

Abstract
Krüppel-like factors (KLF) and specificity proteins (SP) constitute a family of zinc-finger-containing transcription factors that play important roles in a wide range of processes including differentiation and development of various tissues. The human genome possesses 17 KLF genes (KLF1–KLF17) and nine SP genes (SP1–SP9) with diverse functions. We used sequence similarity searches and gene synteny analysis to identify a new putative KLF gene/pseudogene named KLF18 that is present in most of the placental mammals with sequenced genomes. KLF18 is a chromosomal neighbor of the KLF17 gene and is likely a product of its duplication. Phylogenetic analyses revealed that mammalian predicted KLF18 proteins and KLF17 proteins experienced elevated rates of evolution and are grouped with KLF1/KLF2/KLF4 and non-mammalian KLF17. Predicted KLF18 proteins maintain conserved features in the zinc fingers of the SP/KLF family, while possessing repeats of a unique sequence motif in their N-terminal regions. No expression data have been reported for KLF18, suggesting that it either has highly restricted expression patterns and specialized functions, or could have become a pseudogene in extant placental mammals. Besides KLF18 genes/pseudogenes, we identified several KLF18-like genes such as Zfp352, Zfp352-like, and Zfp353 in the genomes of mouse and rat. These KLF18-like genes do not possess introns inside their coding regions, and gene expression data indicate that some of them may function in early embryonic development. They represent further expansions of KLF members in the murine lineage, most likely resulted from several events of retrotransposition and local gene duplication starting from an ancient spliced mRNA of KLF18.
Introduction
Krüppel-like factors (KLF) and specificity proteins (SP) are an important family of transcription factors (SP/KLF family) under extensive research [1–3]. They possess three DNA-binding C2H2-type zinc finger domains, each of which contains two conserved cysteines and two conserved histidines for zinc binding. The three zinc finger domains and the linkers between them are well conserved in the SP/KLF family, with a cysteine-histidine pattern of “CX4CX12HX3HX7CX4CX12HX3HX7CX2CX12HX3H” (Xn: separation of n residues). The separations between the first, second, and third cysteine pairs are four residues, four residues, and two residues, respectively. Such a pattern together with the number of C2H2 domains (three) appears to be a unique feature of SP/KLF members in mammalian genomes compared to the patterns of other known C2H2-domain-containing proteins (based on an analysis of human and mouse C2H2-domain-containing proteins from the SysZNF database [4]). For example, the EGR2 protein has three zinc fingers but exhibits a different pattern of residue separations between cysteine pairs (4, 2, 2 residue separations compared to 4, 4, 2 residue separations in SP/KLF members). Wilms’ tumor protein possesses four C2H2 domains: three of them sharing the same pattern as the SP/KLF proteins (4, 4, 2 residue separations between cysteine pairs) and a C-terminal fourth C2H2 domain with a four-residue separation between the cysteines. The SP/KLF family proteins mainly recognize and bind GC-rich regions such as GC boxes and GT boxes (CACCC boxes). The structure of KLF4 zinc finger domains in complex with DNA [5] revealed conserved residues responsible for specific DNA interactions. Among them are three invariant arginines that use their guanidinium groups to form critical hydrogen bonds with three guanine bases and contribute the most to the DNA-binding specificity of KLFs. In contrast to the high sequence conservation in zinc fingers, the N-terminal regions of KLFs exhibit great sequence variation [3,6]. These regions contain short sequence motifs that mediate the interactions between KLFs and other proteins such as transcription coactivators and corepressors.
The SP/KLF family proteins regulate a diverse array of cellular processes in development, differentiation, and cell death. Some human SP/KLF members have been associated with various diseases [6]. A total of 17 KLF genes (KLF1-KLF17) and nine SP genes (SP1–SP9) are currently annotated in the human genome. Compared to KLF proteins, SPs are characterized by a unique cysteine-rich motif (“CXCPXC”, buttonhead box) in the region N-terminal to the zinc fingers. Several phylogenetic analyses of the SP/KLF family proteins [2,7] suggest that SPs form a monophyletic group and are more closely related to a subgroup of KLF proteins (e.g., KLF9/KLF13/KLF14/KLF16) than to the other KLF proteins. Members of the SP/KLF family differ in their tissue expression patterns and their functions. Some KLF genes, such as KLF3, KLF9 and KLF10, exhibit broad expression patterns, while other members are expressed in restricted tissues. For example, human KLF1 (also named erythroid KLF, or EKLF) is mostly expressed in erythroid cells and regulates their differentiation.
Of the 17 human KLF genes, three members (KLF1, KLF14, and KLF16) appear to be mammalian-specific. The other human KLF genes have orthologs in other vertebrates such as chicken, frog, and teleost fish. SP/KLF members have also been identified in metazoans outside vertebrates, albeit the number of SP/KLF genes in these species is smaller [7,8]. Two rounds of whole-genome duplications in the ancestor of vertebrates [9] may partially explain the increased number of SP/KLF genes in vertebrates. A more recent whole-genome duplication event in the ancestor of teleost fish could have resulted in highly similar KLF pairs in Danio rerio, such as KLF5a/KLF5b and KLF15a/KLF15b.
The most recent mammalian gene assigned to the SP/KLF family, KLF17 [10], was first discovered as a germ cell-specific gene encoding zinc finger protein 393 (Zfp393) in mouse [11]. Human and mouse KLF17 proteins exhibit less sequence similarity compared to other orthologous KLF protein pairs, suggesting that KLF17 has undergone rapid evolution in the mammalian lineage [10]. This relatively high sequence divergence of KLF17s compared to known KLF proteins has delayed the inference of KLF17 as a KLF member [10]. Based on gene synteny, KLF17 was also proposed to exist in non-mammalian species [12]. Specifically, mammalian, chicken, and frog KLF17 genes are sandwiched by the SLC6A9 gene and the DMAP gene [12]. Likewise, a fish KLF17 ortholog can be inferred based on the syntenic similarity of a fish gene [13] (NCBI Gene ID: 65238, previously proposed to be KLF4 [14] and later annotated as Klf4b in the NCBI gene database) to KLF17 genes in other vertebrates.
