

Abstract
ERPs are commonly elicited by semantic and syntactic violations in sentences, leading to proposals that they reflect neural activity underlying ordinary language comprehension. We examined ERPs in an auditory sentence-picture-matching task, using nonanomalous sentences that were either semantically reversible, (boy pushes girl) or irreversible, (boy eats apple). Timelocked to the end of the critical clause, which occurred in the middle of a longer sentence, we observed an enhanced central-posterior positivity in response to the reversible sentences. The topography of this response is consistent with the P600 potential reported in studies of syntactic anomalies and other manipulations related to sentence structure. Following the end of the sentence, during a memory delay period prior to picture onset, reversible sentences also evoked a protracted anterior negativity, predominantly on the left. This negativity was stronger for sentences containing relative clauses compared to simple active sentences, but did not differ between object-embedded and the less complex subject-embedded clauses. The observation of a P600 occurring selectively in reversible sentences supports the interpretation of that potential as reflecting the syntactic processing of thematic relationships, as irreversible sentences contained alternative cues for thematic roles. The left anterior negativity likely reflects later processes of rehearsal and reanalysis of sentence content in working memory.
Keywords: syntax, EEG, language, ERP, N400, LAN, working memory, reanalysis
Introduction
In the cognitive neuroscience of language, a large body of work has been devoted to the study of neural responses to linguistic anomalies. Anomaly paradigms have been very fruitful, yielding robust responses that relate to distinct aspects of sentence processing. For example, the N400 potential is reliably evoked by numerous manipulations involving the semantic expectedness of a given word. Originally observed in response to semantically incongruous sentence endings (Kutas and Hillyard, 1980), it was subsequently elicited in paradigms that do not involve anomalies. N400 amplitude is closely related to the amount of semantic priming of the eliciting word (Holcomb, 1988) and the ease of its integration into the overall sentence meaning (Kutas and Federmeier, 2000). Other ERP responses are typically evoked by anomalies other than semantic expectedness. For example, the Left Anterior Negativity (LAN) and a later centro-parietal positive wave, commonly called “P600,” are often seen in response to syntactic anomalies such as violations of expected word order (Friederici et al., 1996) or inflection (Osterhout and Mobley, 1995). Subsequent studies have also demonstrated P600 effects in a wide variety of contexts that are not, strictly speaking, syntactically anomalous. These include syntactically ambiguous “garden-path” sentences (Osterhout and Holcomb, 1992, Kaan and Swaab, 2003), violations of animacy restrictions (Kuperberg et al., 2003), and various others (see discussion). The observed dissociations between the N400 and P600 responses suggest that these two ERP components may serve as indices of the engagement of distinct neural systems involved in language processing.
Although anomalies serve as useful tools in the experimental study of language, they are by definition highly atypical instances of language input. This raises an important question: do the same neural mechanisms that give rise to anomaly responses also underlie the routine operations of ordinary language comprehension? To answer this question, some researchers have sought to examine ERPs to other nonanomalous linguistic manipulations. This is somewhat more difficult, as anomalies not only produce quantitatively large responses, but the anomaly typically emerges at one unique “violation point” in a sentence. ERP studies of language generally use the onset of a critical word as a timelocking point. When a sentence becomes anomalous or ambiguous (or unambiguous) at a specific word, timelocking of the response to that word’s onset results in good alignment of the neural activity across trials, resulting in a robust ERP signal. However, neural activity related to more subtle manipulations of syntactic and semantic complexity may also emerge over a longer time scale, as the meaning of a sentence is built up over several words. In such cases, evoked potentials that are typically linked to single words, such as the N400 and the P600, may not be discernible even though the same neural networks may be involved in processing sentences over longer time scales.
Thus, studying neural activity on the time scale of portions of sentences (such as phrases and clauses) is a challenge in neuroscience. FMRI integrates activity over several seconds, limiting its usefulness in discriminating sub-sentence phenomena, while ERPs are best at revealing activity linked to individual words within a sentence. In the past, two main approaches have nonetheless been used to study activity over portions of sentences. One is to examine slow shifts in potential that emerge over the course of sentences, superimposed on the signals evoked by each individual word. This approach was used effectively by King and Kutas (1995) using visual presentation, and Muller et al. (1997) using auditory presentation, revealing sustained negativity evoked by increased syntactic complexity during a sentence comprehension task. Specifically, these studies compared two kinds of sentences, containing the more complex object-embedded relative clauses (example 1, traditionally abbreviated “SO” for subject-object word order) and the less complex subject-embedded relative clause (example 2, traditionally abbreviated “SS” for subject-subject word order).
1 – (SO) The reporter who the senator harshly attacked admitted the error.
2 – (SS) The reporter who harshly attacked the senator admitted the error.
The SO sentences violate the usual tendency in English word order to put the agent of an action first, resulting in increased demands for syntactic processing. The studies discussed here revealed ERP activity related to this increased processing demand, and further showed a relationship with individual language comprehension abilities.
A second approach to studying activity sustained over portions of a sentence is to measure oscillatory reactivity in EEG or MEG signals. Changes in oscillatory amplitude may be induced by cognitive demands but require less precise timelocking to stimulus onset than ERPs, making them suitable for activity that evolves over a few seconds, such as responses to sentences presented as natural speech. In a recent study, we used simultaneous EEG and MEG to examine oscillatory changes induced by sentences in a sentence-picture-matching task. This study built on earlier fMRI study (Meltzer et al., 2010), which examined two orthogonal factors related to syntactic processing. One factor was syntactic complexity, contrasting the two types of sentences described above, SO vs. SS. A second factor was semantic reversibility. In a typical reversible sentence, two nouns are assigned the AGENT and PATIENT roles by syntactic cues (generally word order in English), but either noun is a semantic fit for either role, as in (3):
3) The boy is pushing the girl.
In an irreversible sentence, such as (4), semantic constraints make the nouns ineligible to swap thematic roles, such that the meaning of the sentence can be adequately determined without explicit consideration of syntactic cues:
4) The boy is eating the apple.
Using fMRI, we found that reversible sentences, in the context of a sentence-picture-matching task, elicited a greater degree of activation than irreversible sentences throughout the left hemisphere perisylvian language network, as well as in the dorsal premotor and supplementary motor cortices. This effect was additive to an independent effect of syntactic complexity within reversible sentences, which induced greater responses only in frontal regions. Based on findings from this study as well as previous literature (discussed at greater length in Meltzer et al., 2010, and Meltzer and Braun, 2011), we hypothesized that the effects of syntactic complexity observed in frontal regions may relate to reprocessing a difficult sentence in verbal working memory, rather than online comprehension. To test this, we conducted an EEG-MEG study of the same task (Meltzer and Braun, 2011), focusing on oscillatory activity occurring in contrasting time periods: during the sentence presentation, and in a subsequent 3-second memory delay prior to picture presentation (figure 1A). To measure neural activity at the appropriate time scale, sentences were divided up into three portions (figure 1B), and the EEG-MEG signals were timelocked to the onset of the first word of each portion. We hypothesized that differential neural responses would be most prevalent in the third region of the sentence (post-relative clause), because that is the point when all the relevant information to distinguish the conditions has been presented (for example, “The boy who is pushing the…” could be reversible or irreversible depending on the final word of the relative clause).
Figure 1. Task Design.

A: Schematic illustration of trial structure. On each trial, subjects heard a sentence while viewing a fixation cross. After a 3 second delay, two pictures appeared, the target and a foil. B: Illustration of the placement of digital triggers within the sound files of individual sentences, to allow analysis of EEG-MEG data timelocked to specific points within and following the sentence. C: A sample picture set for the reversible sentence “The woman who the man is teaching is very tired right now.” The target shows the correct arrangement. A syntactic foil has the thematic roles of the two named actors switched, while a lexical foil substitutes one of the actors. D: A sample picture set for the nonreversible sentence “The glass that the man is washing has a small chip in it.”
In oscillatory analysis of the EEG-MEG data, we demonstrated that reversibility effects were present both during the post-RC portion of the sentence as well as the subsequent memory delay, whereas syntactic complexity affected activity only during the delay, suggesting that complexity effects are largely driven by posthoc reanalysis of language input in working memory. The oscillatory effects took the form of desynchronization in the alpha and beta frequency bands (approximately 8–30 Hz). Using beamforming analysis of the MEG data, we observed that the reversibility effects localized to left-hemisphere perisylvian regions, whereas the later effects of syntactic complexity localized to bilateral frontal regions, matching the fMRI results obtained from the same subjects.
