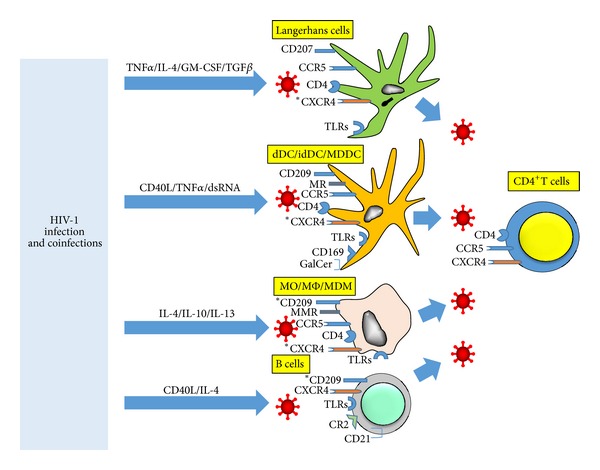
Factors that enhance cis infection of APC also enhance trans infection. Inflammatory cytokines produced in response to HIV-1 infection and co-infections enhance various stages of HIV-1 infection of APC, including maturation of DC and activation of B cells. This in turn is essential for transfer of HIV-1 to CD4+ T cells and conferring them with the ability to produce prodigious amounts of HIV-1. *CXCR4 and CD209 expressed on activated APC.
The expression of CD4 and CCR5, as well as CLR, on LC in endocervical mucosa and foreskin is purported to be a basis for the preferential sexual transmission of R5 HIV-1 [41–44]. The dDC and idDC also express these HIV-1 receptors, although there are little phenotypic data on these human APC. Steady state LC do not constitutively express the other major HIV-1 coreceptor, CXCR4, in vivo, but do so after in vitro culture [33] (Figure 2). Coreceptor expression is upregulated by cytokines IL-4 and TGF-β [45] and GM-CSF [46]. In addition, LC derived by treatment of CD34+ neonatal cord blood stem cells with cytokines (GM-CSF, IL-4, tumor necrosis factor α (TNF-α), and TGF-β) express CD4 and both CCR5 and CXCR4 and can be productively infected with R5 and X4 tropic HIV-1 [45, 47]. HIV-1 replication can be further enhanced by treatment of such cultured LC with recombinant CD40L [48]. These studies support that activated LC, and likely dDC and idDC, can occur in vivo through interactions with activated CD4+ T cells expressing cytokines and CD40L and are thereby rendered permissive for both R5 and X4 HIV-1 infection [48].
Additional evidence supporting the conventional pathway for cis infection of LC by either R5 or X4 strains is that this can be blocked by pretreatment of the cells with natural and synthetic ligands for CCR5 (RANTES; regulated on activation, normally T cell-expressed and secreted) or CXCR4 (SDF-1; stromal cell-derived factor 1) [49, 50], as well as blockers of gp41 fusion [51]. Moreover, LC from persons who are heterozygous for the 32 base pair deletion mutation in CCR5 ORF (delta 32/wt) are less susceptible to HIV-1 infection than LC from CCR5 ORF wt/wt individuals [35].
Activation of LC due to the local cytokine milieu could explain how both CCR5 and CXCR4 are expressed on cervical DC [52]. Furthermore, these cytokines can be induced in LC in response to sexually transmitted infections. Yeast (Candida albicans) and bacterial (Gram negative and positive organisms and their products) infections act as agonists for TLR2/4/5 in vaginal and skin explants and also induce TNFα production [53, 54]. This enhances susceptibility of LC to HIV-1 cis infection. TLR triggering leads to lower levels of the HIV-1 restriction factor, apolipoprotein B mRNA-editing, enzyme-catalytic, polypeptide-like 3G (APOBEC3G), in LC. Interestingly, this phenomenon is not demonstrable in monocyte-derived DC (MDDC).
Herpes simplex virus type 2 (HSV-2), a common sexually transmitted pathogen, enhances both HIV-1 cis infection of LC and trans infection of T cells by LC [55]. This could be related to the protective effect of langerin against HIV-1 cis infection of LC [56], in that HSV-2 competes with HIV-1 for attachment to langerin and also induces a decrease in langerin expression. Interestingly, this effect does not require full cycle replication of HSV-2 in the LC, as it can be mediated by UV-inactivated virus and the TLR3 ligand, poly I:C (synthetic double stranded RNA).
Notably, these studies used different bodily sources and laboratory methods to isolate and develop LC in vitro which likely result in variable properties including state of activation. Early studies indicated that cis infection of APC can vary as to the source of HIV-1 and in vitro conditions [57], which is still being reestablished in the recent literature [58]. Thus, for example, LC captured from epidermal sheets of suction blisters have different phenotypic and functional properties after in vitro culture [59]. Such epidermal-derived LC can be infected with either R5 or X4 strains of HIV-1 and transmit these viruses to T cells within tonsillar tissue explants [60]. Intracellular CXCR4 present in LC [33] is expressed externally through an activation mechanism during in vitro processing. This could explain how CXCR4 is expressed on freshly isolated epidermal LC and that these cells can be infected with X4 HIV-1 [46]. Likewise, activated LC derived from CD34+ stem cells can efficiently trans infect T cells with X4 HIV-1 [61]. This also suggests that lack of CCR5 expression by LC in some studies [62] could be due to vagaries of the in vitro model.
Infection of LC and dDC with HIV-1 has undergone revisionist changes in recent years as different DC subsets have been recognized. Using the human skin explant model, Kawamura and colleagues [63] demonstrated that 2 distinct DC subsets emigrate upon culture, that is, CD1a+langerin+DC-SIGN−LC and CD1a+ langerin−DC-SIGN−dDC, as well as CD1a−langerin−DC-SIGN+ dermal macrophages (Figure 1). CD1a is a transmembrane glycoprotein used to classify phenotypes of tissue DC. It is similar in structure to MHC class I and can heterodimerize with β-2 microglobulin and present lipids to T cells. Only LC could be productively infected with HIV-1 R5. Differences in HIV-1 infection of LC subsets could be related to their levels of activation. Indeed, under steady-state conditions, RNA transcriptome profiles of LC are indicative of less inflammatory, migratory, and T cell stimulatory activity, as well as greater expression of cell adhesion and retention molecules, than dDC [64]. More complexity in DC subsets and HIV-1 infection is evident in that dDC include a CD14+ subpopulation [65] that appears permissive for productive HIV-1 infection [66].
Recent studies provide further evidence that the susceptibility of LC to HIV-1 cis infection and productive replication is dependent on their state of maturation [11, 56]. Immature LC bind and take in HIV-1 via langerin. However, they do not support productive HIV-1 infection, as virus taken up by langerin is internalized into Birbeck granules and destroyed. This protective effect can be overcome by adding larger amounts of HIV-1 to the LC cultures, whereby immature LC become productively infected [67]. Such a natural barrier to HIV-1 infection could be highly significant in preventing HIV-1 transmission. This dichotomy in cis infection of LC with HIV-1 based on langerin and the extracellular concentration of infectious HIV-1 has been challenged by evidence that HIV-1 efficiently infects LC in vitro [68]. Moreover, it is not clear how infection of LC with low levels of HIV-1 can result in trans infection of T cells if virus is destroyed within the LC. One explanation of this paradox is that there is an altered, lower expression of langerin due to activation of LC [61]. This allows virus to enter the LC without being destroyed and traffic to CD1a-containing, multivesicular bodies, where clusters of virions are transmitted to T cells. Once infected by HIV-1, immature LC also become more mature and consequently are more competent in activating CD4+ T cells [69]. This effect on LC maturation could thereby enhance trans infection.
Besides the X4/R5 paradigm of infection patterns of HIV-1, enhanced susceptibility of vaginal LC to infection with the subtype E clade of HIV-1 has been proposed as the basis for a predominance of heterosexual transmission in Thailand [70]. However, this result was not reproducible [71].
HIV-1 cis infection of LC as well as idDC can be blocked by anti-HIV-1 neutralizing and nonneutralizing antibodies. This involves the Fc γ receptor for IgG and IgA, implicating this process in protection of mucosa from HIV-1 infection [68].
2.4. Trans Infection
Early evidence from cocultures of epidermal sheets with T cell lines first suggested that LC can trans infect T cells with HIV-1 [72]. There were concurrent findings that blood-derived DC could trans infect T cells with HIV-1 [73], which are detailed later. This principle regarding LC was confirmed and extended by Pope et al. [74] using an in vitro skin explant model to show presence of stable, nonproliferating conjugates of DC (presumably including LC) and memory CD4+ T cells. Addition of HIV-1 to these mixed DC-T cell cultures resulted in production of virus by 4 days. It was subsequently formally demonstrated by Ayehunie et al. [75], using the basic DC-T cell coculture model still employed today, that HIV-1-infected LC (and also blood purified DC) transmit virus to uninfected CD4+ T cells. Current principles of DC-T cell trans infection established by this early, pioneering study were (a) cocultivation of LC and DC isolated as monocyte-negative, low density cells, with allogeneic CD4+ T cells results in massive virus production; (b) trans infection requires a short, 30 min cell-cell contact; (c) proteolytic striping of cell surface-bound virus from the LC or DC does not decrease the efficiency of virus transmission, suggesting that internalized virus rather than plasma membrane-bound virus is required for trans infection; (d) effective virus transfer by LC or DC requires prior activation of T cells; (e) LC or DC mediate trans infection of T cells at least 100-fold more efficiently than T cell-to-T cell trans infection. The authors concluded that LC and DC could play a key role in the dissemination of virus to T cells they encounter in skin or lymphoid tissue, a hypothesis that is still prevalent almost 20 years later.
We now know that LC can mediate trans infection of CD4+ T cells with both R5 and X4 strains of HIV-1 [49]. This occurs in LC-T cell clusters, primarily involving proliferating, CD4+ memory T cells [51]. Using a foreskin explant model, it was shown that HIV-1 initially forms virologic synapses between keratinocytes and LC [76]. Virus is soon found within migrating LC in the inner foreskin, with presence of LC-T cell conjugates where there was evidence of T cell trans infection.
With the establishment of the LC-T cell trans infection model, it became clear that various aspects of cis infection of LC described earlier significantly affect their capacity to trans infect T cells. Thus, it was first noted that treatment of LC with anti-CD4 receptor Ab, coreceptor ligands, and antiviral agents had no effect on trans infection of T cells [47]. This suggested a second pathway to the CD4/CCR5 receptor, productive infection, where LC capture HIV-1 by other receptors and transmit the virus to T cells. However, later evidence supported that entry via CD4/CCR5 receptors on LC leads to trans infection of CD4+ T cells [35]. This is supported by the blocking of HIV-1 trans infection from LC by inhibitors of R5 coreceptor binding and gp41 fusion [77]. Hence, there could be at least 2 pathways by which HIV-1 trans infects T cells via LC, as is detailed later.
Finally, intriguing data from a hallmark study by Hladik et al. [78] using a vaginal mucosal explant system supported a different pathway model for HIV-1 trans infection by LC. They showed that HIV-1 R5 infection was first established independently in both intraepithelial CD4+ T and LC through a CCR5 binding pathway. HIV-1 fused with and entered T cells, resulting in productive cis infection, whereas uptake by LC involved endocytosis and nonproductive sequestration of virus particles for up to 3 days. Furthermore, virus particles were observed in membrane synapses between the LC and T cells that emigrated from the epithelium. This study therefore provided rare, in situ evidence supporting the virus synapse between DC and T cells as the principal site of HIV-1 trans infection.
3. Blood Myeloid DC
3.1. Cis and Trans Infection In Vivo
A key to our understanding of the role of myeloid DC in both cis and trans infection with HIV-1 R5 and X4 is direct examination of HIV-1 interactions with primary DC and T cells in blood and tissues of HIV-1 infected persons. As with LC, it is difficult but possible to delineate HIV-1 infection in primary DC examined ex vivo. Thus, a portion of myeloid DC in blood of untreated HIV-1 infected persons is positive for proviral HIV-1 DNA, but at much lower levels than CD4+ T cells [79]. Once on long term, virus suppressive ART, proviral DNA, and viral particles are not detectable in myeloid DC or other types of circulating DC [80]. It is much more difficult, however, to directly prove trans infection of endogenous virus from such blood or tissue DC in vivo. The usual approach therefore has been to examine blood myeloid DC in vitro for their capacity to support cis and trans infection with HIV-1.
The first study to definitively demonstrate HIV-1 trans infection in vitro was over 20 years ago using blood myeloid DC [73]. DC were purified by density gradient centrifugation from peripheral blood mononuclear cells (PBMCs) that were depleted of T cells, B cells, monocytes, and natural killer (NK) cells. These immature DC did not support cis infection with HIV-1 X4 virus but resulted in explosive virus replication when mixed with polyclonally activated, autologous CD4+ T cells. Replication of virus was noted within T cells clustered around the DC. Coincident with this breakthrough discovery was a controversy as to whether such blood DC could actually support productive HIV-1 cis infection. Results from several research groups showed high levels of HIV-1 cis replication in cultures of purified blood DC [81–84]. However, using similar blood-derived, myeloid DC models, others found that DC did not support HIV-1 cis infection yet could result in high levels of trans infection of CD4+ T cells [85]. This could relate to the amount of input virus, as productive cis infection was noted in later studies using purified blood CD11c+ myeloid DC and high titers of input R5 or X4 HIV-1 [86]. The integrin CD11c is commonly used to distinguish phenotypes of tissue DC and is involved in their cell-cell adhesion. Importantly, viral replication occurs in a small percentage of circulating CD1c+ (BDCA-1+) myeloid DC that express CD4 and low levels of CCR5 [87]. These virus-infected DC can trans infect CD4+ T cells. Thus, these data using a “natural” form of myeloid DC derived directly from blood suggest their potential for supporting both HIV-1 cis and trans infection in vivo.
Many of the characteristics of trans infection in these early DC studies still pertain to our current understanding. Thus, trans infection could (1) be blocked by antibodies to CD4 and T cell accessory molecule CD80 [85], (2) be augmented by activated CD4+ T cells expressing CD40L [85], proinflammatory cytokines such as IL-2 [88] and nominal antigens [89, 90], and (3) result in extensive, apoptotic death of the T cells within the DC-T cell clusters [89, 91]. However, cis infection of DC was becoming more complex. Although it appeared to require CD4 [90] and R5 or X4 coreceptors [92], as for HIV-1 infection of LC, there was accumulating evidence that HIV-1 infected the DC by more than one pathway. Electron and light microscopy studies suggested that HIV-1 could enter DC by several routes—fusion with the plasma membrane, endocytosis in coated pits, and phagocytosis [93, 94].
Digging deeper into the pathways of DC infection with HIV-1, Cameron et al. [95] showed that after in vitro exposure to HIV-1, viral DNA was more frequent in purified, blood CD11c+ myeloid DC from HIV-1 negative donors than monocytes or resting CD4+ T cells. Infection was more efficient with R5 than X4 strains of HIV-1. Importantly, infection of CD11c+ DC-T cell cocultures with R5 virus resulted in productive, trans infection of CD4+ T cells, whereas cis infection of these DC with HIV-1 was inefficient.
In 2000, Geijtenbeek et al. [96] reported that myeloid DC expressed the CLR DC-SIGN, which served as a receptor on MDDC for HIV-1 trans infection. Clearly the key to delineating the importance of DC-SIGN expression in both HIV-1 cis and trans infection is whether these in vitro findings using MDDC correspond to in vivo activity. Initial evidence showed that blood DC express very little DC-SIGN and that DC-SIGN is expressed on a greater proportion of dermal DC and idDC. Further proof is that DC-SIGN-expressing mononuclear cells are found in Peyer's patches of gut mucosa [97, 98]. Indeed, 1%–5% of these cells in the rectal mucosa from non-HIV-1 infected adults are DC-SIGN+ compared to only 0.01% of blood mononuclear cells [99]. Gurney et al. [100] established that although gut DC express CD4, CD11c and HLA DR, they do not express CCR5 or CXCR4. Interestingly, DC-SIGN is expressed on CD14+ cells of the same mucosal region. This was an early indication that steady state, tissue macrophages express DC-SIGN, in contrast to blood monocytes and monocyte-derived macrophages (see later).
Most important is that R5 HIV-1 preferentially binds to the HLA DR+ DC-SIGN+ mononuclear cells isolated from the gut mucosa and is efficiently transferred to and replicates in CD4+ T cells [100]. This trans infection can be blocked by mannan and anti-DC-SIGN mAb. Interestingly, gut mucosa mononuclear cells in HIV-1 infected subjects express more DC-SIGN than HIV-1 uninfected persons.
3.1.1. Role of Regulatory T Cells (Treg) in Trans Infection
An early suggestion that Treg could be involved in the expression of DC-SIGN was the strong, positive correlation of the IL-10/IL-12 ratios (mRNA) and DC-SIGN cell numbers in the gut tissues. Notably, Treg are an amalgamation of several varieties of naturally resident and pathogen-induced CD4+ T cells [101]. These regulatory cells can inhibit DC and T cell activation and thereby decrease DC-mediated HIV-1 trans infection [102, 103].
A major Treg pathway involves generation of adenosine from extracellular adenosine triphosphate (ATP) and adenosine diphosphate that is mediated by ectoenzymes CD39 and CD73 on Treg membranes [104]. Adenosine binds to the A2A receptor leading to increased cyclic adenosine monophosphate that inhibits T cell differentiation and proliferation. This provides the negative feedback necessary to prevent tissue damage that may result from a persistent inflammatory state. However, it has been noted that extracellular ATP also dampens DC-T cell trans infection [105]. This ATP effect was linked to lower numbers of trans-infected T cells with both X4 and R5 HIV-1 and had no effect on cis infection of the T cells. Evidence suggested that extracellular ATP led to increased lysosomal degradation of internalized virus in the DC. Clearly we need more information on the potential role of Treg and adenosine in HIV-1 trans infection.
3.1.2. Effect of HIV-1 Infection on Capacity of DC to Mediate HIV-1 Trans Infection
A critical point not usually addressed in trans infection studies is whether in vivo infection with HIV-1 alters the capacity of DC to mediate trans infection. Most of this evidence is suggestive and indirect. Nevertheless, various aspects of the number and functional capacity of blood and tissue myeloid DC could bear on their ability to mediate trans infection. Primary, blood myeloid DC from persons with untreated, progressive HIV-1 infection exhibit low T cell stimulation and cytokine/chemokine production [106]. In fact, circulating myeloid DC levels decline very early in blood during acute HIV-1 infection [107]. Pertinent to trans infection is that myeloid DC directly circulating in blood of untreated HIV-1 infected persons are partially activated [108], possibly due to direct stimulation by HIV-1, but still undergo maturation after stimulation in vitro by TLR ligation. Interestingly, these partially activated DC accumulate in lymph nodes of the infected persons, supporting their potential to mediate trans infection. Such blood myeloid DC also exhibit abnormally low production of the inflammatory cytokines IL-6, TNF-alpha, and IL-12 in vitro [109]. However, this depends on the type of stimulation, as DC from HIV-1 infected subjects produce high levels of cytokines and chemokines in response to stimulation with TLR7/8 agonists [107]. They also retain their ability to stimulate allogeneic T cell responses and upregulate DC maturation markers. Contrasting studies show a defect in circulating myeloid DC early in HIV-1 infection, including poor antigen presentation and production of cytokines [110]. Finally, the very low expression of DC-SIGN on blood myeloid DC does not differ in relation to progression of HIV-1 infection [111].
Of special interest is that HIV-1 derived, uridine-rich ssRNA serves as a ligand for TLR7/8 [112], through which it upregulates programmed cell death 1 ligand 1 (PD-L1; CD274) expression on myeloid DC. PD-L1 in turn is an inhibitory signal on DC which triggers PD-1 on CD8+ T cells, thereby impeding their reactivity [113]. This pathway could impact on HIV-1 trans infection and requires further study.
3.1.3. HIV-2 Trans Infection
HIV-2 infection is much less pathogenic than HIV-1 infection [114]. Interestingly, mature MDDC cannot trans infect autologous CD4+ T cells with either R5 or X4 tropic HIV-2 [115]. This block could be due to poor cis infection of DC with HIV-2. It points to the possibility that a lower capacity to mediate trans infection could be involved in slower progression of HIV-1 infection.
4. MDDC and CD34+ Cell-Derived DC
4.1. Cis and Trans Infection
The major breakthroughs in our understanding of HIV-1 cis and trans infection stem from discovery of simple methods for deriving large numbers of myeloid DC from CD34+ bone marrow stem cells using various growth factors [116] and more importantly from blood monocytes using IL-4 and GM-CSF [117]. These in vitro derived DC are presumed to mirror differentiation of myeloid DC in vivo and have many of the properties of blood and tissue dDC and idDC. Indeed, the bulk of our knowledge of HIV-1 trans infection is based on MDDC derived from peripheral blood monocytes by in vitro culture with IL-4 and GM-CSF. This is because of (1) the ease of accessibility and processing, and relative low cost, of deriving large numbers of MDDC from CD14+ blood monocytes, as well as (2) many phenotypic and functional traits of MDDC that are common to natural myeloid DC circulating in blood. Supporting evidence that HIV-1 cis and trans infection mediated by MDDC is a significant gauge of HIV-1 pathogenesis is that differentiation of monocytes into CD1a+ DC in vitro correlates with low CD4+ T cell counts and high viral loads in HIV-1 infected adults [118]. Nevertheless, similar to studies using LC, dDC, and idDC, there are significant differences among techniques used in studies of trans infection in the MDDC-T cell model that could result in discordant results. These include different strains of HIV-1, purification and maturation techniques for MDDC, and types of CD4+ T cell targets (e.g., autologous and allogeneic; continuous cell lines). In particular, several differences in the pathways of HIV-1 cis infection in MDDC are critical in defining the down-stream events of trans infection.
Basic principles of both cis and trans HIV-1 infection of these DC were soon demonstrated and are pertinent today. First, despite early conflicting studies [119, 120], it became apparent that both MDDC and CD34+ precursor cell-derived DC are poorly permissive for productive HIV-1 cis infection [121–123]. The cis infection process requires CD4, which is more efficient via cell associated virus [124] and can be enhanced by virus opsonized with antibody and complement [125, 126], while blocked by IgG alone via binding to FCγR [127]. However, the fate of IgG-opsonized virus depends on several factors including time of exposure to the MDDC and the opsonization pathway used to enter and traffic within the cells [128]. Complement, that is, C5a, can also be triggered in HIV-1 infected T cells, drawing in immature MDDC and resulting in enhanced trans infection [129].
4.1.1. DC Maturity Significantly Affects HIV-1 Trans Infection
The dogma that has emerged over the years is that immature DC in submucosal tissues (see earlier) capture HIV-1 and taxi it to local draining lymphoid tissues, where they undergo maturation that enhances their antigen presenting properties [130]. These mature DC also are most efficient at trans infection of T cells. Indeed, early studies supporting efficient HIV-1 trans infection by immature DC [131, 132] were not confirmed, as mature DC appear more effective at trans infection of CD4+ T cells [133, 134]. This could, however, be dependent on the form of DC maturation. MDDC matured with CD40L, TNFα, and bacterial lipopolysaccharide (LPS) have a respective, decreasing propensity to support HIV-1 cis infection, while all 3 types of mature MDDC promote efficient trans infection [134]. Moreover, mature MDDC and purified blood myeloid DC display increased uptake of HIV-1 and survival of the engulfed virus [133]. Thus, MDDC that are treated with CD40L, or different combinations of LPS, IFN-γ and double stranded synthetic RNA (poly I:C), produce large amounts of IL-12 (termed (DC-1) that polarize CD4+ T cells to differentiate into type 1 helper (Th1) cells [135]. In contrast, MDDC treated with prostaglandin E2 (termed DC-2) promote the induction of type 2 helper (Th2) cells [136]. The DC-1 mediate more efficient trans infection than DC-2 [137].