We combined sequence similarity searches, multiple sequence alignment, phylogenetic reconstruction, and gene synteny analysis for computational identification of new KLF genes/pseudogenes in mammals. We predicted a novel KLF gene or pseudogene, named KLF18, in most of placental mammals with sequenced genomes including human and mouse. Mammalian KLF18 and KLF17 are chromosomal neighbors, and their inferred protein products form a monophyletic group to the exclusion of other known KLF proteins, suggesting that KLF18 resulted from a local gene duplication of KLF17. We propose that KLF18 retrotransposition and local gene duplication resulted in further expansion of KLF members in the murine genomes of mouse and rat, giving rise to the highly diversified Zfp352, Zfp352l, and Zfp353 genes [15].
Materials and Methods
Mammalian genome selection
To study the distribution of predicted KLF18 genes/pseudogenes, we examined 44 mammalian genomes available in the UCSC genome browser [16] as of December 2012. For three species, sheep, hedgehog, and tenrec, we used their latest genome assemblies from NCBI with higher coverage of sequencing than their UCSC versions. In addition, we also analyzed NCBI genome assemblies of three recently sequenced mammalian species from the Afrotheria superorder, which like the Xenarthra superorder, is underrepresented compared to the other two placental mammalian superorders (Euarchontoglires and Laurasiatheria). The total number of mammalian genomes analyzed is 47, consisting of 43 placental mammals and four non-placental mammals (Table S1).
Protein similarity searches and phylogenetic analyses of predicted KLF proteins
BLAST [17] was used to search for close homologs of KLF proteins starting with known human KLF proteins against the nr database in NCBI (e-value cutoff: 1e-10). Multiple sequence alignment of KLF proteins was made by MAFFT [18] (options: --localpair --maxiterate 1000). For phylogenetic analyses, we selected a set of SP/KLF proteins consisting of known human KLF proteins (KLF1-KLF17) and SP proteins (SP1–SP9), three non-mammalian KLF17s (from zebrafish, frog and chicken), three predicted KLF18s (from human, mouse and rat), mouse and rat Zfp352 proteins and their close homologs, and the human Wilms’ tumor protein (WT1) as an out-group (WT1 contains four zinc fingers, three of which exhibit the same pattern as SP/KLF members and have a similar set of DNA-binding specificity residues as SP/KLF proteins). The MOLPHY package [19] was used for phylogenetic reconstruction for the zinc finger regions of these proteins. The JTT amino acid substitution model [20] was used in MOLPHY. The local estimates of bootstrap percentages were obtained by the RELL method [21] (-R option in the ProtML program of MOLPHY). For this dataset, we also used MrBayes [22] to run a Bayesian inference of phylogeny using mixed amino acid substitution model with the invgamma (invariant site + gamma distribution of rate variation) option. A total of 300,000 generations were performed, and the first 150,000 generations (50%) were discarded as burn-in. A consensus tree was obtained for the remaining generations with sampling frequency set to one sample per 100 generations. We also applied MOLPHY to a larger dataset of SP/KLF zinc finger regions consisting of known human and zebrafish SP/KLFs, a larger set of predicted KLF18 proteins, and close homologs of mouse and rat Zfp352 proteins.
Detection of predicted KLF18 genes/pseudogenes
Translated BLAT [23] was used to search for KLF18 genes/pseudogenes for UCSC genomes, and TBLASTN [24] was used for the NCBI genomes. Their chromosomal locations were further confirmed by BLAT/TBLASTN searches of KLF17 and DMAP1, two genes neighboring to the KLF18 locus. For a few species, the pseudogene status of KLF18 was inferred based on the presence of premature stop condons inside the regions encoding zinc fingers. Pre-calculated gene prediction results available in the UCSC genome browser, mostly by GENSCAN [25], were examined in regions corresponding to the predicted KLF18 genes. For genomes where such predictions are not available, FGENESH [26] was used to predict KLF18 genes. TBLASTN was further used to search for missing pieces of zinc finger regions for some predicted KLF18 genes. The gene prediction results are shown in Table S1, and the predicted KLF18 protein sequences are available in Figure S2.
Results and Discussion
KLF18 is a new predicted KLF gene/pseudogene in most of the placental mammals
BLAST sequence similarity searches using zinc finger domains of known human KLF proteins identified several predicted proteins from rabbit annotated as “PREDICTED: mCG120027-like” with e-values ( less than 1e-20) comparable to those of other SP/KLF family proteins. For example, a BLAST search using the human KLF17 protein as the query found a rabbit mCG120027-like protein (Genbank: XP_002715727.1) with an e-value of 5e-25 (score: 118 bits), which is comparable to or better than the e-values of some known KLF proteins (e.g., human KLF8 with an e-value of 1e-24 and human SP4 with an e-value of 4e-24) and better than the e-values of other zinc-finger-containing proteins such as Wilms’ tumor protein (e.g., human Wilms’ tumor protein with an e-value of 6e-19). These predicted rabbit proteins have three C-terminal zinc finger domains with the same cysteine-histidine pattern (“CX4CX12HX3HX7CX4CX12HX3HX7CX2CX12HX3H”) that is a distinct feature of the SP/KLF family proteins. The names of these rabbit proteins indicate orthology to a predicted mouse protein called mCG120027 (GenBank: EDL30545.1). Potential orthologs of mCG120027 from cow (GenBank: DAA31138.1) and a primate Otolemur garnettii (GenBank: XP_003801272.1), both derived from predicted genes, were also among the top BLAST hits of human KLF proteins. Examination of the chromosome locations of these predicted genes revealed that they have conserved gene synteny, as they are all neighbors of KLF17 genes in corresponding genomes and are oriented in a tail-to-tail fashion compared to KLF17 genes. Similarity searches against genome sequences of 47 mammals by translated BLAT/BLAST and gene predictions by GENSCAN and FGENESH (see Materials and Methods) identified a predicted mCG120027-like gene or pseudogene downstream of KLF17 in most of the placental mammals (Figure 1 and Table S1). We name these new predicted genes/pseudogenes (putative orthologs of mouse mCG120027) KLF18.
Figure 1. Chromosome localization and gene synteny of KLF17 and KLF18 in vertebrate genomes.

Chromosome (Chr) or scaffold (Sca.) numbers are shown to the left of the gene order diagrams, with ‘r’ after the chromosome number denoting the reverse strand. In most of the placental mammalian genomes, KLF17 and KLF18 are neighbors arranged in a tail-to-tail fashion, and they are sandwiched by three upstream genes (abbreviations: B: B4GALT2; C: CCDC24; and S: SLC6A9) and three downstream genes (abbreviations: D: DMAP1; E: ERI3; and R: RNF220). Such a gene context for KLF17 is largely preserved in non-mammalian vertebrates including chicken, frog, and zebrafish. Copy number expansions of KLF18 (the number of expanded genes shown beside the brackets) were observed in rat, guinea pig, and rabbit. The aardvark KLF18 with pseudogene evidence is shown with dashed outline. The tree on the left shows the relationships of mammals and other vertebrates. Roots of four major groups (superorders) of placental mammals are shown in circles - E: Euarchontoglires; L: Laurasiatheria; A: Afrotheria; and X: Xenarthra.