Increased perisylvian activation for semantically reversible sentences suggests that these sentences require more extensive processing of syntactic information, in order to select the correct picture. Therefore, ERP responses associated with syntactic processing on the basis of anomaly studies may also be modulated by this factor. To our knowledge, reversible and irreversible sentences have not been directly contrasted in any previous ERP study (but see Wassenaar and Hagoort, 2007, who reported effects of sentence-picture mismatch within the two categories). Therefore, we reanalyzed our EEG data in the time domain to examine ERPs. In our previous report, we focused on the MEG data, as it is more easily subjected to beamforming analysis for source localization. Despite this advantage of MEG data, there are numerous strengths of the ERP technique that are not shared by MEG and deserve independent attention. ERP effects are characterized by a particular polarity, positive or negative, which is consistent across subjects and, to a degree, sensors. Polarity in MEG is highly dependent on sensor coil geometry and head position, and therefore highly variable across subjects due to variable head position within a fixed helmet. For this reason, MEG is typically analyzed in a polarity independent way (such as root-mean-square or time-frequency analysis), and is not easily compared with the existing ERP literature. In the present report, we find that the factor of reversibility does induce ERP responses that closely resemble components seen in previous studies of syntactic anomaly, suggesting that these components may in fact reflect activation of neural networks that are involved in routine processing of syntactic cues, even without anomaly.
METHODS
Subjects
24 healthy volunteers (12 female, age 22–37) were recruited from the local community. All were right-handed monolingual native speakers of English. Subjects gave informed consent (NIH protocol 92-DC-0178) and were financially compensated. EEG and MEG were recorded simultaneously. Oscillatory reactivity measured in EEG and MEG from the same experimental sessions was described previously (Meltzer and Braun, 2011). In the present report, we describe effects seen in event-related potentials derived from the EEG data recorded at the same time.
Materials
252 critical sentences were used for this experiment, in 6 categories, for a 2×3 factorial design (semantic reversibility × syntactic complexity). These were selected from a pool of 540 sentences constructed for this experiment and the parallel fMRI experiment. All sentences involved one or two out of four possible people, namely “the boy, the girl, the man, and the woman.” Reversible sentences (R) involved a human as both subject and object, and were constructed to avoid plausibility biases. Irreversible sentences (I) involved one human and one inanimate object. Syntactic complexity was manipulated by employing three different sentence structures: subject-embedded relative clauses (SS), object-embedded relative clauses (SO), and simple active sentences (AC) without relative clauses. The abbreviations (RSS, RSO, etc.) will be used throughout the paper to denote the six conditions. Based on previous studies, we were primarily interested in the contrast between the two types of embedded clause, which were closely matched in terms of sentence structure and lexical content, making it feasible to timelock the EEG to the onset of the embedded clause and the following segment. The AC condition served as a “filler” to reduce subjects’ habituation to the relative clause structure, but also serves as another condition of interest with a relatively simple syntactic structure.
For each level of reversibility, 45 verbs were chosen for sentence construction. All were chosen for easy imageability to facilitate the production of photographs for target and distractor stimuli. For reversible sentences, in order to avoid an over-reliance on verbs of violence (bite, pinch, kick, etc.), we also used some phrasal verbs (e.g. pick up, whisper to, sneeze on). Phrasal verbs were also used in the irreversible condition (e.g. put away, spread out, flip over). All conditions were matched for total length in words and characters. Reversible sentences were matched item-wise across conditions by using the same ending material in multiple sentences. For the irreversible sentences, it was not possible to use item-wise matching, as the ending material for the object-embedded clauses had to apply to an inanimate object instead of a person. Main verbs used in reversible and irreversible conditions were matched on word frequency, derived from the Brown corpus, (Kucera and Francis, 1967) and familiarity, taken from the MRC Psycholinguistic Database (Wilson, 1988). For other words in the sentence, it was an inevitable consequence of the design that the relative clause portion of the irreversible sentences would have lower-frequency words, given the large number of different inanimate objects occurring in those sentences, whereas “man, woman, boy, girl” are all very frequent words. We compared the mean frequency for open-class words in the second and third portions of the sentences containing relative clauses (SS and SO). As expected, a strong word-frequency difference was present during the relative clause portion of the sentence (mean Rev: 2.17 ×10−4, mean Irrev: 1.49 ×10−4, t = 3.12, p<.001), but no such effect is present during the post-relative clause segment (mean Rev: 2.00 ×10−4, mean Irrev: 1.95 ×10−4, t = 0.392, p=.70). The difference occurring during the relative clause is unlikely to affect our results, as significant differences in ERPs between reversible and irreversible sentences were only found timelocked to the post-relative clause segment and the subsequent memory delay. Furthermore, any effect of word frequency would most likely be in the opposite direction of our reported results, as the irreversible sentences had words of lower frequency, which have consistently been shown to evoke increased brain activation (Chee et al., 2002, Halgren et al., 2002). Both of our reported components, a P600-like positivity and a sustained left anterior negativity, were evoked by reversible sentences, which contained more frequent words. The most likely place for a frequency effect in our study is on the first noun of the sentence, which in ISO sentences is of much lower frequency both in general and within the context of this experiment. A strong N400 effect is indeed found here (see results), but it does not overlap in time with the effects of reversibility emerging later in the sentence.
In all sentences, the critical imageable information was presented in the second portion of the sentence. For the SS and SO conditions, this was the relative clause portion, and the third portion of the sentence, containing the main clause, presented extraneous information that was not depicted in the picture. For the AC conditions, the main clause was in the second sentence portion, while the third portion comprised an attached modifying phrase. For each verb, six sentences were generated, two of each sentence structure, with separate pools of verbs used for reversible and irreversible categories. Example sextets of sentences for two verbs are presented in table 1.
Table 1.
Example sentences in six conditions. Slashes indicate prosodic boundaries, during which a brief pause occurred.
| Code | Reversibility | Syntactic Complexity | Example sentences |
|---|---|---|---|
| RSS | Reversible | Subject-embedded clause | The man/who is teaching the woman/is discussing a hard problem. |
| The woman/who is teaching the man/is very tired right now. | |||
| RSO | Reversible | Object-embedded clause | The man/who the woman is teaching/is discussing a hard problem. |
| The woman/who the man is teaching/is very tired right now. | |||
| RAC | Reversible | Simple Active | The man/is teaching the woman/about a hard math problem. |
| The woman/is teaching the man/about a hard math problem. | |||
| ISS | Irreversible | Subject-embedded clause | The man/who is washing the car/is doing a sloppy job. |
| The woman/who is washing the glass/is not using enough soap. | |||
| ISO | Irreversible | Object-embedded clause | The glass/that the man is washing/has a small chip in it. |
| The car/that the woman is washing/has over 50,000 miles. | |||
| IAC | Irreversible | Simple Active | The man/is washing the glass/in the kitchen sink. |
| The woman/is washing the car/on a Sunday afternoon. |
Sentences were digitally recorded by a female voice artist, sampled at 44.1 kHz, and subsequently edited in Audacity software (http://audacity.sourceforge.net) as individual wav files. Spoken sentences averaged 3.45s in length. Digital triggers were manually inserted into the wav files at the boundaries of the three prosodic segments for each sentence, delineating the pre relative clause segment (“Pre-RC”), the relative clause (“RC”), and the post-relative-clause segment (“Post-RC”), as shown in figure 1B (not that these segment names do not strictly apply to the AC condition, which lacked a relative clause). These triggers were inserted into the EEG acquisition stream to allow for analysis activity timelocked to particular regions of the sentence, and also for activity occurring in the memory delay following the sentence.
For each group of six sentences involving a particular verb, we produced four photographs for the picture-matching task (figure 1C–D). The photographs all involved the same set of four actors (man, woman, boy, and girl). Subjects were familiarized with the actors before the experiment so that they could clearly distinguish them. For a reversible sentence such as “The boy who the girl is pushing hopes to win the race,” the target picture depicted a girl pushing a boy, while the syntactic foil depicted a boy pushing a girl. For trials with syntactic foils, it is necessary to use syntactic information to assign thematic roles correctly, as the same two people are mentioned in the sentence. Lexical foils involved a person not mentioned in the sentence, depicting e.g. a girl pushing a woman, or a woman pushing a girl. For irreversible sentences, only lexical foils were used, but either the human agent or the inanimate patient could be substituted. For reversible sentences, half of the picture pairs involved a syntactic foil, and half a lexical foil. For irreversible sentences, half of the pictures involved a substitution of the human agent, and half of the inanimate patient. Thus, for behavioral purposes, and including the simple active fillers, there were a total of 12 conditions (2 reversibility × 3 complexity × 2 foil type), with 21 trials of each. However, since the subjects did not know what type of foil would appear on each trial, EEG analysis of brain activity occurring prior to picture onset was collapsed across foil types, yielding 42 trials per condition.
Procedure
Subjects were seated in a padded chair inside a magnetically shielded room containing the MEG instrument. Auditory stimuli were delivered through pneumatic tubes ending in foam ear inserts, with the volume adjusted to the individual subject’s preference. Visual stimuli were projected on a screen approximately 0.5m in front of the subject’s face. The trial structure of the task is illustrated in figure 1A. A fixation cross was presented at all times except during picture presentation. Each sentence was followed by a silent delay period of three seconds, during which the relevant information was held in working memory. Next, two pictures appeared on the screen on either side of the fixation cross, which disappeared. Subjects indicated their choice of picture by pressing the left or right button of a fiberoptic response box with the index or middle finger, respectively, of the right hand. Subjects were instructed to make their selection within the time that the pictures remained on the screen (four seconds). Upon picture selection, a green box appeared around the selected picture to confirm that the selection was registered, but no feedback about accuracy was given. After each picture trial, a delay of 2.1–2.25s occurred before the presentation of the next sentence. Subjects were instructed to refrain from head and facial movement, and were informed that minimal blinking during trials was preferred, although they were not asked to completely refrain from blinking. Trials were presented in seven runs of 36 trials each, and subjects were allowed to rest in between runs if desired. Total time including electrode preparation was about 2 hours.