Early studies indicated that immature DC support greater cis infection by HIV-1 than mature MDDC [131, 138, 139]. However, this was challenged by further work showing that cis infection was dependent on R5 and X4 tropism and the type of DC maturation agent used. Thus, both immature and mature MDDC show poor fusion and cis infection by X4 virus [140]. This is not due to differential expression of coreceptors, as immature MDDC express similar amounts of CXCR4 and CCR5 [141]. Indeed, immature MDDC can utilize both tyrosine kinase (Src and SyK) [142] and the protein kinase A and C [143] pathways in supporting HIV-1 cis infection. Nevertheless, immature MDDC are less susceptible to cis infection by X4 compared to R5 HIV-1, which is related to less efficient fusion by X4 virus to immature MDDC. Furthermore, trans infection mediated by mature DC with R5 but not X4 virus is enhanced by CXCL12, the chemokine ligand for CXCR4. This could effectively select for R5 virus infection that predominates in the earlier phase of natural HIV-1 infection [144].
A key issue is that, similar to HIV-1 infection of LC, HIV-1 infection of MDDC can inhibit the maturation process [103, 145]. This includes lack of stimulation of allogeneic T cell responses and expression of cell surface maturation molecules such as MHC class II and CCR7.
Of significance is that trans infection from mature DC to T cells is dependent on levels of CD4, as blocking CD4 on immature MDDC enhances their trans infection with X4 virus [146]. This could be due to redirecting of CD4 biasing virus entry into DC through DC-SIGN and subsequent efficient trans infection of T cells. Mature MDDC also have enhanced HIV-1 endocytosis and are highly effective at concentrating HIV-1 in virologic synapses [147].
4.1.2. Cholesterol and Tetraspanins Play a Key Role in HIV-1 Trans Infection
Cholesterol-enriched surface domains of DC are central to their capacity to mediate trans infection [133]. These are concentrated in lipid rafts, which are cholesterol-sphingolipid floating microdomains involved in cholesterol transport, endocytosis, and signal transduction [148]. The rafts contain HIV-1 receptors including DC-SIGN [149, 150]. Hence, it is not surprising that cholesterol and lipid rafts are involved in HIV-1 binding, entry and subsequent intracellular trafficking in DC [151, 152] which is required for HIV-1 trans infection. Moreover, lipid rafts are linked to the actin cytoskeleton and microtubules, whose integrity is also essential for HIV-1 trans infection [153]. Evidence for a role of cholesterol in HIV-1 trans infection is that enhancing cholesterol efflux in DC by triggering of regulators of cholesterol metabolism, that is, peroxisome proliferator-activated receptor gamma and liver X receptor, decreases HIV-1 trans infection [154]. Furthermore, increasing the level of cellular cholesterol by inhibiting the cholesterol transport protein ATP-binding cassette A1 (ABCA1) in these DC restores their ability to mediate HIV-1 trans infection [154].
Interestingly, host-derived lipids embedded within HIV-1 virions are involved in capture of the virus by DC and subsequent trans infection of T cells. In fact, alpha 2-3 ganglioside GM3 can mediate Env-independent binding to DC through its sialic acid residue [155]. This effect is accentuated in mature DC through sialyllactose in the ganglioside [156]. This process of virus capture by mature DC involves binding of sialic acid-binding Ig-like lectin 1 (Siglec-1, CD169) on the surface of DC to sialyllactose-containing gangliosides on the viral membrane [157]. This provides HIV-1 trans infection pathway mediated by mature DC that is independent of CLR.
Intriguing recent reports indicate that IFN-α acts to inhibit viral infection by reducing sterol metabolic activity [158]. This involves the oxysterol cholesterol-25-hydroxylase, which catalyzes the oxidation of cholesterol [159], thereby altering the cell membrane and inhibiting virus fusion, including HIV-1 [160]. It would be of interest to examine a possible role of this innate immune pathway in inhibiting HIV-1 trans infection.
Mature DC concentrate captured HIV-1 in late endosomal vesicles containing CD63 and CD81 tetraspanins, which are not found in immature DC. In fact, it is apparent that HIV-1 particles colocalize within DC in tetraspanin-rich compartments that include CD9, CD63, CD81, and CD82 [161, 162]. Some of these tetraspanins, particularly CD63, are carried in the viral membrane upon budding from the DC. These data point to the well-documented role of tetraspanins in cis infection of T cells [163, 164] and support a role for tetraspanin-rich microdomains in HIV-1 trans infection via egress of virus from DC to T cells.
4.2. DC-SIGN Receptor
MDDC express the CLR DC-SIGN, which is a major receptor for HIV-1 cis and trans infection [96]. Thus, HIV-1 trans infection by MDDC can be blocked by antibodies to DC-SIGN [96], mannose-binding lectin [165], and interfering RNA specific for DC-SIGN [166, 167]. Virus is being transmitted across a virologic synapse between the DC and T cell after cell-to-cell contact [168]. This unique discovery opened a new, non-CD4/coreceptor dependent pathway for HIV-1 cis and trans infection of T cells by passaging through DC. DC-SIGN-expressing DC retain the ability to trans infect T cells for several days and traffic towards M-tropic (R5) but not T-tropic (X4) HIV-1 [169].
The end result of these early studies was still the finding of poor cis replication of either R5 or X4 HIV-1 in DC. Instead of the conventional fusion and entry by CD4/coreceptors in T cells, carbohydrate regions of gp120 bind to DC-SIGN [170], which is in a tetrameric form [171]. A combination of N-linked glycosylation sites on gp120 are involved in HIV-1 binding to DC-SIGN [172, 173]. Notably, DC-SIGN also functions as a pattern recognition receptor for nonself glycans [174]. Virus entry into DC via DC-SIGN or other CLR does not lead to processing of HIV-1 antigen for MHC class I or II presentation [175]. Internalization motifs on the cytoplasmic tail of DC-SIGN trigger endocytosis and uptake of virus, which ends up in low pH nonlysosomal compartments. Leukocyte-specific protein 1 binds to the cytoplasmic region of DC-SIGN and directs internalized virus to the proteasome [176]. Importantly, downregulation of DC-SIGN by microRNA-155 indicates that this CLR is under tight host control that could affect binding of gp120 and uptake of virus [177].
Glycosylation sites on gp120 are key to binding to DC-SIGN, and thus viruses differing in the V1V2 and V3 variable loops of gp120 are differentially captured by DC-SIGN expressing cells [178]. In fact, R5 virus from patients with end-stage AIDS that does not undergo the switch to X4 virus has reduced ability to bind DC-SIGN [179]. This is due to a lack of potential N-linked glycosylation sites in the gp120 in these late-stage R5 variants. Following entry, HIV-1 sequesters in DC and remains available in an infectious form for trans infection [180, 181]. In fact, MDDC can support infectious R5 virus [182] but not X4 virus [183] for several weeks in culture.
The efficiency of cis infection of DC and subsequent trans infection also involves TLR8 signaling stimulated by HIV-1 encoded ssRNA and DC-SIGN signaling stimulated by gp120 [184]. This initiates transcription of integrated proviral DNA by RNA polymerase II and generation of full length viral transcripts.
R5 viruses are preferentially transmitted to T cells by DC [185], which is not blocked by anti-HIV-1 neutralizing antibodies [186]. Likewise, antibody neutralized, dual-tropic HIV-1 is captured by DC-SIGN and Fc receptors on immature MDDC and can trans infect T cells [187]. Of significance is that trans infection can occur in both activated and resting T cells [188] but requires activated T cells for extensive virus replication. Indeed, trans infection is enhanced during antigen-specific activation of T cells by DC, leading to selective destruction of anti-HIV-1 T cells [189]. Further evidence of the importance of DC-SIGN to trans infection is the enhanced binding to DC-SIGN by complement-opsonized virus, leading to its uptake and trans infection of T cells by immature MDDC [190]. Of note is that uptake of complement-bound virus also involves several integrins [191].
4.3. Dual Pathways of HIV-1 Cis Infection of DC Affect Trans Infection
Data from the earliest investigations of DC-SIGN-expressing MDDC began to clarify our understanding that there are both nonproductive and productive pathways of HIV-1 cis infection of DC that determine the efficiency of trans infection [192]. The bulk of the evidence supports that there are 2 parallel, distinct pathways that have different temporal phases within the DC. The first 24-hour phase of HIV-1 infection is a nonproductive pathway following binding to DC-SIGN and endocytosis. Initiation of this process is enhanced by direct contact of DC with HIV-1 infected cells [124] via HIV-1 Env-mediated virion endocytosis [193], which in this case may not involve DC-SIGN [194]. Endocytosis of cell-associated HIV-1 by DC involves activity of matrix metalloproteinase-9 [195], a collagenase that could be involved with cytoskeletal movements during endocytosis.
HIV-1 infection of DC by the endocytic process is resistant to proteolysis by trypsin [196]. Both viral endocytosis and subsequent trans infection by the DC can be decreased by HIV-1 protease inhibitors [197]. Virus accumulates in nonlysosomal, mildly acidic, endocytic compartments that are rich in DC-SIGN and conducive to maintaining infectivity [198, 199]. Virus is transferred to a virologic synapse formed between the DC and the CD4+ T cell (Figure 3).
Figure 3.
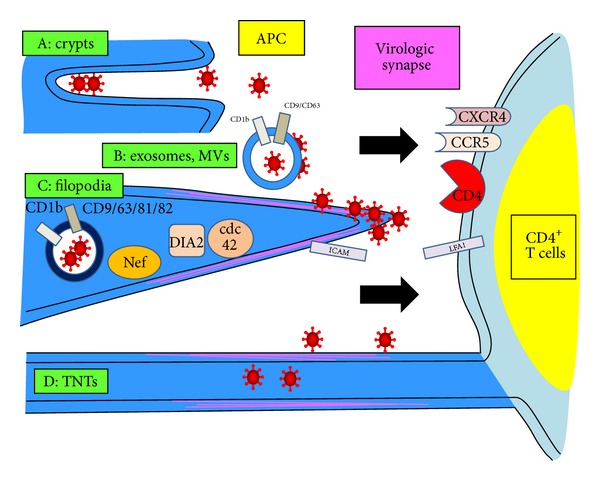
APC crypts, exosomes/microvesicles (MVs), filopodia, and tunneling nanotubes (TNTs) are associated with HIV-1 trans infection of T cells. HIV-1 infected APCs enhance trans infection of T cells by transferring (a) infectious virus within crypts (extracellular invaginations), (b) release of infected exosomes that express CD1b and tetraspanins, and transporting virus harbored within endosomal MVs expressing CD1b and tetraspanins; (c) viral Nef and cellular dia2 and cdc42 proteins rearrange actin filaments in the infected APC membrane to form filopodia; virus buds from the tips to form the virus synapse with the T cells, which involves ICAM/LFA-1 adhesion molecules and CD4/CCR5 (CXCR4) viral receptors. (d) Virus is transported through within TNT conduits and on the outside (“surfing”) of TNTs directly into T cells.
The second phase of infection is where a smaller fraction of HIV-1 enters DC by CD4 receptor/coreceptor-mediated membrane fusion [200]. DC-SIGN could also play a role in this process by interacting with gp120 and thereby increasing the Env binding site for CD4 [201]. By 2 days, this nonendosomal virus undergoes productive, complete cycle replication that is also transmitted via a virologic synapse to T cells.
HIV-1 is therefore transmitted from DC to T cells in two time-dependent phases. This explains how a higher expression DC-SIGN on immature MDDC resulted in non-productive, cis infection [202], yet these MDDC could transfer virus efficiently to T cells. HIV-1 infected, mature MDDC also activate CD4+ T cells, facilitating the process of trans infection [69]. Further distinguishing the two phases of HIV-1 infection in DC is that they differ in mRNA expression, particularly in decreased cathepsin activity during the replicative virus pathway [203]. This could enhance HIV-1 replication by avoiding lysosomal enzymatic degradation.
The endocytosis pathway for trans infection from DC to T cells has been challenged [204]. Thus, the Raji B cell line engineered to express endocytosis-defective DC-SIGN is as effective at mediating trans infection as cells expressing wild-type DC-SIGN [205]. Moreover, immature MDDC required virus fusion and production of infectious virions to trans infect T cells. Others have shown that mature DC concentrate virus particles in nonendosomal, intracellular compartments [206] or on the surface of DC [207].
An intriguing offshoot of the HIV-1-DC endocytic trans infection pathway is that infectious virus particles that are taken into endosomes can be exocytosed and productively infect T cells as an alternate trans pathway [208]. Similar results for efficient trans infection not requiring intact DC have been reported using fractionated membranes or microsomes from HIV-1-infected, dead DC and other cell types [209]. The exosomes express HLA-DR1, MHC-like transmembrane glycoprotein CD1b, and transpanins CD9 and CD63, which are incorporated upon release of virus from the endosomes. Moreover, uptake of such exosomes is enhanced in mature DC, which can then trans infect T cells in a unique manner independent of HIV-1 envelope glycoprotein [210].
The vagaries of the in vitro DC-T cell model used to assess HIV-1 trans infection are evident when comparing these latter results of efficient infection via cell-associated and exosome-associated virus to whole, apoptotic T cells. Thus, feeding of apoptotic, uninfected, activated CD4+ T cells to DC can inhibit both HIV-1 cis and trans infection [211]. While these apoptotic cells actually mature the DC, they also induce virus inhibitory APOBEC3G.
4.4. Non-DC-SIGN Receptors
Antibodies to DC-SIGN and DC-SIGN ligands do not always block cis infection of DC [202], nor do they always block trans infection [212]. Thus, HIV-1 trans infection is not consistently linked to DC-SIGN expression [213–216]. Indeed, circulating blood myeloid DC express little DC-SIGN yet can trans infect T cells [99]. This suggests that HIV-1 trans infection of CD4+ T cells can occur via DC-SIGN-independent mechanisms. Most of these molecules, however, work similar to DC-SIGN by binding carbohydrate residues on gp120, concentrating virus on the cell surface and enhancing virus uptake by DC. Thus, other CLRs can be involved in DC-mediated trans infections. These include DC immunoreceptor (DCIR) [217], which is expressed on all DC subsets [218]. However, the role of DCIR in trans infection is complicated by findings that DCIR expression is induced in CD4+ T cells by HIV-1 infection and that this is also evident as a paracrine-like effect in uninfected CD4+ T cells [219]. Moreover, these DCIR-expressing T cells exhibit enhanced replication of HIV-1. The virus acts on tyrosine and threonine residues of the DCIR immunoreceptor tyrosine-based inhibitory motif, associated with activation of various kinase pathways [220].
Syndecan-3, a DC-specific heparan sulfate proteoglycan, captures HIV-1 through binding of gp120 and thereby enhances both cis and trans infection by MDDC [221]. Surfactant protein A, a collectin family molecule involved in pathogen recognition in the lungs, amniotic fluid, and vaginal tract, enhances HIV-1 trans infection at low, acidic pH similar to the vaginal tract [222]. Finally, the glycosphingolipid galactosyl ceramide (GalCer) is a mucosal epithelial receptor for HIV-1 that binds gp41 and is also expressed on immature MDDC and primary myeloid from blood and mucosal tissue [223]. Blocking with anti-GalCer mAb inhibits HIV-1 trans infection.
5. Role of CD4+ T Cells in DC-Mediated HIV-1 Trans Infection
5.1. CD4+ T Cell Subsets
The efficiency of the APC-to-T cell trans infection pathway is dependent on the type and metabolic state of the CD4+ T cell. There is a complexity in how CD4+ T cells relate to trans infection based on their tissue location and activated, inflammatory state, which has received little attention. CD4+ T helper (TH) cells come in many varieties ranging from proinflammatory (TH1) to anti-inflammatory (TH2) and beyond [224]. Similar to T cell plasticity involved in anticancer immunity [225], the subset and functional capacity of TH cells in virus infection can vary as to the organ tissue and pathogen encountered. Recently it was reported that follicular memory CD4+ T cells defined as CXCR5+PD-1+Bcl-6+ are a major compartment for HIV-1 infection in viremic hosts [226]. The intimate interaction in lymph nodes of APC with follicular CD4+ T cells [227] points to these being major targets for trans infection. These tissue T cells secrete IL-21 that regulates T cell, B cell, and NK cell differentiation and function [228].
Eradication of HIV-1 infected T cells could require recognition and lysis by cytotoxic T lymphocytes (CTLs) [229]. Here we enter the complex world of HIV-1 cis infection of professional APC that is required for induction and maintenance of anti-HIV-1 CTL, as well as trans infection of CD4+ T cells. Indeed, a new, potential aspect of trans infection of CD4+ T cells that should be explored involves its role in the quest for a true or functional cure of HIV-1 infection [230, 231]. ART and immunotherapies that inhibit HIV-1 cis infection may not inhibit HIV-1 trans infection. Different CD4+ T cell subsets, in particular the resting central and transitional memory subsets known to harbor the latent HIV-1 reservoir during ART [232], could be involved in HIV-1 trans infection. Stealth, low level passage of virus between APC and these T cells in lymph nodes and other tissues could be a source of virus persistence in the face of ART.
5.2. Membrane-to-Membrane Contact Is Essential for Trans Infection
Direct contact of infected DC with CD4+ T cells forms a virologic synapse that is central to trans infection [233]. Several elegant microscopy studies have shown that infection of DC by HIV-1 induces dramatic changes in the actin cytoskeleton that enhance DC-T cell interaction [234]. Thus, HIV-1 infection of immature DC activates the membrane cell division control protein 42 homolog which is a regulator of actin polymerization, leading to filopod extensions of the DC cytoplasmic membrane [235]. These filopodia are also dependent on Nef and the formin protein diaphanous 2 that induce polymerization of actin [236]. The filopodia bear a large amount of budding HIV-1 particles on the tips of their long extensions [236]. Filopodia also protrude from CD4+ T cells and express CD4 that is involved in transfer of HIV-1 virions from DC [237].
5.3. T Cell Activation Is Essential for Trans Infection
Highly productive HIV-1 trans infection requires activated CD4+ T cells. Indeed, many studies have used polyclonally activated, allogeneic T cells and continuous T cells lines which presumably undergo further DC-mediated allostimulatory activation. Under conditions of natural infection within the host, HIV-1 effectively takes advantage of the immunologic synapse, that is, MHC-antigen-T cell receptor complex, to form a virologic synapse. This is supported by mathematical models that recapitulate the dynamics of HIV-1 infection, where simultaneous priming and infection of T cells by DC is predicted to drive early HIV-1 infection [238]. Alternatively, the virologic synapse could form by interactions among roaming T cells and tissue-resident or blood DC, without an antecedent, antigen recognition, DC-T cell interaction. Simple contact of uninfected, immature DC can augment HIV-1 replication in infected, resting CD4+ T cells [239]. This occurs in the absence of nominal antigen presentation. However, the basis of this phenomenon is unclear. For example, DC without nominal antigen do not alter HIV-1 proviral DNA in resting CD4+ T cells [240]. Moreover, many of the T cells within the DC-T cell clusters die by apoptosis [241].
DC infected with HIV-1 can express nominal viral antigens that prime T cells in vitro. However, this immunologic process activates the P38MAPK/STAT3 pathway resulting in upregulation of T cell inhibitory molecules in the DC including cytotoxic T-lymphocyte antigen (CTLA-4), TNF-related apoptosis-inducing ligand (TRAIL), lymphocyte-activation gene-3 (LAG3), and T-cell Ig mucin-3 (TIM-3) [242]. Thus, it is possible though not yet proven that during T cell priming by infected DC, HIV-1 trans infection could be diminished.
5.4. Central Role of the Virologic Synapse in Trans Infection
The virologic synapse is where critical virus transmission events occur between the DC and T cell involving conventional HIV-1 CD4 and chemokine receptors on the T cells. Thus, trans infection is accentuated by interaction of various cell adhesion molecules such as LFA-1/ICAM-1 [243] which are upregulated on activated mature DC and are involved in the DC-T cell virologic synapse [137] (Figure 3). HIV-1 trans infection is impaired in MDDC from patients with leukocyte adhesion deficiency type 1 whose T cells lack expression of LFA-1 [244]. There are contradictory data, however, that not all LFA-1 ligands play this role, as ICAM-1 [243] and ICAM-3 [245] have not been confirmed to be involved in HIV-1 trans infection. However, additional evidence that such binding is important in trans infection is that it can be prevented by inhibitors that block the virus-cell fusion process, for example, CD4 attachment blockers (mAb), fusion blockers [T20], and chemokine receptor blockers (RANTES) [246, 247].
Various intracellular processes are operative in forming the DC-T cell virologic synapse that is required for efficient trans infection. Activation of Rho guanine nucleotide-exchange factor LARG in DC after HIV-1 infection is one such factor involved in formation of virus-T cell synapses [248]. During the DC-T cell adhesion process, HIV-1 translocates across the synapse to form virus particles in intracellular compartments in endocytic vesicles of the T cells [2]. Although there is enhanced replication of R5 virus in this microenvironment [185, 249], both R5 and X4 viruses are transmissible to the T cells.
Importantly, the CD4+ effector-memory T cell (Tem) subset is most susceptible to trans infection by R5 HIV-1, which is related to expression of the CCR5 coreceptor [250]. In contrast, CD4+ naïve T cells (Tn) are more susceptible to trans infection with X4 virus in relation to expression of CXCR4.
The efficiency of trans infection is further dependent on expression of HIV-1 nef [233, 251]. Nef enhances this process by promoting HIV-1 cis infection of immature DC and recruitment of T cells to DC [252]. Nef heightens HIV-1 trans infection by upregulating DC-SIGN expression on DC and promoting DC-T cell clustering [146], while enhancing activation and proliferation of CD4+ T cells [253]. However, recent work has found that Nef induces tetherin, a type 2 integral membrane protein that can inhibit budding of virus in HIV-1-infected immature DC [254]. Thus, there appear to be competing inhibitory and enhancing effects of Nef on HIV-1 cis infection of DC that need to be clarified.
5.5. Neutralizing Antibodies Can Enhance Trans Infection
In contrast to early studies suggesting that trans infection could be blocked by neutralizing antibodies [255], later investigations showed that HIV-1 trans infection was protected from such effects [187]. In fact, neutralizing antibodies enhance the ability of both immature and mature MDDC to trans infect T cells with X4 virus [256], suggesting that this process could be involved in the switch from R5 to X4 that occurs during natural, in vivo HIV-1 infection. These seemingly conflicting results could be related to the viral target of the antibody. Thus, neutralizing antibody directed against gp41 is much more effective at blocking trans infection than that directed to gp120 [257]. This is postulated to be due to binding of gp41-specific antibody to the DC plasma membrane prior to the formation of a virologic synapse, positioning them to inhibit virus transfer.
6. NK Cells and DC-Mediated HIV-1 Trans Infection
NK cells have bidirectional interactions with DC where they promote maturation of DC and production of cytokines that activate T cells [258, 259]. This process could therefore bear on HIV-1 trans infection. Indeed, NK cells drive DC maturation [260]. As discussed earlier, mature DC mediate enhanced HIV-1 trans infection of T cells. Some NK cells express CD4 and are susceptible to HIV-1 cis infection. Infection of NK cells by HIV-1 has been controversial, but accumulating data support that a CD4-expressing NK cell subset is permissive for HIV-1 replication [261]. Moreover, HIV-1 infected DC are resistant to NK cell-mediated killing or “editing” due to enhancement of antiapoptotic factors in the DC [262]. This could thereby indirectly augment HIV-1 trans infection mediated by DC. Additional studies are needed to better define NK cell interactions with DC and T cells in relation to HIV-1 trans infection.
7. Coinfections and DC-Mediated HIV-1 Trans Infection
7.1. Bacteria
Given the importance of DC maturation and T cell activation in both cis and trans infection, it is logical that coinfections could affect these processes. This has been studied in the context of an enhancing role of genital coinfections on HIV-1 trans infection in sexual transmission. Bacterial vaginosis is a mildly symptomatic condition, for example, discharge, odor, and pain, occurring when the predominant commensal lactobacilli in the vaginal tract are largely replaced by Gram negative or positive bacteria [263]. It is associated with increased risk for HIV-1 infection [264]. Mucosal fluids from women with bacterial vaginosis activate and mature MDDC [265], although a more recent study has failed to show that this leads to enhanced HIV-1 trans infection [266]. Nevertheless, maturation of genital tract DC could be due to the effects of Gram negative bacterial LPS [135], which also enhances their ability to mediate HIV-1 trans infection [131, 267]. This likely occurs at least in part through binding of LPS to TLR4 [268]. TLR2 triggering of DC, which includes bacterial peptidoglycan and lipoproteins ligands, also enhances HIV-1 trans infection [269]. Finally, some combinations of oral Gram negative bacteria activate the HIV-1 promoter and viral replication in immature DC as well as macrophages [270]. This supports a potential role for polybacteria activating HIV-1 infection in the oral cavity.