Is KLF18 a pseudogene or a protein-coding gene?
KLF18 was predicted to be a protein-coding gene with zinc finger regions for the majority of the examined placental mammals with sequenced genomes (36 out of 43 genomes, Table S1). KLF18 pseudogenes were inferred for four genomes of placental mammals (pig, hedgehog, tenrec, and aardvark) based on premature stop codon mutations or deteriorated zinc fingers. KLF18 zinc finger regions were not detected in only three out of the 43 placental mammal genomes. These three genomes (two primates: mouse lemur and bushbaby, and rock hyrax from the Afrotheria group) have low genome sequencing coverage (less than 3 fold) [27]. Verification of KLF18’s presence in them may require genome sequences of higher quality. For mouse lemur and rock hyrax, we did find regions with significant similarity to the N-terminal regions (containing a repeated motif, described below) of other predicted KLF18 proteins (Table S1 and Figure S2).
Despite the prevalence of KLF18 as a predicted protein-coding gene in the majority of the placental mammals analyzed, sequence database searches did not find evidence of gene expression such as cDNA and expressed sequence tags (ESTs) for these predicted KLF18 genes. Recently, techniques such as RNA-seq [28] and ribosome profiling [29] greatly expanded the data of gene expression. RNA-seq-based data (including ENCODE RNA-seq datasets [30]) supporting KLF18 expression were not found at the UCSC genome browser [16]. We also searched the NCBI Sequence Read Archive (SRA) for potential transcripts of human KLF18 and only found a few spurious hits. The lack of expression data suggests that some or all of these predicted KLF18 genes may not be expressed and may have become pseudogenes. Pseudogene evidence was available for a couple of genomes such as hedgehog and aardvark, as premature stop codons were detected inside the regions corresponding to the C2H2 zinc fingers. However, for the majority of the placental mammal genomes examined, KLF18 was predicted to be a protein-coding gene by GENSCAN or FGENESH, and their predicted coding regions lack deterioration signals commonly found in pseudogenes such as frame shifts and premature stop codons. Moreover, the predicted KLF18 proteins exhibit conserved features in the zinc finger regions as compared to known KLF proteins (Figure 2 and see Figure S1 for the alignment of zinc fingers of all predicted KLF18 proteins derived from analyzed genomes). In particular, the zinc-binding cysteines and histidines are mostly preserved. One exception is the last zinc-binding position in the mouse KLF18 (predicted protein mCG120027) (Figure 2), where the histidine is replaced by a cysteine residue. In a general C2H2 zinc finger consensus sequence, both histidine and cysteine are allowed in such a position, and thus this change may not affect the zinc-binding potential of mCG120027 if it is translated from the mouse predicted KLF18 gene.
Figure 2. Multiple sequence alignment of three zinc fingers of select KLF proteins and Wilms’ tumor proteins.

Two new KLF groups (predicted KLF18 proteins and Zfp352/Zfp352l/Zfp353) are shown above the red line. Known KLF members below the red line are grouped according to frequently well-supported clusters found in separate phylogenetic studies. Conserved cysteines and histidines involved in metal binding are on black background. Three conserved DNA base-interacting arginines are shaded in magenta. Three negatively charged residues interacting with the three arginines are shaded in grey, with connections for interaction pairs shown above the alignment. Negatively charged residues (aspartate and glutamate) and positively charged residues (lysine, arginine, and histidine) are colored red and blue, respectively. Insertion and deletion events are highlighted in cyan. Species name abbreviations are: bt, Bos taurus (cow); cf, Canis familaiaris (domestic dog); ch, Choloepus hoffmanni (two-toed sloth); cj, Callithrix jacchus (common marmoset); cp, Cavia porcellus (guinea pig); dn, Dasypus novemicinctus (nine-banded armadillo); dr, Danio rerio (zebrafish); ec, Equus caballus (horse); hs, Homo sapiens (human); la, Loxodonta Africana (African Savannah elephant); mm, Mus musculus (mouse); oc, Oryctolagus cuniculus (rabbit); rn, Rattus norvegicus (rat); sa, Sorex araneus (common shrew); tb, Tupaia belangeri (tree shrew); tm, Trichechus manatus latirostris (the Floirida manatee); xt, Xenopus tropicalis (Western clawed frog). Species names are colored as follows - black: Euarchontoglires; red: Laurasiatheria; green: Afrotheria; magenta: Xenarthra; and blue: non-mammalian vertebrates.
Besides metal-binding residues, the other parts of the zinc finger domains of the newly predicted KLF18 proteins are also well conserved compared to known KLFs (Figure 2 and Figure S1). Most interestingly, three arginine residues contributing most to the specific interactions with DNA base pairs are conserved in predicted KLF18 proteins, like other SP/KLF members (Figure 2 and Figure S1). These arginines (two in the second zinc finger and one in the third zinc finger, on magenta background in Figure 2) use their side-chain guanidinium groups to make double hydrogen bond interactions with three guanine bases in the consensus GC box/GA box motif (GGCG or GGTG) [5]. Three negatively charge residues (two aspartic acids and one glutamic acid) that help orienting the guanidinium groups of these arginines are also largely preserved in predicted KLF18 proteins (Figure 2 and Figure S1). Therefore, it is likely that predicted KLF18 proteins, if translated, are capable of DNA-binding and recognition of DNA motifs such as GC box and GT box like known KLF proteins.
The coding potential of the predicted exon regions encoding the three zinc fingers of KLF18 was probed by the program PhyloCSF [31] for ten species (human, mouse, rat, guinea pig, rabbit, cow, horse, dog, elephant, and armadillo). PhyloCSF aims to distinguish protein coding regions from non-coding regions based on codon substitution frequencies and does not rely on information of similarity to other proteins. PhyloCSF gave a positive score of 305.9 decibans (a score of N decibans corresponds to a difference of 100.1*N fold), suggesting that the protein coding model is ~1030 times more likely than the non-coding model. Such a score supports the hypothesis that KLF18 or at least its ancestral form is a protein coding gene. If KLF18 is still active in extant mammals, the lack of expression data for KLF18 suggests very low expression levels or tightly controlled spatial or temporal expression patterns.