EEG recording
EEG was recorded from 34 Ag-AgCl electrodes, fixed in an elastic cap in an extended 10–20 arrangement. Additional electrodes were placed on the left and right mastoid. Electrode Cz was placed halfway between the nasion and inion for each subject, and one of three caps was used depending on the individual’s head size. The skin under electrode sites was lightly abraded using NuPrep, and conductive gel was inserted between the electrode and the scalp. Scalp impedances were reduced below 5kΩ. Head localization coils were placed at three fiducial points, the nasion and left and right preauricular points, for MEG head localization and coregistration with structural MRI. Electrode locations and fiducial points were measured with a 3D digitizer (Polhemus, Colchester, VT). EEG was amplified and digitized along with the MEG data at 600 Hz, with an online 125 Hz lowpass filter. EEG recordings were referenced on-line to the left mastoid, and re-referenced offline to the average of the left and right mastoids. EEG data were downsampled offline to 200 Hz (100 Hz lowpass filter), and submitted to independent component analysis to remove effects of blinks and horizontal eye movements (Jung et al., 2000), using the EEGLAB toolbox (Delorme and Makeig, 2004). In each subject, either 2 or 3 components were removed, resulting in satisfactory suppression of ocular artifacts.
EEG statistical analysis
EEG was analyzed in the time-domain, in order to examine traditional Event-related Potential (ERP) components affected by this task. Digital triggers embedded in the stimulus sound files were used to timelock ERPs to the onset of each the three portions of the sentence (see above), as well as the post-sentence memory delay. Because we were timelocking our ERP analysis to word boundaries within a continuous stream of auditory speech input, it was difficult to predict a priori the expected latency of ERP components that may be modulated by the factors of the experiment, precluding statistical tests on pre-defined temporal windows. Most importantly, in our design it is conceivable that significant effects could occur systematically at the time of one of our defined phrase boundaries, but begin before the actual point used for timelocking, which is not usually the case in ERP experiments.
To provide the flexibility necessary to reveal effects of interest while protecting against type-1 error given multiple comparisons across time points and electrodes, we employed a two-stage approach to statistical analysis. First, we conducted a cluster analysis to identify time windows in which the ERPs differed significantly between two key conditions (Maris, 2004, Maris and Oostenveld, 2007). In the second stage, we conducted a standard factorial repeated-measures ANOVA within the time windows identified by the cluster analysis. This second stage allows for a full characterization of differences between all the conditions included in the experiment, but it is not independent of the cluster analysis that identified the periods of significant effects. Therefore, the cluster analysis should be considered the primary one, and the ANOVA serves as an exploratory follow-up to further elucidate the effects and facilitate comparison with previous studies.
Cluster analysis routines were implemented in the Fieldtrip toolbox (http://fieldtrip.fcdonders.nl/), using the approach outlined in Maris and Oostenveld (2007). Briefly, a paired t-test comparing two conditions was conducted at each electrode at each time point, and t-values exceeding a threshold of p<.05 were clustered based on adjacent time bins and neighboring electrodes (distance <6cm). Cluster-based corrected p-values were produced by randomly permuting the assignment of individual subjects values to the two conditions 1000 times, and counting the number permutations in which larger clusters (defined by the total t-values of all timepoints and electrodes exceeding p<.05) were obtained than those in the correct assignment of conditions. As cluster analysis depends on homogenous effects across adjacent time bins, we first smoothed the EEG data using an FIR filter with a cut-off of 5 Hz, in order to increase sensitivity to relatively slow timelocked components such as the N400 and P600. The waveforms displayed in figures 2–3 have undergone this filtering. For comparability to previous studies, the followup ANOVAs were conducted on unfiltered data (bandwidth 0–100 Hz), averaged across time bins within temporal windows identified by the cluster analysis. We also conducted the same ANOVAs on filtered data, and obtained essentially identical results. This is unsurprising, as the conventional procedure of averaging data within time windows extending over several hundreds of milliseconds is itself a strong form of low-pass filtering.
Figure 2. ERP overview, whole trials.
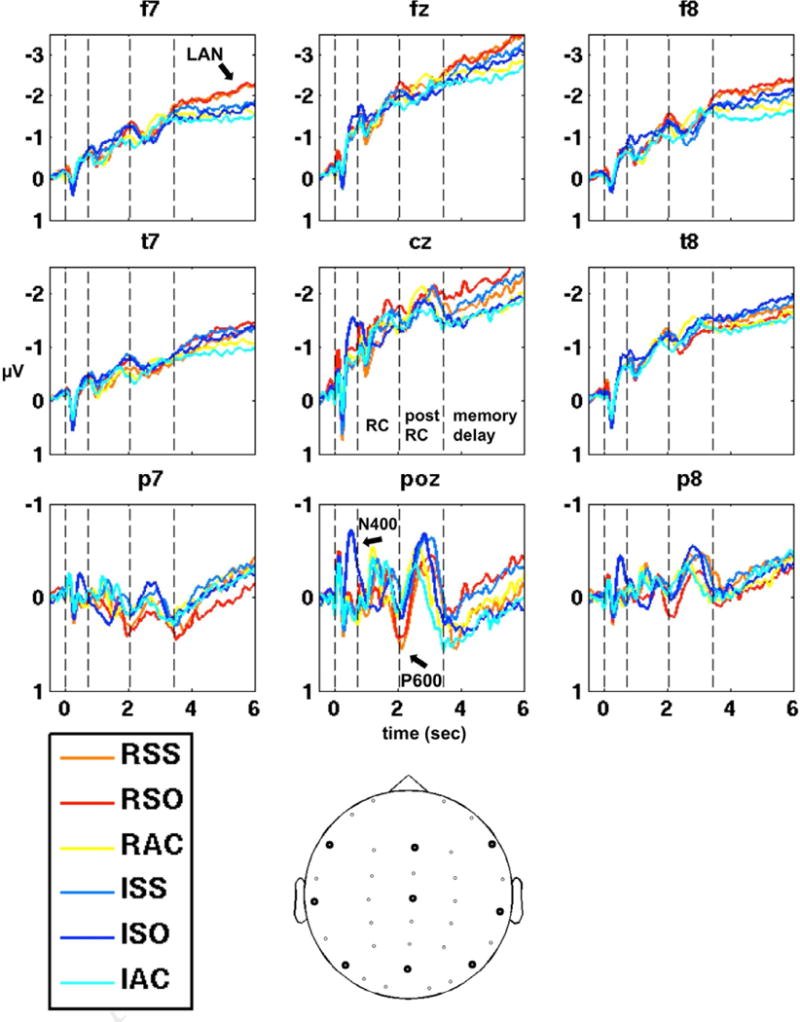
Multi-phrase ERPs timelocked to sentence onset, shown at 9 selected channels (locations plotted at the bottom) in the six sentence conditions. Vertical dotted lines indicate the average positions of the four digital triggers illustrated in figure 1B. Corresponding labels for the same triggers (except the first, for lack of room) are shown in the lower right-hand panel (electrode P8). Although the statistical analyses were done based on ERPs timelocked to individual trigger onsets within each sentence, the temporal overlap of evoked activity across sentences is sufficient that the three major effects are still visible in the multi-phrase ERPs, as indicated by arrows. LAN = Left Anterior Negativity.
Figure 3. ERP significant clusters.