A second context of bacterial coinfection enhancing HIV-1 trans infection is through translocation of bacterial products across the gastrointestinal (GI) tract to the blood, which leads to systemic immune activation. Indeed, there is extensive evidence for early, enhanced HIV-1 replication in resident CD4+ T cells of the GI tract, with lysis of the infected cells and associated disruption of the epithelial-blood barrier [271]. Lamina propria CD4+ T cells are under a steady state of activation and express the primary CD4 receptor and CCR5 and α4β7 coreceptors, making them fertile ground for either cis or trans infection. Moreover, recent studies have revealed that there is selective destruction of the Th17 cell subset of CD4+ T cells in the GI tract [272]. These cells produce IL-17 as well as IL-22 and IL-26 and are important in antimicrobial defense. Lamina propria mononuclear cells including CD1c+ DC are also constitutively activated in relation to chronic TLR signaling by microbial products [273]. These DC enhance productive infection of Th17 cells after activation by E. coli [274]. This is related to MHC class II-restricted Ag presentation to the T cells by the DC.
Tuberculosis is the leading cause of death in HIV-1 infected persons [275]. Although macrophages are the APC primarily targeted by Mycobacterium tuberculosis (MTB), infection of DC is also notable [276]. MTB binds to DC-SIGN by mannose-capped lipoarabinomannan in its cell wall [277]. Soluble factors produced by MTB-infected macrophages lead to partial maturation of DC, with further production of immune mediators by the DC [278]. This macrophage-DC inflammatory milieu is associated with increased HIV-1 trans infection by the DC. Microscopy studies indicate that MTB-HIV-1 coinfection of DC decreases degradation of the virus, resulting in greater sequestration of HIV-1 in the intracellular trans infection pathway [279].
7.2. Fungi
Several diverse types of fungi induce maturation of DC and coincidental enhancement of HIV-1 trans infection. Thus, infection of MDDC with Candida albicans augments HIV trans infection of T cells in relation to increases in expression of CD80 and CD86 on the DC [280]. Cryptococcus neoformans and Penicillium marneffei, both of which can be lethal to HIV-1 infected persons, enhance DC-mediated HIV-1 trans infection in association with increases in DC-T cell conjugates and CD4+ T cell activation [281, 282].
7.3. Parasites
Coinfection of malarial parasites with HIV-1 is common in coendemic areas [283]. Infection with Plasmodium falciparum increases HIV-1 viral load and enhances progression to AIDS [284]. The enhancing effect of malaria on DC-mediated trans infection could be due to hemozoin, a pigment that is released in excess after destruction of the infected erythrocytes. The hemozoin-loaded DC are partially activated, leading to increased activation and trans infection of CD4+ T cells [285, 286].
In contrast to enhancement of HIV-1 trans infection by malarial parasites, leishmania parasites inhibit trans infection by DC [287]. The mechanism of this phenomenon is not clear, although it could be due to competition for binding to DC-SIGN.
8. Blocking of HIV-1 DC-Mediated Trans Infection by Microbicides and Antiviral Drugs
Although much of this review is premised on the hypothesis that DC serve as the major, initial sites of HIV-1 infection in the mucosa, it needs to be acknowledged that this is a controversial concept. Indeed, there is extensive evidence that CD4+ T cells and not DC are the first, primary targets in HIV-1 and SIV sexual transmission models [288]. Other studies, however, support an important role for DC as initial targets in sexual transmission [289] as well as other modes such as mother-to-child transmission [290].
HIV-1 captured on spermatozoa via heparan sulfate can efficiently cis infect MDDC in vitro [291]. Vaginal and ectocervical derived DC also serve as efficient targets for HIV-1 cis infection in vitro [292]. However, seminal plasma could be a natural barrier to HIV-1 trans infection, as it can inhibit this process in vitro, possibly by blocking HIV-1 binding to CLR on the DC by high-mannose N-linked carbohydrates [293]. Also, there is increasing evidence of the importance of cross-talk between non-DC mucosal cells in regulating DC mediated cis and trans infection. Thus, uterine epithelial cells secrete transforming growth factor β that decreases DC-SIGN expression on immature DC, consequently inhibiting trans infection by the DC [294]. In contrast, genital mucosal epithelial cells also produce thymic stromal lymphopoietin that activates DC [295]. This results in increased HIV-1 trans infection of activated CD4+ T cells.
DC infected at these mucosal transmission sites likely spread HIV-1 through trans infection of regional CD4+ T cells. However, systemic infection with HIV-1 could require trafficking of HIV-1 infected DC to draining lymph nodes. Recent evidence in a humanized mouse model indicates that HIV-1-infected CD4+ T cells are very motile, circulating to lymph nodes where they tether to other T cells to form virologic synapses that facilitate cell-to-cell transmission [296]. Similar studies need to be done to assess the role of DC and other professional APC in HIV-1 trafficking and dissemination.
Based on the premise that DC are early targets during sexual transmission of HIV-1, there has been a good deal of interest in assessing inhibition of HIV-1 trans infection by microbicides and antiviral drugs. Trans infection by both R5 and X4 viruses can be at least partially blocked by many different nonnucleoside reverse transcriptase inhibitors (NNRTIs) [297]. Others have reported that RTIs such as azidothymidine do not inhibit trans infection [196]. Both cis infection of DC and DC-mediated trans infection of T cells can be inhibited by protease inhibitor saquinavir [298]. Several HIV-1 entry inhibitors targeting Env (BMS-C, T-1249) or CCR5 (CMPD167) have been shown to block cis infection of DC [299].
Various experimental microbicides act through different inhibitory pathways to decrease HIV-1 cis and trans infection, some of whose mechanisms of action are still unclear. These include amphibian-derived antimicrobial peptides [300], HIV-1 integrase strand transfer inhibitors [301], Miltefosine, a phospholipid drug that induces type I IFN in DC [302], SAMMA, a polymer derived from mandelic acid [303], HIV-1 fusion inhibitor T-1249 [304], the nematode C-type lectin Mermaid, a structural homologue of DC-SIGN [305], sulfonated polyanion PRO 2000 [306], a soluble CD4-linker-DC-SIGN fusion protein [307], Dendron 12, a multimeric glycomimetic DC-SIGN ligand [308], Inmunoferon, a natural glycoconjugate [309], carbohydrate-binding plant lectins [310, 311], oligomannose glycans [312], trimannosides [313, 314], and advanced glycation end products [315].
9. HIV-1 Trans Infection Mediated by Plasmacytoid DC (pDC)
In 1999, Siegal et al. [316] defined a population of lymphoid-derived DC in blood, termed pDC, that were highly potent producers of IFN-α in response to HIV-1 and other viral infections. The pDC were distinct from myeloid DC in morphology and phenotype, clarifying previous work indicating that a natural IFN-producing cell with a dendritic cell phenotype was the major producer of IFN-α [317, 318]. The pDC differ phenotypically from myeloid DC primarily by lack of expression of CD11c and positive expression of the IL-3 receptor, CD123. It is now well established that pDC are central to innate immunity to viral infections through viral DNA and RNA signaling of TLR9 and TLR7, respectively. We also know that pDC have important yet enigmatic roles in both combating and enhancing HIV-1 infection [319].
It appears that pDC are less permissive for productive, cis infection with HIV-1 than are myeloid DC, even though they both express CD4 and HIV-1 coreceptors [86, 320]. Nevertheless, several studies indicate that pDC derived from blood [95, 321] and thymus [322, 323] can support productive HIV-1 infection with X4 or R5 strains, particularly if the cells are prestimulated with CD40L [324]. Moreover, although pDC do not express DC-SIGN in vitro [98], there is evidence of expression of this CLR on pDC subsets in vivo [97].
Given these data on expression of HIV-1 receptors and productive cis infection of pDC, it is not surprising that pDC can trans infect T cells with R5 and X4 HIV-1 [95, 324, 325]. This infection preferentially targets antigen-responding CD4+ T cells [325]. Interestingly, productive cis infection of the pDC is not required for trans infection of T cells with HIV-1 [325].
The ability of pDC to mediate trans infection with HIV-1 has been challenged by work showing that, regardless of their state of activation, pDC derived from the same donors as myeloid DC did not trans infect their T cells [326]. This is explained at least in part by the antiviral effects of the prodigious amounts of IFN-α produced by the pDC. Clearly more information is needed to define whether pDC are able to trans infect T cells with HIV-1.
10. HIV-1 Trans Infection Mediated by Monocytes and Macrophages
Circulating monocytes and their tissue-transformed macrophages are APC that can mediate HIV-1 trans infection of CD4+ T cells similar to DC [13]. This was first reported in 1990 [327], where blood monocyte-derived macrophages (MDMs) transmitted HIV-1 to peripheral blood lymphocytes, presumably CD4+ T cells. Two basic principles of HIV-1 trans infection were established with this pioneering study, that is, that antigen-specific activation of the T cells results in greater trans infection by the MDM and that anti-CD4 and anti-gp120 antibodies block trans infection. It took years for this phenomenon to be extended, as in 1999 Carr and colleagues [328] reported that MDM transmitted HIV-1 to CD4+ T cells much more efficiently than cis infection of T cells. This process required cell contact, occurred in both allogeneic and autologous systems, and was enhanced by preactivation of the T cells. Overall, trans infection mediated by MDM appears as efficient as that mediated by myeloid DC.
10.1. Cis Infection
10.1.1. HIV-1 R5 and X4 Receptors on Monocytes/Macrophages
As with DC, the efficiency of HIV-1 trans infection mediated by monocytes and macrophages is largely dependent on the pathways and proficiency of preceding cis infection in these APCs. Given that blood-derived monocytes/macrophages express CD4 and CCR5, but not CXCR4, they are highly susceptible to R5 HIV-1, albeit with much lower production of infectious virus than cis infection of CD4+ T cells [329]. However, some X4 variants are able to infect monocytes/macrophages. The efficacy of HIV-1 infection largely depends on variations in binding capacity of HIV-1 Env to these coreceptors [330]. It is also apparent that, as with tissue DC and LC, subsets of tissue resident macrophages express both HIV-1 coreceptors in vivo [62, 331, 332]. Tissue macrophages, including those within the genital mucosa and foreskin, are consequently potentially susceptible to cis infection with either R5 or X4 strains of HIV-1 [17, 41, 44, 292]. HIV-1 infection of monocytes/macrophages is further complicated by evidence that circulating monocytes in HIV-1 infected persons harbor many different phenotypes of HIV-1 [333]. These include HIV-1 variants that use multiple combinations of coreceptors that differ from virus in the corresponding CD4+ T cells.
Interestingly, semen contains enhancers of HIV-1 cis replication. For example, prostatic acidic phosphatase (PAP) in seminal fluid forms amyloid fibrils that enhance HIV-1 infection of macrophages [334].
The overall outcome is that cis replication efficiency, that is, production of infectious virus, varies greatly among both R5 and X4 HIV-1 strains but is relatively poor in monocytes/macrophages compared to T cells [335]. This is based on the well-documented yet still controversial process of HIV-1 replication in monocytes/macrophages compared to CD4+ T cells. In contrast to CD4+ T cells, HIV-1 resides and accumulates in specialized compartments within monocytes/macrophages, where new virions form and are eventually released to the extracellular environment [336].
10.1.2. TLR Triggering in Monocytes/Macrophages
Monocytes and macrophages express a variety of TLRs depending on their blood and tissue distribution [337]. These can serve as triggers for monocyte/macrophage activation and, hence, affect HIV-1 cis infection. Thus, the TLR2 ligand PAM3CSK4, a synthetic triacylated lipopeptide mimic of the acylated amino terminus of bacterial lipopeptide, activates HIV-1 transcription in macrophages [338]. This effect can be counteracted and suppressed by nuclear receptor signaling through, for example, the glucocorticoid receptor. Such nuclear receptor signaling also inhibits the proinflammatory response induced by PAM3CSK4 that activates HIV-1 expression in macrophages. Furthermore, HIV-1 may code for its own activation in macrophages, as suggested by single-stranded RNA of HIV-1 inducing uridine-rich TLR 7/8 ligands that stimulate MyD88-dependent monocyte activation [337].
10.1.3. Regulation of Cis Infection in Monocytes/Macrophages
Monocytes are more resistant to productive HIV-1 infection than macrophages, which has been associated with presence of apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3A (APOBEC3A) [339]. This cytidine deaminase may inhibit HIV-1 replication in monocytes through an IFN-α activated deamination of viral DNA [340]. Also, recent genome-wide scanning has revealed that expression of the gene DYRK1A, encoding a kinase that phosphorylates serine and threonine residues, is associated with efficient cis replication of HIV-1 in MDM [341].
Macrophages have been divided into M1 and M2 subsets distinguished by their polarized function, with M1 being activated by proinflammatory factors such as IFN-γ, TNF-α, and LPS, and M2 being activated by anti-inflammatory agents including IL-4, IL-10, and IL-13 [342]. Polarizing cytokines affect HIV-1 replication efficiency in macrophages; IFN-γ and TNF-α induce activation of the M1 subset of MDM, with downregulation of CD4 and consequent resistance to HIV-1 infection [343]. IFN-γ-activated M1 cells can, however, maintain low levels of both R5 and X4 HIV-1 DNA and low virus replication [344]. Regarding M2 macrophages, IL-4-activated M2 cells exhibit resistance to HIV-1 replication at a later, postentry stage. There also are a number of other host proteins and microRNAs known to regulate various stages of HIV-1 cis infection of monocyte/macrophages that are beyond the scope of this review [345].
An exciting, recently discovered mechanism by which macrophages restrict HIV-1 replication is that they express the dideoxynucleotide hydrolase SamHD1 [346, 347]. The actual mechanism of action is not fully clear but involves inhibition of reverse transcription of HIV-1 DNA. Furthermore, the IFN-inducible protein viperin (virus inhibitory protein, endoplasmic reticulum-associated, interferon-inducible) that was first discovered in macrophages [348] inhibits HIV-1 replication [349]. All of these host factors are likely contributing to the low level replication and persistence of HIV-1 infection, as well as its relative noncytopathic effect, in monocytes/macrophages.
Finally, the phenomenon of autophagy has in recent years been shown to be central to HIV-1 cis infection of monocytes/macrophages and therefore likely to affect trans infection of T cells. Autophagy is a catabolic process where cellular components are degraded by lysosomal machinery and thereby has a critical role in antigen processing by APC [350]. Although autophagy also functions as an intracellular host defense mechanism against viruses, it can be usurped by HIV-1 infection, particularly in macrophages and CD4+ T cells [351]. Whereas autophagy is enhanced in bystander, uninfected CD4+ T cells by X4 or R5 Env, leading to apoptosis, uninfected macrophages are resistant to this process [352]. In HIV-1-infected macrophages, the effects of autophagy are more complicated. In fact, early, nondegradative steps in autophagy are required for efficient HIV-1 infection, whereas later, degradative steps are detrimental to HIV-1 replication in macrophages [352, 353].
10.2. Trans Infection
Monocytes and MDM normally do not express DC-SIGN in vitro [96], thus negating this pathway in HIV-1 cis and trans infection. However, MDM treated in vitro with IL-13 [354, 355] or IL-4 [356, 357] express DC-SIGN and, hence, can mediate HIV-1 trans infection. M2a MDM derived by IL-4 treatment express DC-SIGN that facilitates cis infection with both R5 and X4 HIV-1 and subsequent efficient trans infection of T cells [358]. In contrast, M1 MDM derived by TNF-α and IFN-γ treatment express little DC-SIGN and do not efficiently trans infect T cells.
MDM also express the C-type lectin mannose-binding receptor (macrophage mannose receptor (MMR); CD206) which binds mannosylated carbohydrates on gp120 with high efficiency [359]. Virus binding is consequently inhibited by carbohydrate-binding agents such as mannan, D-mannose, EDTA, and soluble mannose-binding lectin [359], as well as antibody specific for MMR [360].
Of particular pertinence to HIV-1 trans infection is that certain resident tissue macrophages [97, 361] express DC-SIGN, supporting their potential for mediating HIV-1 trans infection. It has been suggested that placental CD163+ macrophages (Hofbauer cells) are infected by HIV-1 via their DC-SIGN receptors and that this could represent a pathway for mother-to-child transmission of HIV-1 [362]. However, MMR and not DC-SIGN expression is associated with HIV-1 trans infection of T cells by IL-13-treated MDM that express both [354]. Interestingly, IL-13 also inhibits cis replication of HIV-1 in MDM [363]. This suggests that trans infection mediated by MDM involves other nonreplicative pathways of HIV-1 in these APC. This concept fits with the extensive data that HIV-1 replicates relatively poorly in MDM and tissue resident macrophages [13, 364]. In this regard, the scavenger receptor cysteine-rich protein gp340, which is expressed by macrophages in vivo and MDM in vitro, is associated with HIV-1 cis infection of MDM and their capacity to mediate trans infection [365].
Given the ability of MMR to bind HIV-1 with high efficiency, this pathway has been linked to trans infection of T cells [359]. Thus, HIV-1 trans infection mediated by MDM can be blocked by inhibitors of MMR binding. This process differs, however, from trans infection mediated by DC in that virus does not survive beyond 1 day after binding to MDM. Notably, this latter finding contrasts with evidence that HIV-1 survives for weeks within macrophages and can still trans infect T cells [366]. Regardless, HIV-1 trans infection mediated by monocytes/macrophages is based largely on inefficient cis infection of these APC with HIV-1 virions assembling in late endosomes or multivesicular bodies. This involves tetraspanins including CD63 and CD81 that are constitutively expressed in these microvesicles. The virion-containing vesicles are important components in the trans infection T cell pathway, where they accumulate at the point of the virologic synapse. Efficient trans infection may also involve multiple points of cell contact, termed polysynapses, which are rosette-like structures of an infected macrophage surrounded by many T cells [367]. Finally, expression of the cell adhesion molecule sialoadhesin is upregulated by HIV-1 infection of monocytes, possibly through IFN-α, and is related to enhanced trans infection [368].
Using the HIV-1-infected MDM model, it is evident that movement of the vesicular body to the virologic synapse is dependent on Gag [369]. HIV-1 forming within these vesicles trans infects T cells within 6 hours of cell-to-cell contact [370]. Interestingly, neutralizing antibodies fail to block HIV-1 trans infection of T cells mediated by macrophages [371]. Hence, this could function to enhance HIV-1 pathogenesis.
Another intriguing physical pathway for HIV-1 trans infection of T cells by macrophages is through filopodial bridges or tunneling nanotubes [372]. Murine leukemia virus can hijack such filopodia that extend long distances between cells [373], suggesting that human retroviruses could use similar conduits. In fact, infection of macrophages with HIV-1 induces the formation of nanotubes that contain have HIV-1 particles [374]. HIV-1 can move rapidly between macrophages via endocytic compartments through these actin-rich, membranous nanotubes [375, 376]. Importantly, nanotube-like formation in HIV-1 infected macrophages is driven by Nef and occurs in both systemic and intestinal lymphoid follicles [377]. These actin-propelled nanotubes serve as conduits for transfer of Nef to B cells, where it inhibits immunoglobulin G2 (IgG2) and IgA class switching. It is not yet known if this is involved in macrophage-to-T cell HIV-1 trans infection.
10.3. HIV-1 Trans Infection in Human Colostrum and Milk
Neonatal infection with HIV-1 has largely been prevented by ART prophylaxis during pregnancy [378]. Nevertheless, HIV-1 can be transmitted from mother to child via infected colostrum and milk particularly in resource limited countries [379]. Human colostrum and milk contain macrophages and DC that express DC-SIGN, which therefore could mediate HIV-1 trans infection of CD4+ T cells that are also present [379]. Interestingly, human milk can inhibit HIV-1 trans infection mediated by DC [380]. This occurs by binding of a bile salt-stimulated lipase in the Lewis X (LeX) glycoprotein found in human milk to DC-SIGN on the DC [381]. Human milk can also block HIV-1 trans infection through functional natural IgA and IgG antibodies specific for the carbohydrate recognition domain of DC-SIGN [382]. Strategies to block HIV-1 trans infection in breast milk include stimulation of TLR3 expressed on breast milk macrophages with its double-stranded RNA ligand poly (I:C) [383]. This process may work through the inhibition of DC-SIGN expression by IFN-α and IFN-β that are induced by TLR3 activation.
11. HIV-1 Trans Infection Mediated by B Lymphocytes
Although B lymphocytes are highly activated and modified beginning in the earliest phases of HIV-1 infection, they are not considered a significant, major target for HIV-1 cis infection [384]. Indeed, B cells are difficult to infect with HIV-1 in that they do not express CD4 or CCR5, although they express CXCR4. Nevertheless, B cells derived either from lymphoid tissue or from the peripheral blood of HIV-1-infected persons carry replication-competent X4 or R5 virus [384, 385]. Moreover, binding of HIV-1 immune complexes to CR2 or CD21 on the surface of B cells has been linked to subsequent trans infection of T cells [386–390]. These early studies therefore established that there is transmission of HIV-1 from B cells to T cells involving virus trapped in immune complexes on the surface of the B cells. This is similar to the trapping of HIV-1-antibody complexes by Fc and complement receptors on follicular dendritic cells (FDC) that reside in follicles of secondary lymphoid tissues [391]. Virus infectivity can be maintained on the FDC for months. The FDC serve as a reservoir for HIV-1 that can be genetically diverse from virus in other body compartments [392]. CD4+ T cells trafficking through the lymphatics are infected by the FDC-bound virus, with virus replication being driven by TNF-α produced by the FDC [393].
In 2006, two laboratories discovered that DC-SIGN is expressed on peripheral blood and lymphatic tissue B cells [394, 395], in particular after stimulation with surrogates of activated CD4+ T cells, that is, CD40L and IL-4 [395]. This CLR acts as a portal for endocytosis of both X4 and R5 HIV-1 that does not involve complement receptors or immune complexes [395]. Soon after binding of virus to DC-SIGN, the CLR is internalized and temporarily absent from the B cell surface. This cis infection is non-productive, yet virus can retain its infectivity for at least 2 days within the B cells. Interestingly, most of the captured virions are destroyed in B cell lines engineered to express DC-SIGN [192, 216]. However, a portion of the input virus survives in these cells and can rapidly trans infect T cells [183, 192, 396].
Comparable to DC-SIGN-expressing B cell lines, activated blood and tissue-derived B cells are highly efficient in mediating HIV-1 trans infection of CD4+ T cells [395]. As with DC, HIV-1 trans infection mediated by B cells can be blocked with anti-DC-SIGN mAb as well as mannan. Although B cells express high levels of CXCR4, treatment with anti-CXCR4 mAb prior to addition of HIV-1 to B cells does not inhibit trans infection of the T cells. Thus, although cis infection of B cells does not involve productive replication of infectious virions like that of DC and monocytes/macrophages, B cells are as efficient as these other professional APC in mediating HIV-1 trans infection. This DC-SIGN dependent process of B cell mediated trans infection is not unique to HIV-1, as a similar pathway has been reported for hepatitis C virus trans infection of hepatoma cells [397].
CD40L is expressed on activated CD4+ T cells, which are abundant throughout chronic immune activation that is prevalent in HIV-1 infection. CD40 is a normal constituent on B cell surfaces. Thus, the continuous interaction of CD40L on activated T cells with CD40 on B cells is one pathway of chronic B cell stimulation during HIV-1 infection [384]. Intriguingly, the hyperactive state of B cells in HIV-1 infection has also been linked to CD40L incorporated in HIV-1 virions [398]. Such virions are found in plasma of untreated HIV-1 infected patients and induce Ig as well as IL-6 and other cytokines and chemokines in B cells [399]. Importantly, host-derived CD40L within virions results in increased virus attachment to B cells and more efficient B cell mediated trans infection of autologous CD4+ T cells compared to virions without CD40L [398].