A unique repeated motif in predicted KLF18 proteins
KLF proteins possess N-terminal regions that are highly variable compared to zinc finger regions [3,6]. Closely related KLFs often share certain short sequence motifs for protein-protein interactions inside these regions. For example, KLF3, KLF8, and KLF12 contain a CtBP-binding site with a sequence consensus of “PXDLS” [3,6,32,33], while another closely related group of KLFs (KLF9, KLF10, KLF11, KLF13, KLF14, and KLF16) contain a Sin3A-binding motif that adopts an alpha-helical structure [6,34–36]. Like other KLFs, predicted KLF18 proteins typically possess a long N-terminal region (most of them larger than 300 amino acids, Figure S2). These regions share little sequence similarity to N-terminal regions of known SP/KLF family members. One interesting feature of such regions in predicted KLF18 proteins is the presence of a unique repeated motif exhibiting the pattern of “[YC]x[sE][QH]” (x: any amino acid, s: a small residue such as Gly, Ala, Ser, Thr, Asp, Asn and Pro, Figure S2). For example, the human and mouse predicted KLF18 proteins have 50 and 14 copies of such repeats, respectively (Figure S2). The first position of this four-residue motif has a preference for tyrosine (Y) with less frequent occurrence of cysteine (C), while the last position of this motif is mostly glutamine (Q). Residue preferences were also observed in positions before and after the motif. For example, the three positions before the conserved tyrosine are most frequently occupied by Q, T, and L, respectively (see sequence logo in Figure 3). Consecutive occurrences of a 14-residue segment, consisting of the [YC]x[sE][QH] motif, five residues before it, and five residues after it, are very common, especially in primate species, e.g. human (Figure S2).
Figure 3. Sequence logo of the repeated segments in the N-terminal regions of predicted KLF18 proteins.
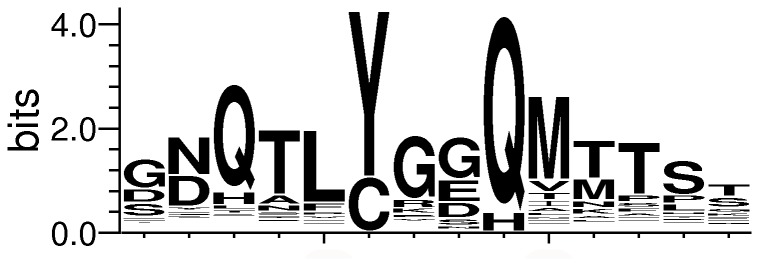
Four-residue sequence segments matching the pattern “[YC]x[sE][QH]” are extracted from predicted KLF18 proteins. These segments were extended by five residues both N-terminally and C-terminally to obtain segments of 14 residues. Sequence logo was generated for the expanded segments by the program WebLogo [44].
Searches of the human proteome with this motif pattern ([YC]x[sE][QH]) found very few proteins with a high density of this motif (motif density is defined as the number of motifs divided by protein length). Although the cysteine-rich keratins have high concentrations of this motif, their motifs have cysteines in the first position as opposed to mostly tyrosine in predicted KLF18 proteins. Another protein with a high density of this motif is RNA-binding protein 14 (GenBank: NP_006319.1). This protein possesses the “[GS]Y[GS]” repeats often found in proteins from RNA granules [37]. The [YC]x[sE][QH] motifs in this protein overlap with the [GS]Y[GS] motifs, with the residue before the tyrosine being a small residue such as glycine and serine. However, the [YC]x[sE][QH] motif in the predicted KLF18 proteins is different from the [GS]Y[GS] motif since the residue before the first position is often a large hydrophobic residue such as leucine (Figure 3 and Figure S2). As repeated patterns in proteins, such as leucine-rich repeats, heat repeats, and beta helices, are often involved in protein-protein interactions, the repeats in the N-terminal regions of predicted KLF18 proteins may also be responsible for interactions with other proteins, and they may serve to recruit transcription coactivators/corepressors to specific chromosomal locations. However, PSI-BLAST [17] and HHpred [38] searches of several these repeated regions (from human, horse, and elephant) did not yield hits with significant scores to known structures.
The origin of KLF18
We found KLF18 in species from all four major groups (superorders) [39] of the placental mammals: Euarchontoglires (including primates such as human and marmoset, rodents such as mouse and rat, and lagomorphs such as rabbit), Laurasiatheria (such as cow, horse, dog, and microbat), Afrotheria (such as elephant and manatee), and Xenarthra (such as armadillo and sloth) (Figure 1, Figure 2, Figure S1, Figure S2, and Table S1). However, sequence similarity searches and gene predictions did not reveal such a gene/pseudogene in non-mammalian vertebrates. KLF18 was also not found in non-placental mammals (marsupials and monostremes) despite the availability of several genomes of marsupials and the platypus, a monotreme (Table S1). The near-universal presence of KLF18 in placental mammals but not other genomes suggests that it may have originated in the last common ancestor of extant placental mammals.
A gene structure feature shared by KLF18, KLF17 and most of the other mammalian KLF genes (except KLF14) is an intron between the coding regions of first zinc finger and the last two zinc fingers. The intronless KLF14 gene is believed to be a product of retrotransposition from its close homolog KLF16 [40]. Presence of an intron in the predicted KLF18 genes suggests that KLF18 is not generated by retrotransposition. On the other hand, the closeness of KLF18 to KLF17 in chromosomal location (Figure 1) suggests that KLF18 could have resulted from a local gene duplication of KLF17. This scenario of KLF18 origin is also supported by phylogenetic analyses (Figure 4, Figure S3, and Figure S4, described below), as mammalian KLF17 and KLF18 form a well-supported group to the exclusion of other KLF proteins.
Figure 4. A phylogenetic tree of SP/KLF proteins with the human Wilms’ tumor protein (human_WT1) as an out-group.

Branch support values 80 or above are in bold. Each protein node is denoted by the species name followed by the protein name.
Phylogenetic positioning of KLF18 and KLF17
Previous phylogenetic studies consistently identified several well-supported groups of vertebrate KLF proteins, such as KLF1/KLF2/KLF4, KLF3/KLF8/KLF12, KLF6/KLF7, KLF10/KLF11, and KLF9/KLF13/KLF14/KLF16 [2,3,6,7,10,13]. However, the groupings among some of these KLF groups and the positions of some KLF members are not consistently recovered in separate studies. The positioning of mammalian KLF17 is not consistent in several phylogenetic studies [6,7,10,13]. Due to elevated evolutionary rate [10], mammalian KLF17s tend to form long branches in phylogenetic reconstructions. In contrast, a recent phylogenetic study revealed that non-mammalian KLF17 members do not form long branches, and they are grouped with KLF1/KLF2/KLF4 [13].