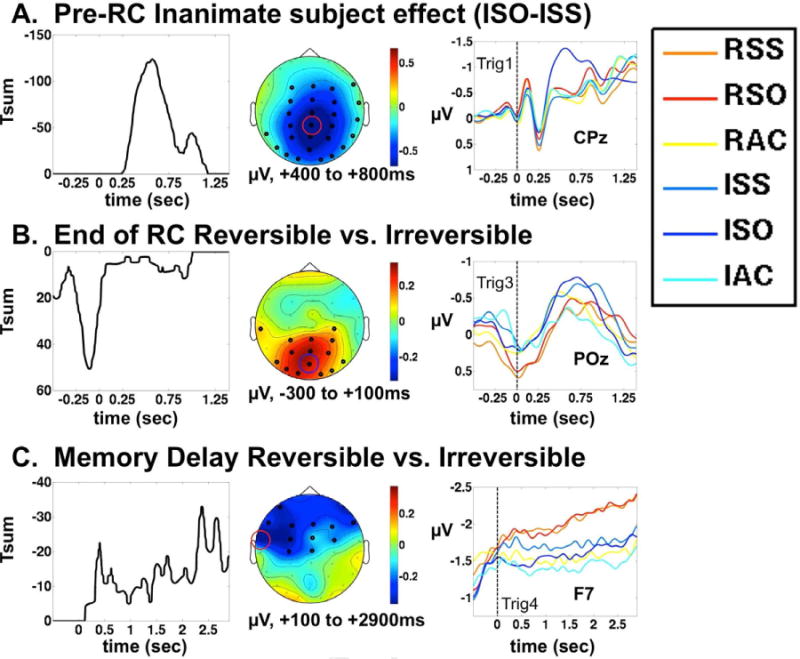
For all three rows, the left panel shows the sum of t-values for the entire cluster at each time point, giving a measure of the temporal evolution of the cluster as a whole. In keeping with the ERP convention used for the waveforms, negativity is plotted upwards. The middle panel is a topographic map of the voltage difference between the two conditions (or sets of conditions), integrated over the time range indicated. The Right panel is the ERP timelocked to a within-trial trigger, shown at a single representative channel, as indicated. The trigger point to which the response is timelocked is indicated (see figure 1 for trigger locations). A: N400 effect timelocked to very beginning of the sentence. This response is evoked by the inanimate subjects of the ISO sentences (e.g. The car vs. the boy). B: P600-like posterior positivity evoked by reversible vs. irreversible sentences, occurring approximately at the border between the second and third sentence segments. For relative-clause containing sentences (SS and SO), the timelocking point is the onset of the first word following the relative clause, e.g. “The woman who the man is teaching IS very tired right now.” C: Left anterior negativity evoked by reversible vs. irreversible sentences during the post-sentence memory delay, prior to picture onset.
Comparisons of interest
The cluster analysis described above is best suited for testing binary contrasts. On the basis of previous work with the same experimental materials, there were two particular contrasts that were expected to modulate neural activity in response to sentences. One is reversibility. For this effect, we compared activity between all reversible and irreversible sentences, collapsing across the different syntactic structures, i.e.
Sentences were contrasted based on timelocking to different triggers within the sentence, as described in the results. We used the onset of the second and third sentence segments, as well as the post-sentence memory delay.
The second contrast of interest was (RSO-RSS). We refer to this as “syntactic complexity,” but strictly speaking, it is syntactic complexity only within the context of reversibility, as it does not include the irreversible trials, nor does it include the simple active sentences. Our previous fMRI and MEG studies revealed large effects of this contrast on the BOLD signal and oscillatory power, whereas the comparable contrast within irreversible sentences (ISO-ISS) did not. Therefore, we elected to test for an effect within reversible sentences using the cluster analysis, and then followup with a full factorial ANOVA on significant time windows to test for any potential effect within the irreversible conditions.
We also conducted one additional cluster analysis, only on the first segment of the sentence, which introduced the subject of the main clause. This was between the irreversible conditions ISO-ISS. This was done because of a lexical difference in this segment between the ISO condition and all other conditions, as described in the results.
Results
Behavioral
Behavioral results from this study have been reported in detail previously (Meltzer and Braun, 2011), including statistical analysis. Given that the present report presents an analysis of ERP data collected at the same time, we briefly summarize the behavior here. Reaction time and error rate for all 6 conditions are given in table 2. There was a strong effect of reversibility on both reaction time (RT) and error rates, with reversible sentences leading to longer RT and more errors. For both behavioral measures, there were also effects of syntactic complexity, and an interaction of reversibility and complexity, such that the RSO sentences caused particular difficulty, with the longest RT and the most errors. Note that these comparisons collapse across foil type. The increased difficulty in RSO sentences is mainly driven by the 50% of trials with syntactic foils, in which the roles of the two people are reversed (see Meltzer and Braun, 2011, for details). In the present report, all ERP effects are taken from time periods prior to the picture-matching event, such that the different foil types are not relevant, but they do affect the behavioral results, which are obtained from the response to the pictures.
Table 2.
Reaction times and error rates for picture selection following the six sentence conditions. Errors include incorrect responses and non-responses.
| condition | rt mean (ms) | error mean |
|---|---|---|
| RSS | 1602 | 1.98% |
| RSO | 1752 | 4.07% |
| RAC | 1526 | 2.18% |
| ISS | 1247 | 0.99% |
| ISO | 1269 | 0.59% |
| IAC | 1246 | 3.17% |
Whole-sentence ERPs
This experiment used auditory sentences spoken at a natural rate. The primary topic of interest in this study is slow potentials that evolve over several words during a relative clause, after a relative clause, and during the memory delay following the sentence. To isolate these temporal segments, we conducted statistical analyses on ERPs timelocked to digital triggers embedded in the sound files at the onset of each individual segment. These analyses revealed three significant effects related to the differences between sentence conditions, which will be detailed below. However, it is nonetheless possible to observe these effects in the multi-phrase ERPs obtained by averaging over the entire time of the trial, timelocked only to the onset of the first word of the sentences, despite the fact that the phrase boundaries do not line up precisely across sentences. For illustrative purposes, we provide an initial overview of the ERP effects by presenting whole-sentence ERPs in all six conditions, from a selection of nine electrodes. The dotted vertical lines within the graphs represent the average time of the triggers dividing the sentence segments. In the first segment, the ISO condition differs from the other 5 conditions, and induces an N400 effect, indicated at electrode POz in figure 3 (although this was not the location of the maximum for the N400, see below). At the end of the second segment (the relative clause), and extending through the third segment, an enhanced posterior positivity is seen in the RSS and RSO conditions, also indicated at electrode POz. This positivity displays characteristics of a P600 response (see discussion), and is so labeled in the figure. Finally, during the memory delay period after the sentences, the same two conditions, RSS and RSO, induce a negativity at anterior electrodes, more strongly on the left, indicated at electrode F7. These three effects (N400, P600, and Left Anterior Negativity) are shown in more detail in the following sections and figures, along with the statistical results establishing their significance.
First sentence segment – N400
The first segment comprised a simple noun phrase, the subject of the sentence. For three out of the four conditions, this was one of the actors “The man,” “the woman,” “the boy,” or “the girl.” Only for the ISO condition did this segment differ, in which case it was an inanimate undergoer, e.g. “the ball,” “the car,” etc. Therefore we compared the ISO condition with the ISS condition. One significant cluster was detected for this comparison, in the form of central-parietal negativity peaking at approximately 500 ms after the onset of the sentence. This corresponds to the classic N400 potential (Kutas and Hillyard, 1980). The left panel of figure 3A displays the sum of t-values in the cluster at each time point (Maris et al., 2007), illustrating the temporal evolution typical of an N400 effect. The middle panel shows the topography of ISOISS from 400 to 800 ms after the sentence onset. Electrodes that are significant at any point within this time window are shown as dark black circles, while other electrodes are shown in light gray. The right panel of figure 3A shows the ERP signals from all six sentence conditions at electrode CPz, showing that the N400 effect is specific to the ISO condition.
Second sentence segment – relative clause
This segment was timelocked to the onset of the relative clause (SS and SO conditions) or the main verb (AC condition). The comparison testing for effects of syntactic complexity, RSO-RSS, did not reveal any significant effects. We tested for a general effect of reversibility using the contrast (RSO+RSS+RAC)–(ISO+ISS+IAC). This revealed a significant cluster of positivity at the beginning of the segment, but visual inspection suggested that this effect was driven by the large N400 effect engendered by sentence-initial words in the ISO condition, extending into the time period of the second segment. Therefore, general effects of reversibility within this sentence segment are better revealed by timelocking to the boundary between the second and third sentence segment. As discussed below, a positivity timelocked to this boundary does extend backwards into the latter portion of the second sentence segment.
Third sentence segment – P600-like positivity
This segment is timelocked to the onset of the third sentence segment, which falls after the relative clause in the SS and SO conditions, and after the main clause in the AC condition. Again, the syntactic complexity comparison RSO-RSS did not reveal any significant effects. However, the reversibility contrast, (RSO+RSS+RAC)–(ISO+ISS+IAC), revealed one significant cluster, a posterior central positivity. This cluster is maximal just before the timelocking trigger, indicating that it is most likely evoked by linguistic information within the previous sentence segment. Therefore, this positivity may be thought of as a response to the end of the relative clause (or main clause for the AC conditions), rather than the beginning of the next segment. In all conditions, this was the sentence segment that presented the critical information relevant to the picture matching task (i.e. who is doing what to whom). Reversible sentences evoked a more positive signal than irreversible ones within this time period.
The temporal evolution of the cluster T-sum value is shown in figure 3B, left panel. Although the cluster persists at some electrodes well into the third sentence segment, the bulk of it peaks and subsides as the second segment ends, between approximately −300 to +100 ms relative to the trigger. The middle panel displays the average ERP topography in that window, with electrodes achieving significance highlighted in black. It is difficult to discuss a “latency” for this cluster, as it is detected on both sides of a point of interest within a continuous auditory sentence, but the long duration and topographic distribution of this effect suggest homology to a P600 component (see discussion). Waveforms at electrode POz are shown in the right panel, illustrating that this positivity appears virtually identical in the RSO and RSS conditions. The RAC condition expresses the effect less strongly, but is still more positive than the irreversible conditions.