An emerging concept in HIV-1 cis and trans infection is the role of cross-talk among DC, B cells and other cell types. Thus, HIV-1 cis infection of DC is enhanced by coculture with CD4+ T or B lymphocytes but not with CD8+ T cells [400]. In addition to potential involvement of CD40-CD40L-related B cell activation in this process, an unexplored area of HIV-1 cis and, hence, trans infection is the role of the tumor necrosis factor receptor superfamily [401]. Specifically, B cell activating factor (BAFF) and proliferation-inducing ligand (APRIL) could be central to HIV-1 interaction with B cells. Interestingly, these factors are produced by DC and function to drive B cell differentiation and survival by both T cell dependent and independent pathways. Thus, HIV-1 infection could act through DC to hyperactivate B cell responses [402], which in turn could affect both HIV-1 cis and trans infection mediated by B cells. Another area of potential interest regarding HIV-1 trans infection mediated by B cells is a possible role of TLR both in vitro and in vivo [403]. For example, autoimmune complexes, which are highly prevalent during HIV-1 infection, can stimulate B cells via TLR7 and TLR9 [404]. TLRs have been associated with B cell activation during HIV-1 infection [384]. These factors need to be assessed in B cell models of HIV-1 cis and trans infection.
B cells have also been implicated as intermediaries in enhanced HIV-1 infection of CD4+ T cells by macrophages [405]. This process involves activation of macrophages via Nef and CD40 ligation, leading to production of soluble CD23 and ICAM. These proteins act on B cells to upregulate expression of CD22, CD58, and CD80, which in turn stimulate CD4+ T cells via CD45, CD2, and CD28, respectively. The final result is enhanced HIV-1 replication in the T cells.
12. Concluding Remarks: Role of HIV-1 Trans Infection in HIV-1 Disease Progression
Despite years of extensive studies on HIV-1 trans infections, there are still major gaps in our understanding of this phenomenon. In particular, there is a paucity of information on the importance HIV-1 trans infection in natural HIV-1 infection and disease. There are inherent difficulties and highly challenging complexities in proving that this process is involved in virus transmission and disease pathogenesis. Nevertheless, studies of trans infection would be of interest in persons who control HIV-1 infection in the absence of ART, who range in definition and terminology as long-term nonprogressors, viral controllers, and elite controllers [406–408]. Myeloid DC from the blood of elite controllers have increased antigen presenting capacity and secrete less proinflammatory cytokines than HIV-1 infected progressors [409], which could affect both HIV-1 cis and trans infection. However, APC from these individuals have not been studied for their ability to trans infect T cells. We can speculate that the efficiency of this process will depend in part on the integrity of functional activity of the APC and CD4+ T cells, that is, their expression of receptors and intracellular and cell-cell pathways known to be involved in both HIV-1 cis and trans infections.
Conflict of Interests
The author reports no conflict of interests associated with this paper.
Acknowledgments
The author thanks Drs. Robbie Mailliard and Giovanna Rappocciolo, Taylor Poston, and Colleen Zaccard for review of this paper. Research in the authors' laboratory was supported by the National Institutes of Health, United States Public Health Service.
References
- 1.Mothes W, Sherer NM, Jin J, Zhong P. Virus cell-to-cell transmission. Journal of Virology. 2010;84(17):8360–8368. doi: 10.1128/JVI.00443-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hübner W, McNerney GP, Chen P, et al. Quantitative 3D video microscopy of HIV transfer across T cell virological synapses. Science. 2009;323(5922):1743–1747. doi: 10.1126/science.1167525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chen BK. T cell virological synapses and HIV-1 pathogenesis. Immunologic Research. 2012;54(1–3):133–139. doi: 10.1007/s12026-012-8320-8. [DOI] [PubMed] [Google Scholar]
- 4.Martin N, Welsch S, Jolly C, Briggs JAG, Vaux D, Sattentau QJ. Virological synapse-mediated spread of human immunodeficiency virus type 1 between T cells is sensitive to entry inhibition. Journal of Virology. 2010;84(7):3516–3527. doi: 10.1128/JVI.02651-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sattentau QJ. Cell-to-cell spread of retroviruses. Viruses. 2010;2(6):1306–1321. doi: 10.3390/v2061306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gupta P, Balachandran R, Ho M, Enrico A, Rinaldo C. Cell-to-cell transmission of human immunodeficiency virus type 1 in the presence of azidothymidine and neutralizing antibody. Journal of Virology. 1989;63(5):2361–2365. doi: 10.1128/jvi.63.5.2361-2365.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sigal A, Kim JT, Balazs AB, et al. Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy. Nature. 2011;477(7362):95–98. doi: 10.1038/nature10347. [DOI] [PubMed] [Google Scholar]
- 8.Sigal A, Baltimore D. As good as it gets? The problem of HIV persistence despite antiretroviral drugs. Cell Host & Microbe. 2012;12(2):132–138. doi: 10.1016/j.chom.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 9.Wilkinson J, Cunningham AL. Mucosal transmission of HIV-1: first stop dendritic cells. Current Drug Targets. 2006;7(12):1563–1569. doi: 10.2174/138945006779025482. [DOI] [PubMed] [Google Scholar]
- 10.Morrow G, Vachot L, Vagenas P, Robbiani M. Current concepts of HIV transmission. Current HIV/AIDS Reports. 2007;4(1):29–35. doi: 10.1007/s11904-007-0005-x. [DOI] [PubMed] [Google Scholar]
- 11.de Jong MAWP, Geijtenbeek TBH. Human immunodeficiency virus-1 acquisition in genital mucosa: langerhans cells as key-players. Journal of Internal Medicine. 2009;265(1):18–28. doi: 10.1111/j.1365-2796.2008.02046.x. [DOI] [PubMed] [Google Scholar]
- 12.McDonald D. Dendritic cells and HIV-1 trans-infection. Viruses. 2010;2(8):1704–1717. doi: 10.3390/v2081704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Waki K, Freed EO. Macrophages and cell-cell spread of HIV-1. Viruses. 2010;2(8):1603–1620. doi: 10.3390/v2081603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wonderlich ER, Barratt-Boyes SM. A dendrite in every pie: myeloid dendritic cells in HIV and SIV infection. Virulence. 2012;3(7):647–653. doi: 10.4161/viru.22491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Teleshova N, Derby N, Martinelli E, Pugach P, Calenda G, Robbiani M. Simian immunodeficiency virus interactions with macaque dendritic cells. Advances in Experimental Medicine and Biology. 2013;762:155–181. doi: 10.1007/978-1-4614-4433-6_6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Romani N, Clausen BE, Stoitzner P. Langerhans cells and more: langerin-expressing dendritic cell subsets in the skin. Immunological Reviews. 2010;234(1):120–141. doi: 10.1111/j.0105-2896.2009.00886.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mesman AW, Geijtenbeek TB. Pattern recognition receptors in HIV transmission. Frontiers in Immunology. 2012;3:p. 59. doi: 10.3389/fimmu.2012.00059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Harman AN, Bye CR, Nasr N, et al. Identification of lineage relationships and novel markers of blood and skin human dendritic cells. The Journal of Immunology. 2013;190(1):66–79. doi: 10.4049/jimmunol.1200779. [DOI] [PubMed] [Google Scholar]
- 19.Belsito DV, Sanchez MR, Baer RL, Valentine F, Thorbecke GJ. Reduced Langerhans’ cell Ia antigen and ATPase activity in patients with the acquired immunodeficiency syndrome. The New England Journal of Medicine. 1984;310(20):1279–1282. doi: 10.1056/NEJM198405173102002. [DOI] [PubMed] [Google Scholar]
- 20.Tschachler E, Groh V, Popovic M. Epidermal Langerhans cells—a target for HTLV-III/LAV infection. Journal of Investigative Dermatology. 1987;88(2):233–237. doi: 10.1111/1523-1747.ep12525402. [DOI] [PubMed] [Google Scholar]
- 21.Kanitakis J, Marchand C, Su H, et al. Immunohistochemical study of normal skin of HIV-1-infected patients shows no evidence of infection of epidermal Langerhans cells by HIV. AIDS Research and Human Retroviruses. 1989;5(3):293–302. doi: 10.1089/aid.1989.5.293. [DOI] [PubMed] [Google Scholar]
- 22.Kanitakis J, Thivolet J. Infection of human epidermal Langerhans’ cells by HIV. AIDS. 1990;4(3):266–268. [PubMed] [Google Scholar]
- 23.Kalter DC, Greenhouse JJ, Orenstein JM, Schnittman SM, Gendelman HE, Meltzer MS. Epidermal Langerhans cells are not principal reservoirs of virus in HIV disease. The Journal of Immunology. 1991;146(10):3396–3404. [PubMed] [Google Scholar]
- 24.Dusserre N, Dezutter-Dambuyant C, Mallet F, et al. In vitro HIV-1 entry and replication in Langerhans cells may clarify the HIV-1 genome detection by PCR in epidermis of seropositive patients. Journal of Investigative Dermatology. 1992;99(5):99S–102S. doi: 10.1111/1523-1747.ep12669977. [DOI] [PubMed] [Google Scholar]
- 25.Compton CC, Kupper TS, Nadire KB. HIV-infected Langerhans cells constitute a significant proportion of the epidermal Langerhans cell population throughout the course of HIV disease. Journal of Investigative Dermatology. 1996;107(6):822–826. doi: 10.1111/1523-1747.ep12330574. [DOI] [PubMed] [Google Scholar]
- 26.Henry M, Uthman A, Ballaun C, Stingl G, Tschachler E. Epidermal Langerhans cells of AIDS patients express HIV-1 regulatory and structural genes. Journal of Investigative Dermatology. 1994;103(4):593–596. doi: 10.1111/1523-1747.ep12396918. [DOI] [PubMed] [Google Scholar]
- 27.Dezutter-Dambuyant C. In vivo and in vitro infection of human Langerhans cells by HIV-1. Advances in Experimental Medicine and Biology. 1995;378:447–451. doi: 10.1007/978-1-4615-1971-3_100. [DOI] [PubMed] [Google Scholar]
- 28.Giannetti A, Zambruno G, Cimarelli A, et al. Direct detection of HIV-1 RNA in epidermal Langerhans cells of HIV-infected patients. Journal of Acquired Immune Deficiency Syndromes. 1993;6(4):329–333. [PubMed] [Google Scholar]
- 29.Cimarelli A, Zambruno G, Marconi A, Girolomoni G, Bertazzoni U, Giannetti A. Quantitation by competitive PCR of HIV-1 proviral DNA in epidermal Langerhans cells of HIV-infected patients. Journal of Acquired Immune Deficiency Syndromes. 1994;7(3):230–235. [PubMed] [Google Scholar]
- 30.McIlroy D, Autran B, Cheynier R, et al. Infection frequency of dendritic cells and CD4+ T lymphocytes in spleens of human immunodeficiency virus-positive patients. Journal of Virology. 1995;69(8):4737–4745. doi: 10.1128/jvi.69.8.4737-4745.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ludewig B, Holzmeister J, Gentile M, et al. Replication pattern of human immunodeficiency virus type 1 in mature Langerhans cells. Journal of General Virology. 1995;76, part 6:1317–1325. doi: 10.1099/0022-1317-76-6-1317. [DOI] [PubMed] [Google Scholar]
- 32.Dittmar MT, Simmons G, Hibbitts S, et al. Langerhans cell tropism of human immunodeficiency virus type 1 subtype A through F isolates derived from different transmission groups. Journal of Virology. 1997;71(10):8008–8013. doi: 10.1128/jvi.71.10.8008-8013.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zaitseva M, Blauvelt A, Lee S, et al. Expression and function of CCR5 and CXCR4 on human Langerhans cells and macrophages: implications for HIV primary infection. Nature Medicine. 1997;3(12):1369–1375. doi: 10.1038/nm1297-1369. [DOI] [PubMed] [Google Scholar]
- 34.Gorry PR, Ancuta P. Coreceptors and HIV-1 pathogenesis. Current HIV/AIDS Reports. 2011;8(1):45–53. doi: 10.1007/s11904-010-0069-x. [DOI] [PubMed] [Google Scholar]
- 35.Kawamura T, Gulden FO, Sugaya M, et al. R5 HIV productively infects langerhans cells, and infection levels are regulated by compound CCR5 polymorphisms. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(14):8401–8406. doi: 10.1073/pnas.1432450100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nestle FO, di Meglio P, Qin JZ, Nickoloff BJ. Skin immune sentinels in health and disease. Nature Reviews Immunology. 2009;9(10):679–691. doi: 10.1038/nri2622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Nestle FO, Zheng XG, Thompson CB, Turka LA, Nickoloff BJ. Characterization of dermal dendritic cells obtained from normal human skin reveals phenotypic and functionally distinctive subsets. The Journal of Immunology. 1993;151(11):6535–6545. [PubMed] [Google Scholar]
- 38.Summers KL, Hock BD, McKenzie JL, Hart DNJ. Phenotypic characterization of five dendritic cell subsets in human tonsils. American Journal of Pathology. 2001;159(1):285–295. doi: 10.1016/S0002-9440(10)61694-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Segura E, Valladeau-Guilemond J, Donnadieu MH, Sastre-Garau X, Soumelis V, Amigorena S. Characterization of resident and migratory dendritic cells in human lymph nodes. The Journal of Experimental Medicine. 2012;209(4):653–660. doi: 10.1084/jem.20111457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gibbings D, Befus AD. CD4 and CD8: an inside-out coreceptor model for innate immune cells. Journal of Leukocyte Biology. 2009;86(2):251–259. doi: 10.1189/jlb.0109040. [DOI] [PubMed] [Google Scholar]
- 41.Hirbod T, Bailey RC, Agot K, et al. Abundant expression of HIV target cells and C-type lectin receptors in the foreskin tissue of young Kenyan men. American Journal of Pathology. 2010;176(6):2798–2805. doi: 10.2353/ajpath.2010.090926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hirbod T, Kaldensjö T, Lopalco L, et al. Abundant and superficial expression of C-type lectin receptors in ectocervix of women at risk of HIV infection. Journal of Acquired Immune Deficiency Syndromes. 2009;51(3):239–247. doi: 10.1097/QAI.0b013e3181a74f89. [DOI] [PubMed] [Google Scholar]
- 43.Hirbod T, Kaldensjo T, Broliden K. In situ distribution of HIV-binding CCR5 and C-type lectin receptors in the human endocervical mucosa. PLoS One. 2011;6(9) doi: 10.1371/journal.pone.0025551.e25551 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kaldensjö T, Petersson P, Tolf A, Morgan G, Broliden K, Hirbod T. Detection of intraepithelial and stromal langerin and CCR5 positive cells in the human endometrium: potential Targets for HIV infection. PLoS One. 2011;6(6) doi: 10.1371/journal.pone.0021344.e21344 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zoeteweij JP, Golding H, Mostowski H, Blauvelt A. Cutting edge: cytokines regulate expression and function of the HIV coreceptor CXCR4 on human mature dendritic cells. The Journal of Immunology. 1998;161(7):3219–3223. [PubMed] [Google Scholar]
- 46.Tohou I, Misery L, Sabido O, et al. Functional HIV CXCR4 coreceptor on human epithelial Langerhans cells and infection by HIV strain X4. Journal of Leukocyte Biology. 2001;70(2):313–321. [PubMed] [Google Scholar]
- 47.Blauvelt A, Asada H, Saville MW, et al. Productive infection of dendritic cells by HIV-1 and their ability to capture virus are mediated through separate pathways. Journal of Clinical Investigation. 1997;100(8):2043–2053. doi: 10.1172/JCI119737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kawamura T, Qualbani M, Thomas EK, Orenstein JM, Blauvelt A. Low levels of productive HIV infection in Langerhans cell-like dendritic cells differentiated in the presence of TGF-beta1 and increased viral replication with CD40 ligand-induced maturation. European Journal of Immunology. 2001;31(2):360–368. doi: 10.1002/1521-4141(200102)31:2<360::aid-immu360>3.0.co;2-x. [DOI] [PubMed] [Google Scholar]
- 49.Sivard P, Berlier W, Picard B, Sabido O, Genin C, Misery L. HIV-1 infection of langerhans cells in a reconstructed vaginal mucosa. Journal of Infectious Diseases. 2004;190(2):227–235. doi: 10.1086/421704. [DOI] [PubMed] [Google Scholar]
- 50.Kawamura T, Bruse SE, Abraha A, et al. PSC-RANTES blocks R5 human immunodeficiency virus infection of Langerhans cells isolated from individuals with a variety of CCR5 diplotypes. Journal of Virology. 2004;78(14):7602–7609. doi: 10.1128/JVI.78.14.7602-7609.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sugaya M, Loré K, Koup RA, Douek DC, Blauvelt A. HIV-infected langerhans cells preferentially transmit virus to proliferating autologous CD4+ memory T cells located within langerhans cell-T cell clusters. The Journal of Immunology. 2004;172(4):2219–2224. doi: 10.4049/jimmunol.172.4.2219. [DOI] [PubMed] [Google Scholar]
- 52.Prakash M, Kapembwa MS, Gotch F, Patterson S. Chemokine receptor expression on mucosal dendritic cells from the endocervix of healthy women. Journal of Infectious Diseases. 2004;190(2):246–250. doi: 10.1086/422034. [DOI] [PubMed] [Google Scholar]
- 53.de Jong MAWP, de Witte L, Oudhoff MJ, Gringhuis SI, Gallay P, Geijtenbeek TBH. TNF-α and TLR agonists increase susceptibility to HIV-1 transmission by human Langerhans cells ex vivo. Journal of Clinical Investigation. 2008;118(10):3440–3452. doi: 10.1172/JCI34721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ogawa Y, Kawamura T, Kimura T, Ito M, Blauvelt A, Shimada S. Gram-positive bacteria enhance HIV-1 susceptibility in Langerhans cells, but not in dendritic cells, via Toll-like receptor activation. Blood. 2009;113(21):5157–5166. doi: 10.1182/blood-2008-10-185728. [DOI] [PubMed] [Google Scholar]
- 55.de Jong MAWP, de Witte L, Taylor ME, Geijtenbeek TBH. Herpes simplex virus type 2 enhances HIV-1 susceptibility by affecting langerhans cell function. The Journal of Immunology. 2010;185(3):1633–1641. doi: 10.4049/jimmunol.0904137. [DOI] [PubMed] [Google Scholar]
- 56.de Witte L, Nabatov A, Pion M, et al. Langerin is a natural barrier to HIV-1 transmission by Langerhans cells. Nature Medicine. 2007;13(3):367–371. doi: 10.1038/nm1541. [DOI] [PubMed] [Google Scholar]
- 57.Platt EJ, Wehrly K, Kuhmann SE, Chesebro B, Kabat D. Effects of CCR5 and CD4 cell surface concentrations on infections by macrophagetropic isolates of human immunodeficiency virus type 1. Journal of Virology. 1998;72(4):2855–2864. doi: 10.1128/jvi.72.4.2855-2864.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Platt EJ, Kozak SL, Durnin JP, Hope TJ, Kabat D. Rapid dissociation of HIV-1 from cultured cells severely limits infectivity assays, causes the inactivation ascribed to entry inhibitors, and masks the inherently high level of infectivity of virions. Journal of Virology. 2010;84(6):3106–3110. doi: 10.1128/JVI.01958-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Girolomoni G, Valle MT, Zacchi V, Costa MG, Giannetti A, Manca F. Cultured human Langerhans’ cells are superior to fresh cells at presenting native HIV-1 protein antigens to specific CD4+ T-cell lines. Immunology. 1996;87(2):310–316. [PMC free article] [PubMed] [Google Scholar]
- 60.Blauvelt A, Glushakova S, Margolis LB. HIV-infected human Langerhans cells transmit infection to human lymphoid tissue ex vivo. AIDS. 2000;14(6):647–651. doi: 10.1097/00002030-200004140-00003. [DOI] [PubMed] [Google Scholar]
- 61.Fahrbach KM, Barry SM, Ayehunie S, Lamore S, Klausner M, Hope TJ. Activated CD34-derived langerhans cells mediate transinfection with human immunodeficiency virus. Journal of Virology. 2007;81(13):6858–6868. doi: 10.1128/JVI.02472-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hladik F, Lentz G, Akridge RE, et al. Dendritic cell-T-cell interactions support coreceptor-independent human immunodeficiency virus type 1 transmission in the human genital tract. Journal of Virology. 1999;73(7):5833–5842. doi: 10.1128/jvi.73.7.5833-5842.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Kawamura T, Koyanagi Y, Nakamura Y, et al. Significant virus replication in langerhans cells following application of HIV to abraded skin: Relevance to occupational transmission of HIV. The Journal of Immunology. 2008;180(5):3297–3304. doi: 10.4049/jimmunol.180.5.3297. [DOI] [PubMed] [Google Scholar]
- 64.Santegoets SJAM, Gibbs S, Kroeze K, et al. Transcriptional profiling of human skin-resident Langerhans cells and CD1a+ dermal dendritic cells: Differential activation states suggest distinct functions. Journal of Leukocyte Biology. 2008;84(1):143–151. doi: 10.1189/jlb.1107750. [DOI] [PubMed] [Google Scholar]
- 65.Banchereau J, Thompson-Snipes L, Zurawski S, et al. The differential production of cytokines by human Langerhans cells and dermal CD14+ DCs controls CTL priming. Blood. 2012;119(24):5742–5749. doi: 10.1182/blood-2011-08-371245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Kwan WH, Helt AM, Marañón C, et al. Dendritic cell precursors are permissive to dengue virus and human immunodeficiency virus infection. Journal of Virology. 2005;79(12):7291–7299. doi: 10.1128/JVI.79.12.7291-7299.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.de Witte L, Nabatov A, Geijtenbeek TBH. Distinct roles for DC-SIGN+-dendritic cells and Langerhans cells in HIV-1 transmission. Trends in Molecular Medicine. 2008;14(1):12–19. doi: 10.1016/j.molmed.2007.11.001. [DOI] [PubMed] [Google Scholar]
- 68.Peressin M, Holl V, Schmidt S, et al. HIV-1 replication in langerhans and interstitial dendritic cells is inhibited by neutralizing and Fc-mediated inhibitory antibodies. Journal of Virology. 2011;85(2):1077–1085. doi: 10.1128/JVI.01619-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Harman AN, Wilkinson J, Bye CR, et al. HIV induces maturation of monocyte-derived dendritic cells and Langerhans cells. The Journal of Immunology. 2006;177(10):7103–7113. doi: 10.4049/jimmunol.177.10.7103. [DOI] [PubMed] [Google Scholar]
- 70.Soto-Ramirez LE, Renjifo B, McLane MF, et al. HIV-1 Langerhans’ cell tropism associated with heterosexual transmission of HIV. Science. 1996;271(5253):1291–1293. doi: 10.1126/science.271.5253.1291. [DOI] [PubMed] [Google Scholar]
- 71.Pope M, Frankel SS, Mascola JR, et al. Human immunodeficiency virus type 1 strains of subtypes B and E replicate in cutaneous dendritic cell-T-cell mixtures without displaying subtype-specific tropism. Journal of Virology. 1997;71(10):8001–8007. doi: 10.1128/jvi.71.10.8001-8007.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Berger R, Gartner S, Rappersberger K, Foster CA, Wolff K, Stingl G. Isolation of human immunodeficiency virus type 1 from human epidermis: virus replication and transmission studies. Journal of Investigative Dermatology. 1992;99(3):271–277. doi: 10.1111/1523-1747.ep12616619. [DOI] [PubMed] [Google Scholar]
- 73.Cameron PU, Freudenthal PS, Barker JM, Gezelter S, Inaba K, Steinman RM. Dendritic cells exposed to human immunodeficiency virus type-1 transmit a vigorous cytopathic infection to CD4+ T cells. Science. 1992;257(5068):383–386. doi: 10.1126/science.1352913. [DOI] [PubMed] [Google Scholar]
- 74.Pope M, Betjes MGH, Romani N, et al. Conjugates of dendritic cells and memory T lymphocytes from skin facilitate productive infection with HIV-1. Cell. 1994;78(3):389–398. doi: 10.1016/0092-8674(94)90418-9. [DOI] [PubMed] [Google Scholar]