We carried out a maximum likelihood phylogenetic reconstruction (see Materials and Methods) for the zinc finger regions of known human KLF proteins, several vertebrate KLF17 proteins, and some predicted KLF18 proteins and their derivatives (Zfp352, Zfp352l, and Zfp353, described below). In this phylogenetic tree generated by MOLPHY [19], mammalian KLF17s and predicted KLF18 proteins all lie within the group of KLF1/KLF2/KLF4 (Figure 4), like the non-mammalian KLF17s. Mammalian KLF17s, predicted KLF18 proteins, and KLF18 derivatives have much longer branch lengths than other KLF members. Phylogenetic reconstructions by Bayesian analysis using the same dataset and by maximum likelihood on a larger dataset yielded similar results (Figure S3 and Figure S4).
Copy number expansions of KLF18
KLF18 was expanded in several genomes of the Glires (rodents and lagomorphs) group, including rat, guinea pig, and rabbit (Figure 1). In each of these genomes, highly similar copies of predicted KLF18 genes were discovered, suggesting that their copy number expansions have occurred recently and independently. The rat genome has four copies of KLF18 on chromosome 5, three of which are near KLF17 (Figure 1). Interestingly, predicted rat KLF18 proteins exhibit a two-residue deletion in each of the first two zinc fingers (Figure 2). Both of such deletions occur between the conserved zinc-binding cysteines (Figure 2). The resulting shorter separations (changed from four residues to two residues) between the conserved cysteines are still allowed in a general zinc finger motif. Such two residue separations are common for C2H2 zinc fingers, e.g., in the third zinc finger of the SP/KLF family members (Figure 2) as well as in members of the SNAIL family [41]. As insertions and deletions rarely occur in zinc fingers of SP/KLF proteins, the deletions in the rat KLF18 are compatible with its elevated evolutionary rate manifested by the long branch length (Figure 4).
The rabbit genome possesses six highly similar tandem predicted KLF18 genes near the KLF17 gene. For guinea pig, we did not identify predicted KLF18 genes in the assembly scaffold (Scaffold 165) that contains KLF17 and its surrounding genes (Figure 1). However, on a separate scaffold (Scaffold 635), we found at least 20 tandem repeats of predicted KLF18 genes. The highly repeated nature of this genome region may have posed challenges for its assembly in the guinea pig genome.
Expansion of KLF members in the murine genomes by retrotransposition and local gene duplication
The top BLAST hits of predicted KLF18 proteins include several mouse and rat proteins named Zfp352, Zfp353, and Zfp352-like in addition to known KLF proteins. The mRNA of the mouse Zfp352 gene (NCBI Gene ID: 236537, previously named 2czf48) was first discovered in a mouse embryonic 2-cell cDNA library [42]. Mouse Zfp353 (NCBI Gene ID: 234203), with high sequence similarity to Zfp352, was later discovered as a gene with expression restricted to lung [15]. The lack of introns inside the coding regions of Zfp352 and Zfp353, coupled with the presence of nearby LINE sequences, raised possibility that these genes are products of two consecutive retrotransposition events [15]. Zfp352 has an intron in the 5’ untranslated region, while Zfp353 does not have any introns at all. It was proposed that Zfp353 is a product of retrotransposition from the mRNA of Zfp352, and Zfp352 is a product of retrotransposition from the mRNA of an unknown gene [15]. Both mouse Zfp352 and Zfp353 encode KLF-like proteins with three C-terminal zinc fingers (Figure 2).
Several close homologs of mouse Zfp352, all without introns in coding regions, were also discovered in rat, but not in other mammalian genomes including non-murine rodents. Therefore, it is likely that Zfp352 originated in the ancestor of the Murinae subfamily. Mouse Zfp352 and rat Zfp352 (NCBI Gene ID: 502968) have conserved gene synteny, as both of them are sandwiched by the upstream Dmrta1 gene and the downstream Elavl2 gene (Figure 5A). Two predicted genes encoding close homologs of rat Zfp352 are located near the Zfp352 gene (Figure 5A). One of them is called Zfp352l (NCBI Gene ID: 298232). Zfp352l and Zfp352 are direct neighbors and are oriented in a tail-to-tail fashion (Figure 5A). The other rat predicted gene (named Zfp352lb here, NCBI Gene ID: 298233) is a direct neighbor of Zfp352l and has the same orientation as Zfp352 (Figure 5A). Rat Zfp352l and Zfp352lb, as close homologs of Zfp352, are likely generated by local gene duplication events.
Figure 5. Gene synteny and a model of evolution for Zfp352, Zfp352l, and Zfp353.

(A) Gene synteny of Zfp352, Zfp352l, and Zfp353 in the mouse and rat genomes. (B) A model of expansion of mouse KLF members. LGD and RT are abbreviations for local gene duplication and retrotransposition, respectively. UrZfp352 represents the ancestor gene of extant Zfp352 and Zfp352l.
TBLASTN searches using the mouse Zfp352 protein as the query against the mouse genome sequences also revealed a region nearby the mouse Zfp352 locus that encodes a putative gene. Similar to rat Zfp352l, this mouse predicted gene is a direct neighbor of Zfp352, and they are arranged in a tail-to-tail fashion (Figure 5A). Therefore, this predicted mouse gene should be an ortholog of the rat gene Zfp352l and is thus named mouse Zfp352l. Although mouse Zfp352l is currently listed as a pseudogene (MGI:3650768, NCBI Gene ID: 619842) in the MGI database and the NCBI gene database, it has evidence of being expressed. Its NCBI UniGene record (Mm.484218) contains one cDNA clone (RIKEN clone 7420403B16, GenBank: AK135677.1) and two ESTs (GenBank: CJ052470.1 and BB706967.1), all of which are from cDNA libraries of fertilized eggs. Interestingly, the mouse Zfp352 gene was found to be expressed in the two-cell stage of the early embryonic development (cDNA GenBank: AF290196.1; EST GenBank: AA414357.1, AA422810.1, and AI642873.1). These limited expression data suggest that mouse Zfp352 and Zfp352l may encode KLF proteins that function in early embryonic development. We did not find the counterpart of rat Zfp352lb in the mouse genome (Figure 5A), suggesting that Zfp352lb either has been lost in the mouse genome or is an invention in the rat genome.