Delay period – post-sentence
This time window comprised the offset of the sentence, through to the end of the delay period, just prior to picture onset. As with the previous time window, we saw no significant effect of syntactic complexity (RSO-RSS), but there was again a significant cluster for the effect of reversibility. During the memory delay period, greater negativity was seen at anterior electrodes (particularly on the left) following reversible sentences, especially in the RSS and RSO conditions. This kind of effect is commonly described as “left anterior negativity” (LAN). Figure 3C (left panel) shows the temporal evolution of the cluster, showing that it continues to grow stronger up to the end of the delay period. The middle panel shows the topography from +100 to +2900 msec, demonstrating the left anterior distribution of the effect. The right panel shows the waveforms at electrode F7 timelocked to sentence offset. We see here that reversible sentences containing relative clauses (RSS and RSO) induce a sustained negativity in the delay period, but this effect is relatively insensitive to the different syntactic structures of subject-embedded and object-embedded clauses. We also observe that the relative clause conditions evoke greater negativity than the simple active sentences, within both categories of reversibility.
Followup ANOVA – P600
Cluster analysis revealed a robust effect of semantic reversibility at the boundary between the second and third sentence segments, in the form of an enhanced central-posterior positivity for reversible sentences. However, no differences between the RSO and RSS conditions were observed, in contrast to some previous ERP studies of relative clause processing (King and Kutas, 1995, Muller et al., 1997). To test for a possible effect of syntactic structure across all three sentence types (including the simple active sentences), we conducted a full factorial ANOVA on the ERP signal in the central-posterior region. Mean signal amplitudes in the time window of −300 to +100 ms relative to trigger 3 were averaged across 7 central-posterior electrodes (shown in figure 4A) and submitted to a 2×3 repeated-measures ANOVA, with reversibility and syntactic structure (SS, SO, and AC) as within-subject factors. The Greenhouse-Geisser adjustment to degrees of freedom was applied to all significance calculations involving the structure factor (not applicable to factors with only two levels). Mean amplitudes are displayed in figure 4B. As expected, given that the scalp region and time window were selected based on the cluster analysis, there was a large effect of reversibility, with reversible sentences evoking more positive amplitudes [F(1,23)=9.782, p<.005]. There was no significant effect of syntactic structure [F(1.89,46)=.782, p=.457] nor any interaction between reversibility and structure [F(1.99,45.71)=1.438, p=.248]. Therefore, we conclude that although this P600-like positivity is sensitive to reversibility, there is no evidence for its sensitivity to syntactic structure.
Figure 4. ROI averages.
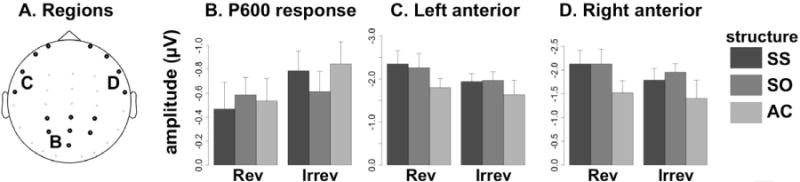
A: Locations of electrodes averaged together for region-of-interest based (ROI) followup ANOVAs. Letters on the scalp map indicate the figure panel in which responses from that region are displayed. B: ERP amplitudes for all 6 sentence conditions in the central-posterior ROI during the time window of the P600 response, −300 to +100ms relative to trigger 3, the border between the second and third sentence segment. In keeping with the ERP convention used for the waveforms, negativity is plotted upwards. C: ERP amplitudes for the left anterior ROI during the time-window of +100 to +2900ms after trigger 4, comprising most of the post-sentence memory delay. D: ERP amplitudes for the right anterior ROI during the same time-window as panel C. Error bars are 95% confidence intervals for repeated measures designs using the method of Morey (2008).
Followup ANOVA – Anterior Negativity
Cluster analysis revealed a second effect also sensitive to semantic reversibility, an enhanced negativity at lateral electrodes during the post-sentence memory delay. Visual inspection of the response (figure 3C) suggests that the RSS and RSO conditions evoke similar enhancements of negativity relative to the other conditions, but do not differ between themselves. To assess this impression statistically and to test for significant differences in laterality, we conducted a 2×3×2 repeated-measures ANOVA, with reversibility, syntactic structure, and laterality (left vs. right) as within-subject factors. Mean signal amplitudes in the time window of +100 to +2900 ms relative to trigger 4 were averaged across 4 lateral anterior electrodes on the left and 4 on the right (shown in figure 4A). Mean amplitudes in the left and right regions are displayed in figure 4C and D. ANOVA revealed significant main effects of reversibility [F(1,23)=4.418, p<.05] and syntactic structure [F(1.89,43.25)=9.075, p=.001], but no interaction between the two [F(1.77,40.79)=.622, p=.523]. There were no significant main effects or interactions involving the laterality effect. Thus, we observe that reversibility and syntactic structure exert parallel, additive effects on anterior negativity during the memory delay. The effect of syntactic structure takes the form of an enhanced negativity for the SS and SO conditions (both containing relative clauses) compared to the simple active sentences, rather than a difference between the two types of relative clause. This enhanced negativity is in fact bilateral, although it appears somewhat stronger on the left.
Followup ANOVA – Third Sentence Segment
The lack of any significant difference between RSO and RSS sentences was somewhat surprising, given the results of Muller et al. (1997), who observed enhanced negativity for RSO sentences at lateral anterior electrodes from the end of the relative clause on to the end of the sentence. To confirm that our previous analyses had not missed such an effect, we conducted one further 2×3×2 ANOVA on mean amplitudes in the time window of +300 to +1000 ms relative to trigger 3 (the “post-RC” segment), on the same two lateral anterior regions used for the analysis of the post-sentence memory delay. This analysis revealed no significant effect of any factor (all p-values above 0.25).
Discussion
Using a sentence-picture-matching task, we measured ERPs during comprehension of nonanomalous sentences, presented as naturalistic speech at an ordinary rate. This experiment was designed to study neural activity emerging over a time scale of several words using sentences that varied in the demands that they place on syntactic comprehension mechanisms. As such, the experimental design was somewhat untraditional. In most ERP studies of sentence processing, a phenomenon of interest emerges with a single word embedded within a sentence, and the EEG signal is timelocked to the critical word. This approach has proven very successful at characterizing neural activity with high temporal resolution for such phenomena as syntactic anomalies (Osterhout and Holcomb, 1992), semantic anomalies (Kutas and Hillyard, 1980), syntactic ambiguity (Frisch et al, 2002), garden-path reanalysis (Osterhout et al., 1994), and others. Syntactic complexity and semantic reversibility are known to be important factors for characterizing cognitive demand in sentence comprehension, but are less easily associated with a critical word, emerging instead on the level of phrases and clauses. The materials in our current experiment were designed primarily for an analysis focused on oscillatory activity, for which precise timelocking of the signal is less critical. Nonetheless, it is possible that analysis of ERPs in the time domain would also yield components that are recognizable from previous language studies, and therefore shed light on the nature of those components.
Most of the sentences in this experiment contained a relative clause. A relative clause is a fairly demanding sentence structure to comprehend and produce, making it an attractive target for psycholinguistic experimentation. The first noun in such sentences carries out two distinct grammatical functions. In all cases, this first noun serves as the subject for the main clause, but it may be either the subject or the object of the relative clause, depending on the syntactic structure. The semantic reversibility of the sentence is not determined until both nouns have been presented, as the post-relative clause portion of the sentence begins. Therefore, we chose to use this point for timelocking of the ERP analysis. For the simple active sentences, this point also corresponds to the point at which the critical information for the sentence has been presented, although in this case it is the end of the main clause rather than the relative clause. For reversible compared to irreversible sentences, we observed an enhanced positivity beginning shortly before this point – as the critical clause is wrapping up. The characteristics of this positivity suggest a homology to the P600 potential that has been reported in numerous other studies of syntactically challenging material. In the remaining discussion, we will review the evidence for this homology, and then discuss the implications of finding the P600 response in a study of nonanomalous sentences. Finally, we will briefly discuss the other ERP components detected in this study and how they affect our interpretation.
P600-like positivity
Typically, an ERP component is defined by up to five criteria: polarity, topography, latency, duration, and the nature of the experimental manipulation that elicits it (although this last criterion is often the topic of novel findings for a known component rather than a means of defining it a priori). In terms of polarity and topography, our observed positivity is congruent with previous reports of the P600, with a bilateral symmetric central-posterior distribution, with a maximum amplitude approximately at electrode Pz. At some electrodes, the duration of the component is quite long, particularly for the RSO and RSS conditions, which evoke a greater positivity for approximately 800–900 ms (figure 3B). This is also consistent with the P600. The P600 is one of the longest-lasting of common ERP components, frequently extending past the end of recorded trials (e.g. Kaan and Swaab, 2003, Osterhout and Mobley, 1995) but not extending all the way to a post-sentence behavioral response (Pakulak and Neville, 2009).