- 75.Ayehunie S, Groves RW, Bruzzese AM, Ruprecht RM, Kupper TS, Langhoff E. Acutely infected Langerhans cells are more efficient than T cells in disseminating HIV type 1 to activated T cells following a short cell-cell contact. AIDS Research and Human Retroviruses. 1995;11(8):877–884. doi: 10.1089/aid.1995.11.877. [DOI] [PubMed] [Google Scholar]
- 76.Ganor Y, Zhou Z, Tudor D, et al. Within 1 h, HIV-1 uses viral synapses to enter efficiently the inner, but not outer, foreskin mucosa and engages Langerhans-T cell conjugates. Mucosal Immunology. 2010;3(5):506–522. doi: 10.1038/mi.2010.32. [DOI] [PubMed] [Google Scholar]
- 77.Sugaya M, Hartley O, Root MJ, Blauvelt A. C34, a membrane fusion inhibitor, blocks HIV infection of Langerhans cells and viral transmission to T cells. Journal of Investigative Dermatology. 2007;127(6):1436–1443. doi: 10.1038/sj.jid.5700736. [DOI] [PubMed] [Google Scholar]
- 78.Hladik F, Sakchalathorn P, Ballweber L, et al. Initial events in establishing vaginal entry and infection by human immunodeficiency virus type-1. Immunity. 2007;26(2):257–270. doi: 10.1016/j.immuni.2007.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Patterson S, English NR, Longhurst H, et al. Analysis of human immunodeficiency virus type 1 (HIV-1) variants and levels of infection in dendritic and T cells from symptomatic HIV-1-infected patients. Journal of General Virology. 1998;79(2):247–257. doi: 10.1099/0022-1317-79-2-247. [DOI] [PubMed] [Google Scholar]
- 80.Otero M, Nunnari G, Leto D, et al. Peripheral blood dendritic cells are not a major reservoir for HIV type 1 in infected individuals on virally suppressive HAART. AIDS Research and Human Retroviruses. 2003;19(12):1097–1103. doi: 10.1089/088922203771881194. [DOI] [PubMed] [Google Scholar]
- 81.Langhoff E, Haseltine WA. Infection of accessory dendritic cells by human immunodeficiency virus type 1. Journal of Investigative Dermatology. 1992;99(5):89S–94S. doi: 10.1111/1523-1747.ep12669964. [DOI] [PubMed] [Google Scholar]
- 82.Langhoff E, Kalland KH, Haseltine WA. Early molecular replication of human immunodeficiency virus type 1 in cultured-blood-derived T helper dendritic cells. Journal of Clinical Investigation. 1993;91(6):2721–2726. doi: 10.1172/JCI116512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Chehimi J, Prakash K, Shanmugam V, et al. CD4-independent infection of human peripheral blood dendritic cells with isolates of human immunodeficiency virus type 1. Journal of General Virology. 1993;74, part 7:1277–1285. doi: 10.1099/0022-1317-74-7-1277. [DOI] [PubMed] [Google Scholar]
- 84.Patterson S, Gross J, English N, Stackpoole A, Bedford P, Knight SC. CD4 expression on dendritic cells and their infection by human immunodeficiency virus. Journal of General Virology. 1995;76, part 5:1155–1163. doi: 10.1099/0022-1317-76-5-1155. [DOI] [PubMed] [Google Scholar]
- 85.Pinchuk LM, Polacino PS, Agy MB, Klaus SJ, Clark EA. The role of CD40 and CD80 accessory cell molecules in dendritic cell-dependent HIV-1 infection. Immunity. 1994;1(4):317–325. doi: 10.1016/1074-7613(94)90083-3. [DOI] [PubMed] [Google Scholar]
- 86.Smed-Sörensen A, Loré K, Vasudevan J, et al. Differential susceptibility to human immunodeficiency virus type 1 infection of myeloid and plasmacytoid dendritic cells. Journal of Virology. 2005;79(14):8861–8869. doi: 10.1128/JVI.79.14.8861-8869.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Granelli-Piperno A, Shimeliovich I, Pack M, Trumpfheller C, Steinman RM. HIV-1 selectively infects a subset of nonmaturing BDCA1-positive dendritic cells in human blood. The Journal of Immunology. 2006;176(2):991–998. doi: 10.4049/jimmunol.176.2.991. [DOI] [PubMed] [Google Scholar]
- 88.Weissman D, Daucher J, Barker T, Adelsberger J, Baseler M, Fauci AS. Cytokine regulation of HIV replication induced by dendritic cell-CD4-positive T cell interactions. AIDS Research and Human Retroviruses. 1996;12(9):759–767. doi: 10.1089/aid.1996.12.759. [DOI] [PubMed] [Google Scholar]
- 89.Cameron PU, Pope M, Gezelter S, Steinman RM. Infection and apoptotic cell death of CD4+ T cells during an immune response to HIV-1-pulsed dendritic cells. AIDS Research and Human Retroviruses. 1994;10(1):61–71. doi: 10.1089/aid.1994.10.61. [DOI] [PubMed] [Google Scholar]
- 90.Cameron PU, Lowe MG, Sotzik F, Coughlan AF, Crowe SM, Shortman K. The interaction of macrophage and non-macrophage tropic isolates of HIV-1 with thymic and tonsillar dendritic cells in vitro. Journal of Experimental Medicine. 1996;183(4):1851–1856. doi: 10.1084/jem.183.4.1851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Weissman D, Li Y, Orenstein JM, Fauci AS. Both a precursor and a mature population of dendritic cells can bind HIV: however, only the mature population that expresses CD80 can pass infection to unstimulated CD4+ T cells. The Journal of Immunology. 1995;155(8):4111–4117. [PubMed] [Google Scholar]
- 92.Granelli-Piperno A, Moser B, Pope M, et al. Efficient interaction of HIV-1 with purified dendritic cells via multiple chemokine coreceptors. Journal of Experimental Medicine. 1996;184(6):2433–2438. doi: 10.1084/jem.184.6.2433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Pudney J, Song MJ. Electron microscopic analysis of HIV-host cell interactions. Tissue and Cell. 1994;26(4):539–550. doi: 10.1016/0040-8166(94)90006-x. [DOI] [PubMed] [Google Scholar]
- 94.Cameron PU, Lowe MG, Crowe SM, et al. Susceptibility of dendritic cells to HIV-1 in vitro. Journal of Leukocyte Biology. 1994;56(3):257–265. doi: 10.1002/jlb.56.3.257. [DOI] [PubMed] [Google Scholar]
- 95.Cameron PU, Handley AJ, Baylis DC, et al. Preferential infection of dendritic cells during human immunodeficiency virus type 1 infection of blood leukocytes. Journal of Virology. 2007;81(5):2297–2306. doi: 10.1128/JVI.01795-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Geijtenbeek TBH, Kwon DS, Torensma R, et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell. 2000;100(5):587–597. doi: 10.1016/s0092-8674(00)80694-7. [DOI] [PubMed] [Google Scholar]
- 97.Soilleux EJ, Morris LS, Leslie G, et al. Constitutive and induced expression of DC-SIGN on dendritic cell and macrophage subpopulations in situ and in vitro. Journal of Leukocyte Biology. 2002;71(3):445–457. [PubMed] [Google Scholar]
- 98.Jameson B, Baribaud F, Pöhlmann S, et al. Expression of DC-SIGN by dendritic cells of intestinal and genital mucosae in humans and rhesus macaques. Journal of Virology. 2002;76(4):1866–1875. doi: 10.1128/JVI.76.4.1866-1875.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Engering A, van Vliet SJ, Geijtenbeek TBH, van Kooyk Y. Subset of DC-SIGN+ dendritic cells in human blood transmits HIV-1 to T lymphocytes. Blood. 2002;100(5):1780–1786. doi: 10.1182/blood-2001-12-0179. [DOI] [PubMed] [Google Scholar]
- 100.Gurney KB, Elliott J, Nassanian H, et al. Binding and transfer of human immunodeficiency virus by DC-SIGN+ cells in human rectal mucosa. Journal of Virology. 2005;79(9):5762–5773. doi: 10.1128/JVI.79.9.5762-5773.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Macatangay BJC, Rinaldo CR. Regulatory T cells in HIV immunotherapy. HIV Therapy. 2010;4(6):639–647. doi: 10.2217/hiv.10.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Wendelsdorf K, Dean G, Hu S, Nordone S, Banks HT. Host immune responses that promote initial HIV spread. Journal of Theoretical Biology. 2011;289:17–35. doi: 10.1016/j.jtbi.2011.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Granelli-Piperno A, Golebiowska A, Trumpfheller C, Siegal FP, Steinman RM. HIV-1-infected monocyte-derived dendritic cells do not undergo maturation but can elicit IL-10 production and T cell regulation. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(20):7669–7674. doi: 10.1073/pnas.0402431101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Moreno-Fernandez ME, Rueda CM, Rusie LK, Chougnet CA. Regulatory T cells control HIV replication in activated T cells through a cAMP-dependent mechanism. Blood. 2011;117(20):5372–5380. doi: 10.1182/blood-2010-12-323162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Barat C, Gilbert C, Imbeault M, Tremblay MJ. Extracellular ATP reduces HIV-1 transfer from immature dendritic cells to CD4+ T lymphocytes. Retrovirology. 2008;5, article 30 doi: 10.1186/1742-4690-5-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Derby N, Martinelli E, Robbiani M. Myeloid dendritic cells in HIV-1 infection. Current Opinion in HIV and AIDS. 2011;6(5):379–384. doi: 10.1097/COH.0b013e3283499d63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Sabado RL, O'Brien M, Subedi A, et al. Evidence of dysregulation of dendritic cells in primary HIV infection. Blood. 2010;116(19):3839–3852. doi: 10.1182/blood-2010-03-273763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Dillon SM, Robertson KB, Pan SC, et al. Plasmacytoid and myeloid dendritic cells with a partial activation phenotype accumulate in lymphoid tissue during asymptomatic chronic HIV-1 infection. Journal of Acquired Immune Deficiency Syndromes. 2008;48(1):1–12. doi: 10.1097/QAI.0b013e3181664b60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Almeida M, Cordero M, Almeida J, Orfao A. Abnormal cytokine production by circulating monocytes and dendritic cells of myeloid origin in ART-treated HIV-1+ patients relates to CD4+ T-cell recovery and HCV Co-infection. Current HIV Research. 2007;5(3):325–336. doi: 10.2174/157016207780636524. [DOI] [PubMed] [Google Scholar]
- 110.Huang J, Yang Y, Al-Mozaini M, et al. Dendritic cell dysfunction during primary HIV-1 infection. The Journal of Infectious Diseases. 2011;204(10):1557–1562. doi: 10.1093/infdis/jir616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Vázquez-Guillén JM, García-Jacobo PJ, Zapata-Benavides P, et al. Expression of DC-SIGN in peripheral blood dendritic cells of patients with typical, slow, and rapid progression to AIDS. Archives of Medical Research. 2009;40(2):132–135. doi: 10.1016/j.arcmed.2008.12.002. [DOI] [PubMed] [Google Scholar]
- 112.Meier A, Alter G, Frahm N, et al. MyD88-dependent immune activation mediated by human immunodeficiency virus type 1-encoded toll-like receptor ligands. Journal of Virology. 2007;81(15):8180–8191. doi: 10.1128/JVI.00421-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Meier A, Bagchi A, Sidhu HK, et al. Upregulation of PD-L1 on monocytes and dendritic cells by HIV-1 derived TLR ligands. AIDS. 2008;22(5):655–658. doi: 10.1097/QAD.0b013e3282f4de23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Campbell-Yesufu OT, Gandhi RT. Update on human immunodeficiency virus (HIV)-2 infection. Clinical Infectious Diseases. 2011;52(6):780–787. doi: 10.1093/cid/ciq248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Duvall MG, Loré K, Blaak H, et al. Dendritic cells are less susceptible to human immunodeficiency virus type 2 (HIV-2) infection than to HIV-1 infection. Journal of Virology. 2007;81(24):13486–13498. doi: 10.1128/JVI.00976-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Szabolcs P, Feller ED, Moore MAS, Young JW. Progenitor recruitment and in vitro expansion of immunostimulatory dendritic cells from human CD34+ bone marrow cells by c-kit-ligand, GM-CSF, and TNFα . Advances in Experimental Medicine and Biology. 1995;378:17–20. doi: 10.1007/978-1-4615-1971-3_4. [DOI] [PubMed] [Google Scholar]
- 117.Romani N, Reider D, Heuer M, et al. Generation of mature dendritic cells from human blood. An improved method with special regard to clinical applicability. Journal of Immunological Methods. 1996;196(2):137–151. doi: 10.1016/0022-1759(96)00078-6. [DOI] [PubMed] [Google Scholar]
- 118.Sacchi A, Cappelli G, Cairo C, et al. Differentiation of monocytes into CD1a- dendritic cells correlates with disease progression in HIV-infected patients. Journal of Acquired Immune Deficiency Syndromes. 2007;46(5):519–528. doi: 10.1097/QAI.0b013e31815b1278. [DOI] [PubMed] [Google Scholar]
- 119.Canque B, Bakri Y, Camus S, Yagello M, Benjouad A, Gluckman JC. The susceptibility to X4 and R5 human immunodeficiency virus-1 strains of dendritic cells derived in vitro from CD34+ hematopoietic progenitor cells is primarily determined by their maturation stage. Blood. 1999;93(11):3866–3875. [PubMed] [Google Scholar]
- 120.Warren MK, Rose WL, Cone JL, Rice WG, Turpin JA. Differential infection of CD34+ cell-derived dendritic cells and monocytes with lymphocyte-tropic and monocyte-tropic HIV-1 strains. The Journal of Immunology. 1997;158(10):5035–5042. [PubMed] [Google Scholar]
- 121.Tsunetsugu-Yokota Y, Akagawa K, Kimoto H, et al. Monocyte-derived cultured dendritic cells are susceptible to human immunodeficiency virus infection and transmit virus to resting T cells in the process of nominal antigen presentation. Journal of Virology. 1995;69(7):4544–4547. doi: 10.1128/jvi.69.7.4544-4547.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Tsunetsugu-Yokota Y, Yasuda S, Sugimoto A, et al. Efficient virus transmission from dendritic cells to CD4+ T cells in response to antigen depends on close contact through adhesion molecules. Virology. 1997;239(2):259–268. doi: 10.1006/viro.1997.8895. [DOI] [PubMed] [Google Scholar]
- 123.Rubbert A, Combadiere C, Ostrowski M, et al. Dendritic cells express multiple chemokine receptors used as coreceptors for HIV entry. The Journal of Immunology. 1998;160(8):3933–3941. [PubMed] [Google Scholar]
- 124.Izquierdo-Useros N, Esteban O, Rodriguez-Plata MT, et al. Dynamic imaging of cell-free and cell-associated viral capture in mature dendritic cells. Traffic. 2011;12(12):1702–1713. doi: 10.1111/j.1600-0854.2011.01281.x. [DOI] [PubMed] [Google Scholar]
- 125.Bajtay Z, Speth C, Erdei A, Dierich MP. Cutting edge: productive HIV-1 infection of dendritic cells via complement receptor type 3 (CR3, CD11b/CD18) The Journal of Immunology. 2004;173(8):4775–4778. doi: 10.4049/jimmunol.173.8.4775. [DOI] [PubMed] [Google Scholar]
- 126.Pruenster M, Wilflingseder D, Bánki Z, et al. C-type lectin-independent interaction of complement opsonized HIV with monocyte-derived dendritic cells. European Journal of Immunology. 2005;35(9):2691–2698. doi: 10.1002/eji.200425940. [DOI] [PubMed] [Google Scholar]
- 127.Holl V, Peressin M, Schmidt S, et al. Efficient inhibition of HIV-1 replication in human immature monocyte-derived dendritic cells by purified anti-HIV-1 IgG without induction of maturation. Blood. 2006;107(11):4466–4474. doi: 10.1182/blood-2005-08-3490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Wilflingseder D, Banki Z, Garcia E, et al. IgG opsonization of HIV impedes provirus formation in and infection of dendritic cells and subsequent long-term transfer to T cells. The Journal of Immunology. 2007;178(12):7840–7848. doi: 10.4049/jimmunol.178.12.7840. [DOI] [PubMed] [Google Scholar]
- 129.Soederholm A, Bánki Z, Wilflingseder D, et al. HIV-1 induced generation of C5a attracts immature dendritic cells and promotes infection of autologous T cells. European Journal of Immunology. 2007;37(8):2156–2163. doi: 10.1002/eji.200636820. [DOI] [PubMed] [Google Scholar]
- 130.Steinman RM. Decisions about dendritic cells: past, present, and future. Annual Review of Immunology. 2012;30:1–22. doi: 10.1146/annurev-immunol-100311-102839. [DOI] [PubMed] [Google Scholar]
- 131.Granelli-Piperno A, Delgado E, Finkel V, Paxton W, Steinman RM. Immature dendritic cells selectively replicate macrophagetropic (M- tropic) human immunodeficiency virus type 1, while mature cells efficiently transmit both M- and T-tropic virus to T cells. Journal of Virology. 1998;72(4):2733–2737. doi: 10.1128/jvi.72.4.2733-2737.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.McDyer JF, Dybul M, Goletz TJ, et al. Differential effects of CD40 ligand/trimer stimulation on the ability of dendritic cells to replicate and transmit HIV infection: evidence for CC- chemokine-dependent and -independent mechanisms. The Journal of Immunology. 1999;162(6):3711–3717. [PubMed] [Google Scholar]
- 133.Izquierdo-Useros N, Blanco J, Erkizia I, et al. Maturation of blood-derived dendritic cells enhances human immunodeficiency virus type 1 capture and transmission. Journal of Virology. 2007;81(14):7559–7570. doi: 10.1128/JVI.02572-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Dong C, Janas AM, Wang JH, Olson WJ, Wu L. Characterization of human immunodeficiency virus type 1 replication in immature and mature dendritic cells reveals dissociable cis- and trans-infection. Journal of Virology. 2007;81(20):11352–11362. doi: 10.1128/JVI.01081-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Mailliard RB, Wankowicz-Kalinska A, Cai Q, et al. α-type-1 polarized dendritic cells: a novel immunization tool with optimized CTL-inducing activity. Cancer Research. 2004;64(17):5934–5937. doi: 10.1158/0008-5472.CAN-04-1261. [DOI] [PubMed] [Google Scholar]
- 136.Jonuleit H, Kühn U, Müller G, et al. Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. European Journal of Immunology. 1997;27(12):3135–3142. doi: 10.1002/eji.1830271209. [DOI] [PubMed] [Google Scholar]
- 137.Sanders RW, de Jong EC, Baldwin CE, Schuitemaker JHN, Kapsenberg ML, Berkhout B. Differential transmission of human immunodeficiency virus type 1 by distinct subsets of effector dendritic cells. Journal of Virology. 2002;76(15):7812–7821. doi: 10.1128/JVI.76.15.7812-7821.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Frank I, Kacani L, Stoiber H, et al. Human immunodeficiency virus type 1 derived from cocultures of immature dendritic cells with autologous T cells carries T-cell-specific molecules on its surface and is highly infectious. Journal of Virology. 1999;73(4):3449–3454. doi: 10.1128/jvi.73.4.3449-3454.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Macdougall THJ, Shattock RJ, Madsen C, Chain BM, Katz DR. Regulation of primary HIV-1 isolate replication in dendritic cells. Clinical and Experimental Immunology. 2002;127(1):66–71. doi: 10.1046/j.1365-2249.2002.01715.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Cavrois M, Neidleman J, Kreisberg JF, Fenard D, Callebaut C, Greene WC. Human immunodeficiency virus fusion to dendritic cells declines as cells mature. Journal of Virology. 2006;80(4):1992–1999. doi: 10.1128/JVI.80.4.1992-1999.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Pion M, Arrighi JF, Jiang J, et al. Analysis of HIV-1-X4 fusion with immature dendritic cells identifies a specific restriction that is independent of CXCR4 levels. Journal of Investigative Dermatology. 2007;127(2):319–323. doi: 10.1038/sj.jid.5700518. [DOI] [PubMed] [Google Scholar]
- 142.Gilbert C, Barat C, Cantin R, Tremblay MJ. Involvement of Src and Syk tyrosine kinases in HIV-1 transfer from dendritic cells to CD4+ T lymphocytes. The Journal of Immunology. 2007;178(5):2862–2871. doi: 10.4049/jimmunol.178.5.2862. [DOI] [PubMed] [Google Scholar]
- 143.Gilbert C, Cantin R, Barat C, Tremblay MJ. Human immunodeficiency virus type 1 replication in dendritic cell-T-cell cocultures is increased upon incorporation of host LFA-1 due to higher levels of virus production in immature dendritic cells. Journal of Virology. 2007;81(14):7672–7682. doi: 10.1128/JVI.02810-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.González N, Bermejo M, Calonge E, et al. SDF-1/CXCL12 production by mature dendritic cells inhibits the propagation of X4-tropic HIV-1 isolates at the dendritic cell-T-cell infectious synapse. Journal of Virology. 2010;84(9):4341–4351. doi: 10.1128/JVI.02449-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Fairman P, Angel JB. The effect of human immunodeficiency virus-1 on monocyte-derived dendritic cell maturation and function. Clinical & Experimental Immunology. 2012;170(1):101–113. doi: 10.1111/j.1365-2249.2012.04628.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Wang JH, Janas AM, Olson WJ, KewalRamani VN, Wu L. CD4 coexpression regulates DC-SIGN-mediated transmission of human immunodeficiency virus type 1. Journal of Virology. 2007;81(5):2497–2507. doi: 10.1128/JVI.01970-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Wang JH, Janas AM, Olson WJ, Wu L. Functionally distinct transmission of human immunodeficiency virus type 1 mediated by immature and mature dendritic cells. Journal of Virology. 2007;81(17):8933–8943. doi: 10.1128/JVI.00878-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Luo C, Wang K, Liu D, Li Y, Zhao QS. The functional roles of lipid rafts in T cell activation, immune diseases and HIV infection and prevention. Cellular and Molecular Immunology. 2008;5(1):1–7. doi: 10.1038/cmi.2008.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Cambi A, de Lange F, van Maarseveen NM, et al. Microdomains of the C-type lectin DC-SIGN are portals for virus entry into dendritic cells. Journal of Cell Biology. 2004;164(1):145–155. doi: 10.1083/jcb.200306112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Caparrós E, Munoz P, Sierra-Filardi E, et al. DC-SIGN ligation on dendritic cells results in ERK and PI3K activation and modulates cytokine production. Blood. 2006;107(10):3950–3958. doi: 10.1182/blood-2005-03-1252. [DOI] [PubMed] [Google Scholar]
- 151.Gummuluru S, Rogel M, Stamatatos L, Emerman M. Binding of human immunodeficiency virus type 1 to immature dendritic cells can occur independently of DC-SIGN and mannose binding C-type lectin receptors via a cholesterol-dependent pathway. Journal of Virology. 2003;77(23):12865–12874. doi: 10.1128/JVI.77.23.12865-12874.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Hatch SC, Archer J, Gummuluru S. Glycosphingolipid composition of Human Immunodeficiency Virus type 1 (HIV-1) particles is a crucial determinant for dendritic cell-mediated HIV-1 trans-infection. Journal of Virology. 2009;83(8):3496–3506. doi: 10.1128/JVI.02249-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Wang JH, Wells C, Wu L. Macropinocytosis and cytoskeleton contribute to dendritic cell-mediated HIV-1 transmission to CD4+ T cells. Virology. 2008;381(1):143–154. doi: 10.1016/j.virol.2008.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Hanley TM, Puryear WB, Gummuluru S, Viglianti GA. PPARγ and LXR signaling inhibit dendritic cell-mediated HIV-1 capture and trans-infection. PLoS Pathogens. 2010;6(7) doi: 10.1371/journal.ppat.1000981.e1000981 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Puryear WB, Yu X, Ramirez NP, Reinhard BM, Gummuluru S. HIV-1 incorporation of host-cell-derived glycosphingolipid GM3 allows for capture by mature dendritic cells. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(19):7475–7480. doi: 10.1073/pnas.1201104109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Izquierdo-Useros N, Lorizate M, Contreras FX, et al. Sialyllactose in viral membrane gangliosides is a novel molecular recognition pattern for mature dendritic cell capture of HIV-1. PLOS Biologyogy. 2012;10(4) doi: 10.1371/journal.pbio.1001315.e1001315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Izquierdo-Useros N, Lorizate M, Puertas MC, et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLOS Biology. 2012;10(12) doi: 10.1371/journal.pbio.1001448.e1001448 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Blanc M, Hsieh WY, Robertson KA, et al. Host defense against viral infection involves interferon mediated down-regulation of sterol biosynthesis. PLOS Biology. 2011;9(3) doi: 10.1371/journal.pbio.1000598.e1000598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Blanc M, Hsieh WY, Robertson KA, et al. The transcription factor STAT-1 couples macrophage synthesis of 25-hydroxycholesterol to the interferon antiviral response. Immunity. 2013;38(1):106–118. doi: 10.1016/j.immuni.2012.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Liu SY, Aliyari R, Chikere K, et al. Interferon-Inducible Cholesterol-25-Hydroxylase Broadly Inhibits Viral Entry by Production of 25-Hydroxycholesterol. Immunity. 2013;38(1):92–105. doi: 10.1016/j.immuni.2012.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Garcia E, Nikolic DS, Piguet V. HIV-1 replication in dendritic cells occurs through a tetraspanin-containing compartment enriched in AP-3. Traffic. 2008;9(2):200–214. doi: 10.1111/j.1600-0854.2007.00678.x. [DOI] [PubMed] [Google Scholar]