Several lines of evidence suggest that KLF18 is the parent gene that gave rise to Zfp352/Zfp352l (intronless in coding regions) by retrotransposition of an ancestral spliced KLF18 mRNA. First, the closest KLF homologs of Zfp352 and Zfp352l are predicted KLF18 proteins. Second, Zfp352 and Zfp352l proteins are grouped with predicted KLF18 proteins in phylogenetic analyses (Figure 4, Figure S3, and Figure S4). Third, predicted KLF18 proteins, Zfp352, and Zfp352l share the repeats containing the common sequence motif [YC]x[sE][QH] that are not found in other KLF proteins (Figure S2). Inference of the ancestral KLF18 mRNA suggests that KLF18 was an actively expressed gene (transcribed and spliced to intronless mRNA) in the ancestor of mouse and rat.
Four new KLF gene/pseudogene members were discovered in the mouse genome: KLF18, Zfp352, Zfp352l, and Zfp353. Their chromosomal locations (Figure 5A) and gene structures suggest that they originated by local gene duplication (LGD) or retrotransposition (RT) in various stages of evolution since the ancestor of placental mammals. The proposed model of KLF expansion in the mouse genome is illustrated in Figure 5B. In this model, the chromosomally close Zfp352 and Zfp352l, both intronless in their coding regions, are mostly likely the results of a local gene duplication of an ancestral gene, named UrZfp352. This ancestral gene UrZfp352 likely resulted from the restrotransposition of the spliced mRNA of the ancestral KLF18 gene. KLF18 itself, being chromosomally close to KLF17, is possibly a product of local gene duplication that occurred in the ancestor of placental mammals. Zfp353, a mouse-specific gene, is not present in rat. Zfp353 is not chromosomally close to Zfp352 (Figure 5A). Its high similarity to Zfp352 and intronless gene structure suggest that Zfp353 aroused recently in the ancestor of mouse via retrotransposition of the Zfp352 mRNA [13] (Figure 5B).
While the KLF18-derived Zfp352, Zfp352l, and Zfp353 genes have been found to be expressed in certain tissues such as early embryos and lung [15,43], no expression data have been reported for the predicted KLF18 genes. The gene or pseudogene status of KLF18 remains to be experimentally investigated. Our analyses suggest that KLF18 could still be an active protein-coding gene in some extant mammals, as supported by consistent protein-coding gene predictions by GENSCAN or FGENESH across the majority of available genomes of placental mammals, conservation of zinc finger motifs including zinc-binding and DNA-binding residues, and the favorable score in protein coding potential analysis. Current unavailability of KLF18 expression data suggests that KLF18 may perform specialized functions with a tight spatial or temporal expression pattern. In the opposite scenario of KLF18 being a pseudogene, it represents an interesting case that an ancestrally active parent gene (KLF18) gave rise to currently active new genes (Zfp352, Zfp352l, and Zfp353) through retrotransposition, while the parent itself became a pseudogene in extant placental mammals.
Supporting Information
Predicted KLF18 genes in mammalian genomes. Most of the genomes are from UCSC genome browser except a few NCBI genomes not in UCSC or of later version than UCSC genomes. In the "Notes of KLF18 prediction" column, genomes where predicted KLF18 was not found are marked by red “N/A”, and inferred KLF18 pseudogenes with stop codon inside zinc finger regions are also marked in red.
(XLSX)
Alignment of zinc fingers of 36 predicted KLF18 proteins. Conserved cysteines and histidines involved in metal binding of zinc fingers are on black background. Three conserved DNA base-interacting arginines are shaded in magenta. Three negatively-charged residues interacting with the three arginines are shaded in dark grey. Substitutions in these conserved residues are colored red. Insertions and deletions are highlighted in cyan. Color coding of species names is as follows - black: Euarchontoglires; red: Laurasiatheria; green: Afrotheria; and magenta: Xenarthra.
(PDF)
Sequences of predicted KLF18 proteins (section 1) and Zfp352/Zfp352l/Zfp353 proteins (section 2). Repeats matching the regular expression of “[YC]x[GASTDNPE][QH]” (x: a single letter) are highlighted in cyan. Zinc finger regions are highlighted in magenta.
(PDF)
A phylogenetic tree of representative SP/KLF proteins with the human Wilms’ tumor protein (human_WT1) as an out-group. This tree was generated by MrBayes. Each protein node is denoted by its species name followed by the protein name.
(PDF)
A phylogenetic tree of SP/KLF proteins generated by MOLPHY. Each protein is denoted by its species name abbreviation followed by the protein name. Species name abbreviations are: bt, Bos taurus (cow); cf, Canis familaiaris (domestic dog); ch, Choloepus hoffmanni (two-toed sloth); cj, Callithrix jacchus (common marmoset); cp, Cavia porcellus (guinea pig); dn, Dasypus novemicinctus (nine-banded armadillo); do, Dipodomys ordii (kangaroo rat); dr, Danio rerio (zebrafish); ec, Equus caballus (horse); hs, Homo sapiens (human); la, Loxodonta Africana (African Savannah elephant); mm, Mus musculus (mouse); oc, Oryctolagus cuniculus (rabbit); rn, Rattus norvegicus (rat); sa, Sorex araneus (common shrew); tb, Tupaia belangeri (tree shrew); xt, Xenopus tropicalis (Western clawed frog).
(PDF)
Acknowledgments
We would like to thank Lisa Kinch for critical reading of the manuscript.