The one criterion in which our observed positivity appears to differ from previous reports of the P600 is latency. Typically, the P600 emerges quite late, approximately 500–600 ms after the onset of a critical word. In our case, it began shortly (~100 ms) before the critical timelocking point at the end of the relative clause. However, this discrepancy is easily attributable to the particular experimental design that we used. Relative clauses that are semantically reversible incur an increased processing cost both as they are being perceived and shortly after (Traxler et al., 2002, 2005), making it likely that the neural correlates of this processing would be detectable during the relative clause presentation. In our sentences, it is difficult to pinpoint exactly when in the sentence that reversibility would begin to exert an effect. However, it is a straightforward prediction that neural activity related to processing a reversible relative clause should extend for several hundred milliseconds after the clause has ended, and this is exactly what we see.
Thus, our observed positivity resembles the P600 commonly reported in studies of syntactic anomaly, although our experimental design makes it difficult to define its latency precisely. We will now consider the implications of observing a P600-like positivity in a context in which it has not previously been reported, namely, following semantically reversible clauses. Observations of P600 activity at points of syntactic anomaly or disambiguation (Osterhout et al., 1994) have led to proposals that this component reflects an extended analysis, or reanalysis, of syntactic information in working memory. Further support for this view comes from a more recent wave of evidence demonstrating P600 responses to a different class of anomalies that may have been considered semantic in nature rather than syntactic, according to traditional distinctions. The concept of animacy has proven to be particularly germane to the nature of these P600 effects. Animacy is explicitly encoded in the grammar of different languages to various degrees (Comrie, 1989) and is a prototypical example of a phenomenon that lies on the interface of semantics and syntax. Several recent studies have demonstrated that some semantic anomalies related to animacy contraints produce not a “semantic” N400 response, but rather a P600. Animacy-related anomalies causing a P600 in English typically involve a mismatch between the thematic roles implied by a syntactic position, and those associated with the animacy of the nouns in the sentence. A canonical example is the following sentence.
5) “Every morning at breakfast the eggs would eat…” (Kuperberg et al., 2003)
Here, the sentence is grammatically well formed, but because eggs are inanimate objects that do not eat, the meaning is semantically anomalous if one interprets the sentence strictly according to the syntactic cues, which assign the thematic role of AGENT to “eggs.” Nonetheless, the verb “eat” elicits a P600 potential, rather than an N400. Follow-up studies have demonstrated that the semantic attraction between “eggs” and “eat” (i.e. the fact that “eggs” are a plausible PATIENT but not AGENT) is not a necessary precondition for the P600 to occur, but that it can be commonly seen in sentences with an inanimate noun anomalously assigned the AGENT role by syntactic cues (Hoeks et al., 2004, Kuperberg et al., 2007, c.f. Kim and Osterhout, 2005). Various interpretations of these findings have been proposed (for review, see Kuperberg, 2007, Bornkessel-Schlesewsky and Schlesewsky, 2008), but authors generally agree that the P600 activity elicited by such violations reflects reanalysis of a sentence in order to determine the proper thematic roles of nouns. Further support for a key role of the P600 in thematic role assignment comes from a previous ERP study of sentence-picture-matching by Wassenaar et al. (2007), which demonstrated a P600 in response to mismatches of thematic roles between pictures and sentences. That response was also followed by an anterior negativity, as we have seen in the present study.
Our observation of a P600-like positivity in response to reversible clauses supports the idea that this component plays a role in comprehension of thematic roles on the basis of syntactic cues. In our irreversible sentences, comprehension of thematic roles is very easy, as both syntactic cues and animacy constraints converge on the same conclusion: the animate noun is the AGENT, and the inanimate noun is the PATIENT. In our reversible sentences, however, both nouns are animate and equally plausible in the AGENT role. Therefore, subjects must rely entirely on syntactic cues to determine which noun is the AGENT. This increased demand for processing of syntactic cues was reflected by a positive deflection in the ERP signal, resembling the P600 reported in previous studies of anomalous materials. Therefore, we suggest that the neural generators of the P600 signal in response to violations of syntactic structure and animacy constraints may also be involved in processing normal, nonanomalous sentences that are characterized by increased demands on syntactic processing.
We note that our experiment made no attempt to discriminate between effects of reversibility and animacy. By definition, all of our reversible sentences involved two animate nouns, whereas the irreversible sentences involved an animate AGENT and an inanimate PATIENT. Some studies have examined irreversible sentences in which an inanimate noun acts on an animate noun, such as “The critic that the film impressed…” or “The hiker that the boulder injured…” (Weckerly and Kutas, 1999; Chen et al., 2006). These sentences do not typically lend themselves well to visualization in a sentence-picture-matching task, so we chose to limit our materials to the more typical animate-AGENT sentences. As our experimental goal was simply to examine activity related to sentences in which semantic constraints do not determine thematic roles, there was no need to dissociate reversibility from animacy type.
Interestingly, we did not observe any modulation of the P600 response related to syntactic complexity (object-embedded vs. subject-embedded relative clauses). However, this is consistent with our previous findings using the same linguistic materials. We found that semantic reversibility induced greater activity in both anterior and posterior language regions, but that syntactic complexity exerted an effect only in anterior regions. The P600 potential has a bilateral posterior distribution, and MEG-based dipole localization has identified posterior perisylvian cortex as its primary site of origin (Service et al, 2007). Therefore, no effect of structural syntactic complexity on the P600 would be expected, since we did not observe such an effect in posterior cortex in either fMRI (Meltzer et al., 2010) or in oscillatory activity measured with MEG (Meltzer and Braun, 2011).
N400
We observed a very typical-looking N400 response following the initial noun of the ISO sentences. Although this effect was not central to the experimental hypotheses, it is useful for comparison, as the latency, duration, and topography clearly differentiate it from the Left Anterior Negativity observed after the sentence (see below). This N400 effect can be explained by two different factors, and both of them likely play a role in its generation. First, sentences in all other conditions begin with a restricted set of words: the man, the boy, the woman, or the girl. In contrast, the ISO sentences begin with the PATIENT inanimate object – the car, the shovel, etc. As the N400 is quite sensitive to semantic priming, including repetition priming (Doyle et al., 1996), it would be highly suppressed in the other conditions, in which the same opening words are repeated many times throughout the experiment. In contrast, the unique words that begin the ISO sentences are less expected, leading to an enhanced N400. A second factor is animacy – these sentences have an inanimate subject, which is somewhat less common in English. Previous studies have shown N400 effects in response to violations of typical constraints of animate-inanimate word order (Weckerly and Kutas, 1999, Philipp et al., 2007).
The N400 response observed for the ISO sentences is the largest in amplitude of the components observed in this experiment, which is mainly attributable to the fact that it is evoked by the first word of the sentence (not counting the initial “the.”) Therefore, the timelocking of this response is better than the other responses that occur in the midst of a continuous stream of natural speech. Nonetheless, the N400 response is over after one second, and the ERP in the ISO condition has returned completely to the level of the other conditions. Therefore, it is unlikely that the unique beginnings of the sentences in the ISO condition (nouns other than “man,” “woman,” etc.) contributed to any of the other components evoked later in the sentences.
Left Anterior Negativity
The final component observed in this study was a prolonged negativity, maximal over the left anterior electrodes, which followed sentences containing semantically reversible relative clauses. This component emerged only after the sentence was completed, and continued throughout the 3-second delay period until the pictures appeared on the screen. The magnitude of the component was sensitive to both reversibility and syntactic structure in parallel, in that it was increased for SS and SO sentences relative to simple actives, but no difference was observed for object-relative vs. subject-relative clauses. Although “left anterior negativity” (LAN) is an accurate description of the topography and polarity of this component, we caution that it may not be homologous to the LAN responses typically reported in studies of syntactic anomalies. Anomaly-related LAN’s typically occur in similar contexts as P600s, but precede the P600 in time and completely return to baseline in a few hundred milliseconds (Kutas and Hillyard, 1983, Gunter et al., 1997, Penke et al., 1997, Coulson et al., 1998). However, anterior negativity extended over several seconds has also been observed in language comprehension paradigms, and seems to reflect increased demands on working memory (Munte et al., 1998, King and Kutas, 1995, Muller et al., 1997, Martin-Loeches et al., 2005). Steinhauer and Drury (2012) present evidence that even the LANs that precede P600 effects may represent a temporally extended process, but the scalp negativity may be cancelled by the subsequent emergence of the P600. Furthermore, it is common to observe a posterior central positivity followed by an extended anterior negativity, in tasks in which reporting of linguistic violations imposes a substantial working memory requirement (Sabourin and Stowe, 2004, Steinhauer et al., 2010). A similar pattern was also observed in encoding and retention phases of a working memory task for consonants (Ruchkin et al., 1990). Thus, our results fit with a general model in which early reanalysis of linguistic information is associated with a posterior positive wave, while the retention of the information over several seconds gives rise to an anterior negativity. In our study, the information in question has to do with the thematic roles of the people mentioned in a sentence.