- 162.Turville SG, Aravantinou M, Stössel H, Romani N, Robbiani M. Resolution of de novo HIV production and trafficking in immature dendritic cells. Nature Methods. 2008;5(1):75–85. doi: 10.1038/nmeth1137. [DOI] [PubMed] [Google Scholar]
- 163.Nydegger S, Khurana S, Krementsov DN, Foti M, Thali M. Mapping of tetraspanin-enriched microdomains that can function as gateways for HIV-1. Journal of Cell Biology. 2006;173(5):795–807. doi: 10.1083/jcb.200508165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Ono A. Relationships between plasma membrane microdomains and HIV-1 assembly. Biology of the Cell. 2010;102(6):335–350. doi: 10.1042/BC20090165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Spear GT, Zariffard MR, Xin J, Saifuddin M. Inhibition of DC-SIGN-mediated trans infection of T cells by mannose-binding lectin. Immunology. 2003;110(1):80–85. doi: 10.1046/j.1365-2567.2003.01707.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Arrighi JF, Pion M, Garcia E, et al. DC-SIGN-mediated infectious synapse formation enhances X4 HIV-1 transmission from dendritic cells to T cells. Journal of Experimental Medicine. 2004;200(10):1279–1288. doi: 10.1084/jem.20041356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Arrighi JF, Pion M, Wiznerowicz M, et al. Lentivirus-mediated RNA interference of DC-SIGN expression inhibits human immunodeficiency virus transmission from dendritic cells to T cells. Journal of Virology. 2004;78(20):10848–10855. doi: 10.1128/JVI.78.20.10848-10855.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Wu L, Martin TD, Han YC, Breun SKJ, KewalRamani VN. Trans-dominant cellular inhibition of DC-SIGN-mediated HIV-1 transmission. Retrovirology. 2004;1, article 14 doi: 10.1186/1742-4690-1-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Lin CL, Sewell AK, Gao GF, Whelan KT, Phillips RE, Austyn JM. Macrophage-tropic HIV induces and exploits dendritic cell chemotaxis. Journal of Experimental Medicine. 2000;192(4):587–593. doi: 10.1084/jem.192.4.587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Kwon DS, Gregorio G, Bitton N, Hendrickson WA, Littman DR. DC-SIGN-mediated internalization of HIV is required for trans-enhancement of T cell infection. Immunity. 2002;16(1):135–144. doi: 10.1016/s1074-7613(02)00259-5. [DOI] [PubMed] [Google Scholar]
- 171.Bernhard OK, Lai J, Wilkinson J, Sheil MM, Cunningham AL. Proteomic analysis of DC-SIGN on dendritic cells detects tetramers required for ligand binding but no association with CD4. Journal of Biological Chemistry. 2004;279(50):51828–51835. doi: 10.1074/jbc.M402741200. [DOI] [PubMed] [Google Scholar]
- 172.Hong PWP, Nguyen S, Young S, Su SV, Lee B. Identification of the optimal DC-SIGN binding site on human immunodeficiency virus type 1 gp120. Journal of Virology. 2007;81(15):8325–8336. doi: 10.1128/JVI.01765-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.van Montfort T, Eggink D, Boot M, et al. HIV-1 N-glycan composition governs a balance between dendritic cell-mediated viral transmission and antigen presentation. The Journal of Immunology. 2011;187(9):4676–4685. doi: 10.4049/jimmunol.1101876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Sato S, St-Pierre C, Bhaumik P, Nieminen J. Galectins in innate immunity: dual functions of host soluble β-galactoside-binding lectins as damage-associated molecular patterns (DAMPs) and as receptors for pathogen-associated molecular patterns (PAMPs) Immunological Reviews. 2009;230(1):172–187. doi: 10.1111/j.1600-065X.2009.00790.x. [DOI] [PubMed] [Google Scholar]
- 175.Sabado RL, Babcock E, Kavanagh DG, et al. Pathways utilized by dendritic cells for binding, uptake, processing and presentation of antigens derived from HIV-1. European Journal of Immunology. 2007;37(7):1752–1763. doi: 10.1002/eji.200636981. [DOI] [PubMed] [Google Scholar]
- 176.Smith AL, Ganesh L, Leung K, Jongstra-Bilen J, Jongstra J, Nabel GJ. Leukocyte-specific protein 1 interacts with DC-SIGN and mediates transport of HIV to the proteasome in dendritic cells. Journal of Experimental Medicine. 2007;204(2):421–430. doi: 10.1084/jem.20061604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Martinez-Nunez RT, Louafi F, Friedmann PS, Sanchez-Eisner T. MicroRNA-155 modulates the pathogen binding ability of dendritic cells (DCs) by down-regulation of DC-specific intercellular adhesion molecule-3 grabbing non-integrin (DC-SIGN) Journal of Biological Chemistry. 2009;284(24):16334–16342. doi: 10.1074/jbc.M109.011601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Nabatov AA, van Montfort T, Geijtenbeek TBH, Pollakis G, Paxton WA. Interaction of HIV-1 with dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin-expressing cells is influenced by gp120 envelope modifications associated with disease progression. FEBS Journal. 2006;273(21):4944–4958. doi: 10.1111/j.1742-4658.2006.05491.x. [DOI] [PubMed] [Google Scholar]
- 179.Borggren M, Repits J, Kuylenstierna C, et al. Evolution of DC-SIGN use revealed by fitness studies of R5 HIV-1 variants emerging during AIDS progression. Retrovirology. 2008;5, article 28 doi: 10.1186/1742-4690-5-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Reece JC, Vardaxis NJ, Marshall JA, Crowe SM, Cameron PU. Uptake of HIV and latex particles by fresh and cultured dendritic cells and monocytes. Immunology and Cell Biology. 2001;79(3):255–263. doi: 10.1046/j.1440-1711.2001.01011.x. [DOI] [PubMed] [Google Scholar]
- 181.Engering A, Geijtenbeek TBH, van Vliet SJ, et al. The dendritic cell-specific adhesion receptor DC-SIGN internalizes antigen for presentation to T cells. The Journal of Immunology. 2002;168(5):2118–2126. doi: 10.4049/jimmunol.168.5.2118. [DOI] [PubMed] [Google Scholar]
- 182.Popov S, Chenine AL, Gruber A, Li PL, Ruprecht RM. Long-term productive human immunodeficiency virus infection of CD1a-sorted myeloid dendritic cells. Journal of Virology. 2005;79(1):602–608. doi: 10.1128/JVI.79.1.602-608.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Nobile C, Petit C, Moris A, et al. Covert human immunodeficiency virus replication in dendritic cells and in DC-SIGN-expressing cells promotes long-term transmission to lymphocytes. Journal of Virology. 2005;79(9):5386–5399. doi: 10.1128/JVI.79.9.5386-5399.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Gringhuis SI, van der Vlist M, van den Berg LM, Den Dunnen J, Litjens M, Geijtenbeek TBH. HIV-1 exploits innate signaling by TLR8 and DC-SIGN for productive infection of dendritic cells. Nature Immunology. 2010;11(5):419–426. doi: 10.1038/ni.1858. [DOI] [PubMed] [Google Scholar]
- 185.David SA, Smith MS, Lopez GJ, et al. Selective transmission of R5-tropic HIV type 1 from dendritic cells to resting CD4+ T cells. AIDS Research and Human Retroviruses. 2001;17(1):59–68. doi: 10.1089/088922201750056799. [DOI] [PubMed] [Google Scholar]
- 186.Ganesh L, Leung K, Loré K, et al. Infection of specific dendritic cells by CCR5-tropic human immunodeficiency virus type 1 promotes cell-mediated transmission of virus resistant to broadly neutralizing antibodies. Journal of Virology. 2004;78(21):11980–11987. doi: 10.1128/JVI.78.21.11980-11987.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.van Montfort T, Nabatov AA, Geijtenbeek TBH, Pollakis G, Paxton WA. Efficient capture of antibody neutralized HIV-1 by cells expressing DC-SIGN and transfer to CD4+ T lymphocytes. The Journal of Immunology. 2007;178(5):3177–3185. doi: 10.4049/jimmunol.178.5.3177. [DOI] [PubMed] [Google Scholar]
- 188.Scales D, Ni H, Shaheen F, Capodici J, Cannon G, Weissman D. Nonproliferating bystander CD4+ T cells lacking activation markers support HIV replication during immune activation. The Journal of Immunology. 2001;166(10):6437–6443. doi: 10.4049/jimmunol.166.10.6437. [DOI] [PubMed] [Google Scholar]
- 189.Moris A, Pajot A, Blanchet F, Guivel-Benhassine F, Salcedo M, Schwartz O. Dendritic cells and HIV-specific CD4+ T cells: HIV antigen presentation, T-cell activation, and viral transfer. Blood. 2006;108(5):1643–1651. doi: 10.1182/blood-2006-02-006361. [DOI] [PubMed] [Google Scholar]
- 190.Bouhlal H, Chomont N, Réquena M, et al. Opsonization of HIV with complement enhances infection of dendritic cells and viral transfer to CD4 T cells in a CR3 and DC-SIGN-dependent manner. The Journal of Immunology. 2007;178(2):1086–1095. doi: 10.4049/jimmunol.178.2.1086. [DOI] [PubMed] [Google Scholar]
- 191.Tjomsland V, Ellegard R, Che K, Hinkula J, Lifson JD, Larsson M. Complement opsonization of HIV-1 enhances the uptake by dendritic cells and involves the endocytic lectin and integrin receptor families. PLoS One. 2011;6(8) doi: 10.1371/journal.pone.0023542.e23542 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Turville SG, Santos JJ, Frank I, et al. Immunodeficiency virus uptake, turnover, and 2-phase transfer in human dendritic cells. Blood. 2004;103(6):2170–2179. doi: 10.1182/blood-2003-09-3129. [DOI] [PubMed] [Google Scholar]
- 193.Muratori C, Bona R, Ruggiero E, et al. DC contact with HIV-1-infected cells leads to high levels of Env-mediated virion endocytosis coupled with enhanced HIV-1 Ag presentation. European Journal of Immunology. 2009;39(2):404–416. doi: 10.1002/eji.200838751. [DOI] [PubMed] [Google Scholar]
- 194.Venkatachari NJ, Alber S, Watkins SC, Ayyavoo V. HIV-1 infection of DC: evidence for the acquisition of virus particles from infected T cells by antigen uptake mechanism. PLoS ONE. 2009;4(10) doi: 10.1371/journal.pone.0007470.e7470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Ruggiero E, Toschi E, Federico M. The activity of matrix metalloproteinase-9 is part of the mechanism of cell-to-cell HIV-1 endocytosis in dendritic cells. Current Drug Discovery Technologies. 2011;8(2):112–118. doi: 10.2174/157016311795563893. [DOI] [PubMed] [Google Scholar]
- 196.Clotet-Codina I, Bosch B, Senserrich J, et al. HIV endocytosis after dendritic cell to T cell viral transfer leads to productive virus infection. Antiviral Research. 2009;83(1):94–98. doi: 10.1016/j.antiviral.2009.03.009. [DOI] [PubMed] [Google Scholar]
- 197.Muratori C, Ruggiero E, Sistigu A, Bona R, Federico M. Human immunodeficiency virus type 1 (HIV-1) protease inhibitors block cell-to-cell HIV-1 endocytosis in dendritic cells. Journal of General Virology. 2009;90, part 11:2777–2787. doi: 10.1099/vir.0.012609-0. [DOI] [PubMed] [Google Scholar]
- 198.Garcia E, Pion M, Pelchen-Matthews A, et al. HIV-1 trafficking to the dendritic cell-T-cell infectious synapse uses a pathway of tetraspanin sorting to the immunological synapse. Traffic. 2005;6(6):488–501. doi: 10.1111/j.1600-0854.2005.00293.x. [DOI] [PubMed] [Google Scholar]
- 199.Neumann AK, Thompson NL, Jacobson K. Distribution and lateral mobility of DC-SIGN on immature dendritic cells—implications for pathogen uptake. Journal of Cell Science. 2008;121, part 5:634–643. doi: 10.1242/jcs.022418. [DOI] [PubMed] [Google Scholar]
- 200.Janas AM, Dong C, Wang JH, Wu L. Productive infection of human immunodeficiency virus type 1 in dendritic cells requires fusion-mediated viral entry. Virology. 2008;375(2):442–451. doi: 10.1016/j.virol.2008.01.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Hijazi K, Wang Y, Scala C, et al. DC-SIGN increases the affinity of HIV-1 envelope glycoprotein interaction with CD4. PLoS One. 2011;6(12) doi: 10.1371/journal.pone.0028307.e28307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Baribaud F, Pöhlmann S, Leslie G, Mortari F, Doms RW. Quantitative expression and virus transmission analysis of DC-SIGN on monocyte-derived dendritic cells. Journal of Virology. 2002;76(18):9135–9142. doi: 10.1128/JVI.76.18.9135-9142.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Harman AN, Kraus M, Bye CR, et al. HIV-1-infected dendritic cells show 2 phases of gene expression changes, with lysosomal enzyme activity decreased during the second phase. Blood. 2009;114(1):85–94. doi: 10.1182/blood-2008-12-194845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Cavrois M, Neidleman J, Greene WC. The Achilles Heel of the Trojan horse model of HIV-1 trans-infection. PLoS Pathogens. 2008;4(6) doi: 10.1371/journal.ppat.1000051.e1000051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Burleigh L, Lozach PY, Schiffer C, et al. Infection of dendritic cells (DCs), not DC-SIGN-mediated internalization of human immunodeficiency virus, is required for long-term transfer of virus to T cells. Journal of Virology. 2006;80(6):2949–2957. doi: 10.1128/JVI.80.6.2949-2957.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Yu HJ, Reuter MA, McDonald D. HIV traffics through a specialized, surface-accessible intracellular compartment during trans-infection of T cells by mature dendritic cells. PLoS Pathogens. 2008;4(8) doi: 10.1371/journal.ppat.1000134.e1000134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Cavrois M, Neidleman J, Kreisberg JF, Greene WC. In vitro derived dendritic cells trans-infect CD4 T cells primarily with surface-bound HIV-1 virions. PLoS pathogens. 2007;3(1):p. e4. doi: 10.1371/journal.ppat.0030004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Wiley RD, Gummuluru S. Immature dendritic cell-derived exosomes can mediate HIV-1 trans infection. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(3):738–743. doi: 10.1073/pnas.0507995103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Sealy R, Jones BG, Surman SL, Hurwitz JL. Short communication: the dead cell: a potent escort for HIV type 1 transinfection. AIDS Research and Human Retroviruses. 2009;25(11):1123–1128. doi: 10.1089/aid.2009.0021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Izquierdo-Useros N, Naranjo-Gómez M, Archer J, et al. Capture and transfer of HIV-1 particles by mature dendritic cells converges with the exosome-dissemination pathway. Blood. 2009;113(12):2732–2741. doi: 10.1182/blood-2008-05-158642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Mohanram V, Johansson U, Sköld AE, et al. Exposure to apoptotic activated CD4+ T cells induces maturation and APOBEC3G- mediated inhibition of HIV-1 infection in dendritic cells. PLoS One. 2011;6(6) doi: 10.1371/journal.pone.0021171.e21171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Boggiano C, Manel N, Littman DR. Dendritic cell-mediated trans-enhancement of human immunodeficiency virus type 1 infectivity is independent of DC-SIGN. Journal of Virology. 2007;81(5):2519–2523. doi: 10.1128/JVI.01661-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Gummuluru S, KewalRamani VN, Emerman M. Dendritic cell-mediated viral transfer to T cells is required for human immunodeficiency virus type 1 persistence in the face of rapid cell turnove. Journal of Virology. 2002;76(21):10692–10701. doi: 10.1128/JVI.76.21.10692-10701.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Wu L, Martin TD, Vazeux R, Unutmaz D, KewalRamani VN. Functional evaluation of DC-SIGN monoclonal antibodies reveals DC-SIGN interactions with ICAM-3 do not promote human immunodeficiency virus type 1 transmission. Journal of Virology. 2002;76(12):5905–5914. doi: 10.1128/JVI.76.12.5905-5914.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Bounou S, Giguère JF, Cantin R, et al. The importance of virus-associated host ICAM-1 in human immunodeficiency virus type 1 dissemination depends on the cellular context. FASEB Journal. 2004;18(11):1294–1296. doi: 10.1096/fj.04-1755fje. [DOI] [PubMed] [Google Scholar]
- 216.Moris A, Nobile C, Buseyne F, Porrot F, Abastado JP, Schwartz O. DC-SIGN promotes exogenous MHC-I-restricted HIV-1 antigen presentation. Blood. 2004;103(7):2648–2654. doi: 10.1182/blood-2003-07-2532. [DOI] [PubMed] [Google Scholar]
- 217.Lambert AA, Gilbert C, Richard M, Beaulieu AD, Tremblay MJ. The C-type lectin surface receptor DCIR acts as a new attachment factor for HIV-1 in dendritic cells and contributes to trans- and cis-infection pathways. Blood. 2008;112(4):1299–1307. doi: 10.1182/blood-2008-01-136473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Klechevsky E, Flamar AL, Cao Y, et al. Cross-priming CD8+ T cells by targeting antigens to human dendritic cells through DCIR. Blood. 2010;116(10):1685–1697. doi: 10.1182/blood-2010-01-264960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Lambert AA, Imbeault ML, Gilbert C, Tremblay MJ. HIV-1 induces DCIR expression in CD4+ T cells. PLoS Pathogens. 2010;6(11) doi: 10.1371/journal.ppat.1001188.e1001188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Lambert AA, Barabé F, Gilbert C, Tremblay MJ. DCIR-mediated enhancement of HIV-1 infection requires the ITIM-associated signal transduction pathway. Blood. 2011;117(24):6589–6599. doi: 10.1182/blood-2011-01-331363. [DOI] [PubMed] [Google Scholar]
- 221.de Witte L, Bobardt M, Chatterji U, et al. Syndecan-3 is a dendritic cell-specific attachment receptor for HIV-1. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(49):19464–19469. doi: 10.1073/pnas.0703747104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Gaiha GD, Dong T, Palaniyar N, Mitchell DA, Reid KBM, Clark HW. Surfactant protein a binds to HIV and inhibits direct infection of cd4+ cells, but enhances dendritic cell-mediated viral transfer. The Journal of Immunology. 2008;181(1):601–609. doi: 10.4049/jimmunol.181.1.601. [DOI] [PubMed] [Google Scholar]
- 223.Magérus-Chatinet A, Yu H, Garcia S, Ducloux E, Terris B, Bomsel M. Galactosyl ceramide expressed on dendritic cells can mediate HIV-1 transfer from monocyte derived dendritic cells to autologous T cells. Virology. 2007;362(1):67–74. doi: 10.1016/j.virol.2006.11.035. [DOI] [PubMed] [Google Scholar]
- 224.Bluestone JA, MacKay CR, O’Shea JJ, Stockinger B. The functional plasticity of T cell subsets. Nature Reviews Immunology. 2009;9(11):811–816. doi: 10.1038/nri2654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Coussens LM, Zitvogel L, Palucka AK. Neutralizing tumor-promoting chronic inflammation: a magic bullet? Science. 2013;339(6117):286–291. doi: 10.1126/science.1232227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Perreau M, Savoye AL, de Crignis E, et al. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. The Journal of Experimental Medicine. 2013;210(1):143–156. doi: 10.1084/jem.20121932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Ma CS, Deenick EK, Batten M, Tangye SG. The origins, function, and regulation of T follicular helper cells. The Journal of Experimental Medicine. 2012;209(7):1241–1253. doi: 10.1084/jem.20120994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Pallikkuth S, Parmigiani A, Pahwa S. The role of interleukin-21 in HIV infection. Cytokine & Growth Factor Reviews. 2012;23(4-5):173–180. doi: 10.1016/j.cytogfr.2012.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Shan L, Deng K, Shroff NS, et al. Stimulation of HIV-1-specific cytolytic T lymphocytes facilitates elimination of latent viral reservoir after virus reactivation. Immunity. 2012;36(3):491–501. doi: 10.1016/j.immuni.2012.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Massanella M, Martinez-Picado J, Blanco J. Attacking the HIV reservoir from the immune and viral perspective. Current HIV/AIDS Reports. 2013;10(1):33–41. doi: 10.1007/s11904-012-0150-8. [DOI] [PubMed] [Google Scholar]
- 231.Deeks SG, Autran B, Berkhout B, et al. Towards an HIV cure: a global scientific strategy. Nature Reviews Immunology. 2012;12(8):607–614. doi: 10.1038/nri3262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Chomont N, El-Far M, Ancuta P, et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nature Medicine. 2009;15(8):893–900. doi: 10.1038/nm.1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Fackler OT, Alcover A, Schwartz O. Modulation of the immunological synapse: a key to HIV-1 pathogenesis? Nature Reviews Immunology. 2007;7(4):310–317. doi: 10.1038/nri2041. [DOI] [PubMed] [Google Scholar]
- 234.Lehmann M, Nikolic DS, Piguet V. How HIV-1 takes advantage of the cytoskeleton during replication and cell-to-cell transmission. Viruses. 2011;3(9):1757–1776. doi: 10.3390/v3091757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Nikolic DS, Lehmann M, Felts R, et al. HIV-1 activates Cdc42 and induces membrane extensions in immature dendritic cells to facilitate cell-to-cell virus propagation. Blood. 2011;118(18):4841–4852. doi: 10.1182/blood-2010-09-305417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Aggarwal A, Iemma TL, Shih I, et al. Mobilization of HIV spread by diaphanous 2 dependent filopodia in infected dendritic cells. PLOS Pathogens. 2012;8(6) doi: 10.1371/journal.ppat.1002762.e1002762 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Felts RL, Narayan K, Estes JD, et al. 3D visualization of HIV transfer at the virological synapse between dendritic cells and T cells. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(30):13336–13341. doi: 10.1073/pnas.1003040107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Hogue IB, Bajaria SH, Fallert BA, Qin S, Reinhart TA, Kirschner DE. The dual role of dendritic cells in the immune response to human immunodeficiency virus type 1 infection. Journal of General Virology. 2008;89, part 9:2228–2239. doi: 10.1099/vir.0.83600-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 239.Barat C, Gilbert C, Tremblay MJ. Efficient replication of human immunodeficiency virus type 1 in resting CD4+ T lymphocytes is induced by coculture with autologous dendritic cells in the absence of foreign antigens. Journal of Virology. 2009;83(6):2778–2782. doi: 10.1128/JVI.01420-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Dai J, Agosto LM, Baytop C, et al. Human immunodeficiency virus integrates directly into naïve resting CD4+ T cells but enters naïve cells less efficiently than memory cells. Journal of Virology. 2009;83(9):4528–4537. doi: 10.1128/JVI.01910-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Lichtner M, Marañón C, Vidalain PO, et al. HIV type 1-infected dendritic cells induce apoptotic death in infected and uninfected primary CD4+ T lymphocytes. AIDS Research and Human Retroviruses. 2004;20(2):175–182. doi: 10.1089/088922204773004897. [DOI] [PubMed] [Google Scholar]