Funding Statement
This work was supported by National Institutes of Health (GM094575 to NVG) and the Welch Foundation (I-1505 to NVG). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Pearson R, Fleetwood J, Eaton S, Crossley M, Bao S (2008) Kruppel-like transcription factors: a functional family. Int J Biochem Cell Biol 40: 1996-2001. doi: 10.1016/j.biocel.2007.07.018. PubMed: 17904406. [DOI] [PubMed] [Google Scholar]
- 2. Suske G, Bruford E, Philipsen S (2005) Mammalian SP/KLF transcription factors: bring in the family. Genomics 85: 551-556. doi: 10.1016/j.ygeno.2005.01.005. PubMed: 15820306. [DOI] [PubMed] [Google Scholar]
- 3. Kaczynski J, Cook T, Urrutia R (2003) Sp1- and Kruppel-like transcription factors. Genome Biol 4: 206. doi: 10.1186/gb-2003-4-2-206. PubMed: 12620113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Ding G, Lorenz P, Kreutzer M, Li Y, Thiesen HJ (2009) SysZNF: the C2H2 zinc finger gene database. Nucleic Acids Res 37: D267-D273. doi: 10.1093/nar/gkn782. PubMed: 18974185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Schuetz A, Nana D, Rose C, Zocher G, Milanovic M et al. (2011) The structure of the Klf4 DNA-binding domain links to self-renewal and macrophage differentiation. Cell Mol Life Sci 68: 3121-3131. doi: 10.1007/s00018-010-0618-x. PubMed: 21290164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. McConnell BB, Yang VW (2010) Mammalian Kruppel-like factors in health and diseases. Physiol Rev 90: 1337-1381. doi: 10.1152/physrev.00058.2009. PubMed: 20959618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Shimeld SM (2008) C2H2 zinc finger genes of the Gli, Zic, KLF, SP, Wilms' tumour, Huckebein, Snail, Ovo, Spalt, Odd, Blimp-1, Fez and related gene families from Branchiostoma floridae. Dev Genes Evol 218: 639-649. [DOI] [PubMed] [Google Scholar]
- 8. Seetharam A, Bai Y, Stuart GW (2010) A survey of well conserved families of C2H2 zinc-finger genes in Daphnia. BMC Genomics 11: 276. doi: 10.1186/1471-2164-11-276. PubMed: 20433734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Meyer A, Schartl M (1999) Gene and genome duplications in vertebrates: the one-to-four (-to-eight in fish) rule and the evolution of novel gene functions. Curr Opin Cell Biol 11: 699-704. doi: 10.1016/S0955-0674(99)00039-3. PubMed: 10600714. [DOI] [PubMed] [Google Scholar]
- 10. van Vliet J, Crofts LA, Quinlan KG, Czolij R, Perkins AC et al. (2006) Human KLF17 is a new member of the Sp/KLF family of transcription factors. Genomics 87: 474-482. doi: 10.1016/j.ygeno.2005.12.011. PubMed: 16460907. [DOI] [PubMed] [Google Scholar]
- 11. Yan W, Burns KH, Ma L, Matzuk MM (2002) Identification of Zfp393, a germ cell-specific gene encoding a novel zinc finger protein. Mech Dev 118: 233-239. doi: 10.1016/S0925-4773(02)00258-7. PubMed: 12351194. [DOI] [PubMed] [Google Scholar]
- 12. Antin PB, Pier M, Sesepasara T, Yatskievych TA, Darnell DK (2010) Embryonic expression of the chicken Kruppel-like (KLF) transcription factor gene family. Dev Dyn 239: 1879-1887. doi: 10.1002/dvdy.22318. PubMed: 20503383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Chen Z, Lei T, Chen X, Zhang J, Yu A et al. (2010) Porcine KLF gene family: Structure, mapping, and phylogenetic analysis. Genomics 95: 111-119. doi: 10.1016/j.ygeno.2009.11.001. PubMed: 19941950. [DOI] [PubMed] [Google Scholar]
- 14. Oates AC, Pratt SJ, Vail B, Yan Y, Ho RK et al. (2001) The zebrafish klf gene family. Blood 98: 1792-1801. doi: 10.1182/blood.V98.6.1792. PubMed: 11535513. [DOI] [PubMed] [Google Scholar]
- 15. Chen HH, Liu TY, Huang CJ, Choo KB (2002) Generation of two homologous and intronless zinc-finger protein genes, zfp352 and zfp353, with different expression patterns by retrotransposition. Genomics 79: 18-23. doi: 10.1006/geno.2001.6664. PubMed: 11827453. [DOI] [PubMed] [Google Scholar]
- 16. Karolchik D, Hinrichs AS, Kent WJ (2012) The UCSC Genome Browser. Curr Protoc Bioinforma Chapter 1: 4 [DOI] [PubMed] [Google Scholar]
- 17. Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25: 3389-3402. doi: 10.1093/nar/25.17.3389. PubMed: 9254694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 30: 3059-3066. doi: 10.1093/nar/gkf436. PubMed: 12136088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Adachi J, Hasegawa M (1996) MOLPHY version 2.3, programs for molecular phylogenetics based on maximum likelihood. Computer Science Monographs 28. The Institute of Statistical Mathematics; pp. 1-150. [Google Scholar]
- 20. Jones DT, Taylor WR, Thornton JM (1992) The rapid generation of mutation data matrices from protein sequences. Comput Appl Biosci 8: 275-282. PubMed: 1633570. [DOI] [PubMed] [Google Scholar]
- 21. Hasegawa M, Kishino H, Saitou N (1991) On the maximum likelihood method in molecular phylogenetics. J Mol Evol 32: 443-445. doi: 10.1007/BF02101285. PubMed: 1904100. [DOI] [PubMed] [Google Scholar]
- 22. Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754-755. doi: 10.1093/bioinformatics/17.8.754. PubMed: 11524383. [DOI] [PubMed] [Google Scholar]
- 23. Kent WJ (2002) BLAT--the BLAST-like alignment tool. Genome Res 12: 656-664. doi: 10.1101/gr.229202. Article published online before March 2002 PubMed; : 11932250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Gertz EM, Yu YK, Agarwala R, Schäffer AA, Altschul SF (2006) Composition-based statistics and translated nucleotide searches: improving the TBLASTN module of BLAST. BMC Biol 4: 41. doi: 10.1186/1741-7007-4-41. PubMed: 17156431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Burge CB, Karlin S (1998) Finding the genes in genomic DNA. Curr Opin Struct Biol 8: 346-354. doi: 10.1016/S0959-440X(98)80069-9. PubMed: 9666331. [DOI] [PubMed] [Google Scholar]