It is somewhat surprising that our observed anterior negativity effect did not differ between the RSO and RSS conditions. Earlier studies of King and Kutas (1995) and Muller et al. (1997), using similar sentence materials in the visual and auditory domains, respectively, observed increased frontal negativity evoked by object-embedded relative clauses, occurring within the sentence itself (as opposed to a post-sentence memory delay as used in our study). We cannot be certain about the reasons for the discrepancy between our findings and the previous two ERP studies that have examined object-relative clauses, but we speculate that it is due to the relative difficulty of the sentences. Muller et al. (1997) attribute the anterior negativity to increased engagement of working memory involved in reanalysis of the sentence. This is evidenced by the fact that subjects with better comprehension performance exhibited more negativity. Overall performance in that study was 77%, and presumably even lower in the object-embedded condition. The sentences were somewhat longer and more abstract than ours, e.g. “The reporter who the senator harshly attacked…” In our study, the comprehension probe was a picture-matching trial, rather than a true/false question. This led us to choose sentences dealing with highly imageable concrete actions, which may have reduced subjects’ need to rely on phonological short-term memory, given the increased availability of a non-verbal visual representation of the sentence’s meaning. Reduced demand on working memory may account for the much higher accuracy obtained by our participants. The increased memory demands of the RSO condition may have been insufficient to elicit a detectable change in ERP amplitude. However, this does not mean that there was no effect of object-embedded clauses for our sentences. In previous studies with the same materials, we observed increased activation in frontal cortex for RSO vs. RSS sentences using both fMRI (Meltzer et al., 2010) and oscillatory analysis of EEG and MEG (Meltzer and Braun, 2011). These findings reinforce the idea that ERPs offer a selective view of certain forms of neural activity that happen to result in a good time-domain average signal, but may be insensitive to certain effects that are discernable with other methods. Of course, such selectivity is inherent to all noninvasive neuroimaging techniques, underscoring the need for multimodal comparisons.
It is also interesting to compare our results with behavioral studies of relative clause processing. Using eye-movement monitoring in self-paced reading Traxler et al. (2002, 2005) demonstrated longer fixations and increased regressions within relative clauses, particularly object-embedded ones. In contrast, we observed increased neural activity in EEG and MEG only after the clauses have been presented. This difference is most likely attributable to task design. In self-paced reading, subjects are free to engage in increased reanalysis of syntactically challenging material as soon as it is encountered. In an externally paced neuroimaging experiment, subjects have no control over the presentation rate. Therefore, neural correlates of increased working memory engagement related to reanalysis are more likely to emerge after the difficult material has been presented.
Conclusion
To summarize, we have observed that semantically reversible clauses in a sentence-picture-matching task evoke two separate ERP responses compared to their irreversible counterparts: 1) a P600-like positivity occurring at the end of the clause, and 2) a left-dominant anterior negativity beginning after the sentence is over and spanning the length of the delay period. Both of these components are interpretable as reflecting increased demands for retention and reanalysis of syntactic information for successful completion of the picture-matching trial, since for the reversible sentences it is not enough to simply remember the lexical items mentioned in the sentence; one must also retain “who did what to whom.”
In an influential review on language and working memory, Caplan and Waters (1999) distinguish between phases of “interpretive” and “post-interpretive” processes. Interpretive processes involve extracting the meaning of a sentence from as it is perceived, whereas post-interpretive processes involve retaining and manipulating that information long enough to complete a secondary task depending on the sentence content (such as picture-matching, enactment, or answering a comprehension probe). However, even interpretation is not always instantaneous, as proper assignment of thematic roles to nouns often depends on further information later in a sentence. Therefore, a process of immediate reanalysis is a necessary part of comprehension of certain syntactic structures, such as relative clauses. Our results suggest that this process occurs more strongly for reversible sentences, in which syntactic information is more important for accurate interpretation, and is reflected by a P600 response. In contrast, the LAN observed here seems to map to the later, post-interpretive processes.
These observations are consistent with contemporary models of the neuroanatomy of working memory. Initial perception and interpretation of sensory stimuli is generally held to occur in primary sensory and association areas in the occipital, parietal, and temporal lobes, while extended retention and manipulation of information involves an interactive exchange of information between these same areas and highly connected portions of the frontal lobe (Romanski, 2004. Postle, 2006, Linden, 2007). The pattern of results emerging from our studies on comprehension of reversible sentences suggests that syntactic comprehension draws on working memory resources in a similar fashion. Using oscillatory analysis of MEG, we observed increased neural activity in posterior perisylvian cortex in response to semantic reversibility as sentences were being presented, and spreading to more frontal regions while the sentences were being retained in short-term memory (Meltzer and Braun, 2011). The present results using ERP analysis suggest that these stages of activity are associated with different scalp components, the early posterior P600 and later anterior LAN. These components have been widely studied in a large variety of anomaly paradigms, but it is commonly assumed that they reflect processes that are constantly active (to a lesser degree) in ordinary language comprehension. The observation of these components in response to nonanomalous sentences provides direct evidence to support that assumption. Semantic reversibility is a property of many utterances that we hear on a daily basis. We have demonstrated here that understanding those utterances draws on some of the same neural systems that are activated in ERP studies of anomalous sentences.
P600 is an event-related potential evoked by syntactic errors in sentences.
P600 may be a general index of neural activity related to syntactic processing.
We detected the P600 and other syntactic ERPs to well-formed sentences.
The ERP reflected syntactic processing necessary to understand the sentence.
Acknowledgments
We thank Joseph McArdle, Susannah Hoffman, Elizabeth Rawson, and Judy Mitchell-Francis for assistance with EEG recording. This work was supported by the intramural program of the National Institute on Deafness and Other Communication Disorders.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bornkessel-Schlesewsky I, Schlesewsky M. An alternative perspective on “semantic P600” effects in language comprehension. Brain Res Rev. 2008;59:55–73. doi: 10.1016/j.brainresrev.2008.05.003. [DOI] [PubMed] [Google Scholar]
- Caplan D, Waters GS. Verbal working memory and sentence comprehension. Behav Brain Sci. 1999;22:77–94. doi: 10.1017/s0140525x99001788. discussion 95–126. [DOI] [PubMed] [Google Scholar]
- Chee MW, Hon NH, Caplan D, Lee HL, Goh J. Frequency of concrete words modulates prefrontal activation during semantic judgments. Neuroimage. 2002;16:259–268. doi: 10.1006/nimg.2002.1061. [DOI] [PubMed] [Google Scholar]
- Chen E, West WC, Waters G, Caplan D. Determinants of bold signal correlates of processing object-extracted relative clauses. Cortex. 2006;42:591–604. doi: 10.1016/s0010-9452(08)70397-6. [DOI] [PubMed] [Google Scholar]
- Comrie B. Language Universals and Linguistic Typology. University of Chicago Press; 1989. (Chapter). [Google Scholar]
- Coulson S, King JW, Kutas M. Expect the unexpected: event-related brain response to morphosyntactic violations. Lang Cogn Proc. 1998;13:21–58. [Google Scholar]
- Delorme A, Makeig S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J Neurosci Methods. 2004;134:9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Doyle MC, Rugg MD, Wells T. A comparison of the electrophysiological effects of formal and repetition priming. Psychophysiology. 1996;33:132–147. doi: 10.1111/j.1469-8986.1996.tb02117.x. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Hahne A, Mecklinger A. Temporal structure of syntactic parsing: early and late event-related brain potential effects. J Exp Psychol Learn Mem Cogn. 1996;22:1219–1248. doi: 10.1037//0278-7393.22.5.1219. [DOI] [PubMed] [Google Scholar]