- 242.Che KF, Shankar EM, Muthu S, et al. P38MAPK/STAT3 pathway signaling regulates expression of inhibitory molecules in T-cells activated by HIV-1 exposed dendritic cells. Molecular Medicine. 2012;18:1169–1182. doi: 10.2119/molmed.2012.00103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Wang JH, Kwas C, Wu L. Intercellular adhesion molecule 1 (ICAM-1), but not ICAM-2 and -3, is important for dendritic cell-mediated human immunodeficiency virus type 1 transmission. Journal of Virology. 2009;83(9):4195–4204. doi: 10.1128/JVI.00006-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Groot F, Kuijpers TW, Berkhout B, de Jong EC. Dendritic cell-mediated HIV-1 transmission to T cells of LAD-1 patients is impaired due to the defect in LFA-1. Retrovirology. 2006;3, article 75 doi: 10.1186/1742-4690-3-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 245.Biggins JE, Biesinger T, Yu Kimata MT, Arora R, Kimata JT. ICAM-3 influences human immunodeficiency virus type 1 replication in CD4+ T cells independent of DC-SIGN-mediated transmission. Virology. 2007;364(2):383–394. doi: 10.1016/j.virol.2007.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Ketas TJ, Klasse PJ, Spenlehauer C, et al. Entry inhibitors SCH-C, RANTES, and T-20 block HIV type 1 replication in multiple cell types. AIDS Research and Human Retroviruses. 2003;19(3):177–186. doi: 10.1089/088922203763315678. [DOI] [PubMed] [Google Scholar]
- 247.Ketas TJ, Frank I, Klasse PJ, et al. Human immunodeficiency virus type 1 attachment, coreceptor, and fusion inhibitors are active against both direct and trans infection of primary cells. Journal of Virology. 2003;77(4):2762–2767. doi: 10.1128/JVI.77.4.2762-2767.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Hodges A, Sharrocks K, Edelmann M, et al. Activation of the lectin DC-SIGN induces an immature dendritic cell phenotype triggering Rho-GTPase activity required for HIV-1 replication. Nature Immunology. 2007;8(6):569–577. doi: 10.1038/ni1470. [DOI] [PubMed] [Google Scholar]
- 249.Yamamoto T, Tsunetsugu-Yokota Y, Mitsuki YY, et al. Selective transmission of R5 HIV-1 over X4 HIV-1 at the dendritic cell-T cell infectious synapse is determined by the T cell activation state. PLoS Pathogens. 2009;5(1) doi: 10.1371/journal.ppat.1000279.e1000279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Groot F, van Capel TMM, Schuitemaker JHN, Berkhout B, de Jong EC. Differential susceptibility of naïve, central memory and effector memory T cells to dendritic cell-mediated HIV-1 transmission. Retrovirology. 2006;3, article 52 doi: 10.1186/1742-4690-3-52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Petit C, Buseyne F, Boccaccio C, Abastado JP, Heard JM, Schwartz O. Nef is required for efficient HIV-1 replication in cocultures of dendritic cells and lymphocytes. Virology. 2001;286(1):225–236. doi: 10.1006/viro.2001.0984. [DOI] [PubMed] [Google Scholar]
- 252.Messmer D, Jacqué JM, Santisteban C, et al. Endogenously expressed nef uncouples cytokine and chemokine production from membrane phenotypic maturation in dendritic cells. The Journal of Immunology. 2002;169(8):4172–4182. doi: 10.4049/jimmunol.169.8.4172. [DOI] [PubMed] [Google Scholar]
- 253.St Gelais C, Coleman CM, Wang JH, Wu L. HIV-1 Nef enhances dendritic cell-mediated viral transmission to CD4+ T cells and promotes T-cell activation. PLoS One. 2012;7(3) doi: 10.1371/journal.pone.0034521.e34521 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 254.Coleman CM, Spearman P, Wu L. Tetherin does not significantly restrict dendritic cell-mediated HIV-1 transmission and its expression is upregulated by newly synthesized HIV-1 Nef. Retrovirology. 2011;8, article 26 doi: 10.1186/1742-4690-8-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 255.Frankel SS, Steinman RM, Michael NL, et al. Neutralizing monoclonal antibodies block human immunodeficiency virus type 1 infection of dendritic cells and transmission to T cells. Journal of Virology. 1998;72(12):9788–9794. doi: 10.1128/jvi.72.12.9788-9794.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.van Montfort T, Thomas AAM, Pollakis G, Paxton WA. Dendritic cells preferentially transfer CXCR4-using human immunodeficiency virus type 1 variants to CD4+ T lymphocytes in trans. Journal of Virology. 2008;82(16):7886–7896. doi: 10.1128/JVI.00245-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Sagar M, Akiyama H, Etemad B, Ramirez N, Freitas I, Gummuluru S. Transmembrane domain membrane proximal external region but not surface unit-directed broadly neutralizing HIV-1 antibodies can restrict dendritic cell-mediated HIV-1 trans-infection. The Journal of Infectious Diseases. 2012;205(8):1248–1257. doi: 10.1093/infdis/jis183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Mailliard RB, Alber SM, Shen H, et al. IL-18-induced CD83+CCR7+ NK helper cells. Journal of Experimental Medicine. 2005;202(7):941–953. doi: 10.1084/jem.20050128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 259.Mailliard RB, Son YI, Redlinger R, et al. Dendritic cells mediate NK cell help for Th1 and CTL responses: two-signal requirement for the induction of NK cell helper function. The Journal of Immunology. 2003;171(5):2366–2373. doi: 10.4049/jimmunol.171.5.2366. [DOI] [PubMed] [Google Scholar]
- 260.Valentin-Torres A, Ramirez Kitchen CM, Haller HS, Bernstein HB. Bidirectional NK/DC interactions promote CD4 expression on NK cells, DC maturation, and HIV infection. Virology. 2012;433(1):203–215. doi: 10.1016/j.virol.2012.06.023. [DOI] [PubMed] [Google Scholar]
- 261.Funke J, Durr R, Dietrich U, Koch J. Natural killer cells in HIV-1 infection: a double-edged sword. AIDS Review. 2011;13(2):67–76. [PubMed] [Google Scholar]
- 262.Melki MT, Saïdi H, Dufour A, Olivo-Marin JC, Gougeon ML. Escape of HIV-1-infected dendritic cells from TRAIL-mediated NK cell cytotoxicity during NK-DC cross-talk—a pivotal role of HMGB1. PLoS Pathogens. 2010;6(4) doi: 10.1371/journal.ppat.1000862.e1000862 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 263.Fredricks DN, Fiedler TL, Marrazzo JM. Molecular identification of bacteria associated with bacterial vaginosis. The New England Journal of Medicine. 2005;353(18):1899–1911. doi: 10.1056/NEJMoa043802. [DOI] [PubMed] [Google Scholar]
- 264.Myer L, Denny L, Telerant R, de Souza M, Wright TC, Jr., Kuhn L. Bacterial vaginosis and susceptibility to HIV infection in South African women: a nested case-control study. Journal of Infectious Diseases. 2005;192(8):1372–1380. doi: 10.1086/462427. [DOI] [PubMed] [Google Scholar]
- 265.St John EP, Martinson J, Simoes JA, Landay AL, Spear GT. Dendritic cell activation and maturation induced by mucosal fluid from women with bacterial vaginosis. Clinical Immunology. 2007;125(1):95–102. doi: 10.1016/j.clim.2007.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.St John EP, Zariffard MR, Martinson JA, Simoes JA, Landay AL, Spear GT. Effect of mucosal fluid from women with bacterial vaginosis on HIV trans-infection mediated by dendritic cells. Virology. 2009;385(1):22–27. doi: 10.1016/j.virol.2008.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.McDonald D, Wu L, Bohks SM, KewalRamani VN, Unutmaz D, Hope TJ. Recruitment of HIV and its receptors to dendritic cell-T cell junctions. Science. 2003;300(5623):1295–1297. doi: 10.1126/science.1084238. [DOI] [PubMed] [Google Scholar]
- 268.Tsan MF, Gao B. Endogenous ligands of Toll-like receptors. Journal of Leukocyte Biology. 2004;76(3):514–519. doi: 10.1189/jlb.0304127. [DOI] [PubMed] [Google Scholar]
- 269.Thibault S, Fromentin R, Tardif MR, Tremblay MJ. TLR2 and TLR4 triggering exerts contrasting effects with regard to HIV-1 infection of human dendritic cells and subsequent virus transfer to CD4+ T cells. Retrovirology. 2009;6, article 42 doi: 10.1186/1742-4690-6-42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 270.Huang CB, Alimova YV, Strange S, Ebersole JL. Polybacterial challenge enhances HIV reactivation in latently infected macrophages and dendritic cells. Immunology. 2011;132(3):401–409. doi: 10.1111/j.1365-2567.2010.03375.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 271.Sandler NG, Douek DC. Microbial translocation in HIV infection: causes, consequences and treatment opportunities. Nature Reviews Microbiology. 2012;10(9):655–666. doi: 10.1038/nrmicro2848. [DOI] [PubMed] [Google Scholar]
- 272.Hunt PW. Th17, gut, and HIV: therapeutic implications. Current Opinion in HIV and AIDS. 2010;5(2):189–193. doi: 10.1097/COH.0b013e32833647d9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 273.Dillon SM, Rogers LM, Howe R, et al. Human intestinal lamina propria CD1c+ dendritic cells display an activated phenotype at steady state and produce IL-23 in response to TLR7/8 stimulation. The Journal of Immunology. 2010;184(12):6612–6621. doi: 10.4049/jimmunol.1000041. [DOI] [PubMed] [Google Scholar]
- 274.Dillon SM, Manuzak JA, Leone AK, et al. HIV-1 infection of human intestinal lamina propria CD4+ T cells in vitro is enhanced by exposure to commensal escherichia coli. The Journal of Immunology. 2012;189(2):885–896. doi: 10.4049/jimmunol.1200681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Harries AD, Lawn SD, Getahun H, Zachariah R, Havlir DV. HIV and tuberculosis—science and implementation to turn the tide and reduce deaths. Journal of the International AIDS Society. 2012;15(2):p. 17396. doi: 10.7448/IAS.15.2.17396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 276.Zuniga J, Torres-Garcia D, Santos-Mendoza T, Rodriguez-Reyna TS, Granados J, Yunis EJ. Cellular and humoral mechanisms involved in the control of tuberculosis. Clinical and Developmental Immunology. 2012;2012:18 pages. doi: 10.1155/2012/193923.193923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Ehlers S. DC-SIGN and mannosylated surface structures of Mycobacterium tuberculosis: a deceptive liaison. European Journal of Cell Biology. 2010;89(1):95–101. doi: 10.1016/j.ejcb.2009.10.004. [DOI] [PubMed] [Google Scholar]
- 278.Mazurek J, Ignatowicz L, Kallenius G, Jansson M, Pawlowski A. Mycobacteria-infected bystander macrophages trigger maturation of dendritic cells and enhance their ability to mediate HIV transinfection. European Journal of Immunology. 2012;42(5):1192–1202. doi: 10.1002/eji.201142049. [DOI] [PubMed] [Google Scholar]
- 279.Reuter MA, Pecora ND, Harding CV, Canaday DH, McDonald D. Mycobacterium tuberculosis promotes HIV trans-infection and suppresses major histocompatibility complex class II antigen processing by dendritic cells. Journal of Virology. 2010;84(17):8549–8560. doi: 10.1128/JVI.02303-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Vachot L, Williams VG, Bess JW, Jr., Lifson JD, Robbiani M. Candida albicans-induced DC activation partially restricts HIV amplification in DCs and increases DC to T-cell spread of HIV. Journal of Acquired Immune Deficiency Syndromes. 2008;48(4):398–407. doi: 10.1097/QAI.0b013e3181776bc7. [DOI] [PubMed] [Google Scholar]
- 281.Qin Y, Li Y, Liu W, et al. Penicillium marneffei-stimulated dendritic cells enhance HIV-1 trans-infection and promote viral infection by activating primary CD4+ T cells. PLoS One. 2011;6(11) doi: 10.1371/journal.pone.0027609.e27609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Qin Y, Li YY, Jiang AP, Jiang JF, Wang JH. Stimulation of Cryptococcus neoformans isolated from skin lesion of AIDS patient matures dendritic cells and promotes HIV-1 trans-infection. Biochemical and Biophysical Research Communications. 2012;423(4):709–714. doi: 10.1016/j.bbrc.2012.06.020. [DOI] [PubMed] [Google Scholar]
- 283.Gonzalez R, Ataide R, Naniche D, Menendez C, Mayor A. HIV and malaria interactions: where do we stand? Expert Review of Anti-Infective Therapy. 2012;10(2):153–165. doi: 10.1586/eri.11.167. [DOI] [PubMed] [Google Scholar]
- 284.Andreani G, Lodge R, Richard D, Tremblay MJ. Mechanisms of interaction between protozoan parasites and HIV. Current Opinion in HIV and AIDS. 2012;7(3):276–282. doi: 10.1097/COH.0b013e32835211e9. [DOI] [PubMed] [Google Scholar]
- 285.Diou J, Tardif MR, Barat C, Tremblay MJ. Dendritic cells derived from hemozoin-loaded monocytes display a partial maturation phenotype that promotes HIV-1 trans-infection of CD4+ T cells and virus replication. The Journal of Immunology. 2010;184(6):2899–2907. doi: 10.4049/jimmunol.0901513. [DOI] [PubMed] [Google Scholar]
- 286.Diou J, Tardif MR, Barat C, Tremblay MJ. Malaria hemozoin modulates susceptibility of immature monocyte-derived dendritic cells to HIV-1 infection by inducing a mature-like phenotype. Cellular Microbiology. 2010;12(5):615–625. doi: 10.1111/j.1462-5822.2009.01420.x. [DOI] [PubMed] [Google Scholar]
- 287.Zhao C, Cantin R, Breton M, Papadopoulou B, Tremblay MJ. DC-SIGN-mediated transfer of HIV-1 is compromised by the ability of Leishmania infantum to exploit DC-SIGN as a ligand. Journal of Infectious Diseases. 2005;191(10):1665–1669. doi: 10.1086/429673. [DOI] [PubMed] [Google Scholar]
- 288.Haase AT. Targeting early infection to prevent HIV-1 mucosal transmission. Nature. 2010;464(7286):217–223. doi: 10.1038/nature08757. [DOI] [PubMed] [Google Scholar]
- 289.Ploquin MJ, Jacquelin B, Jochems SP, Barre-Sinoussi F, Muller-Trutwin MC. Innate immunity in the control of HIV/AIDS: recent advances and open questions. AIDS. 2012;26(10):1269–1279. doi: 10.1097/QAD.0b013e328353e46b. [DOI] [PubMed] [Google Scholar]
- 290.da Silva RC, Segat L, Crovella S. Role of DC-SIGN and L-SIGN receptors in HIV-1 vertical transmission. Human Immunology. 2011;72(4):305–311. doi: 10.1016/j.humimm.2011.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291.Ceballos A, Lenicov FR, Sabatté J, et al. Spermatozoa capture HIV-1 through heparan sulfate and efficiently transmit the virus to dendritic cells. Journal of Experimental Medicine. 2009;206(12):2717–2733. doi: 10.1084/jem.20091579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Shen R, Richter HE, Smith PD. Early HIV-1 target cells in human vaginal and ectocervical mucosa. American Journal of Reproductive Immunology. 2011;65(3):261–267. doi: 10.1111/j.1600-0897.2010.00939.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293.Sabatté J, Ceballos A, Raiden S, et al. Human seminal plasma abrogates the capture and transmission of human immunodeficiency virus type 1 to CD4+ T cells mediated by DC-SIGN. Journal of Virology. 2007;81(24):13723–13734. doi: 10.1128/JVI.01079-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 294.Ochiel DO, Ochsenbauer C, Kappes JC, Ghosh M, Fahey JV, Wira CR. Uterine epithelial cell regulation of DC-SIGN expression inhibits transmitted/founder HIV-1 trans infection by immature dendritic cells. PLoS One. 2010;5(12) doi: 10.1371/journal.pone.0014306.e14306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295.Fontenot D, He H, Hanabuchi S, et al. TSLP production by epithelial cells exposed to immunodeficiency virus triggers DC-mediated mucosal infection of CD4+ T cells. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(39):16776–16781. doi: 10.1073/pnas.0907347106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Murooka TT, Deruaz M, Marangoni F, et al. HIV-infected T cells are migratory vehicles for viral dissemination. Nature. 2012;490(7419):283–287. doi: 10.1038/nature11398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Njai HF, Lewi PJ, Janssen CGM, et al. Pre-incubation of cell-free HIV-1 group M isolates with non-nucleoside reverse transcriptase inhibitors blocks subsequent viral replication in co-cultures of dendritic cells and T cells. Antiviral Therapy. 2005;10(2):255–262. [PubMed] [Google Scholar]
- 298.Stefanidou M, Herrera C, Armanasco N, Shattock RJ. Saquinavir inhibits early events associated with establishment of HIV-1 infection: potential role for protease inhibitors in prevention. Antimicrob Agents Chemother. 2012;56(8):4381–4390. doi: 10.1128/AAC.00399-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Frank I, Robbiani M. Attachment and fusion inhibitors potently prevent dendritic cell-driven HIV infection. Journal of Acquired Immune Deficiency Syndromes. 2011;56(3):204–212. doi: 10.1097/QAI.0b013e3181ff2aa5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300.VanCompernolle SE, Taylor RJ, Oswald-Richter K, et al. Antimicrobial peptides from amphibian skin potently inhibit human immunodeficiency virus infection and transfer of virus from dendritic cells to T cells. Journal of Virology. 2005;79(18):11598–11606. doi: 10.1128/JVI.79.18.11598-11606.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Terrazas-Aranda K, van Herrewege Y, Hazuda D, et al. Human immunodeficiency virus type 1 (HIV-1) integration: a potential target for microbicides to prevent cell-free or cell-associated HIV-1 infection. Antimicrobial Agents and Chemotherapy. 2008;52(7):2544–2554. doi: 10.1128/AAC.01627-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Garg R, Tremblay MJ. Miltefosine represses HIV-1 replication in human dendritic cell/T-cell cocultures partially by inducing secretion of type-I interferon. Virology. 2012;432(2):271–276. doi: 10.1016/j.virol.2012.05.032. [DOI] [PubMed] [Google Scholar]
- 303.Chang TL, Teleshova N, Rapista A, et al. SAMMA, a mandelic acid condensation polymer, inhibits dendritic cell-mediated HIV transmission. FEBS Letters. 2007;581(24):4596–4602. doi: 10.1016/j.febslet.2007.08.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 304.Frank I, Stössel H, Gettie A, et al. A fusion inhibitor prevents spread of immunodeficiency viruses, but not activation of virus-specific T cells, by dendritic cells. Journal of Virology. 2008;82(11):5329–5339. doi: 10.1128/JVI.01987-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305.Nabatov AA, de Jong MAWP, de Witte L, Bulgheresi S, Geijtenbeek TBH. C-type lectin Mermaid inhibits dendritic cell mediated HIV-1 transmission to CD4+ T cells. Virology. 2008;378(2):323–328. doi: 10.1016/j.virol.2008.05.025. [DOI] [PubMed] [Google Scholar]
- 306.Teleshova N, Chang T, Profy A, Klotman ME. Inhibitory effect of PRO 2000, a candidate microbicide, on dendritic cell-mediated human immunodeficiency virus transfer. Antimicrobial Agents and Chemotherapy. 2008;52(5):1751–1758. doi: 10.1128/AAC.00707-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 307.Du T, Hu K, Yang J, et al. Bifunctional CD4-DC-SIGN fusion proteins demonstrate enhanced avidity to gp120 and inhibit HIV-1 infection and dissemination. Antimicrob Agents Chemother. 2012;56(9):4640–4649. doi: 10.1128/AAC.00623-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 308.Berzi A, Reina JJ, Ottria R, et al. A glycomimetic compound inhibits DC-SIGN-mediated HIV infection in cellular and cervical explant models. AIDS. 2012;26(2):127–137. doi: 10.1097/QAD.0b013e32834e1567. [DOI] [PubMed] [Google Scholar]
- 309.Serrano-Gómez D, Martínez-Nunez RT, Sierra-Filardi E, et al. AM3 modulates dendritic cell pathogen recognition capabilities by targeting DC-SIGN. Antimicrobial Agents and Chemotherapy. 2007;51(7):2313–2323. doi: 10.1128/AAC.01289-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Turville SG, Vermeire K, Balzarini J, Schols D. Sugar-binding proteins potently inhibit dendritic cell human immunodeficiency virus type 1 (HIV-1) infection and dendritic-cell-directed HIV-1 transfer. Journal of Virology. 2005;79(21):13519–13527. doi: 10.1128/JVI.79.21.13519-13527.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Balzarini J, van Herrewege Y, Vermeire K, Vanham G, Schols D. Carbohydrate-binding agents efficiently prevent dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin (DC-SIGN)-directed HIV-1 transmission to T lymphocytes. Molecular Pharmacology. 2007;71(1):3–11. doi: 10.1124/mol.106.030155. [DOI] [PubMed] [Google Scholar]
- 312.Wang SK, Liang PH, Astronomo RD, et al. Targeting the carbohydrates on HIV-1: interaction of oligomannose dendrons with human monoclonal antibody 2G12 and DC-SIGN. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(10):3690–3695. doi: 10.1073/pnas.0712326105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Sattin S, Daghett A, Thépaut M, et al. Inhibition of DC-SIGN-mediated HIV infection by a linear trimannoside mimic in a tetravalent presentation. ACS Chemical Biology. 2010;5(3):301–312. doi: 10.1021/cb900216e. [DOI] [PubMed] [Google Scholar]
- 314.Dehuyser L, Schaeffer E, Chaloin O, Mueller CG, Baati R, Wagner A. Synthesis of novel mannoside glycolipid conjugates for inhibition of HIV-1 trans-infection. Bioconjug Chem. 2012;23(9):1731–1739. doi: 10.1021/bc200644d. [DOI] [PubMed] [Google Scholar]
- 315.Nasreddine N, Borde C, Gozlan J, Bélec L, Maréchal V, Hocini H. Advanced glycation end products inhibit both infection and transmission in trans of HIV-1 from monocyte-derived dendritic cells to autologous T cells. The Journal of Immunology. 2011;186(10):5687–5695. doi: 10.4049/jimmunol.0902517. [DOI] [PubMed] [Google Scholar]
- 316.Siegal FP, Kadowaki N, Shodell M, et al. The nature of the principal Type 1 interferon-producing cells in human blood. Science. 1999;284(5421):1835–1837. doi: 10.1126/science.284.5421.1835. [DOI] [PubMed] [Google Scholar]
- 317.Ferbas JJ, Toso JF, Logar AJ, Navratil JS, Rinaldo CR. CD4+ blood dendritic cells are potent producers of IFN-α in response to in vitro HIV-1 infection. The Journal of Immunology. 1994;152(9):4649–4662. [PubMed] [Google Scholar]
- 318.Fitzgerald-Bocarsly P, Dai J, Singh S. Plasmacytoid dendritic cells and type I IFN: 50 years of convergent history. Cytokine and Growth Factor Reviews. 2008;19(1):3–19. doi: 10.1016/j.cytogfr.2007.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319.Fitzgerald-Bocarsly P, Jacobs ES. Plasmacytoid dendritic cells in HIV infection: striking a delicate balance. Journal of Leukocyte Biology. 2010;87(4):609–620. doi: 10.1189/jlb.0909635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Tel J, van der Leun AM, Figdor CG, Torensma R, de Vries IJ. Harnessing human plasmacytoid dendritic cells as professional APCs. Cancer Immunol Immunother. 2012;61(8):1279–1288. doi: 10.1007/s00262-012-1210-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Patterson S, Rae A, Hockey N, Gilmour J, Gotch F. Plasmacytoid dendritic cells are highly susceptible to human immunodeficiency virus type 1 infection and release infectious virus. Journal of Virology. 2001;75(14):6710–6713. doi: 10.1128/JVI.75.14.6710-6713.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322.Schmitt N, Nugeyre MT, Scott-Algara D, et al. Differential susceptibility of human thymic dendritic cell subsets to X4 and R5 HIV-1 infection. AIDS. 2006;20(4):533–542. doi: 10.1097/01.aids.0000210607.63138.bc. [DOI] [PubMed] [Google Scholar]