- 26. Salamov AA, Solovyev VV (2000) Ab initio gene finding in Drosophila genomic DNA. Genome Res 10: 516-522. doi: 10.1101/gr.10.4.516. PubMed: 10779491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Lindblad-Toh K, Garber M, Zuk O, Lin MF, Parker BJ et al. (2011) A high-resolution map of human evolutionary constraint using 29 mammals. Nature 478: 476-482. doi: 10.1038/nature10530. PubMed: 21993624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Wang Z, Gerstein M, Snyder M (2009) RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10: 57-63. doi: 10.1038/nrg2484. PubMed: 19015660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Ingolia NT, Ghaemmaghami S, Newman JR, Weissman JS (2009) Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science 324: 218-223. doi: 10.1126/science.1168978. PubMed: 19213877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. ENCODE Project Consortium (2011) A user's guide to the encyclopedia of DNA elements (ENCODE). PLOS Biol 9: e1001046 PubMed: 21526222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Lin MF, Jungreis I, Kellis M (2011) PhyloCSF: a comparative genomics method to distinguish protein coding and non-coding regions. Bioinformatics 27: i275-i282. doi: 10.1093/bioinformatics/btr209. PubMed: 21685081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Turner J, Crossley M (1998) Cloning and characterization of mCtBP2, a co-repressor that associates with basic Kruppel-like factor and other mammalian transcriptional regulators. EMBO J 17: 5129-5140. doi: 10.1093/emboj/17.17.5129. PubMed: 9724649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. van Vliet J, Turner J, Crossley M (2000) Human Kruppel-like factor 8: a CACCC-box binding protein that associates with CtBP and represses transcription. Nucleic Acids Res 28: 1955-1962. doi: 10.1093/nar/28.9.1955. PubMed: 10756197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Kaczynski J, Zhang JS, Ellenrieder V, Conley A, Duenes T et al. (2001) The Sp1-like protein BTEB3 inhibits transcription via the basic transcription element box by interacting with mSin3A and HDAC-1 co-repressors and competing with Sp1. J Biol Chem 276: 36749-36756. doi: 10.1074/jbc.M105831200. PubMed: 11477107. [DOI] [PubMed] [Google Scholar]
- 35. Zhang JS, Moncrieffe MC, Kaczynski J, Ellenrieder V, Prendergast FG et al. (2001) A conserved alpha-helical motif mediates the interaction of Sp1-like transcriptional repressors with the corepressor mSin3A. Mol Cell Biol 21: 5041-5049. doi: 10.1128/MCB.21.15.5041-5049.2001. PubMed: 11438660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Kaczynski JA, Conley AA, Fernandez Zapico M, Delgado SM, Zhang JS et al. (2002) Functional analysis of basic transcription element (BTE)-binding protein (BTEB) 3 and BTEB4, a novel Sp1-like protein, reveals a subfamily of transcriptional repressors for the BTE site of the cytochrome P4501A1 gene promoter. Biochem J 366: 873-882. PubMed: 12036432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Kato M, Han TW, Xie S, Shi K, Du X et al. (2012) Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell 149: 753-767. doi: 10.1016/j.cell.2012.04.017. PubMed: 22579281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Söding J, Biegert A, Lupas AN (2005) The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res 33: W244-W248. doi: 10.1093/nar/gki162. PubMed: 15980461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Asher RJ, Helgen KM (2010) Nomenclature and placental mammal phylogeny. BMC Evol Biol 10: 102. doi: 10.1186/1471-2148-10-102. PubMed: 20406454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Parker-Katiraee L, Carson AR, Yamada T, Arnaud P, Feil R et al. (2007) Identification of the imprinted KLF14 transcription factor undergoing human-specific accelerated evolution. PLOS Genet 3: e65. doi: 10.1371/journal.pgen.0030065. PubMed: 17480121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Nieto MA (2002) The snail superfamily of zinc-finger transcription factors. Nat Rev Mol Cell Biol 3: 155-166. doi: 10.1038/nrm757. PubMed: 11994736. [DOI] [PubMed] [Google Scholar]
- 42. Choo KB, Chen HH, Cheng WT, Chang HS, Wang M (2001) In silico mining of EST databases for novel pre-implantation embryo-specific zinc finger protein genes. Mol Reprod Dev 59: 249-255. doi: 10.1002/mrd.1029. PubMed: 11424210. [DOI] [PubMed] [Google Scholar]
- 43. Liu TY, Chen HH, Lee KH, Choo KB (2003) Display of different modes of transcription by the promoters of an early embryonic gene, Zfp352, in preimplantation embryos and in somatic cells. Mol Reprod Dev 64: 52-60. doi: 10.1002/mrd.10218. PubMed: 12420299. [DOI] [PubMed] [Google Scholar]
- 44. Crooks GE, Hon G, Chandonia JM, Brenner SE (2004) WebLogo: a sequence logo generator. Genome Res 14: 1188-1190. doi: 10.1101/gr.849004. PubMed: 15173120. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Predicted KLF18 genes in mammalian genomes. Most of the genomes are from UCSC genome browser except a few NCBI genomes not in UCSC or of later version than UCSC genomes. In the "Notes of KLF18 prediction" column, genomes where predicted KLF18 was not found are marked by red “N/A”, and inferred KLF18 pseudogenes with stop codon inside zinc finger regions are also marked in red.
(XLSX)
Alignment of zinc fingers of 36 predicted KLF18 proteins. Conserved cysteines and histidines involved in metal binding of zinc fingers are on black background. Three conserved DNA base-interacting arginines are shaded in magenta. Three negatively-charged residues interacting with the three arginines are shaded in dark grey. Substitutions in these conserved residues are colored red. Insertions and deletions are highlighted in cyan. Color coding of species names is as follows - black: Euarchontoglires; red: Laurasiatheria; green: Afrotheria; and magenta: Xenarthra.
(PDF)
Sequences of predicted KLF18 proteins (section 1) and Zfp352/Zfp352l/Zfp353 proteins (section 2). Repeats matching the regular expression of “[YC]x[GASTDNPE][QH]” (x: a single letter) are highlighted in cyan. Zinc finger regions are highlighted in magenta.
(PDF)
A phylogenetic tree of representative SP/KLF proteins with the human Wilms’ tumor protein (human_WT1) as an out-group. This tree was generated by MrBayes. Each protein node is denoted by its species name followed by the protein name.
(PDF)
A phylogenetic tree of SP/KLF proteins generated by MOLPHY. Each protein is denoted by its species name abbreviation followed by the protein name. Species name abbreviations are: bt, Bos taurus (cow); cf, Canis familaiaris (domestic dog); ch, Choloepus hoffmanni (two-toed sloth); cj, Callithrix jacchus (common marmoset); cp, Cavia porcellus (guinea pig); dn, Dasypus novemicinctus (nine-banded armadillo); do, Dipodomys ordii (kangaroo rat); dr, Danio rerio (zebrafish); ec, Equus caballus (horse); hs, Homo sapiens (human); la, Loxodonta Africana (African Savannah elephant); mm, Mus musculus (mouse); oc, Oryctolagus cuniculus (rabbit); rn, Rattus norvegicus (rat); sa, Sorex araneus (common shrew); tb, Tupaia belangeri (tree shrew); xt, Xenopus tropicalis (Western clawed frog).
(PDF)