- Frisch S, Schlesewsky M, Saddy D, Alpermann A. The P600 as an indicator of syntactic ambiguity. Cognition. 2002;85:B83–92. doi: 10.1016/s0010-0277(02)00126-9. [DOI] [PubMed] [Google Scholar]
- Gunter TC, Stowe LA, Mulder G. When syntax meets semantics. Psychophysiology. 1997;34:660–676. doi: 10.1111/j.1469-8986.1997.tb02142.x. [DOI] [PubMed] [Google Scholar]
- Halgren E, Dhond RP, Christensen N, Van Petten C, Marinkovic K, Lewine JD, Dale AM. N400-like magnetoencephalography responses modulated by semantic context, word frequency, and lexical class in sentences. Neuroimage. 2002;17:1101–1116. doi: 10.1006/nimg.2002.1268. [DOI] [PubMed] [Google Scholar]
- Hoeks JC, Stowe LA, Doedens G. Seeing words in context: the interaction of lexical and sentence level information during reading. Brain Res Cogn Brain Res. 2004;19:59–73. doi: 10.1016/j.cogbrainres.2003.10.022. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Automatic and attentional processing: an event-related brain potential analysis of semantic priming. Brain Lang. 1988;35:66–85. doi: 10.1016/0093-934x(88)90101-0. [DOI] [PubMed] [Google Scholar]
- Jung TP, Makeig S, Westerfield M, Townsend J, Courchesne E, Sejnowski TJ. Removal of eye activity artifacts from visual event-related potentials in normal and clinical subjects. Clin Neurophysiol. 2000;111:1745–1758. doi: 10.1016/s1388-2457(00)00386-2. [DOI] [PubMed] [Google Scholar]
- Kaan E, Swaab TY. Repair, revision, and complexity in syntactic analysis: an electrophysiological differentiation. J Cogn Neurosci. 2003;15:98–110. doi: 10.1162/089892903321107855. [DOI] [PubMed] [Google Scholar]
- Kim A, Osterhout L. The independence of combinatory semantic processing: evidence from event-related potentials. J Mem Lang. 2005;52:205–225. [Google Scholar]
- King JW, Kutas M. Who did what and when? Using word- and clause related ERPs to monitor working memory usage in reading. J Cogn Neurosci. 1995;7:378–397. doi: 10.1162/jocn.1995.7.3.376. [DOI] [PubMed] [Google Scholar]
- Kucera H, Francis W. Computational Analysis of Present-Day American English. Providence, RI: Brown University Press; 1967. (Chapter). [Google Scholar]
- Kuperberg GR. Neural mechanisms of language comprehension: challenges to syntax. Brain Res. 2007;1146:23–49. doi: 10.1016/j.brainres.2006.12.063. [DOI] [PubMed] [Google Scholar]
- Kuperberg GR, Holcomb PJ, Sitnikova T, Greve D, Dale AM, Caplan D. Distinct patterns of neural modulation during the processing of conceptual and syntactic anomalies. J Cogn Neurosci. 2003;15:272–293. doi: 10.1162/089892903321208204. [DOI] [PubMed] [Google Scholar]
- Kuperberg GR, Kreher DA, Sitnikova T, Caplan DN, Holcomb PJ. The role of animacy and thematic relationships in processing active English sentences: evidence from event-related potentials. Brain Lang. 2007;100:223–237. doi: 10.1016/j.bandl.2005.12.006. [DOI] [PubMed] [Google Scholar]
- Kutas M, Federmeier KD. Electrophysiology reveals semantic memory use in language comprehension. Trends Cogn Sci. 2000;4:463–470. doi: 10.1016/s1364-6613(00)01560-6. [DOI] [PubMed] [Google Scholar]
- Kutas M, Hillyard SA. Reading senseless sentences: brain potentials reflect semantic incongruity. Science. 1980;207:203–205. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Kutas M, Hillyard SA. Event-related brain potentials to grammatical errors and semantic anomalies. Mem Cognit. 1983;11:539–550. doi: 10.3758/bf03196991. [DOI] [PubMed] [Google Scholar]
- Linden DE. The working memory networks of the human brain. Neuroscientist. 2007;13:257–267. doi: 10.1177/1073858406298480. [DOI] [PubMed] [Google Scholar]
- Maris E. Randomization tests for ERP topographies and whole spatiotemporal data matrices. Psychophysiology. 2004;41:142–151. doi: 10.1111/j.1469-8986.2003.00139.x. [DOI] [PubMed] [Google Scholar]
- Maris E, Oostenveld R. Nonparametric statistical testing of EEG- and MEG-data. J Neurosci Methods. 2007;164:177–190. doi: 10.1016/j.jneumeth.2007.03.024. [DOI] [PubMed] [Google Scholar]
- Martin-Loeches M, Munoz F, Casado P, Melcon A, Fernandez-Frias C. Are the anterior negativities to grammatical violations indexing working memory? Psychophysiology. 2005;42:508–519. doi: 10.1111/j.1469-8986.2005.00308.x. [DOI] [PubMed] [Google Scholar]
- Meltzer JA, Braun AR. An EEG-MEG Dissociation between Online Syntactic Comprehension and Post Hoc Reanalysis. Front Hum Neurosci. 2011;5:10. doi: 10.3389/fnhum.2011.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meltzer JA, McArdle JJ, Schafer RJ, Braun AR. Neural aspects of sentence comprehension: syntactic complexity, reversibility, and reanalysis. Cereb Cortex. 2010;20:1853–1864. doi: 10.1093/cercor/bhp249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morey RD. Confidence intervals from normalized data: A correction to Cousineau (2005) Tutorials in Quantitative Methods for Psychology. 2008;4:61–64. [Google Scholar]
- Muller HM, King JW, Kutas M. Event-related potentials elicited by spoken relative clauses. Brain Res Cogn Brain Res. 1997;5:193–203. doi: 10.1016/s0926-6410(96)00070-5. [DOI] [PubMed] [Google Scholar]
- Munte TF, Schiltz K, Kutas M. When temporal terms belie conceptual order. Nature. 1998;395:71–73. doi: 10.1038/25731. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Holcomb PJ. Event-related brain potentials ellcited by syntactic anomaly. J Mem Lang. 1992;31:785–806. [Google Scholar]
- Osterhout L, Holcomb PJ, Swinney DA. Brain potentials elicited by garden-path sentences: evidence of the application of verb information during parsing. J Exp Psychol Learn Mem Cogn. 1994;20:786–803. doi: 10.1037//0278-7393.20.4.786. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Mobley LA. Event-related potentials elicited by failure to agree. Journal of Memory and Language. 1995;34:739–773. [Google Scholar]
- Pakulak E, Neville HJ. Proficiency Differences in Syntactic Processing of Monolingual Native Speakers Indexed by Event-related Potentials. J Cogn Neurosci. 2009 doi: 10.1162/jocn.2009.21393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penke M, Weyerts H, Gross M, Zander E, Munte TF, Clahsen H. How the brain processes complex words: an event-related potential study of German verb inflections. Brain Res Cogn Brain Res. 1997;6:37–52. doi: 10.1016/s0926-6410(97)00012-8. [DOI] [PubMed] [Google Scholar]
- Philipp M, Bornkessel-Schlesewsky I, Bisang W, Schlesewsky M. The role of animacy in the real time comprehension of Mandarin Chinese: Evidence from auditory event-related brain potentials. Brain Lang. 2007 doi: 10.1016/j.bandl.2007.09.005. [DOI] [PubMed] [Google Scholar]
- Postle BR. Working memory as an emergent property of the mind and brain. Neuroscience. 2006;139:23–38. doi: 10.1016/j.neuroscience.2005.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romanski LM. Domain specificity in the primate prefrontal cortex. Cogn Affect Behav Neurosci. 2004;4:421–429. doi: 10.3758/cabn.4.4.421. [DOI] [PubMed] [Google Scholar]
- Ruchkin DS, Johnson R, Jr, Canoune H, Ritter W. Short-term memory storage and retention: an event-related brain potential study. Electroencephalogr Clin Neurophysiol. 1990;76:419–439. doi: 10.1016/0013-4694(90)90096-3. [DOI] [PubMed] [Google Scholar]
- Sabourin L, Stowe L. Memory effects in syntactic ERP tasks. Brain Cogn. 2004;55:392–395. doi: 10.1016/j.bandc.2004.02.056. [DOI] [PubMed] [Google Scholar]
- Service E, Helenius P, Maury S, Salmelin R. Localization of syntactic and semantic brain responses using magnetoencephalography. J Cogn Neurosci. 2007;19:1193–1205. doi: 10.1162/jocn.2007.19.7.1193. [DOI] [PubMed] [Google Scholar]
- Steinhauer K, Drury JE. On the early left-anterior negativity (ELAN) in syntax studies. Brain Lang. 2012;120:135–162. doi: 10.1016/j.bandl.2011.07.001. [DOI] [PubMed] [Google Scholar]
- Steinhauer K, Drury JE, Portner P, Walenski M, Ullman MT. Syntax, concepts, and logic in the temporal dynamics of language comprehension: evidence from event-related potentials. Neuropsychologia. 2010;48:1525–1542. doi: 10.1016/j.neuropsychologia.2010.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traxler M, Morris R, Seely R. Processing subject and object relative clauses: Evidence from eye movements. J Mem Lang. 2002;47:69–90. [Google Scholar]
- Traxler MJ, Williams RS, Blozis SA, Morris RK. Working memory, animacy, and verb class in the processing of relative clauses. Journal of Memory and Language. 2005;53:204–224. [Google Scholar]
- Wassenaar M, Hagoort P. Thematic role assignment in patients with Broca’s aphasia: sentence-picture matching electrified. Neuropsychologia. 2007;45:716–740. doi: 10.1016/j.neuropsychologia.2006.08.016. [DOI] [PubMed] [Google Scholar]
- Weckerly J, Kutas M. An electrophysiological analysis of animacy effects in the processing of object relative sentences. Psychophysiology. 1999;36:559–570. [PubMed] [Google Scholar]
- Wilson M. MRC Psycholinguistic Database: Machine-usable dictionary, version 2.00. Behavior Research Methods. 1988;20:6–10. [Google Scholar]