- 323.Evans VA, Lal L, Akkina R, et al. Thymic plasmacytoid dendritic cells are susceptible to productive HIV-1 infection and efficiently transfer R5 HIV-1 to thymocytes in vitro. Retrovirology. 2011;8, article 43 doi: 10.1186/1742-4690-8-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Fong L, Mengozzi M, Abbey NW, Herndier BG, Engleman EG. Productive infection of plasmacytoid dendritic cells with human immunodeficiency virus type 1 is triggered by CD40 ligation. Journal of Virology. 2002;76(21):11033–11041. doi: 10.1128/JVI.76.21.11033-11041.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Loré K, Smed-Sörensen A, Vasudevan J, Mascola JR, Koup RA. Myeloid and plasmacytoid dendritic cells transfer HIV-1 preferentially to antigen-specific CD4+ T cells. Journal of Experimental Medicine. 2005;201(12):2023–2033. doi: 10.1084/jem.20042413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 326.Groot F, van Capel TMM, Kapsenberg ML, Berkhout B, de Jong EC. Opposing roles of blood myeloid and plasmacytoid dendritic cells in HIV-1 infection of T cells: transmission facilitation versus replication inhibition. Blood. 2006;108(6):1957–1964. doi: 10.1182/blood-2006-03-010918. [DOI] [PubMed] [Google Scholar]
- 327.Mann DL, Gartner S, Le Sane F, Buchow H, Popovic M. HIV-1 transmission and function of virus-infected monocytes/macrophages. The Journal of Immunology. 1990;144(6):2152–2158. [PubMed] [Google Scholar]
- 328.Carr JM, Hocking H, Li P, Burrell CJ. Rapid and efficient cell-to-cell transmission of human immunodeficiency virus infection from monocyte-derived macrophages to peripheral blood lymphocytes. Virology. 1999;265(2):319–329. doi: 10.1006/viro.1999.0047. [DOI] [PubMed] [Google Scholar]
- 329.Duncan CJ, Sattentau QJ. Viral determinants of HIV-1 macrophage tropism. Viruses. 2011;3(11):2255–2279. doi: 10.3390/v3112255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 330.Goodenow MM, Collman RG. HIV-1 coreceptor preference is distinct from target cell tropism: a dual-parameter nomenclature to define viral phenotypes. Journal of Leukocyte Biology. 2006;80(5):965–972. doi: 10.1189/jlb.0306148. [DOI] [PubMed] [Google Scholar]
- 331.Soilleux EJ, Coleman N. Expression of DC-SIGN in human foreskin may facilitate sexual transmission of HIV. Journal of Clinical Pathology. 2004;57(1):77–78. doi: 10.1136/jcp.57.1.77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 332.McClure C, Bowman C, Geary I, Ryan C, Ball J, Eley A. HIV-1 co-receptor expression and epithelial immune cells of the cervix in asymptomatic women attending a genitourinary medicine clinic. HIV Medicine. 2013;14(2):108–114. doi: 10.1111/hiv.12002. [DOI] [PubMed] [Google Scholar]
- 333.Xu Y, Zhu H, Wilcox CK, et al. Blood monocytes harbor HIV type 1 strains with diversified phenotypes including macrophage-specific CCR5 virus. Journal of Infectious Diseases. 2008;197(2):309–318. doi: 10.1086/524847. [DOI] [PubMed] [Google Scholar]
- 334.Münch J, Rücker E, Ständker L, et al. Semen-derived amyloid fibrils drastically enhance HIV infection. Cell. 2007;131(6):1059–1071. doi: 10.1016/j.cell.2007.10.014. [DOI] [PubMed] [Google Scholar]
- 335.Koppensteiner H, Brack-Werner R, Schindler M. Macrophages and their relevance in human immunodeficiency virus type I infection. Retrovirology. 2012;9, article 82 doi: 10.1186/1742-4690-9-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.Chu H, Wang JJ, Qi M, et al. The intracellular virus-containing compartments in primary human macrophages are largely inaccessible to antibodies and small molecules. PLoS One. 2012;7(5) doi: 10.1371/journal.pone.0035297.e35297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337.Schneberger D, Aharonson-Raz K, Singh B. Monocyte and macrophage heterogeneity and Toll-like receptors in the lung. Cell and Tissue Research. 2011;343(1):97–106. doi: 10.1007/s00441-010-1032-2. [DOI] [PubMed] [Google Scholar]
- 338.Hanley TM, Viglianti GA. Nuclear receptor signaling inhibits HIV-1 replication in macrophages through multiple trans-repression mechanisms. Journal of Virology. 2011;85(20):10834–10850. doi: 10.1128/JVI.00789-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 339.Peng G, Greenwell-Wild T, Nares S, et al. Myeloid differentiation and susceptibility to HIV-1 are linked to APOBEC3 expression. Blood. 2007;110(1):393–400. doi: 10.1182/blood-2006-10-051763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 340.Thielen BK, McNevin JP, McElrath MJ, Hunt BVS, Klein KC, Lingappa JR. Innate immune signaling induces high levels of TC-specific deaminase activity in primary monocyte-derived cells through expression of APOBEC3A isoforms. Journal of Biological Chemistry. 2010;285(36):27753–27766. doi: 10.1074/jbc.M110.102822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 341.Bol SM, Moerland PD, Limou S, et al. Genome-wide association study identifies single nucleotide polymorphism in DYRK1A associated with replication of HIV-1 in monocyte-derived macrophages. PLoS One. 2011;6(2) doi: 10.1371/journal.pone.0017190.e17190 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 342.Sica A, Mantovani A. Macrophage plasticity and polarization: in vivo veritas. The Journal of Clinical Investigation. 2012;122(3):787–795. doi: 10.1172/JCI59643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 343.Cassol E, Cassetta L, Rizzi C, Alfano M, Poli G. M1 and M2a polarization of human monocyte-derived macrophages inhibits HIV-1 replication by distinct mechanisms. The Journal of Immunology. 2009;182(10):6237–6246. doi: 10.4049/jimmunol.0803447. [DOI] [PubMed] [Google Scholar]
- 344.Saïdi H, Magri G, Carbonneil C, Nasreddine N, Réquena M, Bélec L. IFN-γ-activated monocytes weakly produce HIV-1 but induce the recruitment of HIV-sensitive T cells and enhance the viral production by these recruited T cells. Journal of Leukocyte Biology. 2007;81(3):642–653. doi: 10.1189/jlb.0406278. [DOI] [PubMed] [Google Scholar]
- 345.Cobos-Jimenez V, Booiman T, Hamann J, Kootstra NA. Macrophages and HIV-1. Current Opinion in HIV and AIDS. 2011;6(5):385–390. doi: 10.1097/COH.0b013e3283497203. [DOI] [PubMed] [Google Scholar]
- 346.Laguette N, Sobhian B, Casartelli N, et al. SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature. 2011;474(7353):654–657. doi: 10.1038/nature10117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 347.Hrecka K, Hao C, Gierszewska M, et al. Vpx relieves inhibition of HIV-1 infection of macrophages mediated by the SAMHD1 protein. Nature. 2011;474(7353):658–661. doi: 10.1038/nature10195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 348.Seo JY, Yaneva R, Cresswell P. Viperin: a multifunctional, interferon-inducible protein that regulates virus replication. Cell Host Microbe. 2011;10(6):534–539. doi: 10.1016/j.chom.2011.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 349.Nasr N, Maddocks S, Turville SG, et al. HIV-1 infection of human macrophages directly induces viperin which inhibits viral production. Blood. 2012;120(4):778–788. doi: 10.1182/blood-2012-01-407395. [DOI] [PubMed] [Google Scholar]
- 350.Levine B, Mizushima N, Virgin HW. Autophagy in immunity and inflammation. Nature. 2011;469(7330):323–335. doi: 10.1038/nature09782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 351.Borel S, Espert L, Biard-Piechaczyk M. Macroautophagy regulation during HIV-1 infection of CD4+ T cells and macrophages. Frontiers in Immunology. 2012;3:p. 97. doi: 10.3389/fimmu.2012.00097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 352.Espert L, Varbanov M, Robert-Hebmann V, et al. Differential role of autophagy in CD4 T cells and macrophages during X4 and R5 HIV-1 infection. PLoS One. 2009;4(6) doi: 10.1371/journal.pone.0005787.e5787 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353.Kyei GB, Dinkins C, Davis AS, et al. Autophagy pathway intersects with HIV-1 biosynthesis and regulates viral yields in macrophages. Journal of Cell Biology. 2009;186(2):255–268. doi: 10.1083/jcb.200903070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354.Chehimi J, Luo Q, Azzoni L, et al. HIV-1 transmission and cytokine-induced expression of DC-SIGN in human monocyte-derived macrophages. Journal of Leukocyte Biology. 2003;74(5):757–763. doi: 10.1189/jlb.0503231. [DOI] [PubMed] [Google Scholar]
- 355.Rappocciolo G, Jenkins FJ, Hensler HR, et al. DC-SIGN is a receptor for human herpesvirus 8 on dendritic cells and macrophages. The Journal of Immunology. 2006;176(3):1741–1749. doi: 10.4049/jimmunol.176.3.1741. [DOI] [PubMed] [Google Scholar]
- 356.Martinez FO, Helming L, Gordon S. Alternative activation of macrophages: an immunologic functional perspective. Annual Review of Immunology. 2009;27:451–483. doi: 10.1146/annurev.immunol.021908.132532. [DOI] [PubMed] [Google Scholar]
- 357.Cassol E, Cassetta L, Alfano M, Poli G. Macrophage polarization and HIV-1 infection. Journal of Leukocyte Biology. 2010;87(4):599–608. doi: 10.1189/jlb.1009673. [DOI] [PubMed] [Google Scholar]
- 358.Cassol E, Cassetta L, Rizzi C, Gabuzda D, Alfano M, Poli G. Dendritic cell-specific ICAM-3 grabbing nonintegrin mediates HIV-1 infection of and transmission by M2a-polarized macrophages in vitro. AIDS. 2013;27(5):707–716. doi: 10.1097/QAD.0b013e32835cfc82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 359.Nguyen DG, Hildreth JEK. Involvement of macrophage mannose receptor in the binding and transmission of HIV by macrophage. European Journal of Immunology. 2003;33(2):483–493. doi: 10.1002/immu.200310024. [DOI] [PubMed] [Google Scholar]
- 360.Pollicita M, Schols D, Aquaro S, et al. Carbohydrate-binding agents (CBAs) inhibit HIV-1 infection in human primary monocyte-derived macrophages (MDMs) and efficiently prevent MDM-directed viral capture and subsequent transmission to CD4+ T lymphocytes. Virology. 2008;370(2):382–391. doi: 10.1016/j.virol.2007.08.033. [DOI] [PubMed] [Google Scholar]
- 361.Granelli-Piperno A, Pritsker A, Pack M, et al. Dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin/CD209 is abundant on macrophages in the normal human lymph node and is not required for dendritic cell stimulation of the mixed leukocyte reaction. The Journal of Immunology. 2005;175(7):4265–4273. doi: 10.4049/jimmunol.175.7.4265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 362.Boily-Larouche G, Milev MP, Zijenah LS, et al. Naturally-occurring genetic variants in human DC-SIGN increase HIV-1 capture, cell-transfer and risk of mother-to-child transmission. PLoS One. 2012;7(7) doi: 10.1371/journal.pone.0040706.e40706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 363.Creery D, Weiss W, Graziani-Bowering G, et al. Differential regulation of CXCR4 and CCR5 expression by interleukin (IL)-4 and IL-13 is associated with inhibition of chemotaxis and human immunodeficiency virus (HIV) type 1 replication but not HIV entry into human monocytes. Viral Immunology. 2006;19(3):409–423. doi: 10.1089/vim.2006.19.409. [DOI] [PubMed] [Google Scholar]
- 364.Salazar-Gonzalez JF, Salazar MG, Keele BF, et al. Genetic identity, biological phenotype, and evolutionary pathways of transmitted/founder viruses in acute and early HIV-1 infection. Journal of Experimental Medicine. 2009;206(6):1273–1289. doi: 10.1084/jem.20090378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 365.Cannon G, Yi Y, Ni H, et al. HIV envelope binding by macrophage-expressed gp340 promotes HIV-1 infection. The Journal of Immunology. 2008;181(3):2065–2070. doi: 10.4049/jimmunol.181.3.2065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 366.Sharova N, Swingler C, Sharkey M, Stevenson M. Macrophages archive HIV-1 virions for dissemination in trans. EMBO Journal. 2005;24(13):2481–2489. doi: 10.1038/sj.emboj.7600707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 367.Rudnicka D, Feldmann J, Porrot F, et al. Simultaneous cell-to-cell transmission of human immunodeficiency virus to multiple targets through polysynapses. Journal of Virology. 2009;83(12):6234–6246. doi: 10.1128/JVI.00282-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 368.Rempel H, Calosing C, Sun B, Pulliam L. Sialoadhesin expressed on IFN-induced monocytes binds HIV-1 and enhances infectivity. PLoS One. 2008;3(4) doi: 10.1371/journal.pone.0001967.e1967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 369.Gousset K, Ablan SD, Coren LV, et al. Real-time visualization of HIV-1 GAG trafficking in infected macrophages. PLoS Pathogens. 2008;4(3) doi: 10.1371/journal.ppat.1000015.e1000015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370.Groot F, Welsch S, Sattentau QJ. Efficient HIV-1 transmission from macrophages to T cells across transient virological synapses. Blood. 2008;111(9):4660–4663. doi: 10.1182/blood-2007-12-130070. [DOI] [PubMed] [Google Scholar]
- 371.Koppensteiner H, Banning C, Schneider C, Hohenberg H, Schindler M. Macrophage internal HIV-1 is protected from neutralizing antibodies. Journal of Virology. 2012;86(5):2826–2836. doi: 10.1128/JVI.05915-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 372.Kimura S, Hase K, Ohno H. The molecular basis of induction and formation of tunneling nanotubes. Cell and Tissue Research. 2013;352(1):67–76. doi: 10.1007/s00441-012-1518-1. [DOI] [PubMed] [Google Scholar]
- 373.Sherer NM, Lehmann MJ, Jimenez-Soto LF, Horensavitz C, Pypaert M, Mothes W. Retroviruses can establish filopodial bridges for efficient cell-to-cell transmission. Nature Cell Biology. 2007;9(3):310–315. doi: 10.1038/ncb1544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 374.Eugenin EA, Gaskill PJ, Berman JW. Tunneling nanotubes (TNT) are induced by HIV-infection of macrophages: a potential mechanism for intercellular HIV trafficking. Cellular Immunology. 2009;254(2):142–148. doi: 10.1016/j.cellimm.2008.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 375.Kadiu I, Gendelman HE. Human immunodeficiency virus type 1 endocytic trafficking through macrophage bridging conduits facilitates spread of infection. Journal of NeuroImmune Pharmacology. 2011;6(4):658–675. doi: 10.1007/s11481-011-9298-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 376.Kadiu I, Gendelman HE. Macrophage bridging conduit trafficking of HIV-1 through the endoplasmic reticulum and golgi network. Journal of Proteome Research. 2011;10(7):3225–3238. doi: 10.1021/pr200262q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 377.Xu W, Santini PA, Sullivan JS, et al. HIV-1 evades virus-specific IgG2 and IgA responses by targeting systemic and intestinal B cells via long-range intercellular conduits. Nature Immunology. 2009;10(9):1008–1017. doi: 10.1038/ni.1753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 378.Davis JA, Yawetz S. Management of HIV in the pregnant woman. Clinical Obstetrics and Gynecology. 2012;55(2):531–540. doi: 10.1097/GRF.0b013e31824f3ae1. [DOI] [PubMed] [Google Scholar]
- 379.van de Perre P, Rubbo PA, Viljoen J, et al. HIV-1 reservoirs in breast milk and challenges to elimination of breast-feeding transmission of HIV-1. Science Translational Medicine. 2012;4(143) doi: 10.1126/scitranslmed.3003327.143sr3 [DOI] [PubMed] [Google Scholar]
- 380.Naarding MA, Ludwig IS, Groot F, et al. Lewis X component in human milk binds DC-SIGN and inhibits HIV-1 transfer to CD4+ T lymphocytes. Journal of Clinical Investigation. 2005;115(11):3256–3264. doi: 10.1172/JCI25105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 381.Naarding MA, Dirac AM, Ludwig IS, et al. Bile salt-stimulated lipase from human milk binds DC-SIGN and inhibits human immunodeficiency virus type 1 transfer to CD4+ T cells. Antimicrobial Agents and Chemotherapy. 2006;50(10):3367–3374. doi: 10.1128/AAC.00593-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 382.Requena M, Bouhlal H, Nasreddine N, et al. Inhibition of HIV-1 transmission in trans from dendritic cells to CD4+ T lymphocytes by natural antibodies to the CRD domain of DC-SIGN purified from breast milk and intravenous immunoglobulins. Immunology. 2008;123(4):508–518. doi: 10.1111/j.1365-2567.2007.02717.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 383.Yagi Y, Watanabe E, Watari E, et al. Inhibition of DC-SIGN-mediated transmission of human immunodeficiency virus type 1 by Toll-like receptor 3 signalling in breast milk macrophages. Immunology. 2010;130(4):597–607. doi: 10.1111/j.1365-2567.2010.03264.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 384.Moir S, Fauci AS. B cells in HIV infection and disease. Nature Reviews Immunology. 2009;9(4):235–245. doi: 10.1038/nri2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 385.Clapham PR, McKnight A. Cell surface receptors, virus entry and tropism of primate lentiviruses. Journal of General Virology. 2002;83, part 8:1809–1829. doi: 10.1099/0022-1317-83-8-1809. [DOI] [PubMed] [Google Scholar]
- 386.Moir S, Malaspina A, Li Y, et al. B cells of HIV-1-infected patients bind virions through CD21-complement interactions and transmit infectious virus to activated T cells. Journal of Experimental Medicine. 2000;192(5):637–646. doi: 10.1084/jem.192.5.637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 387.Malaspina A, Moir S, Nickle DC, et al. Human immunodeficiency virus type 1 bound to B cells: relationship to virus replicating in CD4+ T cells and circulating in plasma. Journal of Virology. 2002;76(17):8855–8863. doi: 10.1128/JVI.76.17.8855-8863.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 388.Jakubik JJ, Saifuddin M, Takefman DM, Spear GT. Immune complexes containing human immunodeficiency virus type 1 primary isolates bind to lymphoid tissue B lymphocytes and are infectious for T lymphocytes. Journal of Virology. 2000;74(1):552–555. doi: 10.1128/jvi.74.1.552-555.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 389.Döpper S, Wilflingseder D, Prodinger WM, et al. Mechanism(s) promoting HIV-1 infection of primary unstimulated T lymphocytes in autologous B cell/T cell co-cultures. European Journal of Immunology. 2003;33(8):2098–2107. doi: 10.1002/eji.200323932. [DOI] [PubMed] [Google Scholar]
- 390.Jakubik JJ, Saifuddin M, Takefman DM, Spear GT. B lymphocytes in lymph nodes and peripheral blood are important for binding immune complexes containing HIV-1. Immunology. 1999;96(4):612–619. doi: 10.1046/j.1365-2567.1999.00304.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 391.Smith BA, Gartner S, Liu Y, et al. Persistence of infectious HIV on follicular dendritic cells. The Journal of Immunology. 2001;166(1):690–696. doi: 10.4049/jimmunol.166.1.690. [DOI] [PubMed] [Google Scholar]
- 392.Keele BF, Tazi L, Gartner S, et al. Characterization of the follicular dendritic cell reservoir of human immunodeficiency virus type 1. Journal of Virology. 2008;82(11):5548–5561. doi: 10.1128/JVI.00124-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 393.Thacker TC, Zhou X, Estes JD, et al. Follicular dendritic cells and human immunodeficiency virus type 1 transcription in CD4+ T cells. Journal of Virology. 2009;83(1):150–158. doi: 10.1128/JVI.01652-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 394.He B, Qiao X, Klasse PJ, et al. HIV-1 envelope triggers polyclonal Ig class switch recombination through a CD40-independent mechanism involving BAFF and C-type lectin receptors. The Journal of Immunology. 2006;176(7):3931–3941. doi: 10.4049/jimmunol.176.7.3931. [DOI] [PubMed] [Google Scholar]
- 395.Rappocciolo G, Piazza P, Fuller CL, et al. DC-SIGN on B lymphocytes is required for transmission of HIV-1 to T lymphocytes. PLoS Pathogens. 2006;2(7):p. e70. doi: 10.1371/journal.ppat.0020070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 396.Wu L, Martin TD, Carrington M, KewalRamani VN. Raji B cells, misidentified as THP-1 cells, stimulate DC-SIGN-mediated HIV transmission. Virology. 2004;318(1):17–23. doi: 10.1016/j.virol.2003.09.028. [DOI] [PubMed] [Google Scholar]
- 397.Stamataki Z, Shannon-Lowe C, Shaw J, et al. Hepatitis C virus association with peripheral blood B lymphocytes potentiates viral infection of liver-derived hepatoma cells. Blood. 2009;113(3):585–593. doi: 10.1182/blood-2008-05-158824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 398.Martin G, Roy J, Barat C, Ouellet M, Gilbert C, Tremblay MJ. Human immunodeficiency virus type 1-associated CD40 ligand transactivates B lymphocytes and promotes infection of CD4+ T cells. Journal of Virology. 2007;81(11):5872–5881. doi: 10.1128/JVI.02542-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 399.Imbeault M, Ouellet M, Giguère K, et al. Acquisition of host-derived CD40L by HIV-1 in vivo and its functional consequences in the B-cell compartment. Journal of Virology. 2011;85(5):2189–2200. doi: 10.1128/JVI.01993-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 400.Holl V, Xu K, Peressin M, et al. Stimulation of HIV-1 replication in immature dendritic cells in contact with primary CD4 T or B lymphocytes. Journal of Virology. 2010;84(9):4172–4182. doi: 10.1128/JVI.01567-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 401.Rickert RC, Jellusova J, Miletic AV. Signaling by the TNFR superfamily in B-cell biology and disease. Immunological Reviews. 2011;244(1):115–133. doi: 10.1111/j.1600-065X.2011.01067.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 402.Poudrier J, Chagnon-Choquet J, Roger M. Influence of dendritic cells on B-cell responses during HIV infection. Clinical and Developmental Immunology. 2012;2012:10 pages. doi: 10.1155/2012/592187.592187 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 403.Browne EP. Regulation of B-cell responses by Toll-like receptors. Immunology. 2012;136(4):370–379. doi: 10.1111/j.1365-2567.2012.03587.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 404.Uccellini MB, Avalos AM, Marshak-Rothstein A, Viglianti GA. Toll-like receptor-dependent immune complex activation of B cells and dendritic cells. Methods in Molecular Biology. 2009;517:363–380. doi: 10.1007/978-1-59745-541-1_22. [DOI] [PubMed] [Google Scholar]
- 405.Swingler S, Brichacek B, Jacque JM, Ulich C, Zhou J, Stevenson M. HIV-1 Nef intersects the macrophage CD40L signalling pathway to promote resting-cell infection. Nature. 2003;424(6945):213–219. doi: 10.1038/nature01749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 406.Walker BD. Elite control of HIV Infection: implications for vaccines and treatment. Topics in HIV Medicine. 2007;15(4):134–136. [PubMed] [Google Scholar]
- 407.Okulicz JF, Lambotte O. Epidemiology and clinical characteristics of elite controllers. Current Opinion in HIV and AIDS. 2011;6(3):163–168. doi: 10.1097/COH.0b013e328344f35e. [DOI] [PubMed] [Google Scholar]
- 408.Autran B, Descours B, Avettand-Fenoel V, Rouzioux C. Elite controllers as a model of functional cure. Current Opinion in HIV and AIDS. 2011;6(3):181–187. doi: 10.1097/COH.0b013e328345a328. [DOI] [PubMed] [Google Scholar]
- 409.Huang J, Burke PS, Cung TDH, et al. Leukocyte immunoglobulin-like receptors maintain unique antigen-presenting properties of circulating myeloid dendritic cells in HIV-1-infected elite controllers. Journal of Virology. 2010;84(18):9463–9471. doi: 10.1128/JVI.01009-10. [DOI] [PMC free article] [PubMed] [Google Scholar]